A Small Aromatic Compound Has Antifungal Properties and Potential Anti-Inflammatory Effects against Intestinal Inflammation


 , , and
, , and 
Abstract
1. Introduction
2. Results
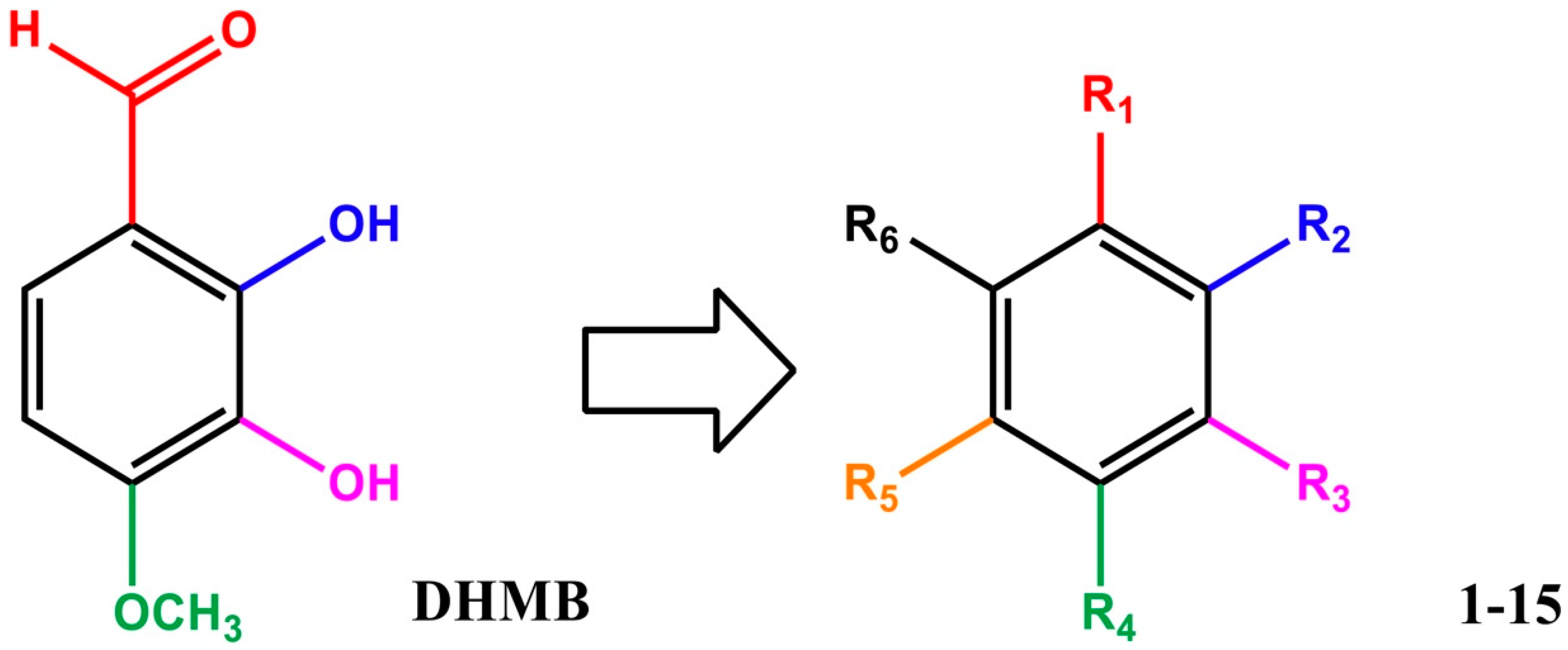
2.1. Structure and Characterisation of DHMB
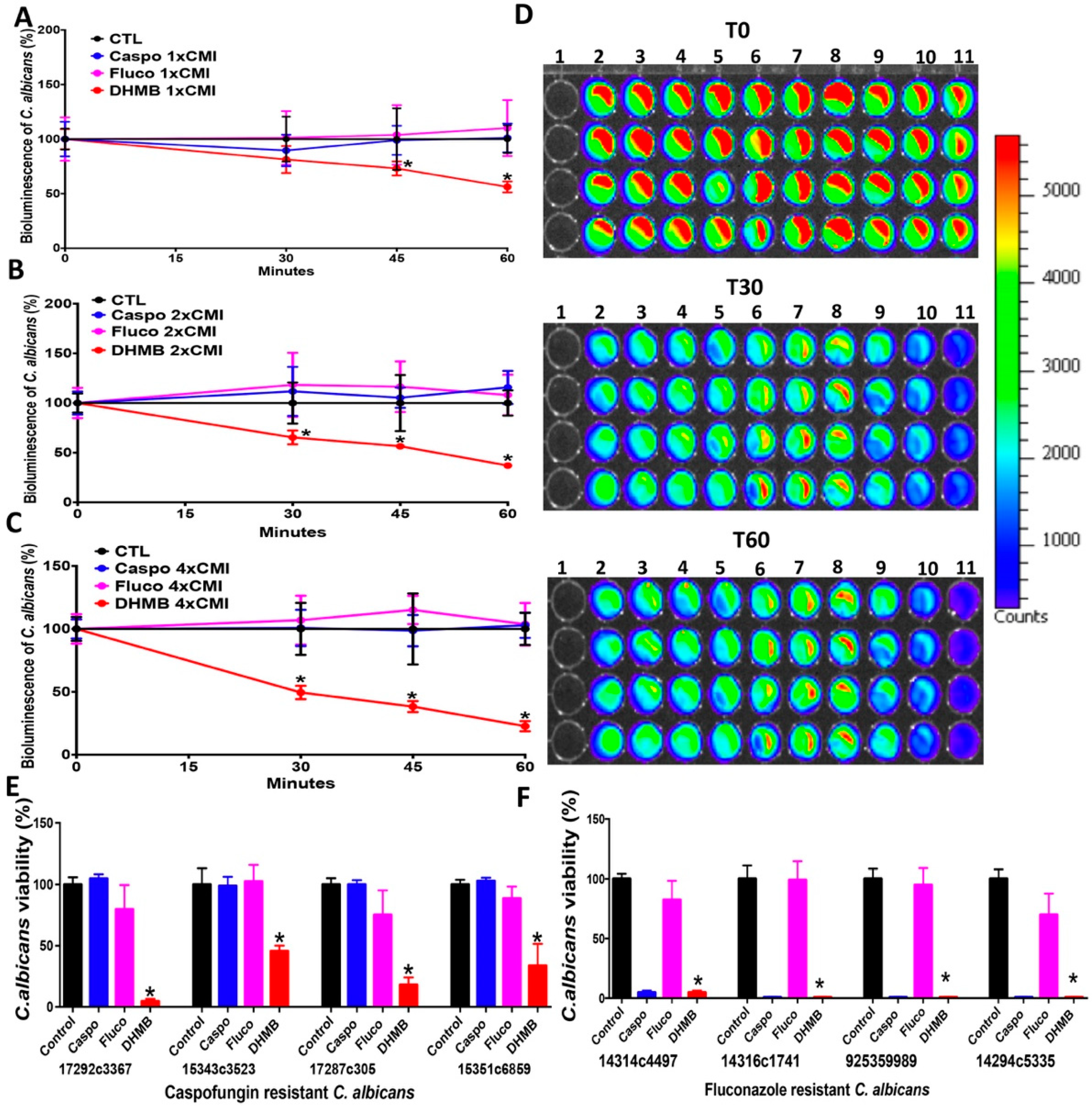
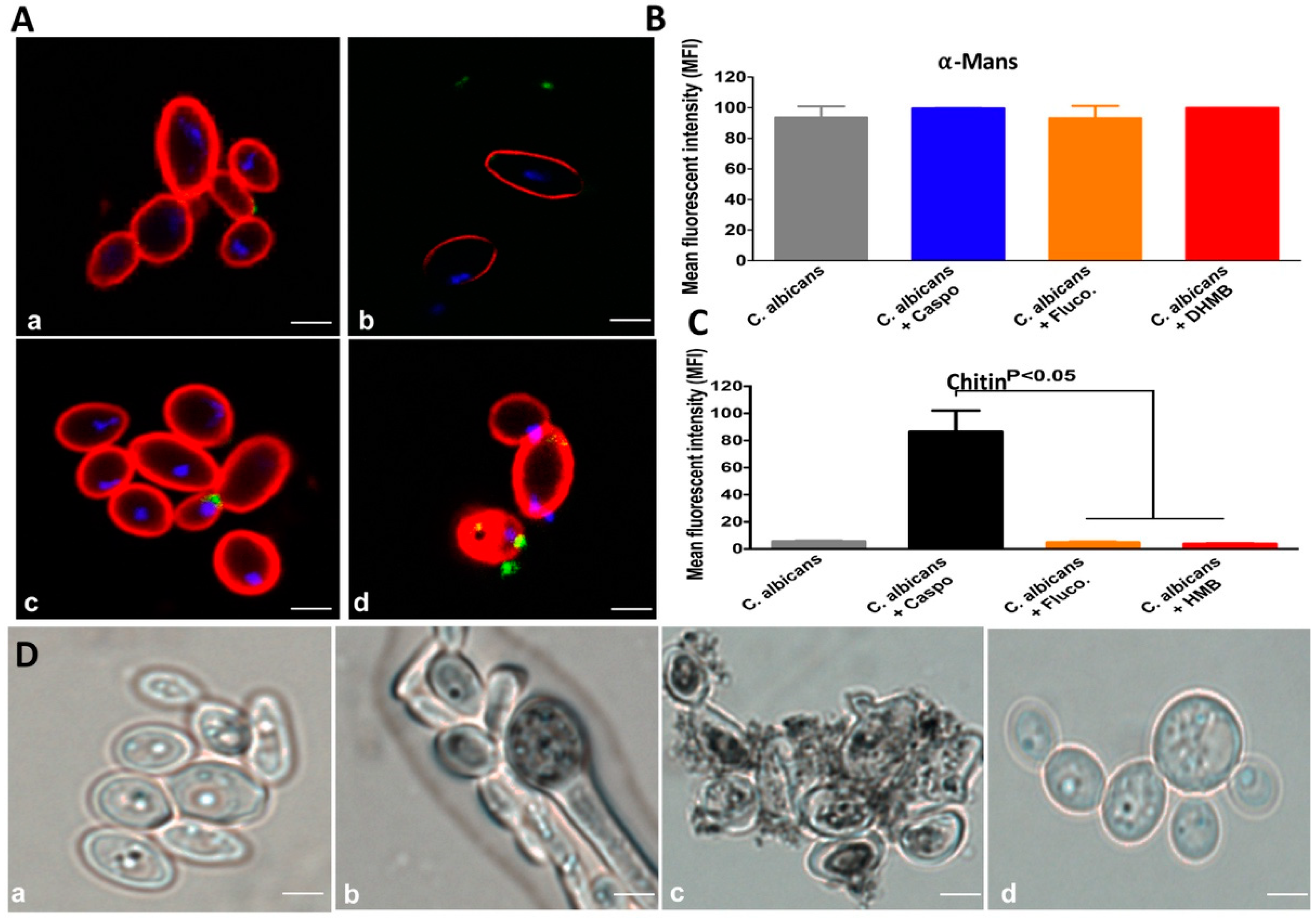
2.2. In Vitro Antifungal Activity of DHMB against C. albicans
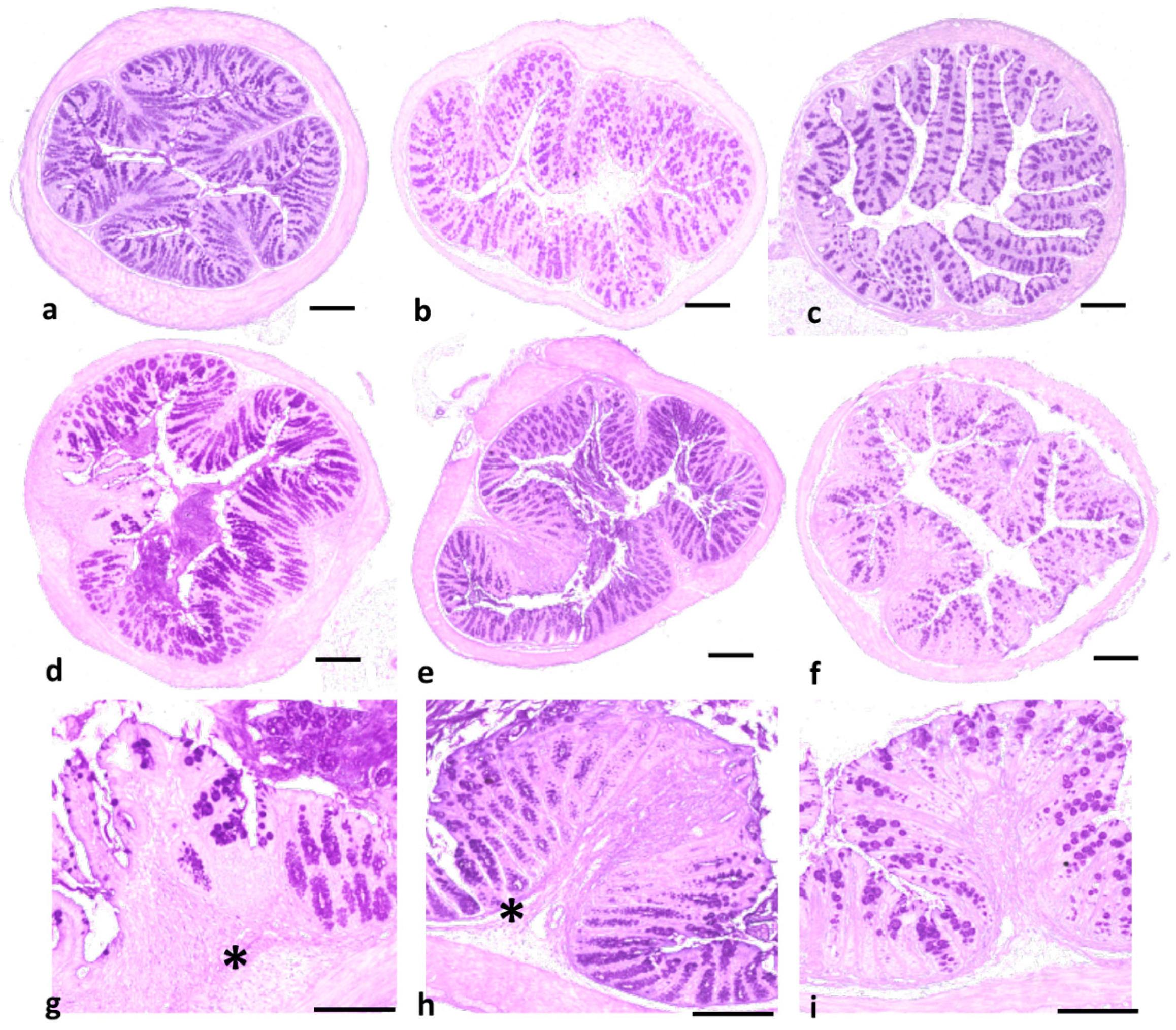
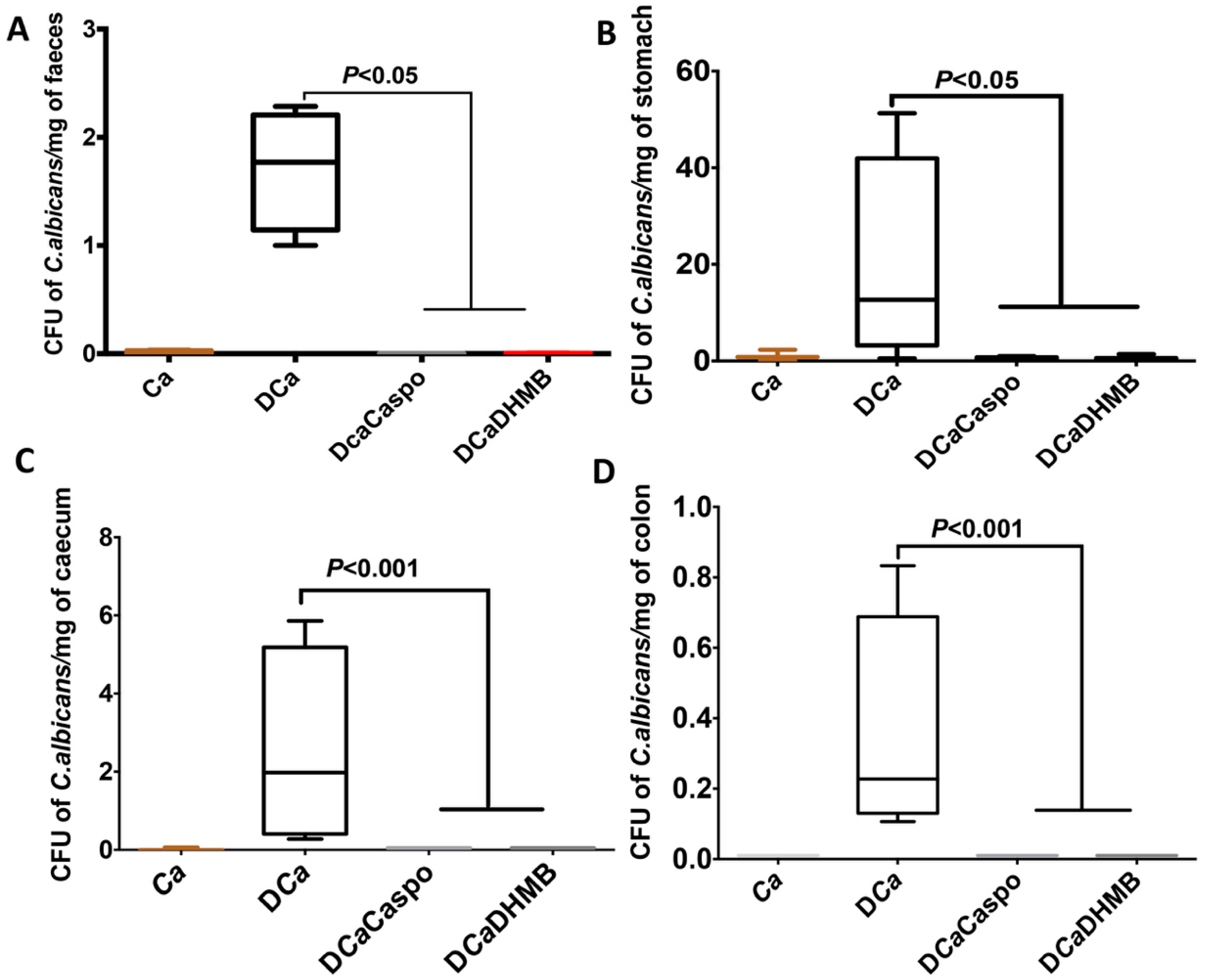
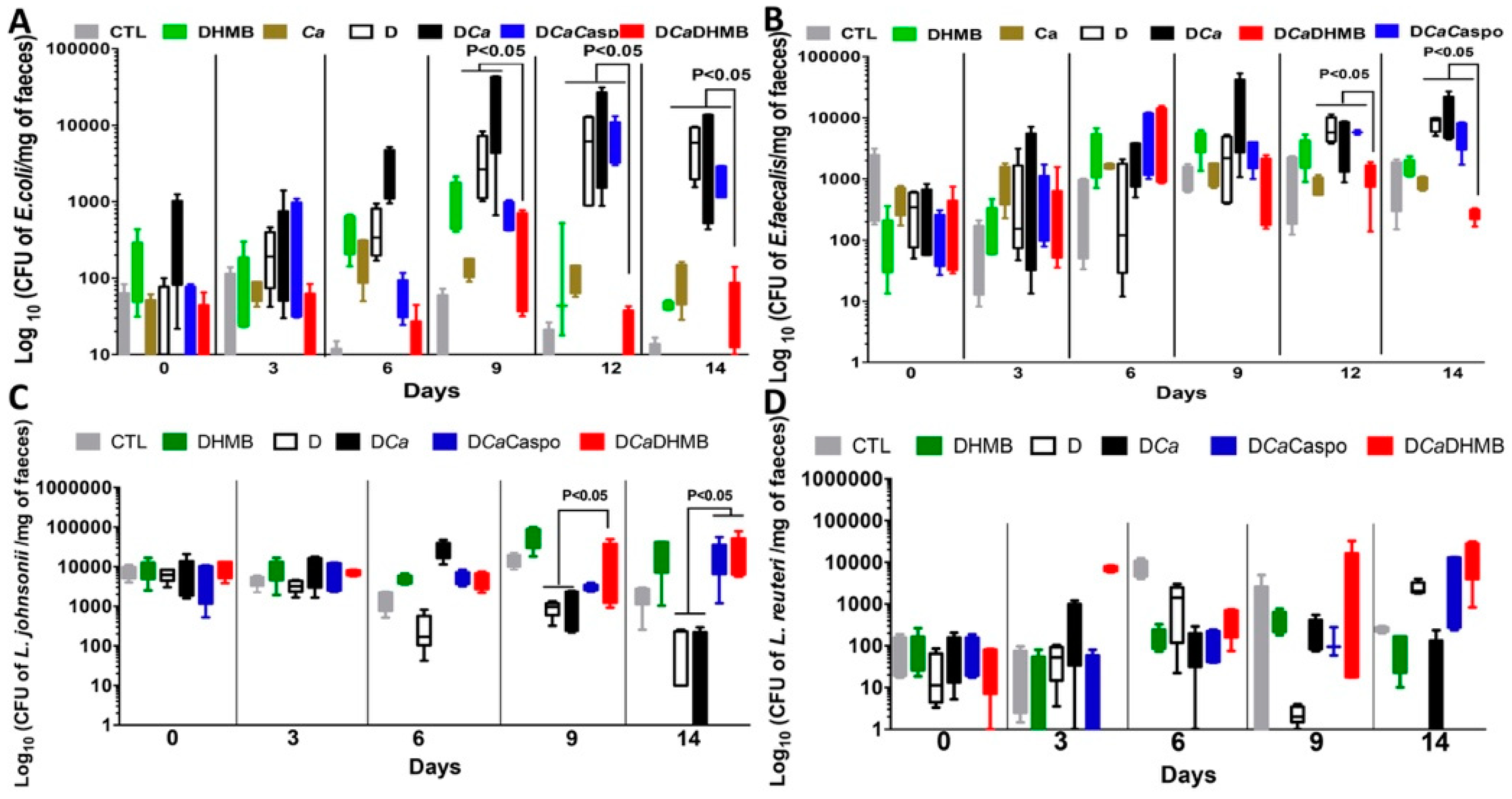
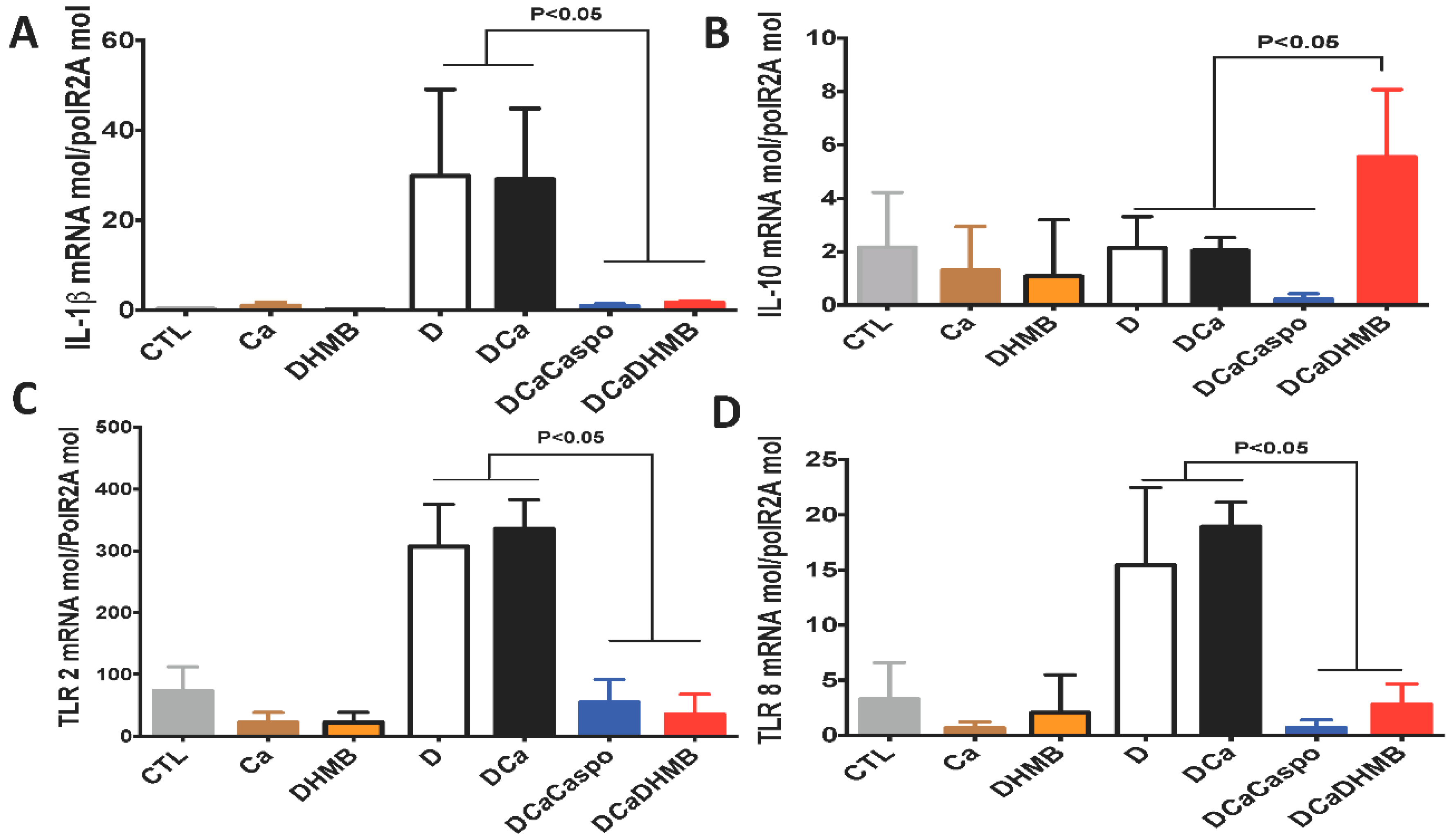
2.3. Effect of DHMB on C. albicans Colonisation and Intestinal Inflammation in DSS-Induced Colitis Model
3. Discussion
4. Materials and Methods
4.1. C. albicans Strains and Growth Conditions
4.2. Antifungal Compounds
4.3. Fungal Viability Assays
4.4. Flow Cytometry and Confocal Microscopy
4.5. Animals
4.6. C. albicans Challenge and Induction of Colitis
4.7. Determination of Clinical and Histological Scores of Inflammation
4.8. Real-Time mRNA Quantification of Pro-Inflammatory Cytokines and Innate Immune Receptors
5. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Baumgart, D.C.; Sandborn, W.J. Inflammatory bowel disease: Clinical aspects and established and evolving therapies. Lancet 2007, 369, 1641–1657. [Google Scholar] [CrossRef]
- Oyri, S.F.; Muzes, G.; Sipos, F. Dysbiotic gut microbiome: A key element of Crohn’s disease. Comp. Immunol. Microbiol. Infect. Dis. 2015, 43, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Gevers, D.; Kugathasan, S.; Denson, L.A.; Vazquez-Baeza, Y.; Van Treuren, W.; Ren, B.; Schwager, E.; Knights, D.; Song, S.J.; Yassour, M.; et al. The treatment-naive microbiome in new-onset Crohn’s disease. Cell Host Microbe 2014, 15, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Darfeuille-Michaud, A.; Neut, C.; Barnich, N.; Lederman, E.; Di Martino, P.; Desreumaux, P.; Gambiez, L.; Joly, B.; Cortot, A.; Colombel, J.F. Presence of adherent Escherichia coli strains in ileal mucosa of patients with Crohn’s disease. Gastroenterology 1998, 115, 1405–1413. [Google Scholar] [CrossRef]
- Jawhara, S.; Thuru, X.; Standaert-Vitse, A.; Jouault, T.; Mordon, S.; Sendid, B.; Desreumaux, P.; Poulain, D. Colonization of mice by Candida albicans is promoted by chemically induced colitis and augments inflammatory responses through galectin-3. J. Infect. Dis. 2008, 197, 972–980. [Google Scholar] [CrossRef] [PubMed]
- Jawhara, S.; Poulain, D. Saccharomyces boulardii decreases inflammation and intestinal colonization by Candida albicans in a mouse model of chemically-induced colitis. Med. Mycol. 2007, 45, 691–700. [Google Scholar] [CrossRef] [PubMed]
- Standaert-Vitse, A.; Sendid, B.; Joossens, M.; Francois, N.; Vandewalle-El Khoury, P.; Branche, J.; Van Kruiningen, H.; Jouault, T.; Rutgeerts, P.; Gower-Rousseau, C.; et al. Candida albicans colonization and ASCA in familial Crohn’s disease. Am. J. Gastroenterol. 2009, 104, 1745–1753. [Google Scholar] [CrossRef]
- Poulain, D.; Sendid, B.; Standaert-Vitse, A.; Fradin, C.; Jouault, T.; Jawhara, S.; Colombel, J.F. Yeasts: Neglected pathogens. Dig. Dis. 2009, 27 (Suppl. 1), 104–110. [Google Scholar] [CrossRef]
- Charlet, R.; Pruvost, Y.; Tumba, G.; Istel, F.; Poulain, D.; Kuchler, K.; Sendid, B.; Jawhara, S. Remodeling of the Candida glabrata cell wall in the gastrointestinal tract affects the gut microbiota and the immune response. Sci. Rep. 2018, 8, 3316. [Google Scholar] [CrossRef]
- Ardizzone, S.; Cassinotti, A.; Manes, G.; Porro, G.B. Immunomodulators for all patients with inflammatory bowel disease? Ther. Adv. Gastroenterol. 2010, 3, 31–42. [Google Scholar] [CrossRef]
- Campieri, M. New steroids and new salicylates in inflammatory bowel disease: A critical appraisal. Gut 2002, 50 (Suppl. 3), III43–III46. [Google Scholar] [CrossRef] [PubMed]
- Wiederhold, N.P. Antifungal resistance: Current trends and future strategies to combat. Infect. Drug Resist. 2017, 10, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Ostrosky-Zeichner, L.; Casadevall, A.; Galgiani, J.N.; Odds, F.C.; Rex, J.H. An insight into the antifungal pipeline: Selected new molecules and beyond. Nat. Rev. Drug Discov. 2010, 9, 719–727. [Google Scholar] [CrossRef] [PubMed]
- Almabruk, K.H.; Chang, J.H.; Mahmud, T. Total Synthesis of (+/−)-Isoperbergins and Correction of the Chemical Structure of Perbergin. J. Nat. Prod. 2016, 79, 2391–2396. [Google Scholar] [CrossRef] [PubMed]
- Pettit, G.R.; Singh, S.B.; Niven, M.L.; Hamel, E.; Schmidt, J.M. Isolation, structure, and synthesis of combretastatins A-1 and B-1, potent new inhibitors of microtubule assembly, derived from Combretum caffrum. J. Nat. Prod. 1987, 50, 119–131. [Google Scholar] [CrossRef]
- McKittrick, B.A.; Stevenson, R. A simple synthesis of the natural benzofuranoquinone, acamelin. J. Chem. Soc. Perkin Trans. 1 1983, 10, 2423–2424. [Google Scholar] [CrossRef]
- Spath, E.; Dobrovolny, E. Natural coumarins. XLII. Synthesis of fraxetin, fraxidin and isofraxidin. Berichte der Deutschen Chemischen Gesellschaft 1938, 71B, 1831–1836. [Google Scholar] [CrossRef]
- Cervi, A.; Aillard, P.; Hazeri, N.; Petit, L.; Chai, C.L.; Willis, A.C.; Banwell, M.G. Total syntheses of the coumarin-containing natural products pimpinellin and fraxetin using Au(I)-catalyzed intramolecular hydroarylation (IMHA) chemistry. J. Org. Chem. 2013, 78, 9876–9882. [Google Scholar] [CrossRef]
- Wang, L.; Yin, Z.Q.; Zhang, L.H.; Ye, W.C.; Zhang, X.Q.; Shen, W.B.; Zhao, S.X. Chemical constituents from root barks of Periploca sepium. China J. Chin. Mater. Med. 2007, 32, 1300–1302. [Google Scholar]
- Tokiwa, T.; Harada, K.; Matsumura, T.; Tukiyama, T. Oriental medicinal herb, Periploca sepium, extract inhibits growth and IL-6 production of human synovial fibroblast-like cells. Biol. Pharm. Bull. 2004, 27, 1691–1693. [Google Scholar] [CrossRef]
- Chu, S.S.; Jiang, G.H.; Liu, W.L.; Liu, Z.L. Insecticidal activity of the root bark essential oil of Periploca sepium Bunge and its main component. Nat. Prod. Res. 2012, 26, 926–932. [Google Scholar] [CrossRef] [PubMed]
- Morris-Jones, R.; Gomez, B.L.; Diez, S.; Uran, M.; Morris-Jones, S.D.; Casadevall, A.; Nosanchuk, J.D.; Hamilton, A.J. Synthesis of melanin pigment by Candida albicans in vitro and during infection. Infect. Immun. 2005, 73, 6147–6150. [Google Scholar] [CrossRef] [PubMed]
- Eisenman, H.C.; Duong, R.; Chan, H.; Tsue, R.; McClelland, E.E. Reduced virulence of melanized Cryptococcus neoformans in Galleria mellonella. Virulence 2014, 5, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Qualls, J.E.; Kaplan, A.M.; van Rooijen, N.; Cohen, D.A. Suppression of experimental colitis by intestinal mononuclear phagocytes. J. Leukoc. Biol. 2006, 80, 802–815. [Google Scholar] [CrossRef]
- Kokkotou, E.; Moss, A.C.; Torres, D.; Karagiannides, I.; Cheifetz, A.; Liu, S.; O’Brien, M.; Maratos-Flier, E.; Pothoulakis, C. Melanin-concentrating hormone as a mediator of intestinal inflammation. Proc. Natl. Acad. Sci. USA 2008, 105, 10613–10618. [Google Scholar] [CrossRef]
- Doyle, T.C.; Nawotka, K.A.; Kawahara, C.B.; Francis, K.P.; Contag, P.R. Visualizing fungal infections in living mice using bioluminescent pathogenic Candida albicans strains transformed with the firefly luciferase gene. Microb. Pathog. 2006, 40, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, B.; Martins, M.A.; Bonfietti, L.X.; Szeszs, M.W.; Jacobs, J.; Garcia, C.; Melhem, M.S. Species distribution and antifungal susceptibility profile of Candida isolates from bloodstream infections in Lima, Peru. J. Med. Microbiol. 2014, 63 Pt 6, 855–860. [Google Scholar] [CrossRef]
- Choteau, L.; Parny, M.; Francois, N.; Bertin, B.; Fumery, M.; Dubuquoy, L.; Takahashi, K.; Colombel, J.F.; Jouault, T.; Poulain, D.; et al. Role of mannose-binding lectin in intestinal homeostasis and fungal elimination. Mucosal Immunol. 2016, 9, 767–776. [Google Scholar] [CrossRef]
- Sendid, B.; Francois, N.; Standaert, A.; Dehecq, E.; Zerimech, F.; Camus, D.; Poulain, D. Prospective evaluation of the new chromogenic medium CandiSelect 4 for differentiation and presumptive identification of the major pathogenic Candida species. J. Med. Microbiol. 2007, 56 Pt 4, 495–499. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entry | Compound | R1 | R2 | R3 | R4 | R5 | R6 | % Inhibition at 32 µg/mL a |
|---|---|---|---|---|---|---|---|---|
| 1 | DHMB | CHO | OH | OH | OCH3 | H | H | 100 |
| 2 | 1 | H | OH | OH | OCH3 | H | H | 3 |
| 3 | 2 |  | OH | OCH3 | OCH3 | H | H | 13 |
| 4 | 3 |  | OH | OH | OCH3 | H | H | 27 |
| 5 | 4 | OH | OCOCH2Cl | OCH3 | H | H | 6 | |
| 6 | 5 | OCOCH2Cl | OCOCH2Cl | OCH3 | H | H | 4 | |
| 7 | 6 |  | H | 2 | ||||
| 8 | 7 | CHO | OH | OH | OH | H | H | 4 |
| 9 | 8 | H | OH | OH | OCH3 | H | COOH | 2 |
| 10 | 9 | H | OCH3 | OCH3 | OCH3 | H | COOH | 2 |
| 11 | 10 | H | OCH3 | OH | OCH3 | H | COOH | 0 |
| 12 | 11 | H | OCH3 | OCH3 | OCH3 | H | CH2-COOH | 1 |
| 13 | 12 | H | OCH3 | OCH3 | OCH3 | H | CH2-OH | 2 |
| 14 | 13 | H | OCH3 | OH | OCH3 | H | CHO | 1 |
| 15 | 14 | H | OCH3 | OH | OCH3 | H | COCH3 | 4 |
| 16 | 15 | H | OCH3 | OH | OCH3 | H | H | 0 |
| Strains | Description | MIC Caspofungin (µg/mL) | MIC Fluconazole (µg/mL) | MIC DHMB (µg/mL) | Ref. |
|---|---|---|---|---|---|
| C. albicans ATCC 90028 | Wild-type | 0.03 | 0.5 | 8 | This study |
| Bioluminescent C. albicans | C. albicans strain CAI4 (ura3::imm434/ura3::imm434) | 0.03 | 0.5 | 8 | [26] |
| C. albicans 14314c4497 | Anal, fluconazole resistant | 0.06 | 128 | 80 | This study |
| C. albicans 14316c1746 | Bronchoalveolar lavage, fluconazole resistant | 0.03 | 128 | 80 | This study |
| C. albicans 92535989 | Tracheal secretion, fluconazole resistant | 0.06 | 64 | 80 | This study |
| C. albicans 14294c5335 | Stools, fluconazole resistant | 0.06 | 5 | 80 | This study |
| C. albicans 17292c3367 | Venous catheter, caspofungin resistant | 8 | 0.5 | 80 | This study |
| C. albicans 15343c3523 | Blood, caspofungin resistant | 2 | 0.5 | 80 | This study |
| C. albicans 17287c305 | Blood, caspofungin resistant | 8 | 0.5 | 80 | This study |
| C. albicans 15351c6859 | Venous catheter, caspofungin resistant | 4 | 1 | 80 | This study |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bortolus, C.; Billamboz, M.; Charlet, R.; Lecointe, K.; Sendid, B.; Ghinet, A.; Jawhara, S. A Small Aromatic Compound Has Antifungal Properties and Potential Anti-Inflammatory Effects against Intestinal Inflammation. Int. J. Mol. Sci. 2019, 20, 321. https://doi.org/10.3390/ijms20020321
Bortolus C, Billamboz M, Charlet R, Lecointe K, Sendid B, Ghinet A, Jawhara S. A Small Aromatic Compound Has Antifungal Properties and Potential Anti-Inflammatory Effects against Intestinal Inflammation. International Journal of Molecular Sciences. 2019; 20(2):321. https://doi.org/10.3390/ijms20020321
Chicago/Turabian StyleBortolus, Clovis, Muriel Billamboz, Rogatien Charlet, Karine Lecointe, Boualem Sendid, Alina Ghinet, and Samir Jawhara. 2019. "A Small Aromatic Compound Has Antifungal Properties and Potential Anti-Inflammatory Effects against Intestinal Inflammation" International Journal of Molecular Sciences 20, no. 2: 321. https://doi.org/10.3390/ijms20020321
APA StyleBortolus, C., Billamboz, M., Charlet, R., Lecointe, K., Sendid, B., Ghinet, A., & Jawhara, S. (2019). A Small Aromatic Compound Has Antifungal Properties and Potential Anti-Inflammatory Effects against Intestinal Inflammation. International Journal of Molecular Sciences, 20(2), 321. https://doi.org/10.3390/ijms20020321