Holothuria Leucospilota Polysaccharides Ameliorate Hyperlipidemia in High-Fat Diet-Induced Rats via Short-Chain Fatty Acids Production and Lipid Metabolism Regulation


Abstract
1. Introduction
2. Results and Discussion
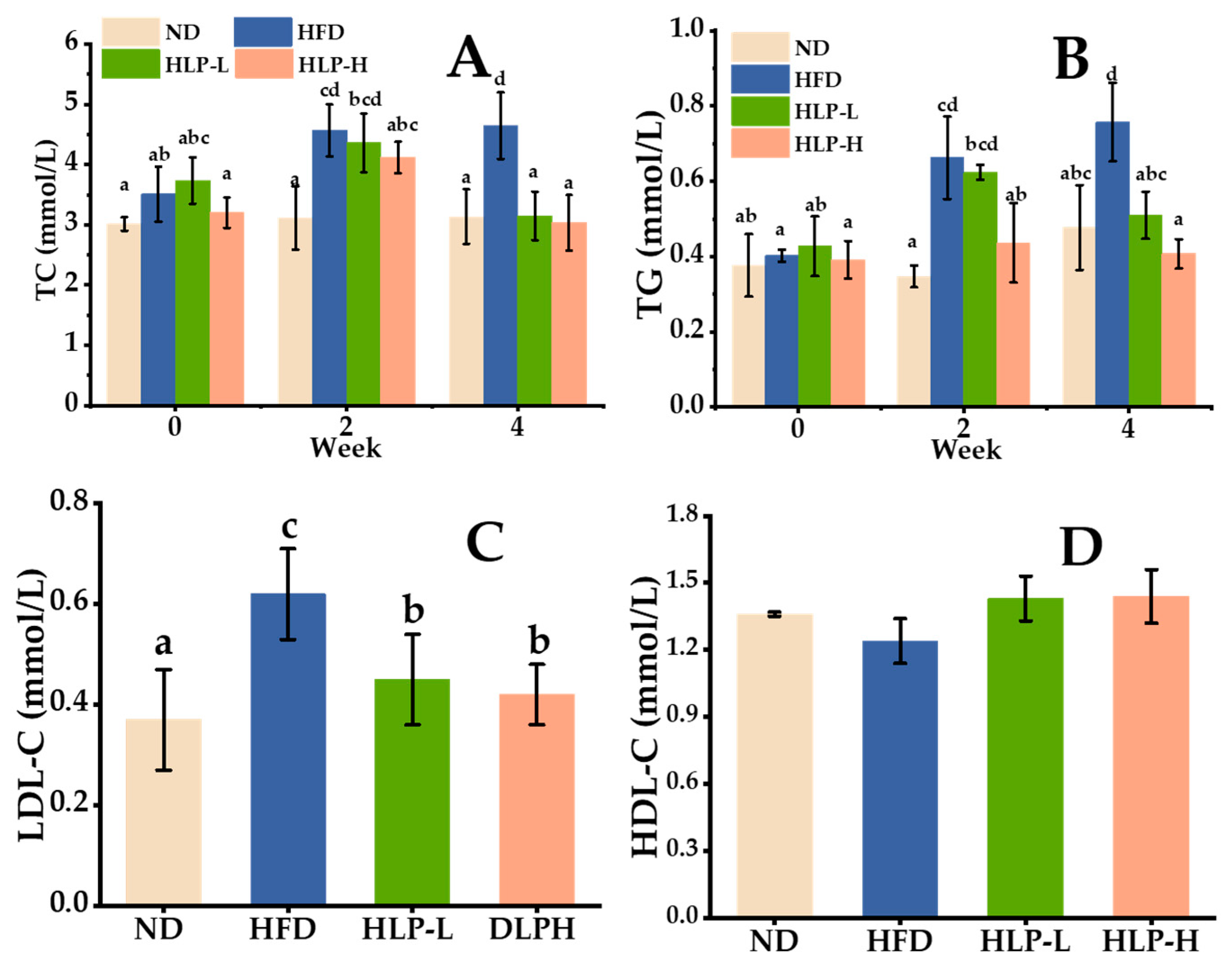
2.1. The Effects of HLP on Normalizing Serum Lipid Metabolic Disorders
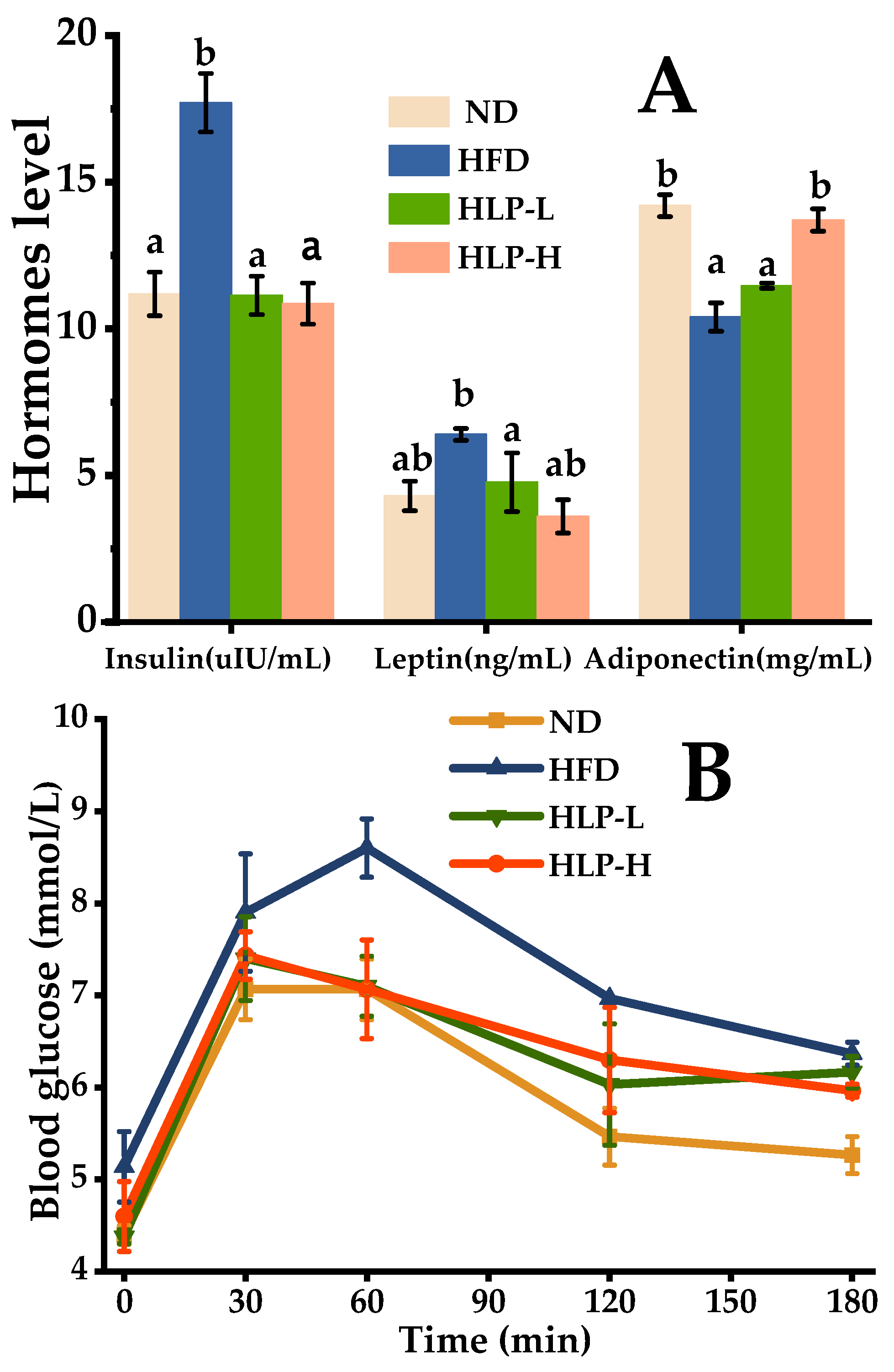
2.2. HLP Alleviated the Hormone Levels in Hyperlipidemia Rats
2.3. HLP Inhibit Inflammation in Hyperlipidemia Rats
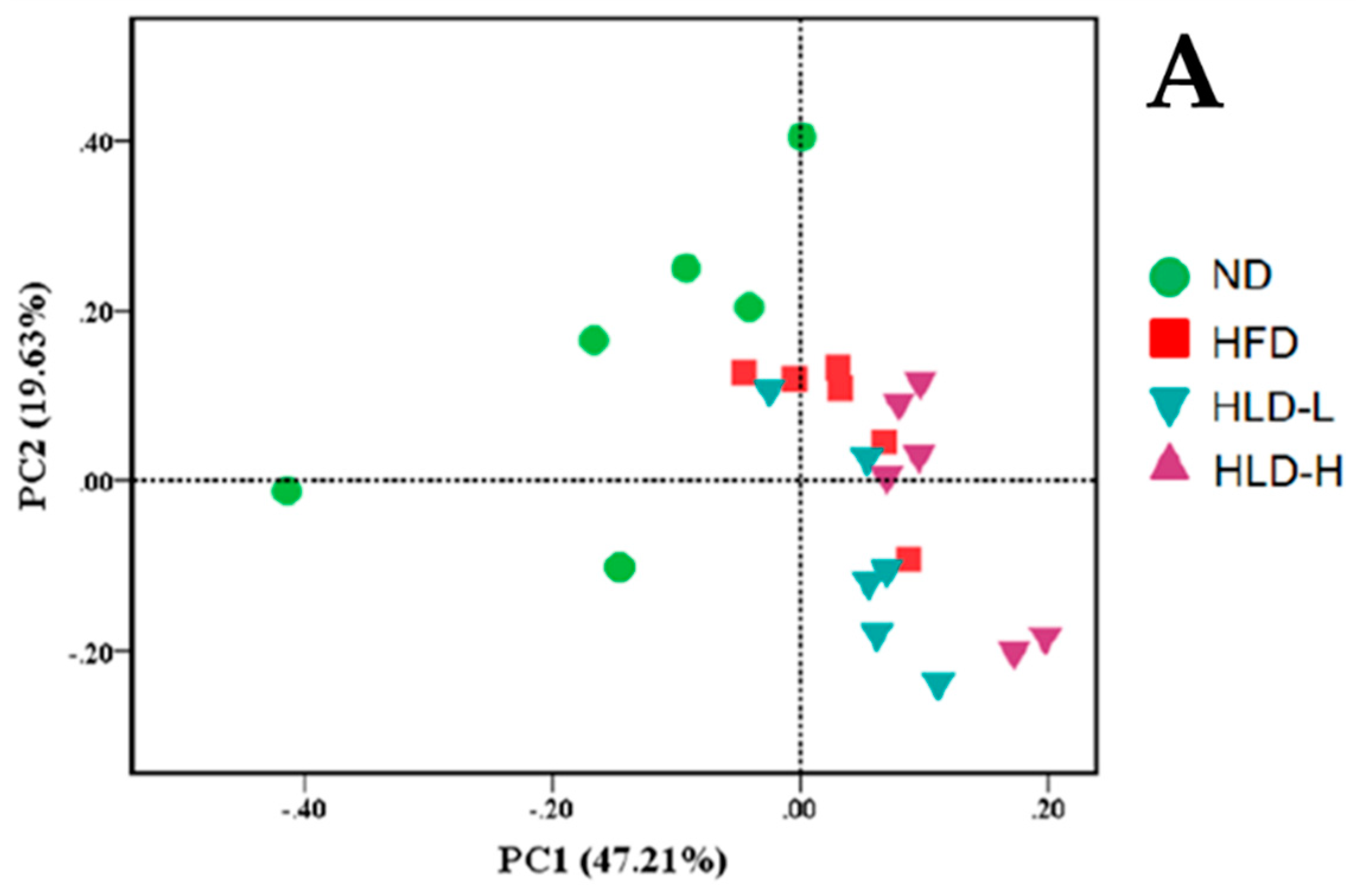
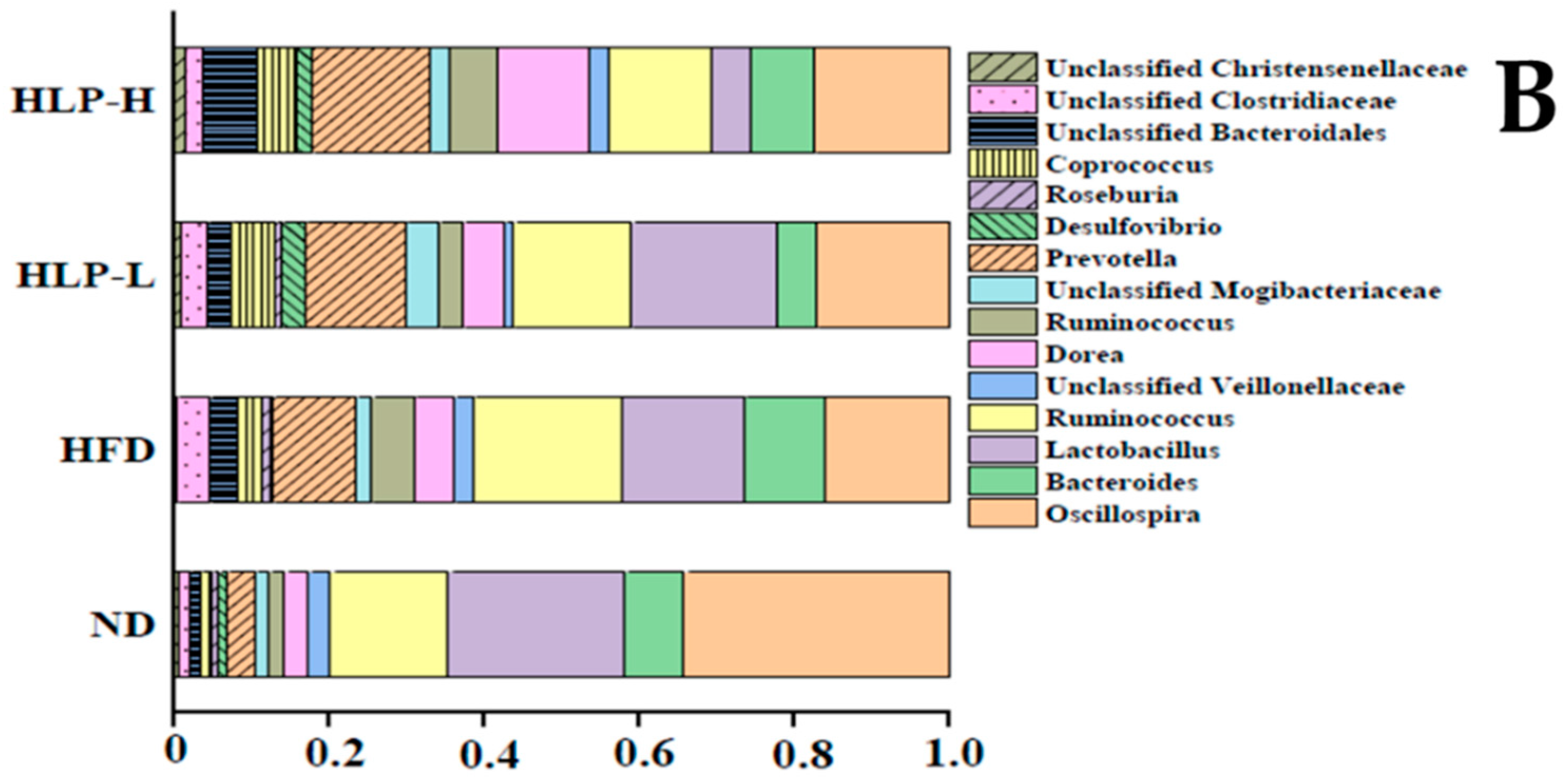
2.4. The Changes of Gut Microbiota in Hyperlipidemia Rats
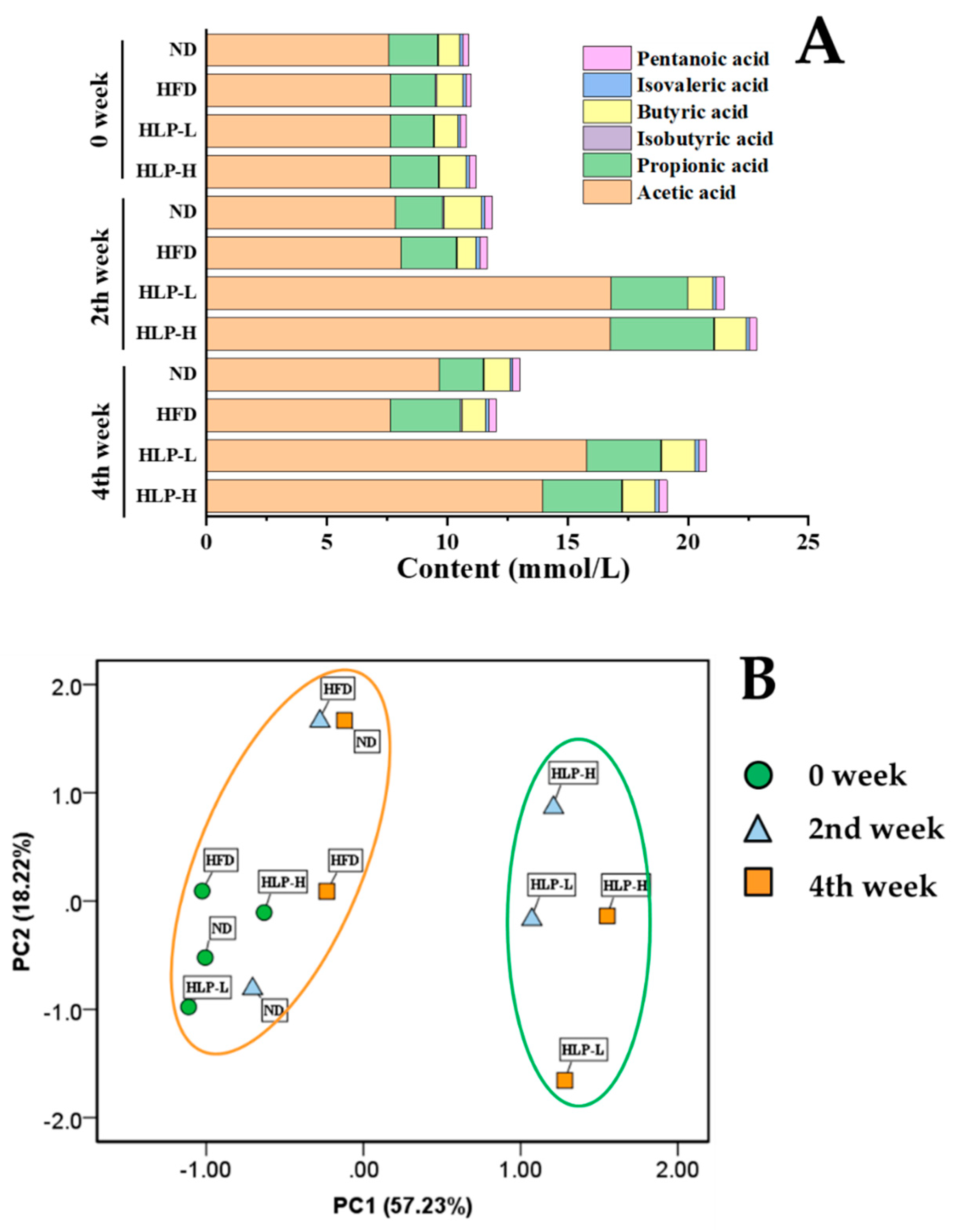
2.5. HLP Altered the SCFA Profile
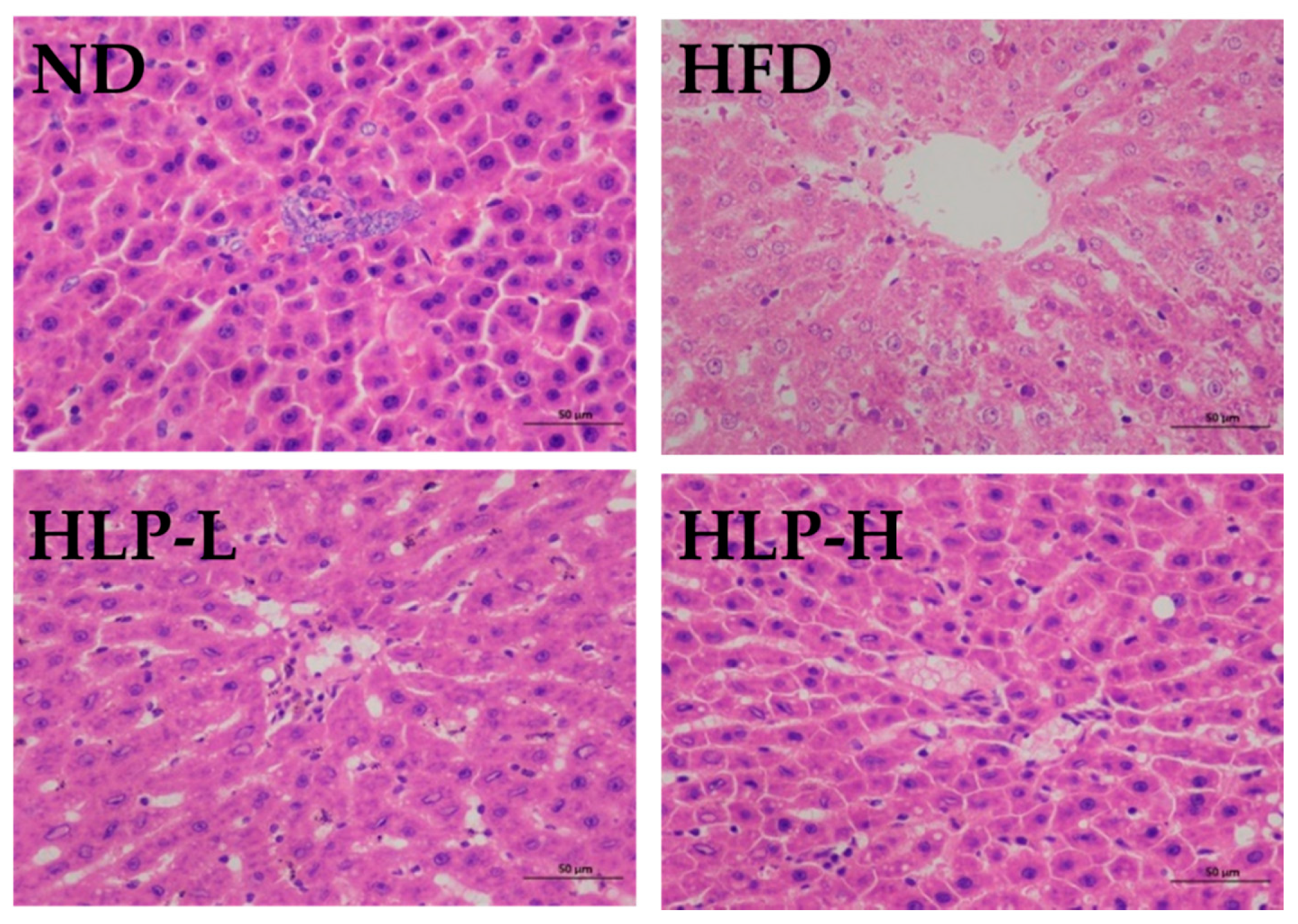
2.6. The Effects of HLP Inhibited Liver Lesions
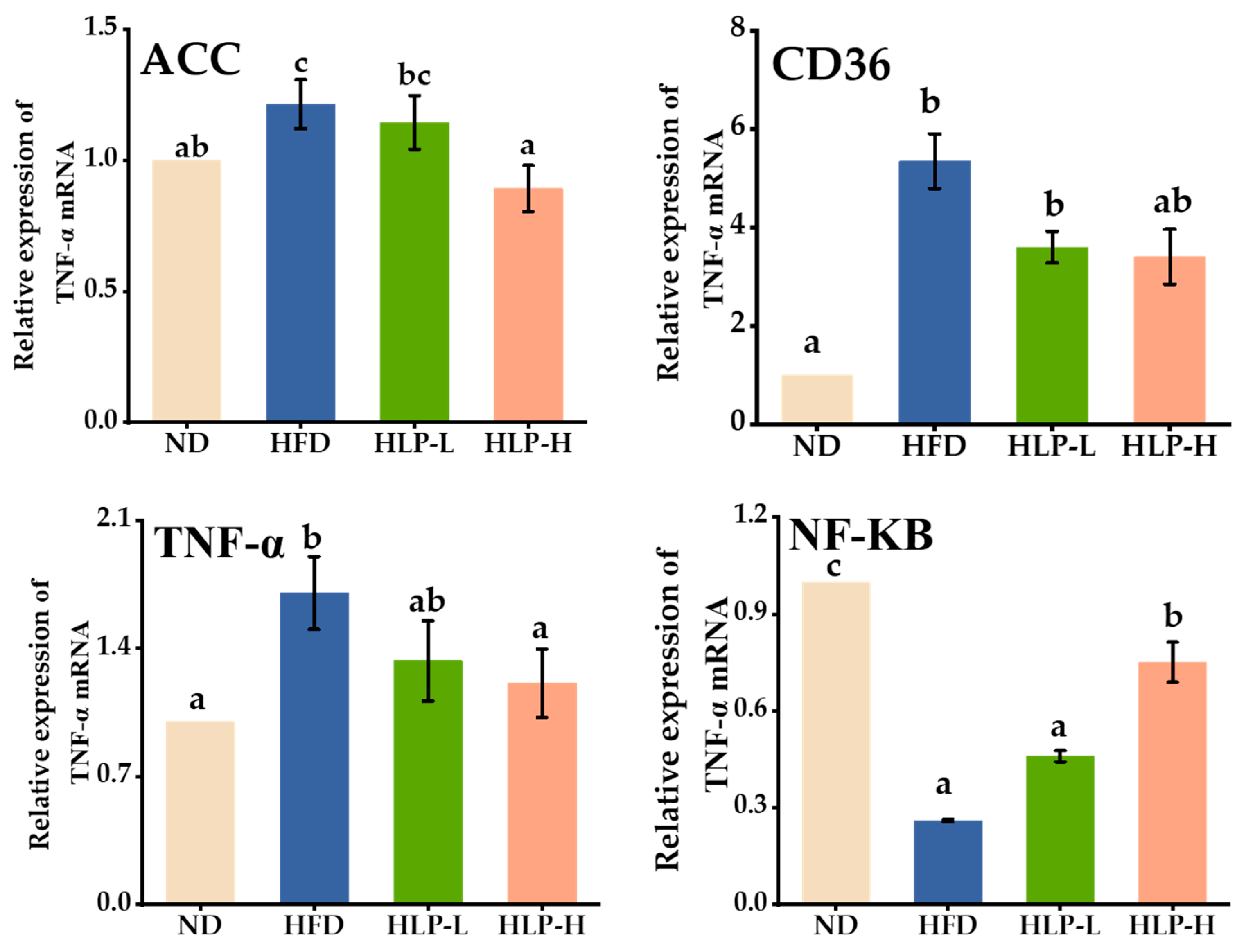
2.7. HLP Regulated Lipogenesis-Related Genes in Hyperlipidemia Rats of Liver
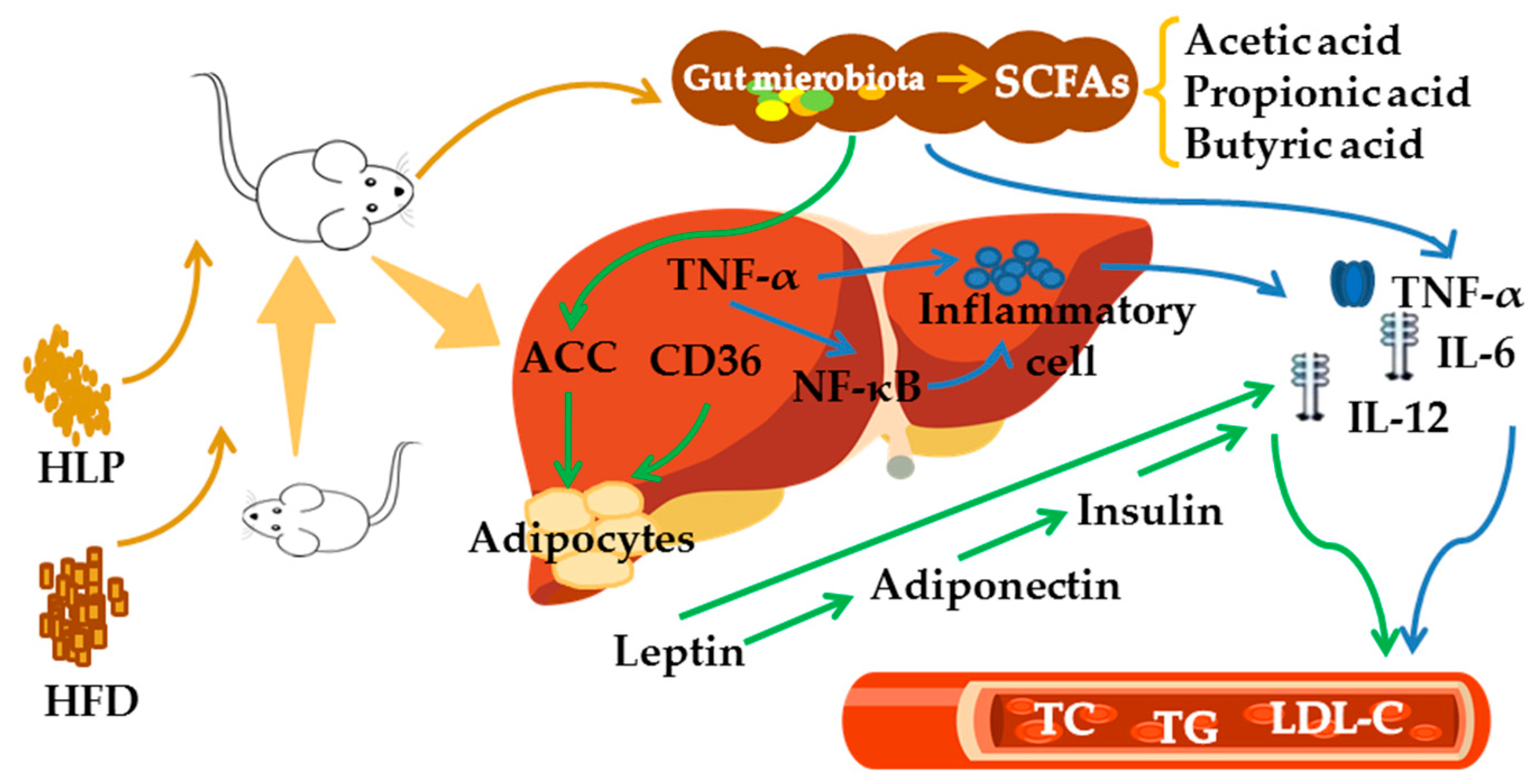
2.8. HLP Alleviate Hyperlipidemia via Enhancing SCFAs to Regulate Lipid Metabolism
3. Materials and Methods
3.1. Materials
3.2. HLP Preparation
3.3. Animals and Ethics Statement
3.4. Experimental Design
3.5. Glucose Tolerance Test
3.6. Serum Biochemical Analysis
3.7. The Gut Microbiota Analysis by 16S rDNA High-Throughput Sequencing
3.8. SCFAs Analysis
3.9. Histological Examinations
3.10. RNA Preparation and Real-Time PCR
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| ACC | CAACCACTACGGCATGACTCA | CGCAGAAGCAGCCCATTACTT |
| CD 36 | TGAGCCTTCACTGTCTGTTGGAAC | AGGCTGTTGAGCACACCTTGAAC |
| TNF-α | GCATGATCCGAGATGTGGAACTGG | CGCCACGAGCAGGAATGAGAAG |
| NF-κB | TGTGGTGGAGGACTTGCTGAGG | AGTGCTGCCTTGCTGTTCTTGAG |
Appendix B
| Groups | 0 Week | 1th Week | 2nd Week | 3rd Week | 4th Week |
|---|---|---|---|---|---|
| ND | 199.51 ± 18.85 a | 227.56 ± 14.03 a | 246.26 ± 12.36 a | 241.92 ± 11.44 a | 229.05 ± 18.44 a |
| HFD | 205.14 ± 16.19 a | 239.49 ± 17.58 b | 271.30 ± 16.84 c | 262.30 ± 17.08 b | 252.04 ± 16.93 c |
| HLP-L | 199.85 ± 14.16 a | 240.26 ± 13.44 b | 261.65 ± 12.78 b | 261.57 ± 15.63 ab | 242.39 ± 16.95 b |
| HLP-H | 197.56 ± 13.97 a | 230.26 ± 13.39 a | 249.40 ± 17.03 a | 251.02 ± 17.79 ab | 241.77 ± 13.95 b |
References
- De Costa, G.; Park, A. Hyperlipidaemia. Medicine 2017, 45, 579–582. [Google Scholar] [CrossRef]
- Xu, P.; Zhai, Y.; Wang, J. The Role of PPAR and Its Cross-Talk with CAR and LXR in Obesity and Atherosclerosis. Int. J. Mol. Sci. 2018, 19, 1260. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zeng, F.; Li, S.; Liu, Y.; Gong, S.; Lv, X.; Zhang, J.; Liu, B. Spirulina active substance mediated gut microbes improve lipid metabolism in high-fat diet fed rats. J. Funct. Foods 2019, 59, 215–222. [Google Scholar] [CrossRef]
- Guarner, F. Decade in review-gut microbiota: The gut microbiota era marches on. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 647–649. [Google Scholar] [CrossRef] [PubMed]
- Yerlikaya, F.H.; Mehmet, Ö. Aberrant expression of miRNA profiles in high-fat and high-sucrose fed rats. Clin. Nutr. Exp. 2019, 20. [Google Scholar] [CrossRef]
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The re-emergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xue, C.; Yin, L.A.; Tang, Q.; Yu, G.; Chai, W. Comparison of structures and anticoagulant activities of fucosylated chondroitin sulfates from different sea cucumbers. Carbohyd. Polym. 2011, 83, 688–696. [Google Scholar] [CrossRef]
- Wang, J.; Hu, S.; Jiang, W.; Song, W.; Cai, L.; Wang, J. Fucoidan from sea cucumber may improve hepatic inflammatory response and insulin resistance in mice. Int. Immunopharmacol. 2016, 31, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Jiang, H.; Cai, C.; Hao, J.; Li, G.; Yu, G. Gut microbiota fermentation of marine polysaccharides and its effects on intestinal ecology: An overview. Carbohydr. Polym. 2018, 179, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, X.; Jiang, H.; Cai, C.; Li, G.; Hao, J.; Yu, G. Marine polysaccharides attenuate metabolic syndrome by fermentation products and altering gut microbiota: An overview. Carbohydr. Polym. 2018, 195, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Gangadaran, S.; Cheema, S.K. A high fat diet enriched with sea cucumber gut powder provides cardio-protective and anti-obesity effects in C57BL/6 mice. Food Res. Int. 2017, 99, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Qi, H.; Ji, X.; Liu, S.; Feng, D.; Dong, X.; He, B.; Srinivas, J.; Yu, C. Antioxidant and anti-dyslipidemic effects of polysaccharidic extract from sea cucumber processing liquor. Electron. J. Biotechnol. 2017, 28, 1–6. [Google Scholar] [CrossRef]
- Li, S.; Li, J.; Zhi, Z.; Wei, C.; Wang, W.; Ding, T.; Ye, X.; Hu, Y.; Linhardt, R.J.; Chen, S. Macromolecular properties and hypolipidemic effects of four sulfated polysaccharides from sea cucumbers. Carbohydr. Polym. 2017, 173, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Chang, Y.; He, M.; Wang, J.; Wang, Y.; Xue, C. Fucosylated chondroitin sulfate from sea cucumber improves insulin sensitivity via activation of PI3K/PKB pathway. J. Food Sci. 2014, 79, H1424–H1429. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Zhu, B.; Sun, Y.; Ai, C.; Wu, S.; Wang, L.; Song, S.; Liu, X. Sulfated polysaccharide from sea cucumber modulates the gut microbiota and its metabolites in normal mice. Int. J. Biol. Macromol. 2018, 120, 502–512. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Li, C.; Zheng, Q.; Wu, J.; Zhu, K.; Shen, X.; Cao, J. Effect of simulated gastrointestinal digestion in vitro on the antioxidant activity, molecular weight and microstructure of polysaccharides from a tropical sea cucumber (Holothuria leucospilota). Food Hydrocolloid. 2019, 89, 735–741. [Google Scholar] [CrossRef]
- Zhang, W.; Lu, Y.; Xu, B.; Wu, J.; Zhang, L.; Gao, M.; Zheng, S.; Wang, A.; Zhang, C.; Chen, L.; et al. Acidic mucopolysaccharide from holothuria leucospilota has antitumor effect by inhibiting angiogenesis and tumor cell invasion in vivo and in vitro. Cancer Biol. Ther. 2014, 8, 1489–1499. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Surhio, M.M.; Wang, Y.; Xu, P.; Shah, F.; Li, J.; Ye, M. Antihyperlipidemic and hepatoprotective properties of selenium modified polysaccharide from Lachnum sp. Int. J. Biol. Macromol. 2017, 99, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Korolenko, T.A.; Johnston, T.P.; Machova, E.; Bgatova, N.P.; Lykov, A.P.; Goncharova, N.V.; Nescakova, Z.; Shintyapina, A.B.; Maiborodin, I.V.; Karmatskikh, O.L. Hypolipidemic effect of mannans from C. albicans serotypes a and B in acute hyperlipidemia in mice. Int. J. Biol. Macromol. 2018, 107, 2385–2394. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Fang, J.-L.; Yuan, K.; Jin, S.-H.; Guo, Y. Ameliorative effect of purified anthocyanin from Lycium ruthenicum on atherosclerosis in rats through synergistic modulation of the gut microbiota and NF-κB/SREBP-2 pathways. J. Funct. Foods 2019, 59, 223–233. [Google Scholar] [CrossRef]
- Vaclav, V.; Jana, V. Effects of yeast-derived beta-glucans on blood cholesterol and macrophage functionality. J. Immunotoxicol. 2009, 6, 30–35. [Google Scholar]
- Malik, S.U.F.; Mahmud, Z.; Alam, J.; Islam, M.S.; Azad, A.K. Relationship among obesity, blood lipids and insulin resistance in Bangladeshi adults. Diabetes Metab. Syndr. 2019, 13, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Stanley, T.L.; Grinspoon, S.K. Effects of growth hormone-releasing hormone on visceral fat, metabolic, and cardiovascular indices in human studies. Growth Horm. IGF Res. 2015, 25, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Vekic, J.; Zeljkovic, A.; Stefanovic, A.; Jelic-Ivanovic, Z.; Spasojevic-Kalimanovska, V. Obesity and dyslipidemia. Metabolism 2019, 92, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhao, J.; Wang, J.; Li, J.; Ouyang, K.; Wang, W. Effects of Cyclocarya paliurus polysaccharide on lipid metabolism-related genes DNA methylation in rats. Int. J. Biol. Macromol. 2019, 123, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.; Kataria, M.A.; Saini, V.; Yadav, A. Role of leptin and adiponectin in insulin resistance. Clinica Chimica Acta 2013, 417, 80–84. [Google Scholar] [CrossRef] [PubMed]
- El-Shinnawy, N.A.; Elhalem, S.S.A.; Haggag, N.Z.; Badr, G. Ameliorative role of camel whey protein and rosuvastatin on induced dyslipidemia in mice. Food Funct. 2018, 9, 1038–1047. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Han, X.; Li, R.; Zhao, W.; Bai, B.; Yan, C.; Dong, X. Anti-atherosclerosis of oligomeric proanthocyanidins from Rhodiola rosea on rat model via hypolipemic, antioxidant, anti-inflammatory activities together with regulation of endothelial function. Phytomedicine 2018, 51, 171–180. [Google Scholar] [CrossRef]
- Song, L.; Schindler, C. IL-6 and the acute phase response in murine atherosclerosis. Atherosclerosis 2004, 177, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Chen, H.; Xu, Q.; Han, X.; Zhao, Y.; Song, X.; Zhao, T.; Ye, L. The effect of di-2-ethylhexyl phthalate on inflammation and lipid metabolic disorder in rats. Ecotoxicol. Environ. Saf. 2019, 170, 391–398. [Google Scholar] [CrossRef]
- Nakandakari, S.C.B.R.; Muñoz, V.R.; Kuga, G.K.; Gaspar, R.C.; Sant’Ana, M.R.; Pavan, I.C.B.; da Silva, L.G.S.; Morelli, A.P.; Simabuco, F.M.; da Silva, A.S.R.; et al. Short-term high-fat diet modulates several inflammatory, ER stress, and apoptosis markers in the hippocampus of young mice. Brain Behav. Immun. 2019, 79, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Lu, J.; Wang, Y.; Gu, W.; Yang, X.; Yu, J. Intake of total saponins and polysaccharides from Polygonatum kingianum affects the gut microbiota in diabetic rats. Phytomedicine 2017, 26, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Konikoff, T.; Gophna, U. Oscillospira: A Central, Enigmatic Component of the Human Gut Microbiota. Trends Microbiol. 2016, 24, 523–524. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.G.; Wang, T.L.; Yu, S.M.; Liang, J.; Kuang, H.X. Structural characteristics and hepatoprotective potential of Aralia elata root bark polysaccharides and their effects on SCFAs produced by intestinal flora metabolism. Carbohydr. Polym. 2019, 207, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Ai, C.; Duan, M.; Ma, N.; Sun, X.; Yang, J.; Wen, C.; Sun, Y.; Zhao, N.; Song, S. Sulfated polysaccharides from pacific abalone reduce diet-induced obesity by modulating the gut microbiota. J. Funct. Foods 2018, 47, 211–219. [Google Scholar] [CrossRef]
- Wang, Y.; Fei, Y.; Liu, L.; Xiao, Y.; Pang, Y.; Kang, J.; Wang, Z. Polygonatum odoratum Polysaccharides Modulate Gut Microbiota and Mitigate Experimentally Induced Obesity in Rats. Int. J. Mol. Sci. 2018, 19, 3587. [Google Scholar] [CrossRef]
- Cao, J.; Devaraj, S. Recent AHA/ACC guidelines on cholesterol management expands the role of the clinical laboratory. Clinica Chimica Acta 2019, 495, 82–84. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yang, H.; Lu, Y. Capsaicin alleviates lipid metabolism disorder in high beef fat-fed mice. J. Funct. Foods 2019, 60, 103444. [Google Scholar] [CrossRef]
- Ren, R.; Gong, J.; Zhao, Y.; Zhuang, X.; Ye, Y.; Huang, F.; Lin, W. Sulfated polysaccharide from Enteromorpha prolifera suppresses SREBP-1c and ACC expression to lower serum triglycerides in high-fat diet-induced hyperlipidaemic rats. J. Funct. Foods 2018, 40, 722–728. [Google Scholar] [CrossRef]
- Jay, A.G.; Hamilton, J.A. The enigmatic membrane fatty acid transporter CD36: New insights into fatty acid binding and their effects on uptake of oxidized LDL. Prostaglandins Leukot. Essent. Fatty Acids 2018, 138, 64–70. [Google Scholar] [CrossRef]
- Shirpoor, A.; Heshmati, E.; Kheradmand, F.; Gharalari, F.H.; Chodari, L.; Naderi, R.; Majd, F.N.; Samadi, M. Increased hepatic FAT/CD36, PTP1B and decreased HNF4A expression contributes to dyslipidemia associated with ethanol-induced liver dysfunction: Rescue effect of ginger extract. Biomed. Pharmacother. 2018, 105, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, K.; Karin, M. NF-kappaB, inflammation, immunity and cancer: Coming of age. Nat. Rev. Immunol. 2018, 18, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Nepomuceno, R.; Vallerini, B.d.F.; da Silva, R.L.; Corbi, S.C.T.; Bastos, A.d.S.; dos Santos, R.A.; Takahashi, C.S.; Regina P Orrico, S.; Scarel-Caminaga, R.M. Systemic expression of genes related to inflammation and lipid metabolism in patients with dyslipidemia, type 2 diabetes mellitus and chronic periodontitis. Diabetes Metab. Syndr. 2019, 13, 2715–2722. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Wang, Y.; Han, Y.; Zhao, J.; Han, F.; Song, G.; Jiang, P.; Miao, H. Effects of Whole-Grain Rice and Wheat on Composition of Gut Microbiota and Short-Chain Fatty Acids in Rats. J. Agric. Food Chem. 2018, 66, 6326–6335. [Google Scholar] [CrossRef] [PubMed]








| Groups | TNF-α (pg/mL) | IL-6 (pg/mL) | IL-10 (pg/mL) | IL-12 (pg/mL) |
|---|---|---|---|---|
| ND | 84.71 ± 1.48 b | 151.97 ± 6.73 b | 13.22 ± 4.64 ab | 24.30 ± 4.79 a |
| HFD | 117.63 ± 19.46 c | 207.38 ± 24.50 c | 9.65 ± 0.80 a | 34.71 ± 3.50 b |
| HLP-L | 48.91 ± 16.11 a | 140.44 ± 7.67 b | 15.97 ± 3.81 ab | 25.32 ± 4.40 a |
| HLP-H | 42.82 ± 7.49 a | 138.05 ± 20.72 a | 17.49 ± 3.85 b | 21.89 ± 6.66 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, Y.; Liu, Q.; Zhao, F.; Cao, J.; Shen, X.; Li, C. Holothuria Leucospilota Polysaccharides Ameliorate Hyperlipidemia in High-Fat Diet-Induced Rats via Short-Chain Fatty Acids Production and Lipid Metabolism Regulation. Int. J. Mol. Sci. 2019, 20, 4738. https://doi.org/10.3390/ijms20194738
Yuan Y, Liu Q, Zhao F, Cao J, Shen X, Li C. Holothuria Leucospilota Polysaccharides Ameliorate Hyperlipidemia in High-Fat Diet-Induced Rats via Short-Chain Fatty Acids Production and Lipid Metabolism Regulation. International Journal of Molecular Sciences. 2019; 20(19):4738. https://doi.org/10.3390/ijms20194738
Chicago/Turabian StyleYuan, Yiqiong, Qibing Liu, Fuqiang Zhao, Jun Cao, Xuanri Shen, and Chuan Li. 2019. "Holothuria Leucospilota Polysaccharides Ameliorate Hyperlipidemia in High-Fat Diet-Induced Rats via Short-Chain Fatty Acids Production and Lipid Metabolism Regulation" International Journal of Molecular Sciences 20, no. 19: 4738. https://doi.org/10.3390/ijms20194738
APA StyleYuan, Y., Liu, Q., Zhao, F., Cao, J., Shen, X., & Li, C. (2019). Holothuria Leucospilota Polysaccharides Ameliorate Hyperlipidemia in High-Fat Diet-Induced Rats via Short-Chain Fatty Acids Production and Lipid Metabolism Regulation. International Journal of Molecular Sciences, 20(19), 4738. https://doi.org/10.3390/ijms20194738