
The Evolution of Total Phenolic Compounds and Antioxidant Activities during Ripening of Grapes (Vitis vinifera L., cv. Tempranillo) Grown in Semiarid Region: Effects of Cluster Thinning and Water Deficit

 ,
, 
Abstract
:1. Introduction
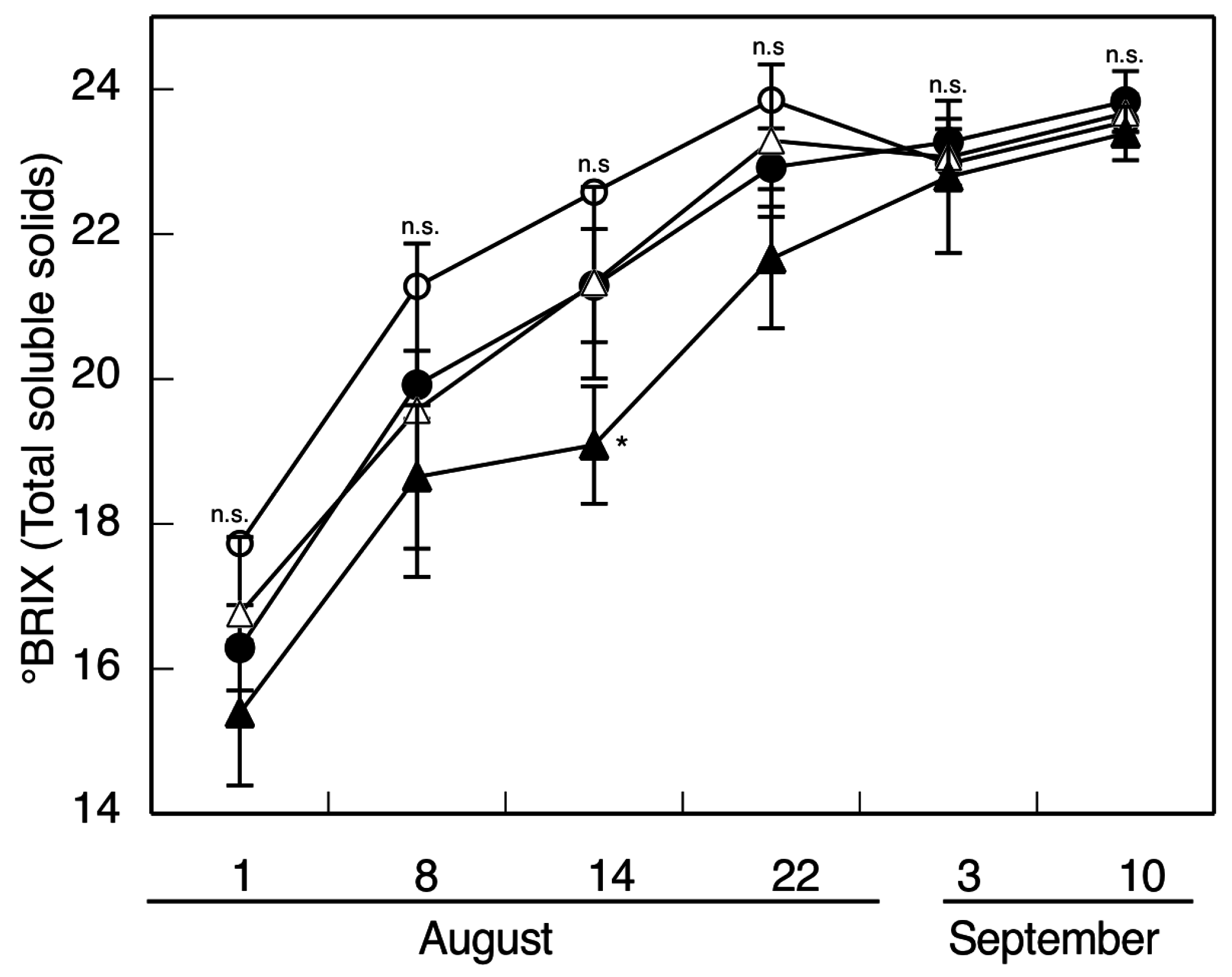
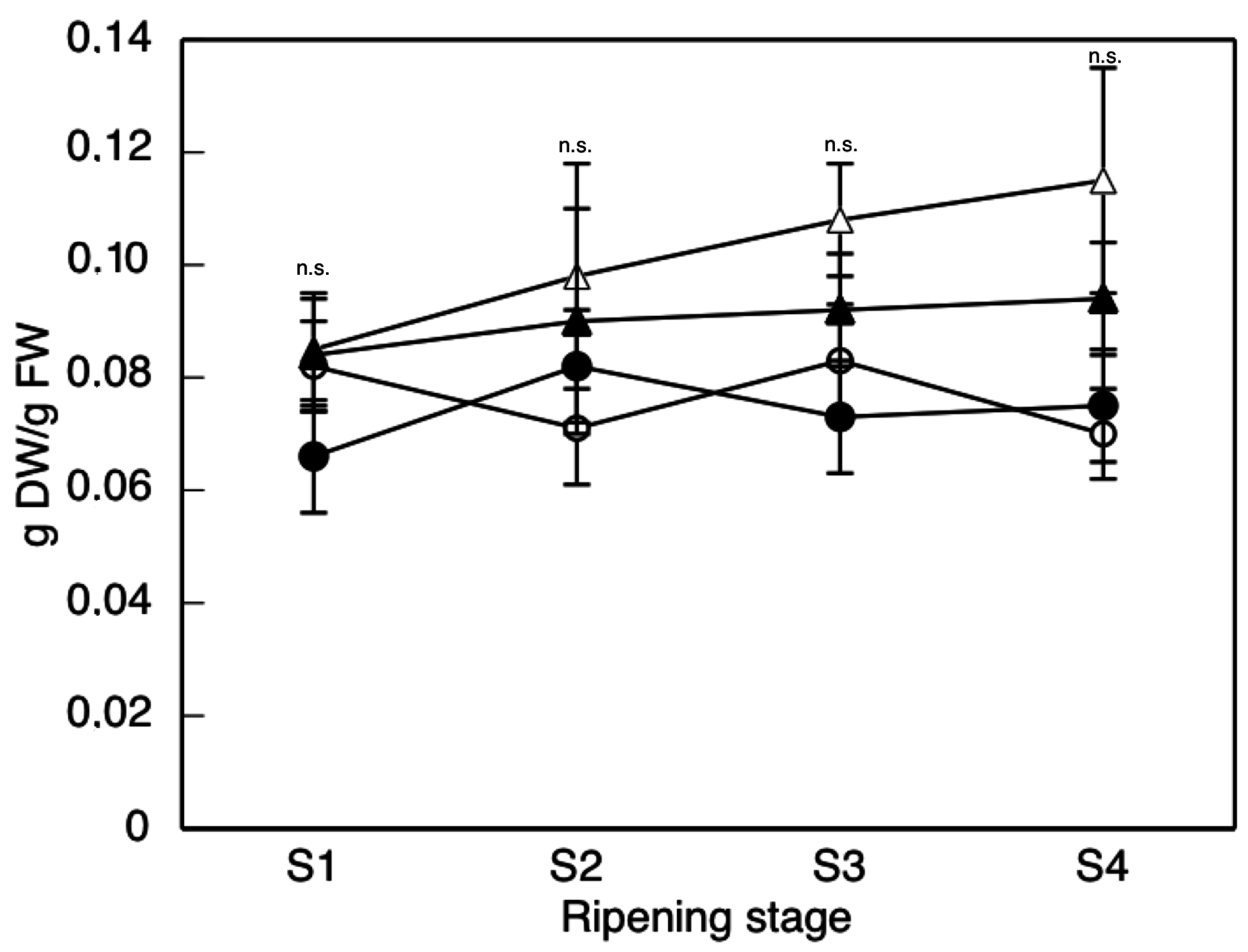
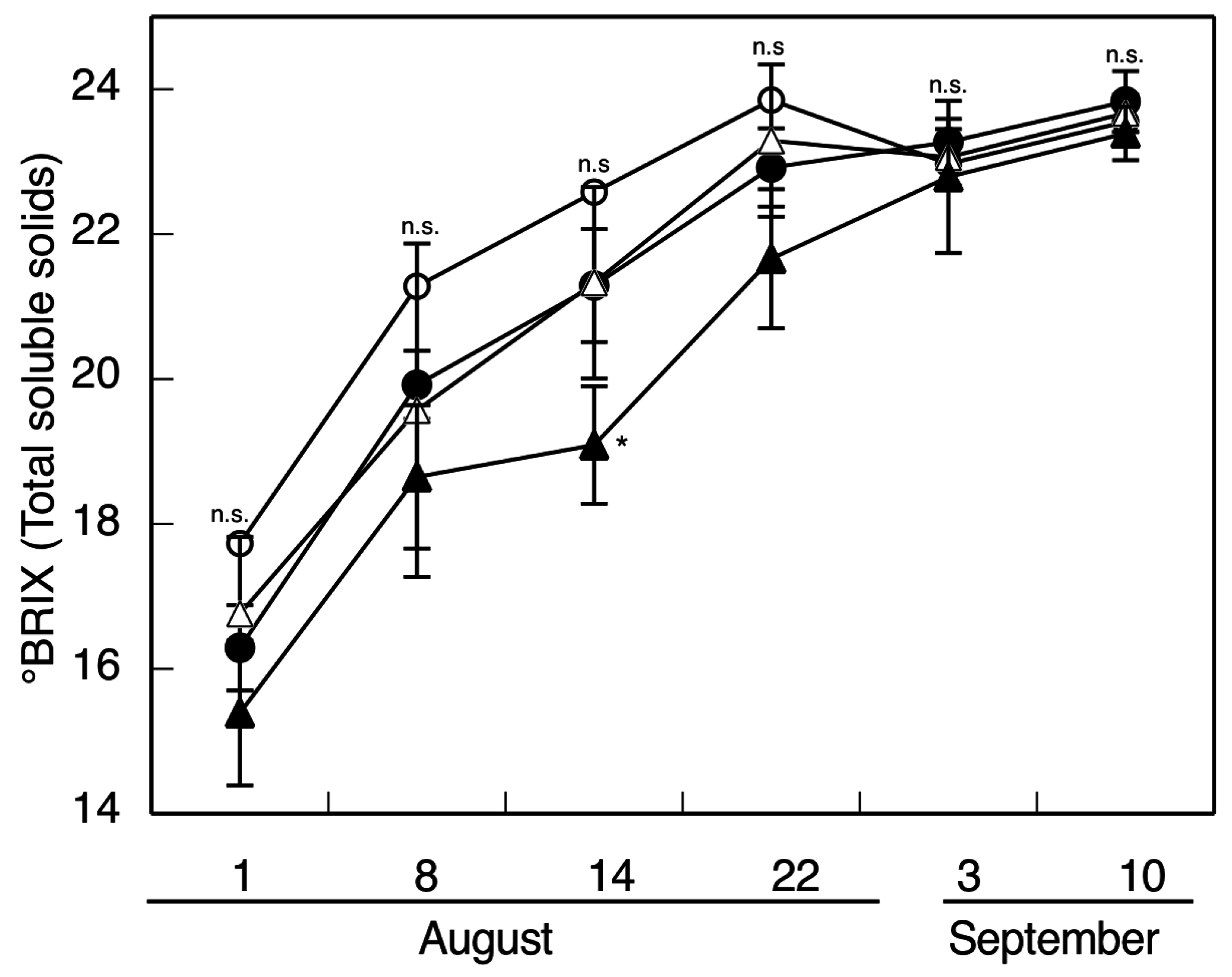
2. Results and Discussion
3. Material and Methods
3.1. Plant Material and Experimental Design
3.2. Biochemical Assay
3.3. Enzymatic Activities
3.4. Statistical Analyses
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chaves, M.M.; Zarrouk, O.; Francisco, R.; Costa, J.M.; Santos, T.; Regalado, A.P.; Rodrigues, M.L.; Lopes, C.M. Grapevine under deficit irrigation—Hints from physiological and molecular data. Ann. Bot. 2010, 105, 661–676. [Google Scholar] [CrossRef] [PubMed]
- Diago, M.P.; Vilanova, M.; Blanco, J.A.; Tardaguila, J. Effects of mechanical thinning on fruit and wine composition and sensory attributes of Grenache and Tempranillo varieties (Vitis vinifera L.). Aust. J. Grape Wine Res. 2010, 16, 314–326. [Google Scholar] [CrossRef]
- Reynard, J.S.; Zufferey, V.; Nicol, G.C.; Murisier, F. Vine water status as a parameter of the “terroir” effect under the non-irrigated conditions of the Vaud viticultural area (Switzerland). J. Int. Sci. Vigne Vin 2011, 45, 139–147. [Google Scholar] [CrossRef]
- Zsofi, Z.; Toth, E.; Rusjan, D.; Balo, B. Terroir aspects of grape quality in a cool climate wine region: Relationship between water deficit, vegetative growth and berry sugar concentration. Sci. Hortic. 2011, 127, 494–499. [Google Scholar] [CrossRef]
- Reynolds, A.G.; Schlosser, J.; Sorokowsky, D.; Roberts, R.; Willwerth, J.; de Savigny, C. Magnitude of viticultural and enological effects. II. Relative impacts of cluster thinning and yeast strain on composition and sensory attributes of Chardonnay musqué. Am. J. Enol. Viticult. 2007, 58, 25–41. [Google Scholar]
- Petri, P.R.; Clingeleffer, P.R. Crop thinning (hand versus mechanical), grape maturity and anthocyanin concentration: Outcomes from irrigated Cabernet Sauvignon (Vitis vinifera L.) in a warm climate. Am. J. Grape Wine 2006, 12, 21–29. [Google Scholar] [CrossRef]
- Valdés, M.E.; Moreno, D.; Gamero, E.; Uriarte, D.; Prieto, M.H.; Manzano, R.; Picón, J.; Intrigliolo, D.S. Effects of cluster thinning and irrigation amount on water relations, growth, yield and fruit and wine composition of Tempranillo grapes in Extremadura (Spain). J. Int. Sci. Vigne Vin 2009, 43, 67–76. [Google Scholar]
- Gil, M.; Esteruelas, M.; González, E.; Kontoudakis, N.; Jiménez, J.; Fort, F.; Zamora, F. Effect of two different treatments for reducing grape yield in Vitis vinifera cv. Syrah on wine composition and quality: Berry thinning versus cluster thinning. J. Agric. Food Chem. 2013, 61, 4968–4978. [Google Scholar] [CrossRef] [PubMed]
- Guidoni, S.; Allara, P.; Schubert, A. Effect of cluster thinning on berry skin anthocyanin composition of Vitis vinifera cv. Nebbiolo. Am. J. Enol. Viticult. 2002, 53, 224–226. [Google Scholar]
- Peña-Neira, A.; Cáceres, A.; Pastenes, C. Low molecular weight phenolic and anthocyanin composition of grape skins from cv. Syrah (Vitis vinifera L.) in the Maipo Valley (Chile): Effect of clusters thinning and vineyard yield. Food Sci. Technol. Int. 2007, 13, 153–158. [Google Scholar] [CrossRef]
- Gil-Muñoz, R.R.; Vila-Lopez, J.I.; Fernandez, J.I.; Martinez-Cutil-Las, A. Effects of cluster thinning on anthocyanin extractability and chromatic parameters of Syrah and Tempranillo grapes and wines. J. Int. Sci. Vigne Vin 2009, 43, 45–53. [Google Scholar] [CrossRef]
- Keller, M.L.; Mills, L.; Wample, R.; Spayd, S. Cluster thinning effects on three deficit-irrigated Vitis vinifera cultivars. Am. J. Enol. Viticult. 2005, 56, 91–103. [Google Scholar]
- Kok, D. Influences of pre- and post-veraison cluster thinning treatments on grape composition variables and monoterpenes levels of Vitis vinifera L. cv. Sauvignon Blanc. J. Food Agric. Environ. 2011, 9, 22–26. [Google Scholar]
- Bubola, M.; Persuric, D.; Ganic, K.K. Impact of cluster thinning on productive characteristics and wine phenolic composition of cv Merlot. J. Food Agric. Environ. 2011, 9, 36–39. [Google Scholar]
- Uriarte, D.; Intrigliolo, D.S.; Mancha, L.A.; Picón-Toro, J.; Valdes, E.; Prieto, M.H. Interactive Effects of Irrigation and Crop Level on Tempranillo Vines in a Semi-Arid Climate. Am. J. Enol. Viticult. 2014, 66, 101–111. [Google Scholar] [CrossRef]
- Preszeler, T.; Schmit, T.M.; Heuvel, J.E.V. A model to establish economically sustainable cluster-thinning practices. Am. J. Enol. Viticult. 2010, 61, 140–146. [Google Scholar]
- Tardáguila, J.; Petrie, P.R.; Poni, S.; Diago, M.P.; de Toda, F.M. Effects of mechanical thinning on yield and fruit composition of Tempranillo and Grenache grapes trained to a vertical shoot-positioned canopy. Am. J. Enol. Viticult. 2008, 59, 412–417. [Google Scholar]
- Roby, G.; Matthews, M.A. Relative proportion of seed, skin and flesh in ripe berries from Cabernet Sauvignon grapevines grown in a vineyard either well irrigated on under water deficit. Aust. J. Grape Wine Res. 2004, 10, 74–82. [Google Scholar] [CrossRef]
- Esteban, M.A.; Villanueva, M.J.; Lisarrague, J.R. Effect of irrigation on changes in the anthocyanin composition of the skin of cv. Tempranillo (Vitis vinifera L.) grape berries during ripening. J. Sci. Food Agric. 2001, 81, 409–420. [Google Scholar] [CrossRef]
- Reynolds, A.G.; Lowrey, W.D.; Tomek, L.; Hakimi, J.; de Savigny, C. Influence of irrigation on vine performance, fruit composition, and wine quality of Chardonnay in a cool, humid climate. Am. J. Enol. Viticult. 2007, 58, 217–228. [Google Scholar]
- Santesteban, L.G.; Miranda, C.; Royo, J.B. Regulated deficit irrigation effects on growth, yield, grape quality and individual anthocyanin composition in Vitis vinifera cv. Tempranillo. Agric. Water Manag. 2011, 98, 1171–1179. [Google Scholar] [CrossRef]
- Gamero, E.; Moreno, D.; Vilanova, M.; Uriarte, D.; Prieto, M.H.; Valdés, M.E. Effect of bunch thinning and water stress on chemical and sensory characteristic of Tempranillo wines. Aust. J. Grape Wine Res. 2014, 20, 394–400. [Google Scholar] [CrossRef]
- Gamero, E.; Moreno, D.; Talaverano, I.; Prieto, M.H.; Guerra, M.T.; Valdés, M.E. Effects of irrigation and cluster thinning on Tempranillo grape and wine composition. S. Afr. J. Enol. Viticult. 2014, 35, 196–204. [Google Scholar] [CrossRef]
- Moreno, D.; Vilanova, M.; Gamero, E.; Intrigliolo, D.S.; Talaverano, M.I.; Uriarte, D.; Valdés, M.E. Effects of preflowering leaf removal on phenolic composition of Tempranillo cv. in semi-arid terroir of western Spain. Am. J. Enol. Viticult. 2014, 66, 204–211. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive Oxygen Species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Physiol. Mol. Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Kawano, T. Roles of the reactive oxygen species generating peroxidase reactions in plant defense and growth induction. Plant Cell Rep. 2003, 21, 829–837. [Google Scholar] [PubMed]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, A.; Romojaro, F.; Gómez, J.M.; Llanos, M.R.; Sevilla, F. Antioxidant systems and their relationship with the response of pepper fruits to storage at 20 °C. J. Sci. Food Agric. 2003, 51, 6293–6299. [Google Scholar] [CrossRef] [PubMed]
- Garrido, I.; Llerena, J.L.; Valdés, M.E.; Mancha, L.A.; Uriarte, D.; Prieto, M.H.; Espinosa, F. Effects of defoliation and water restriction on total phenols and antioxidant activities in grapes during ripening. J. Int. Sci. Vigne Vin 2014, 48, 31–42. [Google Scholar]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Grace, S.C. Phenolics as antioxidants. In Antioxidants and Reactive Oxygen Species in Plants; Smirnoff, N., Grace, S.C., Eds.; Blackwell: Oxford, UK, 2005; pp. 141–168. [Google Scholar]
- Fortes, A.M.; Agudelo-Romero, P.; Silva, M.S.; Ali, K.; Sousa, L.; Maltese, F.; Choi, Y.H.; Grimplet, J.; Martinez-Zapater, J.M.; Verpoorte, R.; et al. Transcript and metabolite analysis in Trincadeira cultivar reveals novel information regarding the dynamics of grape ripening. BMC Plant Biol. 2011, 11. [Google Scholar] [CrossRef] [PubMed]
- Grace, S.C.; Logan, B.A. Energy dissipation and radical scavenging by the plant phenylpropanoid pathway. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2000, 355, 1499–1510. [Google Scholar] [CrossRef] [PubMed]
- Landbo, A.K.; Meyer, A.S. Ascorbic acid improves the antioxidant activity of European grape juices by improving the juices’ ability to inhibit lipid peroxidation of human LDL in vitro. Int. J. Food Sci. Technol. 2001, 36, 727–735. [Google Scholar] [CrossRef]
- Jordao, A.M.; Ricardo-da-Silva, J.M.; Laureano, O. Evolution of catechins and oligomeric procyanidins during grape maturation of Castelao Francês and Touriga Francesa. Am. J. Enol. Viticult. 2001, 52, 230–234. [Google Scholar]
- Ryan, D.; Antolovich, M.; Prenzler, P.; Robards, K.; Lavee, S. Biotransformations of phenolic compounds in Olea europaea L. Sci. Hortic. 2002, 92, 147–176. [Google Scholar] [CrossRef]
- Pastore, C.; Zenoni, S.; Fasoli, M.; Pezzotti, M.; Tornielli, G.B.; Filippetti, I. Selective defoliation affects plant growth, fruit transcriptional ripening program and flavonoid metabolism in grapevine. BMC Plant Biol. 2013, 13. [Google Scholar] [CrossRef] [PubMed]
- Bowen, P.; Bogdanoff, C.; Estergaard, B.; Marsh, S. Response of Merlot Grapevines to Drip and Sprinkler Irrigation in the Okanagan Valley; British Columbia Wine Grape Council Report; British Columbia Wine Grape Council: Peachland, BC, Canada, 2005; p. 6. [Google Scholar]
- Tardáguila, J.; Blanco, J.A.; Poni, S.; Diago, M.P. Mechanical yield regulation in winegrapes: Comparison of early defoliation and crop thinning. Aust. J. Grape Wine Res. 2012, 18, 344–352. [Google Scholar] [CrossRef]
- Girona, J.; Marsal, J.; Mata, M.; del Campo, J.; Basile, B. Phenological sensitivity of berry growth and composition of Tempranillo grapevines (Vitis vinifera L.) to water stress. Aust. J. Grape Wine Res. 2009, 15, 268–277. [Google Scholar] [CrossRef]
- Dokoozlian, N.K.; Hirschfelt, D.J. The influence of clusterthinning at various stages of fruit development on flames seedless table grapes. Am. J. Enol. Viticult. 1995, 46, 429–436. [Google Scholar]
- Ojeda, H.; Andary, C.; Kraeva, E.; Carbonneau, A.; Deloire, A. Influence of pre- and postveraison water deficit on synthesis and concentration of skin phenolic compounds during berry growth of Vitis vinifera cv. Shiraz. Am. J. Enol. Viticult. 2002, 53, 261–267. [Google Scholar]
- Intrigliolo, D.S.; Castel, J.R. Interactive effects of deficit irrigation and shoot and cluster thinning on grapevine cv. Tempranillo. Water relations, vine performance and Berry and wine composition. Irrig. Sci. 2011, 29, 443–454. [Google Scholar] [CrossRef]
- Yildirim, Ö.; Denli, Y.; Ara, S.; Söylemezoğlu, G. Active oxygen scavenging enzyme activities and glutathione, ascorbic acid and lipid peroxidation levels in developing Vitis vinifera L. leaves and berries. Biotechnol. Biotechnol. Equip. 2003, 17, 114–122. [Google Scholar] [CrossRef]
- Rogiers, S.Y.; Kumar, G.N.M.; Knowles, N.R. Maturation and ripening of fruit of Amelanchier alnifolia Nutt. are accompained by increasing oxidative stress. Ann. Bot. 1998, 81, 203–211. [Google Scholar] [CrossRef]
- Negri, A.S.; Prinsi, B.; Rossoni, M.; Failla, O.; Scienza, A.; Cocucci, M.; Espen, L. Proteome changes in the skin of the grape cultivar Barbera among difference stages of ripening. BMC Genom. 2008. [Google Scholar] [CrossRef] [PubMed]
- Kärkönen, A.; Warinowski, T.; Teeri, T.H.; Simola, L.K.; Fry, S.C. On the mechanism of apoplastic H2O2 production during lignin formation and elicitation in culture spruce cells—Peroxidases after elicitation. Planta 2009, 203, 553–567. [Google Scholar] [CrossRef] [PubMed]
- Picón-Toro, J.; González-Dugo, V.; Uriarte, D.; Mancha, L.A.; Testi, L. Effect of canopy size and water stress over the crop coefficient of a Tempranillo vineyard in south-western Spain. Irrig. Sci. 2012, 30, 419–432. [Google Scholar] [CrossRef]
- Williams, L.E.; Trout, T.J. Relationships among vine and soil based measures of water status in a Thompson Seedless vineyard in response to high frequency drip irrigation. Am. J. Enol. Viticult. 2005, 56, 357–366. [Google Scholar]
- Williams, L.E.; Baeza, P. Relationships among Ambient Temperature and Vapor Pressure Deficit and Leaf and Stem Water Potentials of Fully Irrigated, Field-Grown Grapevines. Am. J. Enol. Viticult. 2007, 58, 171–181. [Google Scholar]
- International Organization of Vine and Wine. Compendium of International Methods of Wine and Must Analysis; International Organization of Vine and Wine: Paris, France, 2012; Volume 2. [Google Scholar]
- Singleton, V.L.; Salgues, M.; Zaya, J.; Troudsale, E. Caftaric acid disappearance and conversion to products of enzymatic oxidation in grape must and wine. Am. J. Enol. Viticult. 1985, 36, 50–56. [Google Scholar]
- Kim, D.; Jeong, S.W.; Leo, C.Y. Antioxidant capacity of phenolic phytochemicals from various cultivars of plums. Food Chem. 2003, 81, 321–326. [Google Scholar] [CrossRef]
- Gálvez, M.; Martín-Cordero, C.; Houghton, P.J.; Ayuso, M.J. Antioxidant activity of methanol extracts obtained from Plantago species. J. Agric. Food Chem. 2008, 53, 1927–1933. [Google Scholar] [CrossRef] [PubMed]
- Madhava Rao, K.V.; Sresty, T.V.S. Antioxidative parameters in the seedling of pigempea (Cajanus cajan L. Milspaugh) in response to Zn and Ni stresses. Plant Sci. 2000, 157, 113–128. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–259. [Google Scholar] [CrossRef]
- Ngo, T.T.; Lenhoff, H.M. A sensitive and versatile chromogenic assay for peroxidase and peroxidase-coupled reactions. Anal. Biochem. 1980, 195, 389–397. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Thipyapong, P.; Hunt, M.D.; Steffens, J.C. Systemic wound induction of potato (Solanum tuberosum) polyphenol oxidase. Phytochemistry 1995, 40, 673–676. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Treatment | Ripening Stage | Total Phenols (µg Caffeic Acid g−1 FW) | Phenylpropanoid Glycosides (µg Verbascoside g−1 FW) | Total Flavonoids (µg Rutin g−1 FW) |
|---|---|---|---|---|
| Rainfed unthinned (H1) | S1 | 919.2 ± 177.0 ab | 2075.1 ± 212.6 b | 1133.9 ± 206.3 b |
| S2 | 757.7 ± 64.1 a | 1356.6 ± 147.0 a | 788.6 ± 80.0 a | |
| S3 | 838.0 ± 136.0 ab | 1450.8 ± 175.5 a | 1139.3 ± 208.1 b | |
| S4 | 997.6 ± 161.0 b | 1735.7 ± 141.3 ab | 1519.0 ± 286.5 b | |
| Irrigated unthinned (H2) | S1 | 810.7 ± 189.0 b | 1548.8 ± 290.0 | 803.2 ± 256.3 b |
| S2 | 556.4 ± 49.4 a | 1512.6 ± 201.0 | 577.0 ± 38.1 a | |
| S3 | 928.6 ± 99.6 b | 1673.9 ± 253.4 | 1231.1 ± 148.6 c | |
| S4 | 1038.9 ± 191.3 b | 1842.2 ± 324.9 | 1489.6 ± 401.0 c | |
| Rainfed and thinned (L1) | S1 | 1558.4 ± 172.5 b | 2504.3 ± 218.0 c | 1688.2 ± 319.5 b |
| S2 | 1052.7 ± 128.4 a | 1965.1 ± 303.5 b | 1223.2 ± 101.0 a | |
| S3 | 1010.7 ± 174.5 a | 1870.6 ± 275.6 ab | 1531.5 ± 129.6 b | |
| S4 | 953.5 ± 59.4 a | 1541.1 ± 113.5 a | 1179.8 ± 124.5 a | |
| Irrigated and thinned (L2) | S1 | 804.2 ± 169.0 | 1624.5 ± 425.0 | 927.4 ± 216.4 a |
| S2 | 1035.2 ± 177.5 | 1803.6 ± 316.4 | 1141.6 ± 202.8 ab | |
| S3 | 939.0 ± 158.0 | 1735.0 ± 124.3 | 1168.4 ± 246.0 ab | |
| S4 | 1115.4 ± 146.0 | 1865.6 ± 6.3 | 1502.8 ± 167.6 b |
| Treatment | Ripening Stage | Lipid Peroxidation (nmol MDA g−1 FW) | PPO (UPPO mg−1 Protein) | SOD (USOD mg−1 Protein) | POX (UPOX mg−1 Protein) | CA-POX (UCA-POX mg−1 Protein) |
|---|---|---|---|---|---|---|
| Rainfed unthinned (H1) | S1 | 47.2 ± 14.9 a | 3143.2 ± 325.0 c | 62.7 ± 5.6 | 9.4 ± 3.1 b | 53.9 ± 15.0 |
| S2 | 79.5 ± 13.5 b | 253.7 ± 37.3 a | 76.9 ± 9.8 | 3.7 ± 0.9 a | 28.6 ± 8.7 | |
| S3 | 86.6 ± 21.9 b | 1314.5 ± 682.0 b | 86.5 ± 20.1 | 4.6 ± 1.0 a | 50.3 ± 17.3 | |
| S4 | 75.6 ± 9.1 b | 921.5 ± 42.0 b | 104.9 ± 38.5 | 3.4 ± 1.0 a | 62.9 ± 9.7 | |
| Irrigated unthinned (H2) | S1 | 46.2 ± 14.2 a | 1862.8 ± 402.3 c | 103.6 ± 25.9 a | 4.9 ± 1.5 b | 69.9 ± 18.3 |
| S2 | 80.1 ± 16.0 b | 315.2 ± 136.0 a | 141.9 ± 17.2 b | 6.6 ± 1.9 b | 67.8 ± 15.1 | |
| S3 | 71.3 ± 15.8 ab | 1363.3 ± 184.0 c | 142.3 ± 27.9 b | 4.4 ± 0.7 b | 70.4 ± 12.4 | |
| S4 | 93.7 ± 2.9 b | 656.9 ± 166.0 b | 103.6 ± 4.2 a | 2.0 ± 0.1 a | 45.7 ± 15.6 | |
| Rainfed thinned (L1) | S1 | 71.0 ± 34.0 a | 2461.8 ± 497.0 b | 49.5 ± 7.6 | 5.1 ± 0.6 b | 61.4 ± 19.0 |
| S2 | 130.3 ± 19.1 b | 2704.0 ± 358.0 b | 62.3 ± 19.0 | 2.1 ± 0.3 a | 72.5 ± 18.9 | |
| S3 | 121.4 ± 26.2 ab | 805.4 ± 224.2 a | 65.0 ± 15.7 | 3.8 ± 1.2 ab | 61.7 ± 14.7 | |
| S4 | 98.8 ± 9.2 a | 1471.5 ± 697.3 ab | 47.8 ± 1.1 | 3.0 ± 0.6 a | 51.8 ± 19.8 | |
| Irrigated thinned (L2) | S1 | 39.9 ± 21.5 a | 2322.4 ± 328.5 b | 94.9 ± 5.0 | 5.6 ± 1.3 b | 71.9 ± 28.6 |
| S2 | 72.4 ± 20.0 ab | 2992.5 ± 130.0 c | 76.1 ± 25.4 | 2.3 ± 1.1 a | 44.1 ± 6.5 | |
| S3 | 78.6 ± 20.4 ab | 1134.3 ± 282.4 a | 80.7 ± 18.2 | 2.7 ± 0.9 a | 72.4 ± 26.1 | |
| S4 | 97.9 ± 19.6 b | 1273.5 ± 329.0 a | 73.8 ± 15.5 | 1.7 ± 0.5 a | 52.7 ± 8.0 |
| Month | Rainfall (mm) | Tmax (°C) | Tmin (°C) | T (°C) | % HR max | % HR min | % HR | Solar Radiation (MJ·m−2·day−1) | Net Solar Radiation (MJ·m−2·day−1) |
|---|---|---|---|---|---|---|---|---|---|
| January | 14.80 | 14.31 | −0.47 | 5.88 | 83.96 | 52.68 | 81.47 | 9.34 | 2.43 |
| February | 2.80 | 15.63 | −3.06 | 5.93 | 85.32 | 24.79 | 55.95 | 14.81 | 4.58 |
| March | 6.40 | 20.99 | 4.43 | 12.75 | 79.75 | 24.27 | 51.11 | 17.74 | 7.70 |
| April | 50.60 | 18.41 | 7.46 | 12.71 | 87.61 | 40.57 | 66.81 | 17.60 | 9.68 |
| May | 37.80 | 27.50 | 12.26 | 19.91 | 86.28 | 27.70 | 57.60 | 24.35 | 13.45 |
| June | 0.00 | 30.54 | 14.68 | 22.67 | 82.79 | 22.90 | 52.43 | 28.12 | 15.52 |
| July | 0.00 | 32.78 | 14.96 | 23.93 | 79.75 | 19.39 | 48.70 | 29.51 | 15.72 |
| August | 0.20 | 32.56 | 15.00 | 24.20 | 83.40 | 22.25 | 51.27 | 25.38 | 13.19 |
| September | 47.60 | 29.24 | 13.75 | 21.49 | 83.87 | 29.48 | 56.80 | 18.25 | 8.89 |
| October | 62.20 | 23.03 | 10.58 | 16.48 | 91.49 | 42.64 | 72.56 | 13.05 | 5.52 |
| November | 98.00 | 17.03 | 7.68 | 12.08 | 94.23 | 59.38 | 81.37 | 7.99 | 2.86 |
| December | 58.00 | 13.70 | 4.56 | 8.70 | 98.50 | 72.30 | 90.88 | 6.12 | 1.98 |
| Annual | 378.40 | 22.98 | 8.49 | 15.56 | 87.25 | 36.53 | 63.92 | 17.69 | 8.46 |
| Growth cycle | 136.20 | 28.51 | 13.02 | 20.82 | 83.95 | 27.05 | 55.60 | 23.87 | 12.74 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garrido, I.; Uriarte, D.; Hernández, M.; Llerena, J.L.; Valdés, M.E.; Espinosa, F. The Evolution of Total Phenolic Compounds and Antioxidant Activities during Ripening of Grapes (Vitis vinifera L., cv. Tempranillo) Grown in Semiarid Region: Effects of Cluster Thinning and Water Deficit. Int. J. Mol. Sci. 2016, 17, 1923. https://doi.org/10.3390/ijms17111923
Garrido I, Uriarte D, Hernández M, Llerena JL, Valdés ME, Espinosa F. The Evolution of Total Phenolic Compounds and Antioxidant Activities during Ripening of Grapes (Vitis vinifera L., cv. Tempranillo) Grown in Semiarid Region: Effects of Cluster Thinning and Water Deficit. International Journal of Molecular Sciences. 2016; 17(11):1923. https://doi.org/10.3390/ijms17111923
Chicago/Turabian StyleGarrido, Inmaculada, David Uriarte, Marcos Hernández, José Luis Llerena, María Esperanza Valdés, and Francisco Espinosa. 2016. "The Evolution of Total Phenolic Compounds and Antioxidant Activities during Ripening of Grapes (Vitis vinifera L., cv. Tempranillo) Grown in Semiarid Region: Effects of Cluster Thinning and Water Deficit" International Journal of Molecular Sciences 17, no. 11: 1923. https://doi.org/10.3390/ijms17111923
APA StyleGarrido, I., Uriarte, D., Hernández, M., Llerena, J. L., Valdés, M. E., & Espinosa, F. (2016). The Evolution of Total Phenolic Compounds and Antioxidant Activities during Ripening of Grapes (Vitis vinifera L., cv. Tempranillo) Grown in Semiarid Region: Effects of Cluster Thinning and Water Deficit. International Journal of Molecular Sciences, 17(11), 1923. https://doi.org/10.3390/ijms17111923