
Anti-Melanogenic Activities of Heracleum moellendorffii via ERK1/2-Mediated MITF Downregulation



Abstract
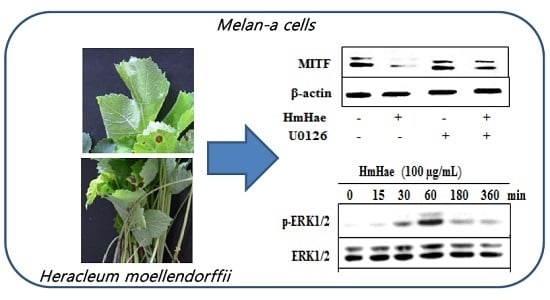
:
1. Introduction
2. Results
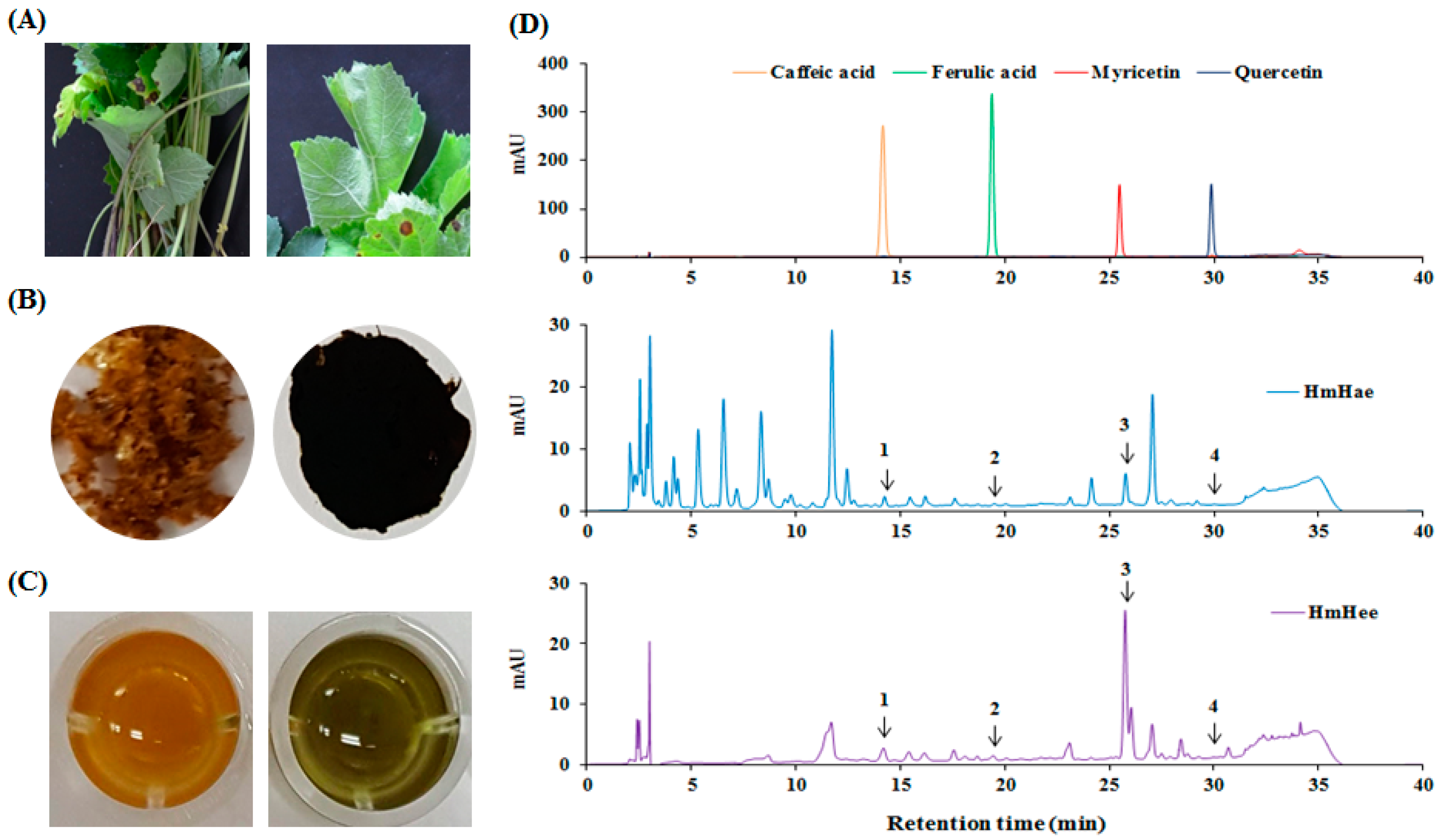
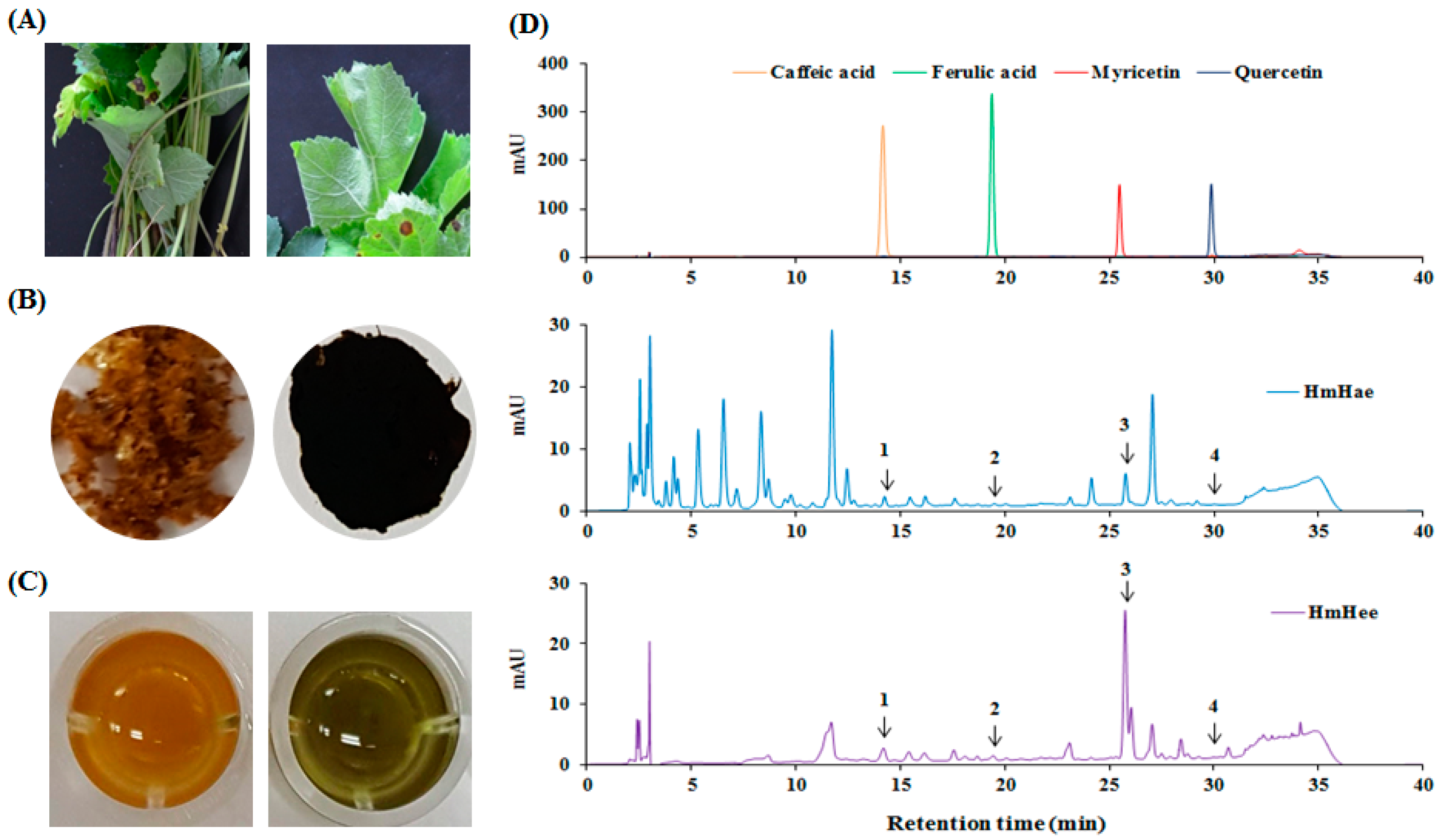
2.1. Chemical Compounds of HmHe by HPLC
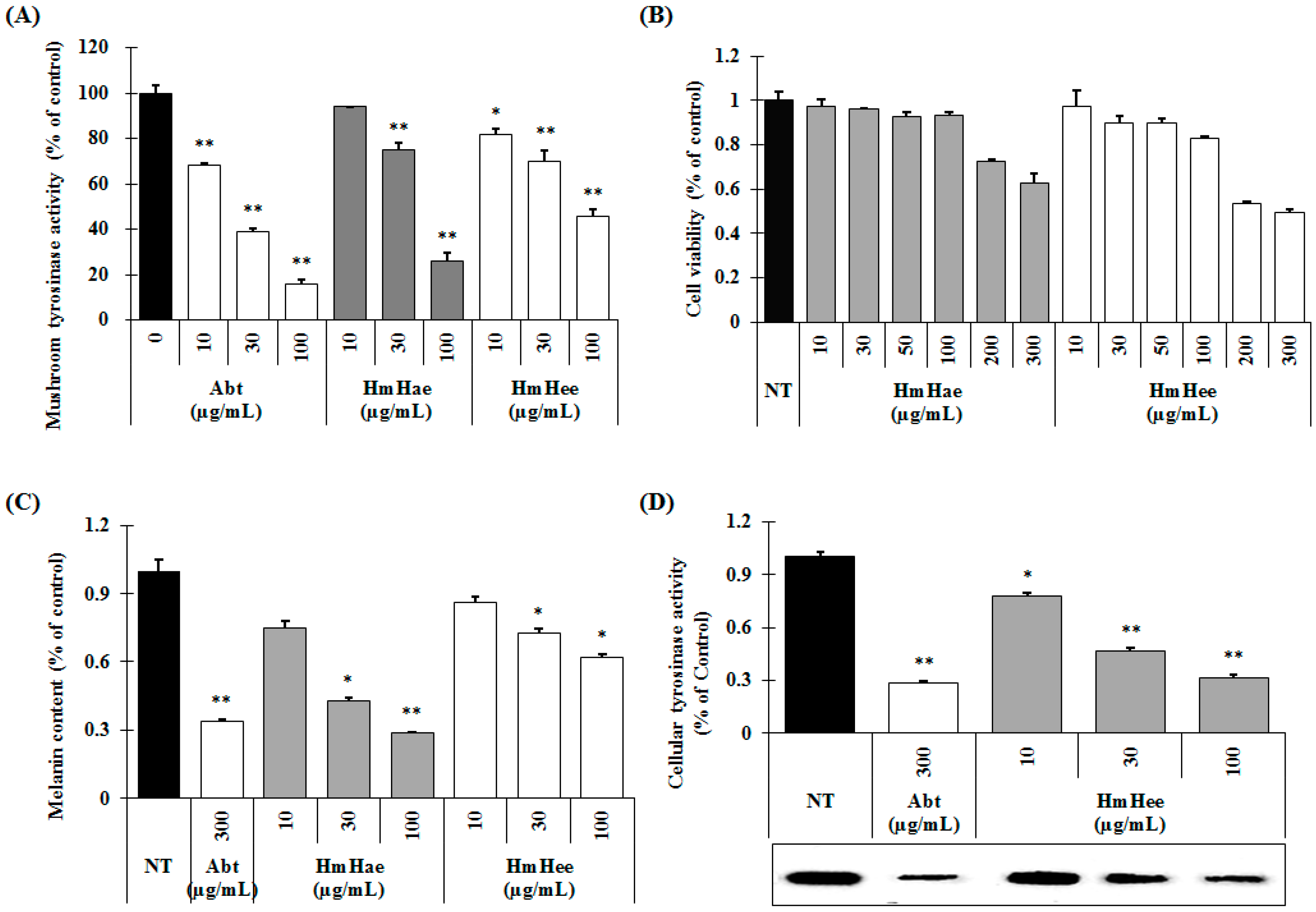
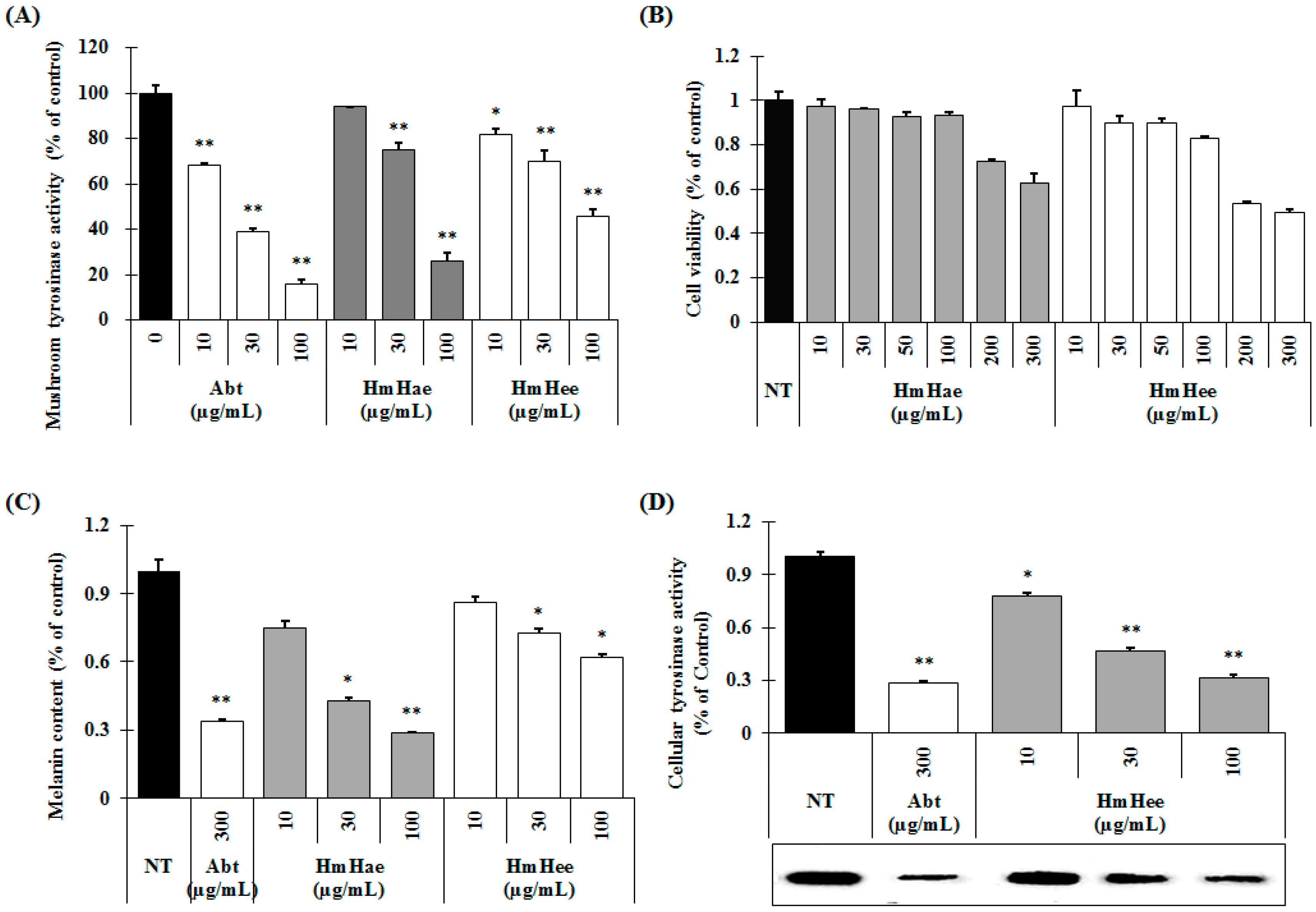
2.2. Effects of HmHe on Mushroom Tyrosinase Activity, Melanin Production and Intracellular Tyrosinase Activity
2.3. Effects of HmHe on Expression of Melanogenesis-Related Proteins
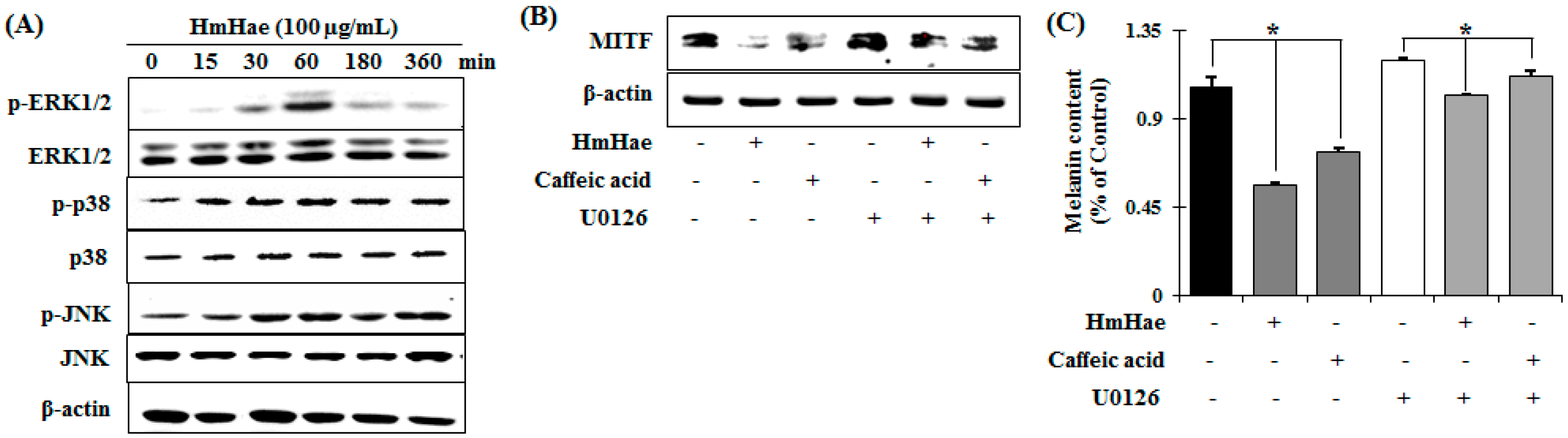
2.4. Effects of HmHe on ERK1/2 Signaling Pathways
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Extraction
4.2. Drugs and Chemicals
4.3. Chemical Compound Analysis by HPLC-DAD (High-Performance Liquid Chromatography with Diode-Array Detection)
4.4. Cell Culture and Cell Viability Assay
4.5. Measurement of Mushroom Tyrosinase Activity
4.6. Melanogenesis Inhibitory Assay in Melan-a Cells
4.7. Analysis of Intracellular Tyrosinase Activity by Zymography
4.8. Analysis of mRNA Expressions
4.9. Preparation of Cell Lysates and Western Blotting
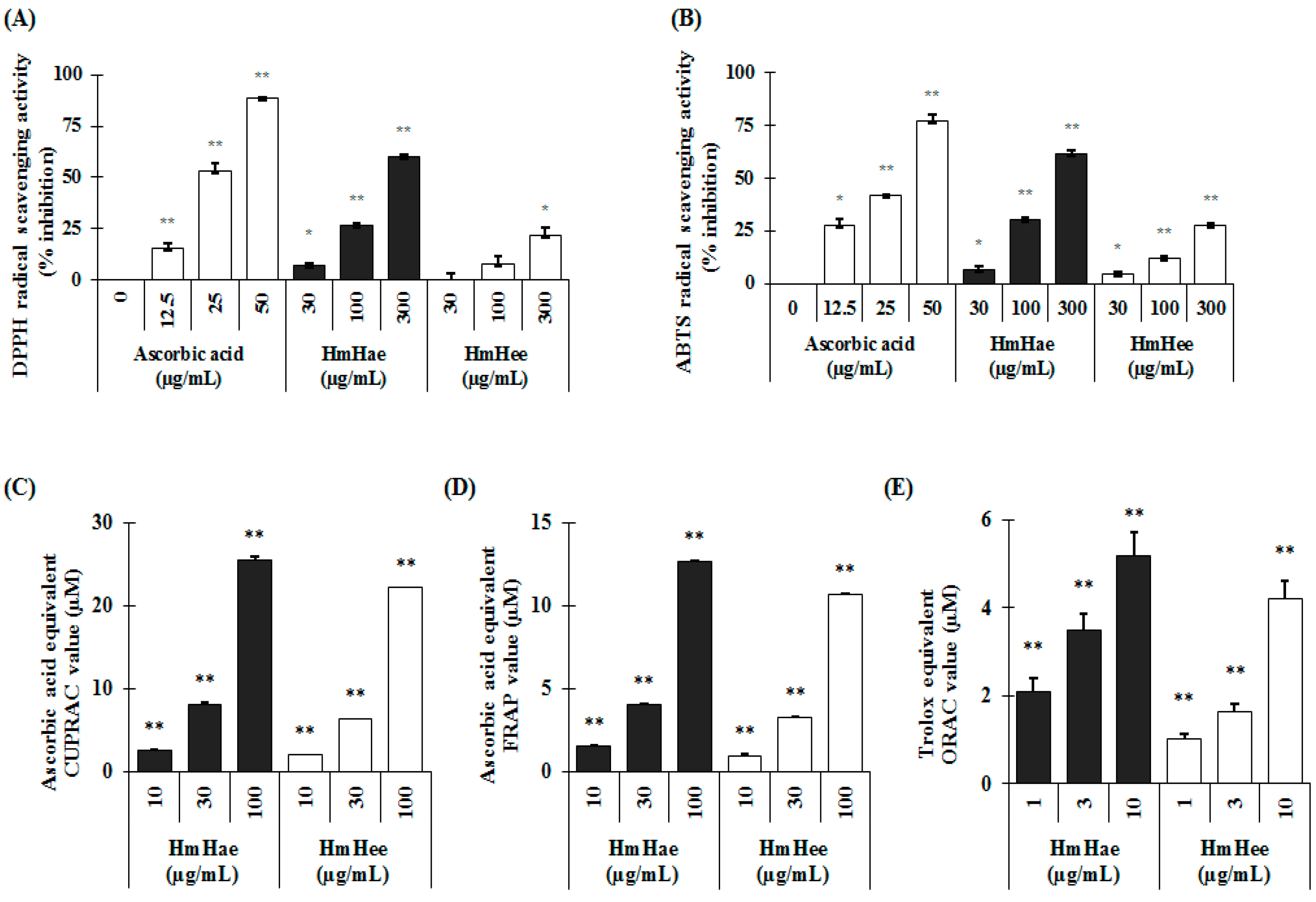
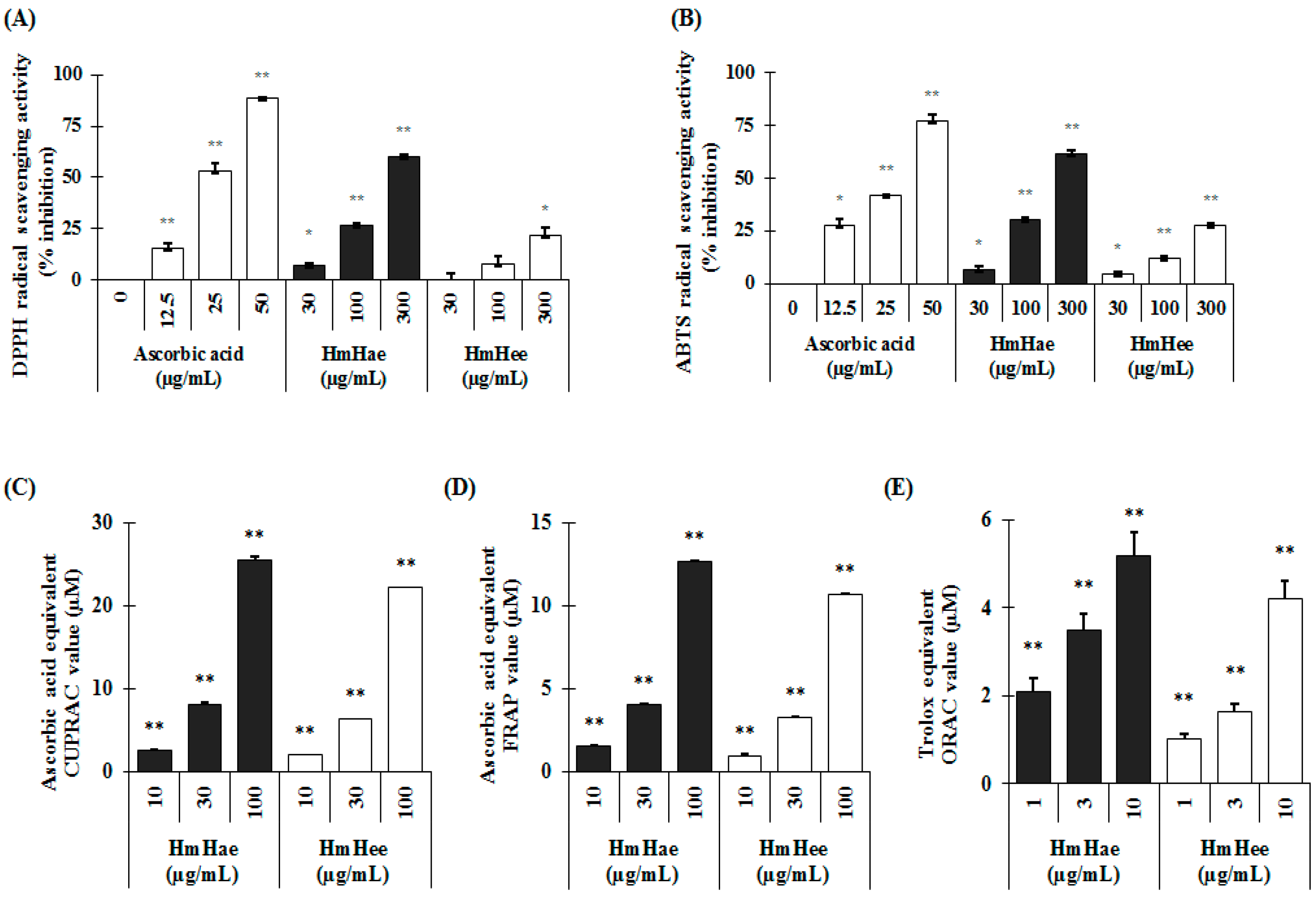
4.10. In Vitro Antioxidant Assays
4.11. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Miyamura, Y.I.; Coelho, S.G.; Wolber, R.; Miller, S.A.; Wakamatsu, K.; Zmudzka, B.Z.; Ito, S.; Smuda, C.; Passeron, T.; Choi, W.; et al. Regulation of human skin pigmentation and responses to ultraviolet radiation. Pigment Cell Res. 2007, 20, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Jimbow, K.; Park, J.S.; Kato, F.; Hirosaki, K.; Toyofuku, K.; Hua, C.; Yamashita, T. Assembly, target-signaling and intracellular transport of tyrosinase gene family proteins in the initial stage of melanosome biogenesis. Pigment Cell Res. 2000, 13, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.Y.; Fisher, D.E. Melanocyte biology and skin pigmentation. Nature 2007, 445, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Wakamatsu, K. Chemistry of mixed melanogenesis—Pivotal roles of dopaquinone. Photochem. Photobiol. 2008, 84, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Cao, J.; Wang, H.; Zhang, J.; Zhu, Z.; Bai, R.; Hao, H.; He, X.; Fan, R.; Dong, C. Nitric oxide enhances the sensitivity of alpaca melanocytes to respond to α-melanocyte-stimulating hormone by up-regulating melanocortin-1 receptor. Biochem. Biophys. Res. Commun. 2010, 396, 849–853. [Google Scholar] [CrossRef] [PubMed]
- Horikoshi, T.; Nakahara, M.; Kaminaga, H.; Sasaki, M.; Uchiwa, H.; Miyachi, Y. Involvement of nitric oxide in UVB-induced pigmentation in guinea pig skin. Pigment Cell Res. 2000, 13, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Weller, R. Nitric oxide: A key mediator in cutaneous physiology. Clin. Exp. Dermatol. 2003, 28, 511–514. [Google Scholar] [CrossRef] [PubMed]
- García-Gavín, J.; González-Vilas, D.; Fernández-Redondo, V.; Toribio, J. Pigmented contact dermatitis due to kojic acid. A paradoxical side effect of a skin lightener. Contact Dermat. 2010, 62, 63–64. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.L.; Liu, R.H.; Sheu, J.N.; Chen, S.T.; Sinchaikul, S.; Tsay, G.J. Toxicogenomics of A375 human malignant melanoma cells treated with arbutin. J. Biomed. Sci. 2007, 14, 87–105. [Google Scholar] [CrossRef] [PubMed]
- Bang, J.E.; Choi, H.Y.; Kim, S.I. Anti-oxidative activity and chemical composition of various Heracleum moellendorffii Hance extracts. Korean J. Food Preserv. 2009, 16, 765–771. [Google Scholar]
- Park, H.J.; Nugroho, A.; Jung, B.; Won, Y.H.; Jung, Y.J.; Kim, W.B.; Choi, J.S. Isolation and quantitative analysis of flavonoids with peroxynitrite-scavenging effect from the young leaves of Heracleum moellendorffii. Korean J. Plant Res. 2010, 23, 393–398. [Google Scholar]
- Nakano, Y.; Matsunaga, H.; Saita, T.; Mori, M.; Katano, M.; Okabe, H. Antiproliferative constituents in Umbelliferae plants II. Screening for polyacetylenes in some Umbelliferae plants, and isolation of panaxynol and falcarindiol from the root of Heracleum moellendorffii. Biol. Pharm. Bull. 1998, 21, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.L.; Goh, S.H.; Ho, S.H. Screening of Chinese medicinal herbs for bioactivity against Sitophilus zeamais Motschulsky and Tribolium castaneum (Herbst). J. Stored Prod. Res. 2007, 43, 290–296. [Google Scholar] [CrossRef]
- Chu, S.S.; Cao, J.; Liu, Q.Z.; Du, S.S.; Deng, Z.W.; Liu, Z.L. Chemical composition and insecticidal activity of Heracleum moellendorffii Hance essential oil. Chemija 2012, 23, 108–112. [Google Scholar]
- Wu, L.C.; Chang, L.H.; Chen, S.H.; Fan, N.C.; Ho, J.A. Antioxidant activity and melanogenesis inhibitory effect of the acetonic extract of Osmanthus fragrans: A potential natural and functional food flavor additive. Food Sci. Technol. 2009, 42, 1513–1519. [Google Scholar] [CrossRef]
- Dewanto, V.; Wu, X.Z.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Lee, W.J.; Chang, S.E.; Lee, G.Y. Hesperidin, a popular antioxidant inhibits melanogenesis via ERK1/2 mediated MITF degradation. Int. J. Mol. Sci. 2015, 16, 18384–18395. [Google Scholar] [CrossRef] [PubMed]
- Hearing, V.J. Determination of melanin synthetic pathways. J. Investig. Dermatol. 2011, 17, E8–E11. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.; Hughes, J.; Hong, M.; Jia, Q.; Orndorff, S. Modulation of melanogenesis by aloesin: A competitive inhibitor of tyrosinase. Pigment Cell Res. 2002, 15, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.S.; Wei, C.I.; Marshall, M.R. Inhibition mechanism of Kojic acid on polyphenol oxidase. J. Agric. Food Chem. 1991, 39, 1897–1901. [Google Scholar] [CrossRef]
- Mastsuda, M.; Murata, K.; Fukuhama, A.; Naruto, S.; Fujita, T.; Uwaya, A.; Isami, F.; Matsuda, H. Inhibitory effects of constituents of Morinda citrifolia seeds on elastase and tyrosinase. J. Nat. Med. 2009, 63, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Kim, D.S.; Lee, H.K.; Kwon, S.B.; Lee, S.; Ryoo, I.J.; Kim, W.G.; Yoo, I.D.; Park, K.C. Long-term suppression of tyrosinase by terrein via tyrosinase degradation and its decreased expression. Exp. Dermatol. 2009, 18, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.B.; Babiarz, L.; Liebel, F.; Roydon-Price, E.; Kizoulis, M.; Gendimenico, G.J.; Fisher, D.E.; Seiberg, M. Modulation of microphthalmia-associated transcription factor gene expression alters skin pigmentation. J. Investig. Dermatol. 2002, 119, 1330–1340. [Google Scholar] [CrossRef] [PubMed]
- Englaro, W.; Bertolotto, C.; Busca, R.; Brunet, A.; Pages, G.; Ortonne, J.P.; Ballotti, R. Inhibition of the mitogen-activated protein kinase pathway triggers B16 melanoma cell differentiation. J. Biol. Chem. 1998, 273, 9966–9970. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Hwang, E.S.; Lee, J.E.; Kim, S.Y.; Kwon, S.B.; Park, K.C. Sphingosine-1-phosphate decreases melanin synthesis via sustained ERK activation and subsequent MITF degradation. J. Cell Sci. 2003, 116, 1699–1706. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Kang, K.S.; Yokozawa, T. The anti-melanogenic effect of pycnogenol by its anti-oxidative actions. Food Chem. Toxicol. 2008, 46, 2466–2471. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.L.; Wang, B.S.; Chang, L.W.; Wu, H.C.; Huang, S.L.; Huang, M.H. Antioxidant and antityrosinase activity of aqueous extracts of green asparagus. Food Chem. 2011, 127, 141–146. [Google Scholar]
- Hanamura, T.; Uchida, E.; Aoki, H. Skin-lightening effect of a polyphenol extract from acerola (Malpighia emarginata DC.) fruit on UV-induced pigmentation. Biosci. Biotechnol. Biochem. 2008, 72, 3211–3218. [Google Scholar] [CrossRef] [PubMed]
- Michihara, A.; Morita, S.; Hirokawa, Y.; Ago, S.; Akasaki, K.; Tsuji, H. Deltatocotrienol causes decrease of melanin content in mouse melanoma cells. J. Health Sci. 2009, 55, 314–318. [Google Scholar] [CrossRef]
- Ohgidani, M.; Komizu, Y.; Goto, K.; Ueoka, R. Antimelanogenic and antioxidative effects of residual powders from Shochu distillation remnants. Food Chem. 2012, 132, 2140–2143. [Google Scholar] [CrossRef]
- Baek, S.H.; Nam, I.J.; Kwak, H.S.; Kim, K.C.; Lee, S.H. Cellular anti-melanogenic effects of a Euryale ferox seed extract ethyl acetate fraction via the lysosomal degradation machinery. Int. J. Mol. Sci. 2015, 16, 9217–9235. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, Y.; Oka, M.; Tsunakawa, M.; Tomita, K.; Hatori, M.; Yamamoto, H.; Kamei, H.; Miyaki, T.; Konishi, M.; Oki, T. Melanostatin, a new melanin synthesis inhibitor. Production, isolation, chemical properties, structure and biological activity. J. Antibiot. 1991, 44, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Bennett, D.C.; Cooper, P.J.; Hart, I.R. A line of non-tumorigenic mouse melanocytes, syngeneic with the B16 melanoma and requiring a tumour promoter for growth. Int. J. Cancer 1987, 39, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Takahashi, H.; Iraha, R.; Toriyama, M. Down-regulation of tyrosinase expression by acetylsalicylic acid in murine B16 melanoma. Biol. Pharm. Bull. 2008, 31, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Elias, P.M.; Menon, G.; Wetzel, B.K.; Williams, J.W. Evidence that stress to the epidermal barrier influenced the development of pigmentation in humans. Pigment Cell Melanoma Res. 2009, 22, 420–434. [Google Scholar] [CrossRef] [PubMed]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol. 2000, 132, 365–386. [Google Scholar] [PubMed]
- Nanjo, F.; Goto, K.; Seto, R.; Suzuki, M.; Sakai, M.; Hara, Y. Scavenging effects of tea catechins and their derivatives on 1,1-diphenyl-2-picrylhydrazyl radical. Free Radic. Biol. Med. 1996, 21, 895–902. [Google Scholar] [CrossRef]
- Noguchi, N.; Yamashita, H.; Gotoh, N.; Yamamoto, Y.; Numano, R. 2,2′-Azobis (4-Methoxy-2,4-Dimethylvaleronitrile), a new lipid-soluble azo initiator: Application to oxidations of lipids and low-density lipoprotein in solution and in aqueous dispersions. Free Radic. Biol. Med. 1998, 24, 259–268. [Google Scholar] [CrossRef]
- Apak, R.; Guclu, K.; Ozyurek, M.; Karademir, S.E. Novel total antioxidant capacity index for dietary polyphenols and vitamins C and E, using their cupric ion reducing capability in the presence of neocuproine: CUPRAC method. J. Agric. Food Chem. 2004, 52, 7970–7981. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Huang, D.; Hampsch-Woodill, M.; Flanagan, J.A.; Deemer, E.K. Analysis of antioxidant activities of common vegetables employing oxygen radical absorbance capacity (ORAC) and ferric reducing antioxidant power (FRAP) assays: A comparative study. J. Agric. Food Chem. 2002, 50, 3122–3128. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Standards | Contents (mg/L) | ||
|---|---|---|---|---|
| 280 nm | 340 nm | Average 1 | ||
| HmHae 2 | Caffeic acid | 13.16 | 12.41 | 12.80 |
| Ferulic acid | 3.03 | 1.16 | 2.01 | |
| Myricetin | 103.95 | 96.12 | 100.04 | |
| Quercetin | 0.81 | 0.44 | 0.63 | |
| HmHee 3 | Caffeic acid | 25.98 | 21.90 | 23.9 |
| Ferulic acid | 7.55 | 3.45 | 5.50 | |
| Myricetin | 446.19 | 408.84 | 427.52 | |
| Quercetin | 1.23 | 1.36 | 1.30 | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alam, M.B.; Seo, B.-J.; Zhao, P.; Lee, S.-H. Anti-Melanogenic Activities of Heracleum moellendorffii via ERK1/2-Mediated MITF Downregulation. Int. J. Mol. Sci. 2016, 17, 1844. https://doi.org/10.3390/ijms17111844
Alam MB, Seo B-J, Zhao P, Lee S-H. Anti-Melanogenic Activities of Heracleum moellendorffii via ERK1/2-Mediated MITF Downregulation. International Journal of Molecular Sciences. 2016; 17(11):1844. https://doi.org/10.3390/ijms17111844
Chicago/Turabian StyleAlam, Md Badrul, Bum-Ju Seo, Peijun Zhao, and Sang-Han Lee. 2016. "Anti-Melanogenic Activities of Heracleum moellendorffii via ERK1/2-Mediated MITF Downregulation" International Journal of Molecular Sciences 17, no. 11: 1844. https://doi.org/10.3390/ijms17111844
APA StyleAlam, M. B., Seo, B.-J., Zhao, P., & Lee, S.-H. (2016). Anti-Melanogenic Activities of Heracleum moellendorffii via ERK1/2-Mediated MITF Downregulation. International Journal of Molecular Sciences, 17(11), 1844. https://doi.org/10.3390/ijms17111844