Melatonin Improves Oocyte Maturation and Mitochondrial Functions by Reducing Bisphenol A-Derived Superoxide in Porcine Oocytes In Vitro

 ,
, 
Abstract
1. Introduction
2. Results
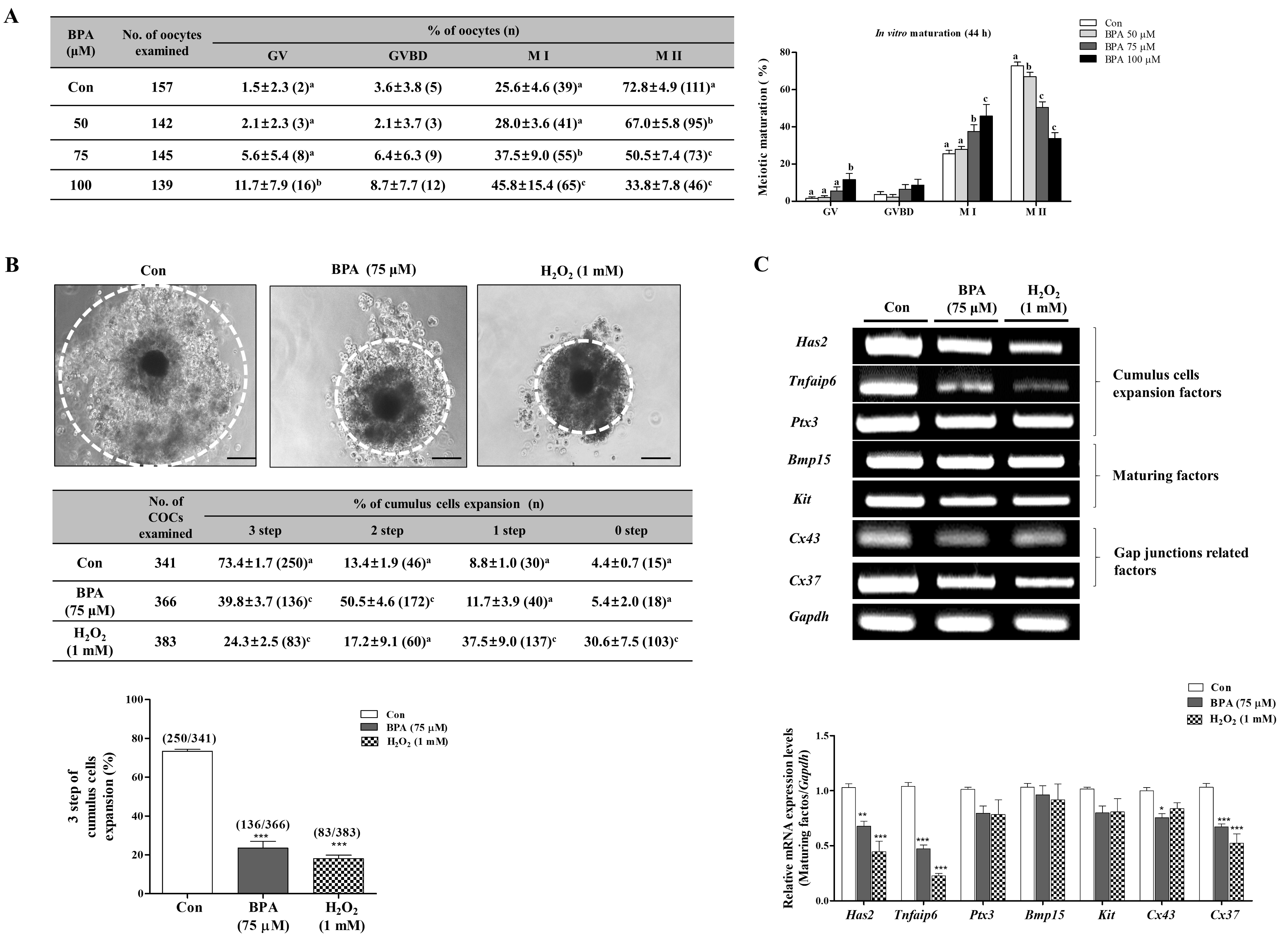
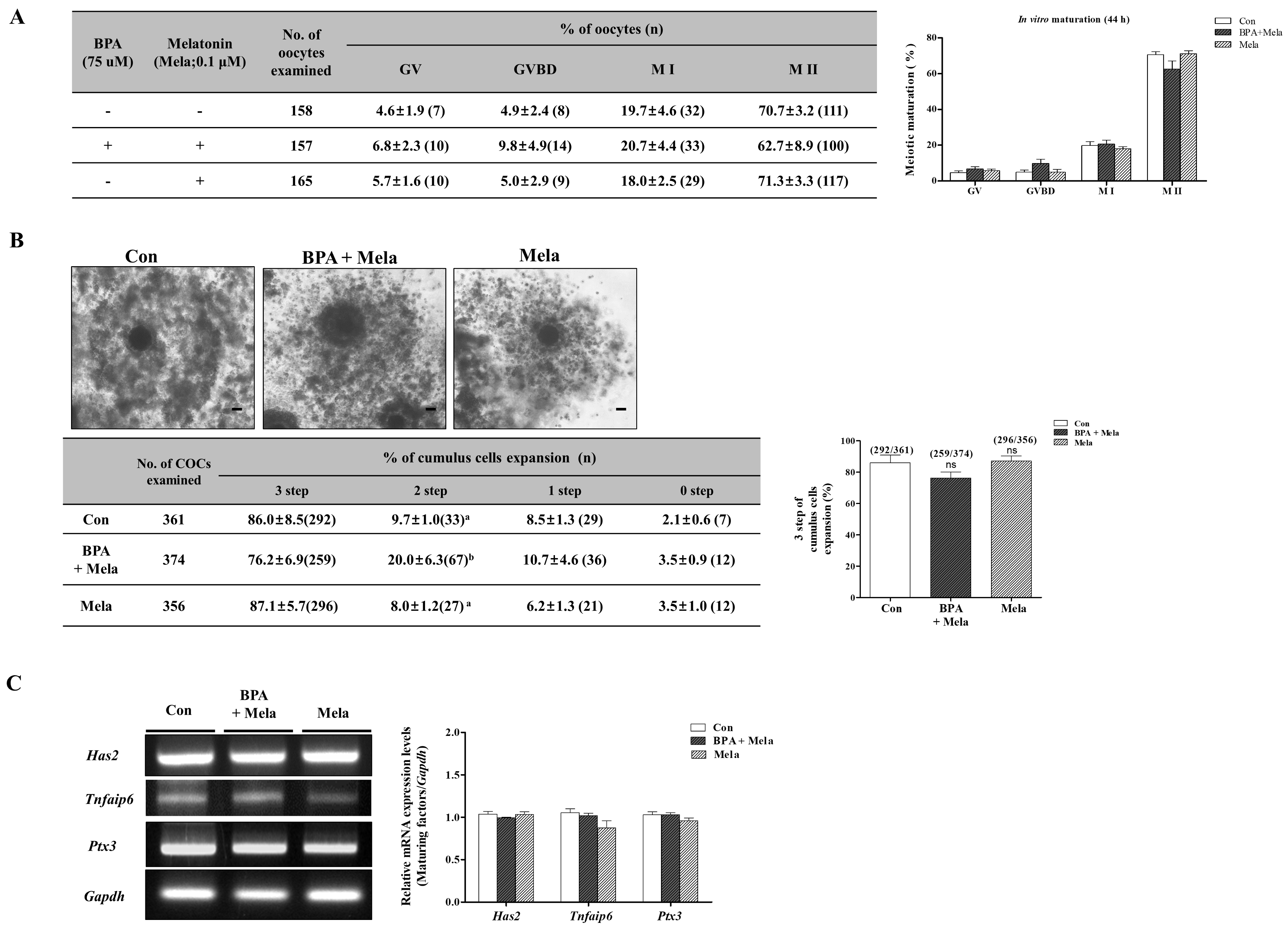
2.1. Effect of BPA Exposure on Meiotic Maturation and Cumulus cell Expansion in Porcine Oocytes
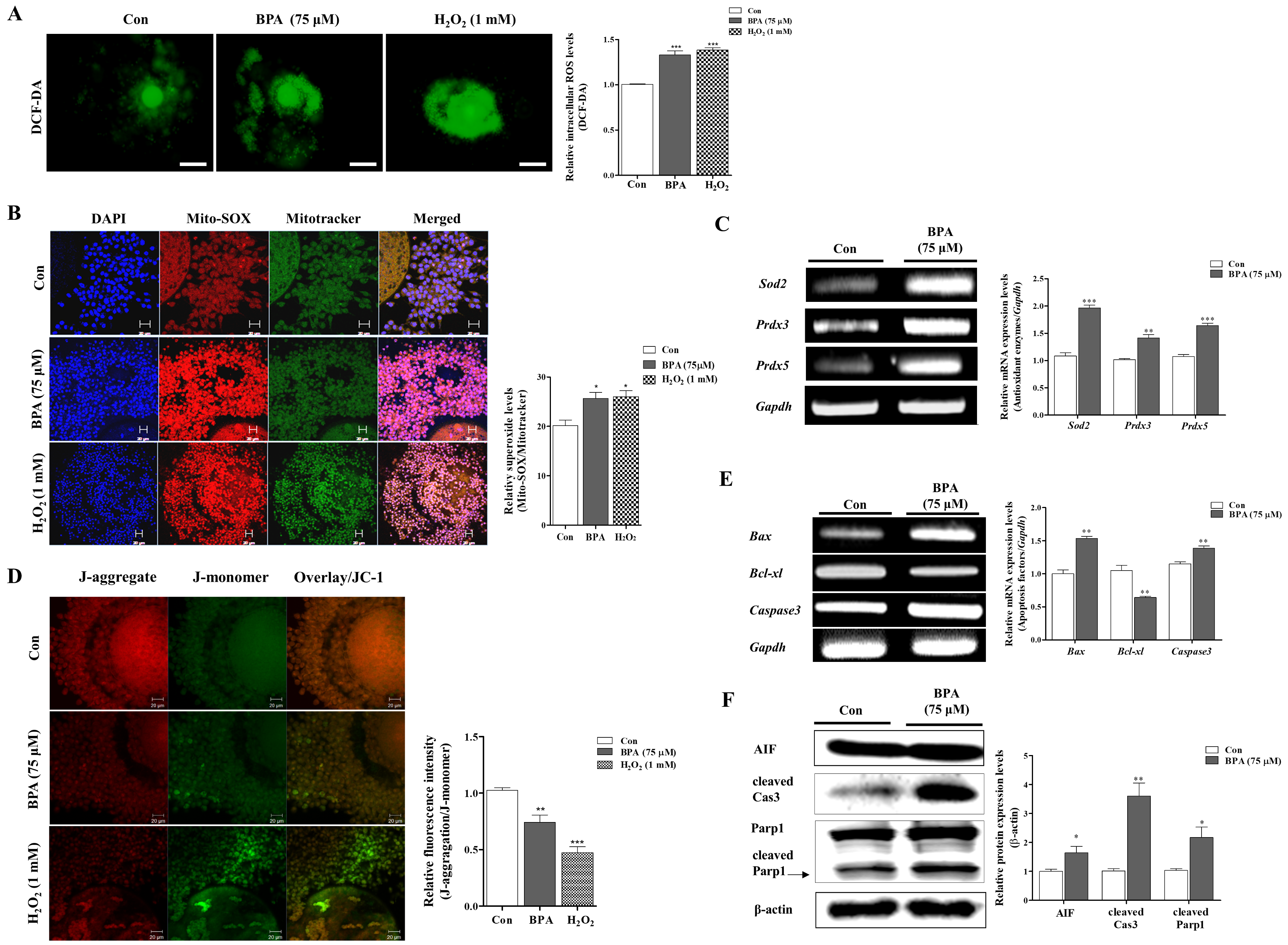
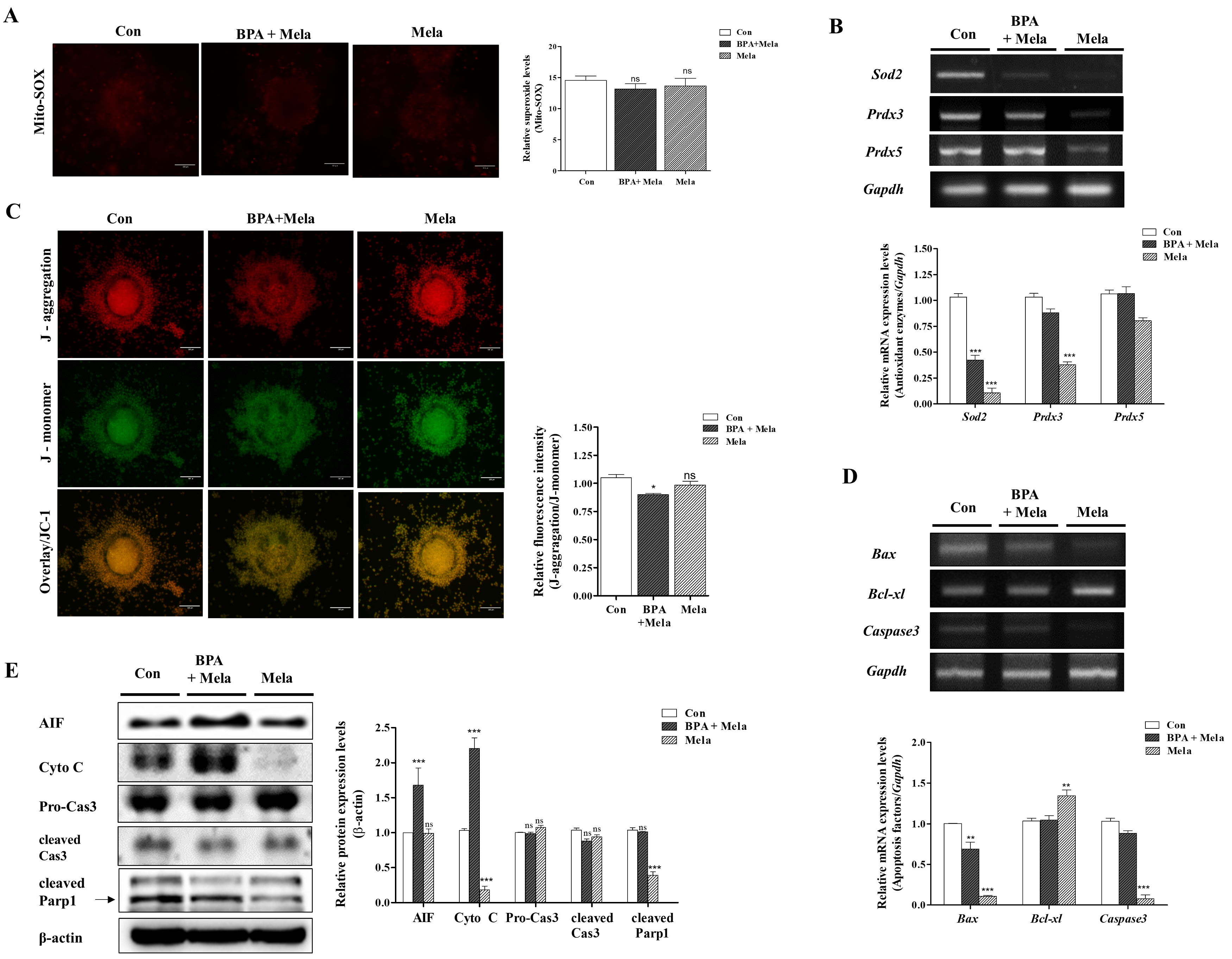
2.2. Measurement of BPA-Induced ROS Production, Mitochondrial Function, and Apoptosis in Porcine Matured COCs
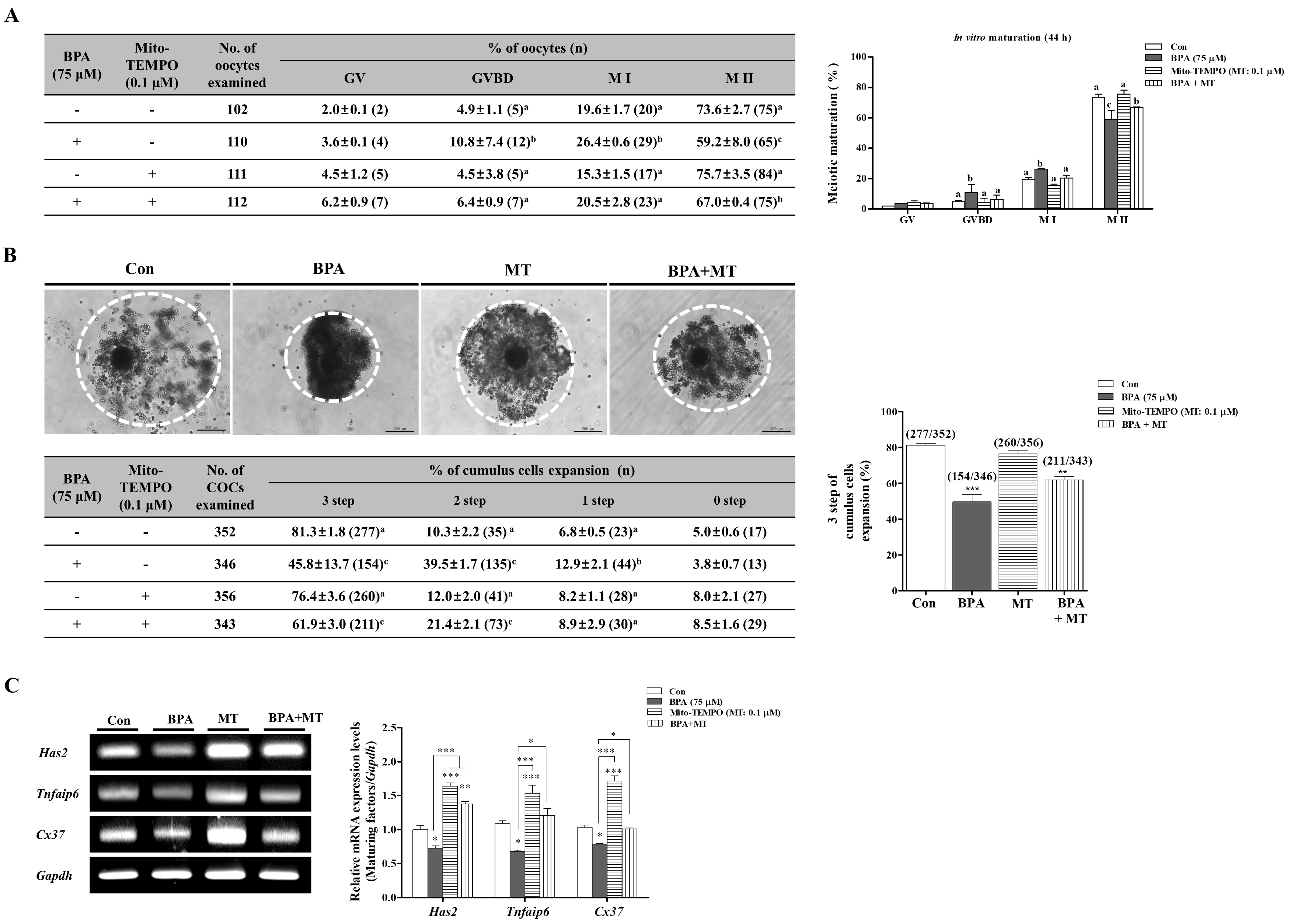
2.3. BPA-Induced ROS Production or Mitochondrial Damage in Porcine COCs Is Recovered by Mito-TEMPO, a Specific Superoxide Scavenger
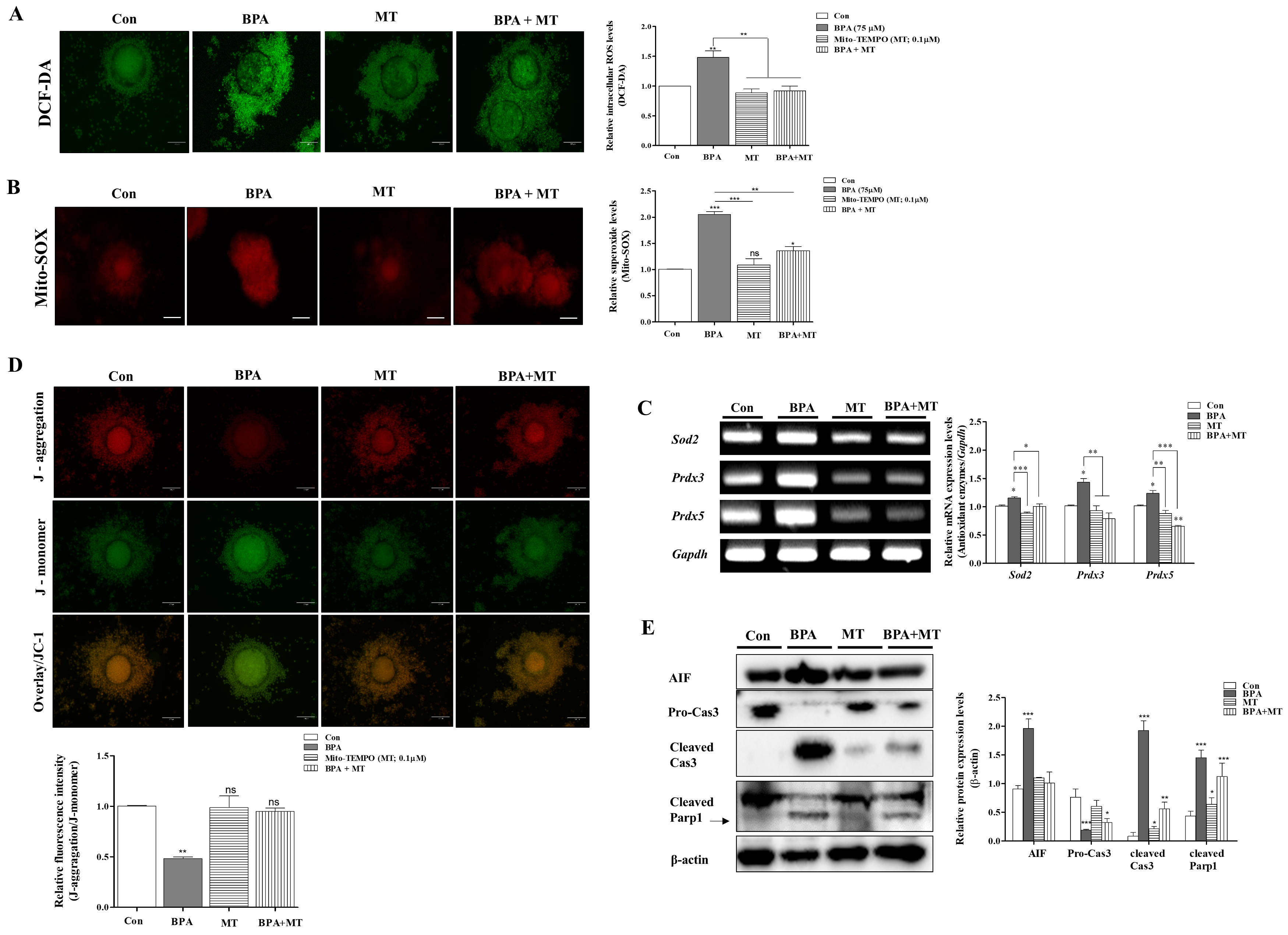
2.4. Protective Mito-TEMPO-Like Effects of Melatonin Against BPA-Induced ROS Production and Mitochondrial Damage in Porcine COCs
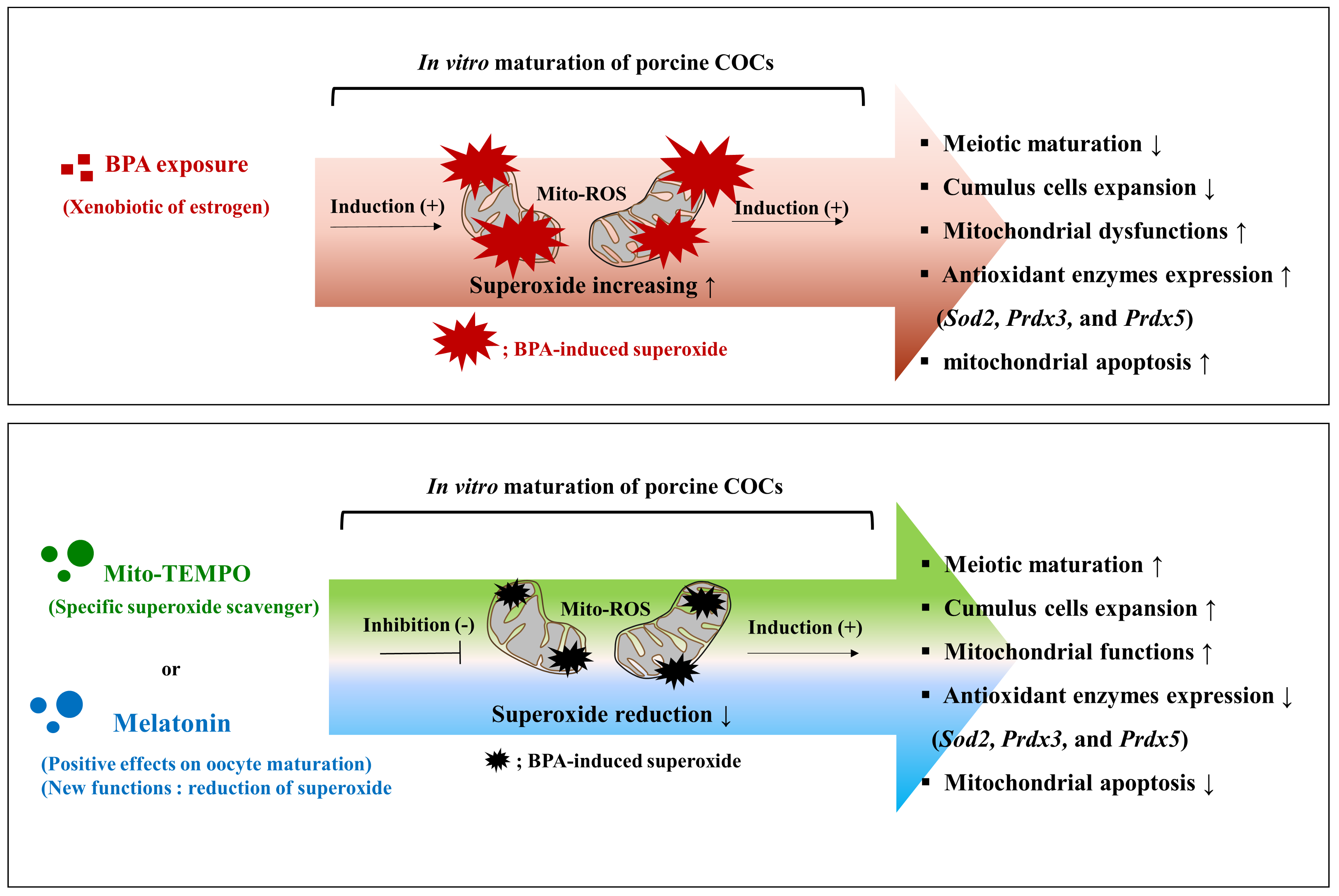
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. In Vitro Maturation (IVM)
4.3. Assessments of Cumulus Cell Expansion and Acetic-Orcein Staining
4.4. RNA Extraction and Reverse Transcription (RT)—PCR
4.5. Staining of DCF-DA, Mito-SOX and Mitotracker
4.6. Measurement of Mitochondrial Membrane Potential (MMP)
4.7. Protein Extraction and Western Blot Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fujii, J.; Iuchi, Y.; Okada, F. Fundamental roles of reactive oxygen species and protective mechanisms in the female reproductive system. Reprod. Biol. Endocrinol. RB&E 2005, 3, 43. [Google Scholar]
- Guerin, P.; El Mouatassim, S.; Menezo, Y. Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Tatemoto, H.; Muto, N.; Sunagawa, I.; Shinjo, A.; Nakada, T. Protection of porcine oocytes against cell damage caused by oxidative stress during in vitro maturation: Role of superoxide dismutase activity in porcine follicular fluid. Biol. Reprod. 2004, 71, 1150–1157. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.A.; Bilodeau, J.F.; Sirard, M.A. Antioxidant requirements for bovine oocytes varies during in vitro maturation, fertilization and development. Theriogenology 2003, 59, 939–949. [Google Scholar] [CrossRef]
- Tiwari, M.; Prasad, S.; Tripathi, A.; Pandey, A.N.; Singh, A.K.; Shrivastav, T.G.; Chaube, S.K. Involvement of reactive oxygen species in meiotic cell cycle regulation and apoptosis in mammalian oocytes. React. Oxyg. Species 2016, 1, 110–116. [Google Scholar] [CrossRef]
- Whitaker, B.D.; Knight, J.W. Mechanisms of oxidative stress in porcine oocytes and the role of anti-oxidants. Reprod. Fertil. Dev. 2008, 20, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, G.M.; Morado, S.A.; Soto, M.P.; Dalvit, G.C.; Cetica, P.D. The control of reactive oxygen species influences porcine oocyte in vitro maturation. Reprod. Domest. Anim. 2015, 50, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Sturmey, R.G.; Leese, H.J. Energy metabolism in pig oocytes and early embryos. Reproduction 2003, 126, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Goud, A.P.; Goud, P.T.; Diamond, M.P.; Gonik, B.; Abu-Soud, H.M. Reactive oxygen species and oocyte aging: Role of superoxide, hydrogen peroxide, and hypochlorous acid. Free Radic. Biol. Med. 2008, 44, 1295–1304. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Dumollard, R.; Rossbach, A.; Lai, F.A.; Swann, K. Redistribution of mitochondria leads to bursts of ATP production during spontaneous mouse oocyte maturation. J. Cell. Physiol. 2010, 224, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Torner, H.; Brussow, K.P.; Alm, H.; Ratky, J.; Pohland, R.; Tuchscherer, A.; Kanitz, W. Mitochondrial aggregation patterns and activity in porcine oocytes and apoptosis in surrounding cumulus cells depends on the stage of pre-ovulatory maturation. Theriogenology 2004, 61, 1675–1689. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.J.; Liu, C.; Duan, W.X.; Xu, S.C.; He, M.D.; Chen, C.H.; Wang, Y.; Zhou, Z.; Yu, Z.P.; Zhang, L.; et al. Melatonin ameliorates bisphenol A-induced DNA damage in the germ cells of adult male rats. Mutat. Res. 2013, 752, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Kurian, J.R.; Keen, K.L.; Kenealy, B.P.; Garcia, J.P.; Hedman, C.J.; Terasawa, E. Acute Influences of Bisphenol A Exposure on Hypothalamic Release of Gonadotropin-Releasing Hormone and Kisspeptin in Female Rhesus Monkeys. Endocrinology 2015, 156, 2563–2570. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.; Bianchi, M.; Lux-Lantos, V.; Libertun, C. Neonatal exposure to bisphenol a alters reproductive parameters and gonadotropin releasing hormone signaling in female rats. Environ. Health Perspect. 2009, 117, 757–762. [Google Scholar] [CrossRef] [PubMed]
- Mlynarcikova, A.; Nagyova, E.; Fickova, M.; Scsukova, S. Effects of selected endocrine disruptors on meiotic maturation, cumulus expansion, synthesis of hyaluronan and progesterone by porcine oocyte-cumulus complexes. Toxicol. In Vitro Int. J. Publ. Assoc. BIBRA 2009, 23, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Murray, T.J.; Maffini, M.V.; Ucci, A.A.; Sonnenschein, C.; Soto, A.M. Induction of mammary gland ductal hyperplasias and carcinoma in situ following fetal bisphenol A exposure. Reprod. Toxicol. 2007, 23, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Mlynarcikova, A.; Kolena, J.; Fickova, M.; Scsukova, S. Alterations in steroid hormone production by porcine ovarian granulosa cells caused by bisphenol A and bisphenol A dimethacrylate. Mol. Cell. Endocrinol. 2005, 244, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Urriola-Munoz, P.; Lagos-Cabre, R.; Patino-Garcia, D.; Reyes, J.G.; Moreno, R.D. Bisphenol-A and Nonylphenol Induce Apoptosis in Reproductive Tract Cancer Cell Lines by the Activation of ADAM17. Int. J. Mol. Sci. 2018, 19, 2238. [Google Scholar] [CrossRef] [PubMed]
- Bilancio, A.; Bontempo, P.; Di Donato, M.; Conte, M.; Giovannelli, P.; Altucci, L.; Migliaccio, A.; Castoria, G. Bisphenol A induces cell cycle arrest in primary and prostate cancer cells through EGFR/ERK/p53 signaling pathway activation. Oncotarget 2017, 8, 115620–115631. [Google Scholar] [CrossRef] [PubMed]
- Di Donato, M.; Cernera, G.; Giovannelli, P.; Galasso, G.; Bilancio, A.; Migliaccio, A.; Castoria, G. Recent advances on bisphenol-A and endocrine disruptor effects on human prostate cancer. Mol. Cell. Endocrinol. 2017, 457, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Han, J.; Duan, X.; Xiong, B.; Cui, X.S.; Kim, N.H.; Liu, H.L.; Sun, S.C. The toxic effects and possible mechanisms of Bisphenol A on oocyte maturation of porcine in vitro. Oncotarget 2016, 7, 32554–32565. [Google Scholar] [CrossRef] [PubMed]
- Huc, L.; Lemarie, A.; Gueraud, F.; Helies-Toussaint, C. Low concentrations of bisphenol A induce lipid accumulation mediated by the production of reactive oxygen species in the mitochondria of HepG2 cells. Toxicol. In Vitro Int. J. Publ. Assoc. BIBRA 2012, 26, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Ni, R.; Cao, T.; Xiong, S.; Ma, J.; Fan, G.C.; Lacefield, J.C.; Lu, Y.; Le Tissier, S.; Peng, T. Therapeutic inhibition of mitochondrial reactive oxygen species with mito-TEMPO reduces diabetic cardiomyopathy. Free Radic. Biol. Med. 2016, 90, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.G.; Park, H.J.; Kim, J.W.; Jung, J.M.; Kim, M.J.; Jegal, H.G.; Kim, I.S.; Kang, M.J.; Wee, G.; Yang, H.Y.; et al. Mito-TEMPO improves development competence by reducing superoxide in preimplantation porcine embryos. Sci. Rep. 2018, 8, 10130. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Park, S.M.; Lee, E.; Kim, J.H.; Jeong, Y.I.; Lee, J.Y.; Park, S.W.; Kim, H.S.; Hossein, M.S.; Jeong, Y.W.; et al. Anti-apoptotic effect of melatonin on preimplantation development of porcine parthenogenetic embryos. Mol. Reprod. Dev. 2008, 75, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- El-Raey, M.; Geshi, M.; Somfai, T.; Kaneda, M.; Hirako, M.; Abdel-Ghaffar, A.E.; Sosa, G.A.; El-Roos, M.E.; Nagai, T. Evidence of melatonin synthesis in the cumulus oocyte complexes and its role in enhancing oocyte maturation in vitro in cattle. Mol. Reprod. Dev. 2011, 78, 250–262. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.M.; Tian, X.Z.; Zhou, G.B.; Wang, L.; Gao, C.; Zhu, S.E.; Zeng, S.M.; Tian, J.H.; Liu, G.S. Melatonin exists in porcine follicular fluid and improves in vitro maturation and parthenogenetic development of porcine oocytes. J. Pineal Res. 2009, 47, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Othman, A.I.; Edrees, G.M.; El-Missiry, M.A.; Ali, D.A.; Aboel-Nour, M.; Dabdoub, B.R. Melatonin controlled apoptosis and protected the testes and sperm quality against bisphenol A-induced oxidative toxicity. Toxicol. Ind. Health 2016, 32, 1537–1549. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Takasaki, A.; Taketani, T.; Tanabe, M.; Kizuka, F.; Lee, L.; Tamura, I.; Maekawa, R.; Aasada, H.; Yamagata, Y.; et al. The role of melatonin as an antioxidant in the follicle. J. Ovarian Res. 2012, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, R.B.; Lane, M.; Thompson, J.G. Oocyte-secreted factors: Regulators of cumulus cell function and oocyte quality. Hum. Reprod. Update 2008, 14, 159–177. [Google Scholar] [CrossRef] [PubMed]
- Kidder, G.M.; Mhawi, A.A. Gap junctions and ovarian folliculogenesis. Reproduction 2002, 123, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Park, J.Y.; Kim, J.W.; Yang, S.G.; Jung, J.M.; Kim, M.J.; Kang, M.J.; Cho, Y.H.; Wee, G.; Yang, H.Y.; et al. Melatonin improves the meiotic maturation of porcine oocytes by reducing endoplasmic reticulum stress during in vitro maturation. J. Pineal Res. 2018, 64, e12458. [Google Scholar] [CrossRef] [PubMed]
- Krisher, R.L.; Brad, A.M.; Herrick, J.R.; Sparman, M.L.; Swain, J.E. A comparative analysis of metabolism and viability in porcine oocytes during in vitro maturation. Anim. Reprod. Sci. 2007, 98, 72–96. [Google Scholar] [CrossRef] [PubMed]
- Boruszewska, D.; Sinderewicz, E.; Kowalczyk-Zieba, I.; Grycmacher, K.; Woclawek-Potocka, I. The effect of lysophosphatidic acid during in vitro maturation of bovine cumulus-oocyte complexes: Cumulus expansion, glucose metabolism and expression of genes involved in the ovulatory cascade, oocyte and blastocyst competence. Reprod. Biol. Endocrinol. RB&E 2015, 13, 44. [Google Scholar]
- Dragovic, R.A.; Ritter, L.J.; Schulz, S.J.; Amato, F.; Thompson, J.G.; Armstrong, D.T.; Gilchrist, R.B. Oocyte-secreted factor activation of SMAD 2/3 signaling enables initiation of mouse cumulus cell expansion. Biol. Reprod. 2007, 76, 848–857. [Google Scholar] [CrossRef] [PubMed]
- Richard, S.; Baltz, J.M. Prophase I arrest of mouse oocytes mediated by natriuretic peptide precursor C requires GJA1 (connexin-43) and GJA4 (connexin-37) gap junctions in the antral follicle and cumulus-oocyte complex. Biol. Reprod. 2014, 90, 137. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, R.B.; Ritter, L.J.; Armstrong, D.T. Oocyte-somatic cell interactions during follicle development in mammals. Anim. Reprod. Sci. 2004, 82–83, 431–446. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, Y.; Suzuki, K.; Yoneda, A.; Watanabe, T. Effects of oxygen concentration and antioxidants on the in vitro developmental ability, production of reactive oxygen species (ROS), and DNA fragmentation in porcine embryos. Theriogenology 2004, 62, 1186–1197. [Google Scholar] [CrossRef] [PubMed]
- Combelles, C.M.; Gupta, S.; Agarwal, A. Could oxidative stress influence the in-vitro maturation of oocytes? Reprod. Biomed. Online 2009, 18, 864–880. [Google Scholar] [CrossRef]
- Takahashi, M. Oxidative stress and redox regulation on in vitro development of mammalian embryos. J. Reprod. Dev. 2012, 58, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Barbonetti, A.; Castellini, C.; Di Giammarco, N.; Santilli, G.; Francavilla, S.; Francavilla, F. In vitro exposure of human spermatozoa to bisphenol A induces pro-oxidative/apoptotic mitochondrial dysfunction. Reprod. Toxicol. 2016, 66, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Luo, C.; Li, Q.; Chen, S.; Hu, Y. Mitochondrion-mediated apoptosis is involved in reproductive damage caused by BPA in male rats. Environ. Toxicol. Pharmacol. 2014, 38, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchester, L.C.; Reiter, R.J.; Plummer, B.F.; Limson, J.; Weintraub, S.T.; Qi, W. Melatonin directly scavenges hydrogen peroxide: A potentially new metabolic pathway of melatonin biotransformation. Free Radic. Biol. Med. 2000, 29, 1177–1185. [Google Scholar] [CrossRef]
- Mehrzadi, S.; Safa, M.; Kamrava, S.K.; Darabi, R.; Hayat, P.; Motevalian, M. Protective mechanisms of melatonin against hydrogen-peroxide-induced toxicity in human bone-marrow-derived mesenchymal stem cells. Can. J. Physiol. Pharmacol. 2017, 95, 773–786. [Google Scholar] [CrossRef] [PubMed]
- Chetsawang, B.; Putthaprasart, C.; Phansuwan-Pujito, P.; Govitrapong, P. Melatonin protects against hydrogen peroxide-induced cell death signaling in SH-SY5Y cultured cells: Involvement of nuclear factor kappa B, Bax and Bcl-2. J. Pineal Res. 2006, 41, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Jiang, X.; Liu, Y. Degradation of bisphenol A and formation of hydrogen peroxide induced by glow discharge plasma in aqueous solutions. J. Hazard. Mater. 2008, 154, 1106–1114. [Google Scholar] [CrossRef] [PubMed]
- Leem, Y.H.; Oh, S.; Kang, H.J.; Kim, J.H.; Yoon, J.; Chang, J.S. BPA-toxicity via superoxide anion overload and a deficit in beta-catenin signaling in human bone mesenchymal stem cells. Environ. Toxicol. 2017, 32, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Ooe, H.; Taira, T.; Iguchi-Ariga, S.M.; Ariga, H. Induction of reactive oxygen species by bisphenol A and abrogation of bisphenol A-induced cell injury by DJ-1. Toxicol. Sci. Off. J. Soc. Toxicol. 2005, 88, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, M.; Tamura, H.; Taketani, T.; Okada, M.; Lee, L.; Tamura, I.; Maekawa, R.; Asada, H.; Yamagata, Y.; Sugino, N. Melatonin protects the integrity of granulosa cells by reducing oxidative stress in nuclei, mitochondria, and plasma membranes in mice. J. Reprod. Dev. 2015, 61, 35–41. [Google Scholar] [CrossRef] [PubMed]
- El-Missiry, M.A.; Othman, A.I.; Al-Abdan, M.A.; El-Sayed, A.A. Melatonin ameliorates oxidative stress, modulates death receptor pathway proteins, and protects the rat cerebrum against bisphenol-A-induced apoptosis. J. Neurol. Sci. 2014, 347, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Dai, X.; Lu, Y.; Miao, Y.; Zhou, C.; Cui, Z.; Liu, H.; Xiong, B. Melatonin protects oocyte quality from Bisphenol A-induced deterioration in the mouse. J. Pineal Res. 2017, 62, e12396. [Google Scholar] [CrossRef] [PubMed]
- Petters, R.M.; Wells, K.D. Culture of pig embryos. J. Reprod. Fertil. Suppl. 1993, 48, 61–73. [Google Scholar] [PubMed]
- Salavati, M.; Ghafari, F.; Zhang, T.; Fouladi-Nashta, A.A. Effects of oxygen concentration on in vitro maturation of canine oocytes in a chemically defined serum-free medium. Reproduction 2012, 144, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.H.; Park, H.J.; Kim, J.W.; Park, J.Y.; Park, S.Y.; Yang, S.G.; Kim, C.H.; Yoon, S.B.; Kim, S.U.; Chang, K.T.; et al. Tyrphostin A9 improves blastocyst development in porcine embryos through induction of dynamin-related protein 1-dependent mitochondrial fission. Mitochondrion 2017, 35, 80–86. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequences | Tm °C | Gene Bank Accession No. | Base Pairs |
|---|---|---|---|---|
| Has2 | F(5′–3′): TGGCTGTACAATGCGATGTG R(5′–3′): TGGGTGGTGTGATTTTCACC | 55 | (NM_214053.1) | 402 |
| Tnfaip6 | F(5′–3′): TCTTCCTGTGGGAAGAGGCT R(5′–3′): GTCCGTCTGAACAGAAGCGA | 55 | (NM_001159607.1) | 337 |
| Ptx3 | F(5′–3′): TCAGTGCCTGCATTTGGGTC R(5′–3′): TTCTGAACAAGGGCATGTAG | 58 | (NM_001244783.1) | 225 |
| Bmp15 | F(5′–3′): GGATCCAGAAAAGCACAACC R(5′–3′): GCATTTCATCCCTGGACACT | 55 | (NM_001005155.1) | 227 |
| Kit | F(5′–3′): TGTATTCACAGAGACTTGGCGG R(5′–3′): TTACGTGGTCAAAGGAAACG | 55 | (NM_001044525.1) | 124 |
| Cx43 | F(5′–3′): ACTGAGCCCCTCCAAAGACT R(5′–3′): GGCTAATTACAGTGCCGAGC | 55 | (NM_001244212.1) | 191 |
| Cx37 | F(5′–3′): TTCCTTGAGAAGCTGCTGGA R(5′–3′): CATCTCCCACATCCGCTACT | 55 | (NM_001244224.1) | 217 |
| Gapdh | F(5′–3′): TCGGAGTGAACGGATTTC R(5′–3′): CCTGGAAGATGGTGATGG | 53.7 | (NM_001206359.1) | 230 |
| Genes | Primer Sequences | Tm °C | Gene Bank Accession No. | Base Pairs |
|---|---|---|---|---|
| Gpx1 | F(5′–3′): CACAACGGTGCGGGACTA R(5′–3′): GTCTCCAGTGTGTCGCAATG | 54 | (NM_214201.1) | 326 |
| Catalase | F(5′–3′): CGAAGGCGAAGGTGTTTG R(5′–3′): GGATATGGATCGCACACT | 50 | (NM_214301.2) | 374 |
| Sod1 | F(5′–3′): GCCAAAGGATCAAGAGAGGC R(5′–3′): GTCGTTTGGCCTGTGGTGTA | 55 | (NM_001190422.1) | 226 |
| Sod2 | F(5′–3′): GCAGCTCGAGCAGGAATCTGG R(5′–3′): ACGCGGCCTACGTGAACAA | 59.7 | (NM_214127.2) | 163 |
| Prdx1 | F(5′–3′): AGAAGCAAGGAGGACTGGGA R(5′–3′): GCCTGATGTCCAGAAGAGCA | 55 | (XM_003128042.3) | 300 |
| Prdx2 | F(5′–3′): CACCTGGCTTGGATCAACAC R(5′–3′): TCCAGGCCTTCCAGTACACA | 55 | (NM_001244474.1) | 249 |
| Prdx3 | F(5′–3′): AGTGGATTCCCACTTCAGCC R(5′–3′): AACCCATGGAGAAGTCTGCC | 55.1 | (NM_001244531.1) | 290 |
| Prdx4 | F(5′–3′): AGTTTACCCATCTGGCCTGG R(5′–3′): GTCCTGCTGGTTGGAAACCT | 55 | (XM_005673497.2) | 296 |
| Prdx5 | F(5’–3’): ACCTTCCAGGGTTTGTGGAG R(5′–3′): CCTGAATGTGGAGCCAGATG | 55 | (NM_214144.1) | 285 |
| Prdx6 | F(5′–3′): ATGCCTGTGACAGCTCGTGT R(5′-3′): ACCAAAGAGCTCCCATCTGG | 55.2 | (NM_214408.1) | 263 |
| Gapdh | F(5′–3′): TCGGAGTGAACGGATTTC R(5′–3′): CCTGGAAGATGGTGATGG | 53.7 | (NM_001206359.1) | 230 |
| Genes | Primer Sequences | Tm °C | Gene Bank Accession No. | Base Pairs |
|---|---|---|---|---|
| Bax | F(5′–3′): AAGCGCATTGGAGATGAACT R(5′–3′): CTGGACTTCCTTCGAGATCG | 50 | (XM_003127290.4) | 251 |
| Bcl-xl | F(5′–3′): AGGGCATTCAGTGACCTGAC R(5′–3′): CACCTAGAGCCTTGGATCCA | 55 | (NM_214285.1) | 242 |
| Caspase3 | F(5′–3′): GAGGCAGACTTCTTGTATGC R(5′–3′): TTCCATGTATTGTGTCCATGC | 50 | (NM_214131.1) | 238 |
| Gapdh | F(5′–3′): TCGGAGTGAACGGATTTC R(5′–3′): CCTGGAAGATGGTGATGG | 53.7 | (NM_001206359.1) | 230 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, H.-J.; Park, S.-Y.; Kim, J.-W.; Yang, S.-G.; Kim, M.-J.; Jegal, H.-G.; Kim, I.-S.; Choo, Y.-K.; Koo, D.-B. Melatonin Improves Oocyte Maturation and Mitochondrial Functions by Reducing Bisphenol A-Derived Superoxide in Porcine Oocytes In Vitro. Int. J. Mol. Sci. 2018, 19, 3422. https://doi.org/10.3390/ijms19113422
Park H-J, Park S-Y, Kim J-W, Yang S-G, Kim M-J, Jegal H-G, Kim I-S, Choo Y-K, Koo D-B. Melatonin Improves Oocyte Maturation and Mitochondrial Functions by Reducing Bisphenol A-Derived Superoxide in Porcine Oocytes In Vitro. International Journal of Molecular Sciences. 2018; 19(11):3422. https://doi.org/10.3390/ijms19113422
Chicago/Turabian StylePark, Hyo-Jin, Soo-Yong Park, Jin-Woo Kim, Seul-Gi Yang, Min-Ji Kim, Ho-Guen Jegal, In-Su Kim, Young-Kug Choo, and Deog-Bon Koo. 2018. "Melatonin Improves Oocyte Maturation and Mitochondrial Functions by Reducing Bisphenol A-Derived Superoxide in Porcine Oocytes In Vitro" International Journal of Molecular Sciences 19, no. 11: 3422. https://doi.org/10.3390/ijms19113422
APA StylePark, H.-J., Park, S.-Y., Kim, J.-W., Yang, S.-G., Kim, M.-J., Jegal, H.-G., Kim, I.-S., Choo, Y.-K., & Koo, D.-B. (2018). Melatonin Improves Oocyte Maturation and Mitochondrial Functions by Reducing Bisphenol A-Derived Superoxide in Porcine Oocytes In Vitro. International Journal of Molecular Sciences, 19(11), 3422. https://doi.org/10.3390/ijms19113422