

The Extracts Derived from Artemisia japonica Thunb. Leaves Mitigate Oxidative Stress and Inflammatory Response Induced by LPS in RAW264.7 Cells through Modulation of the Nrf2/HO-1 Signaling Pathway

Abstract

1. Introduction
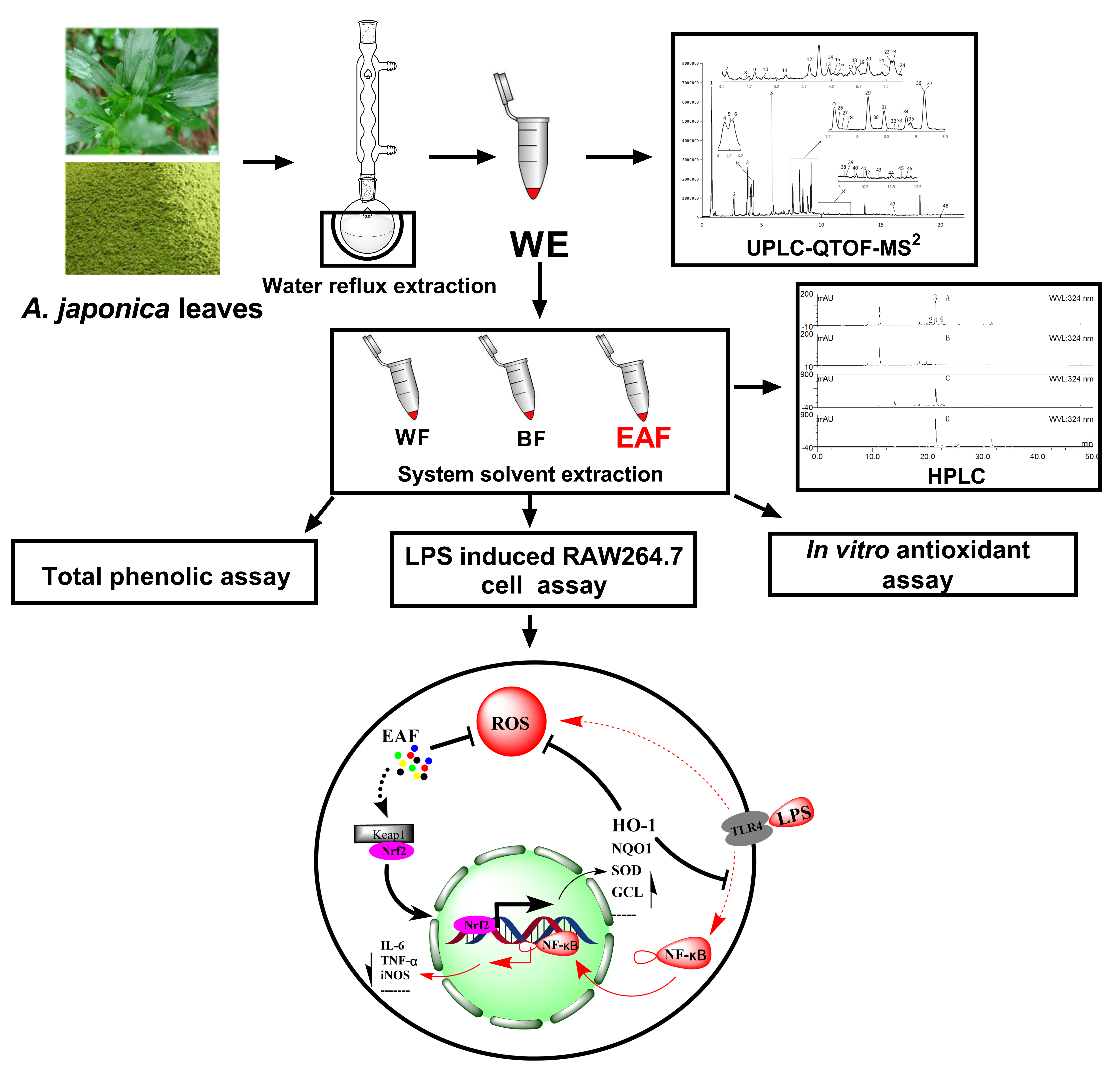
2. Results and Discussion
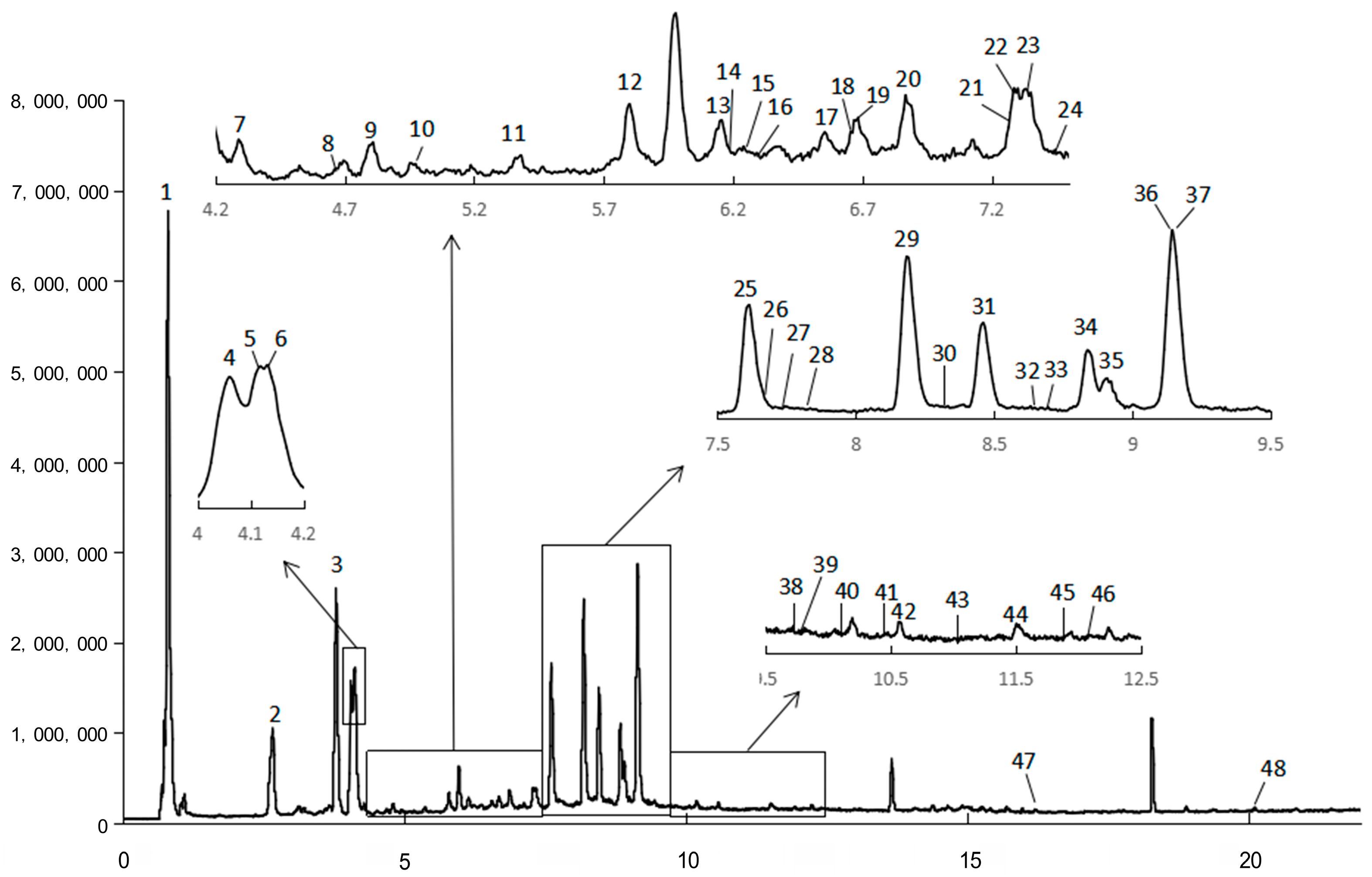
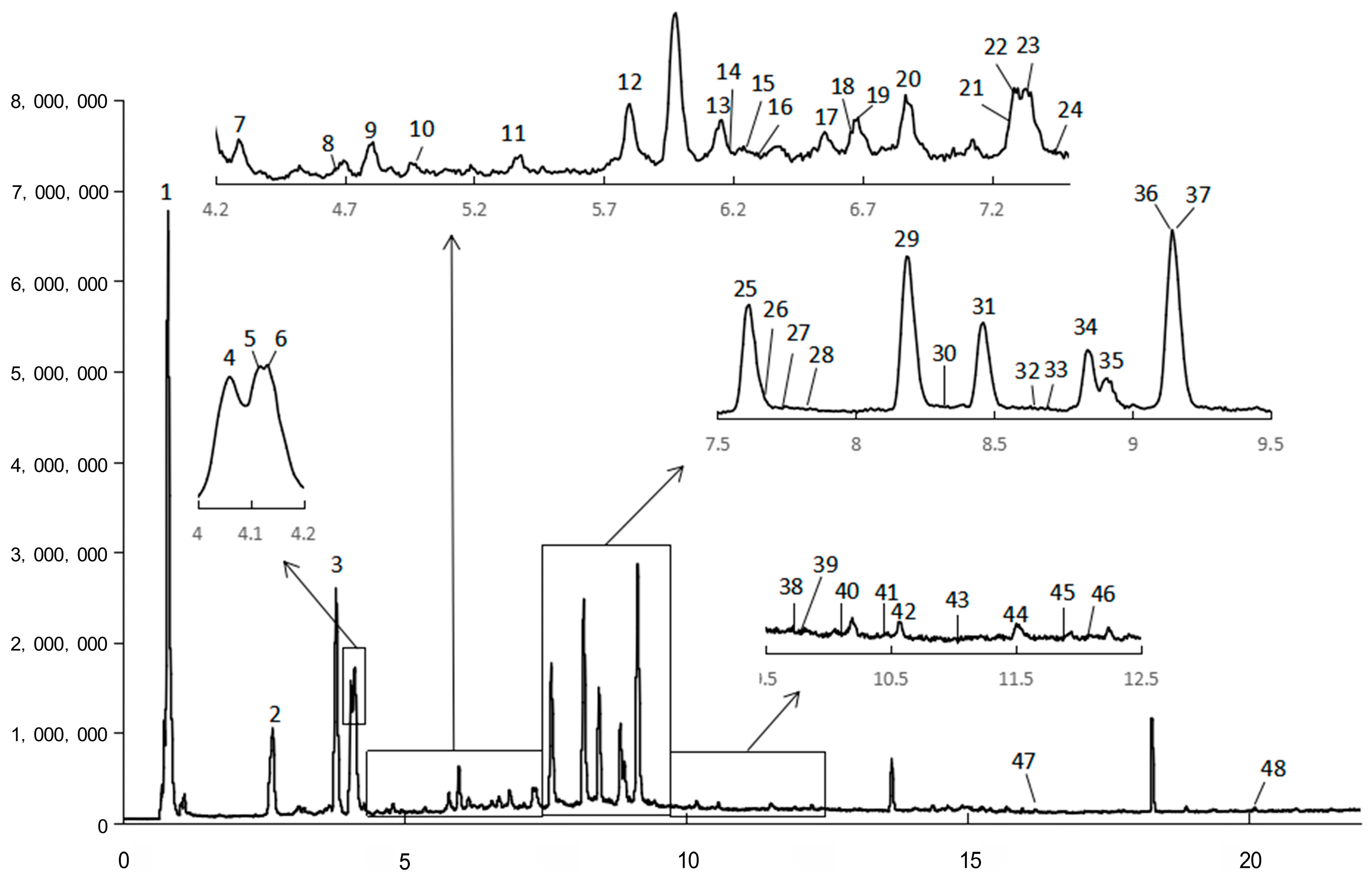
2.1. Chemical Composition
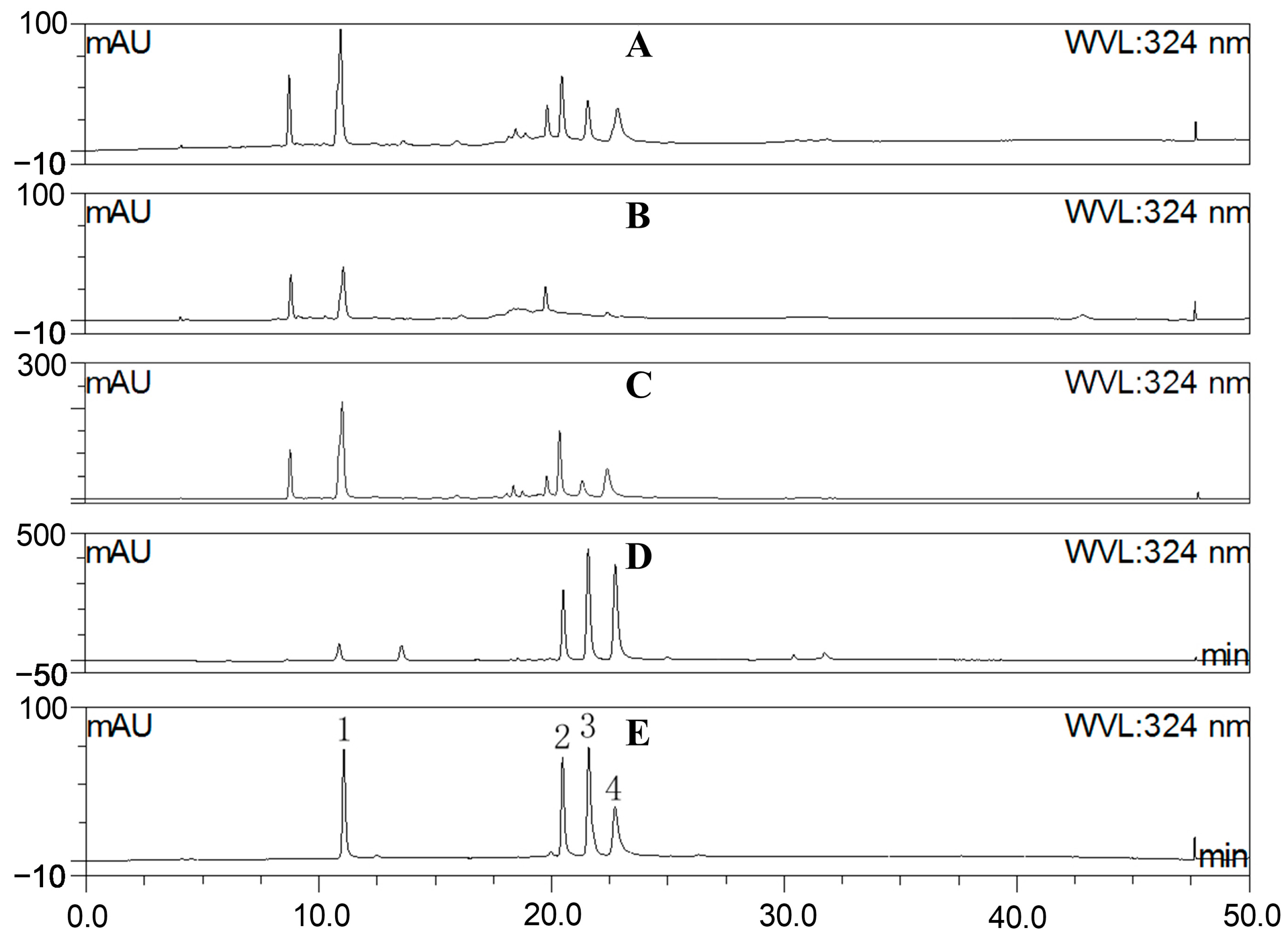
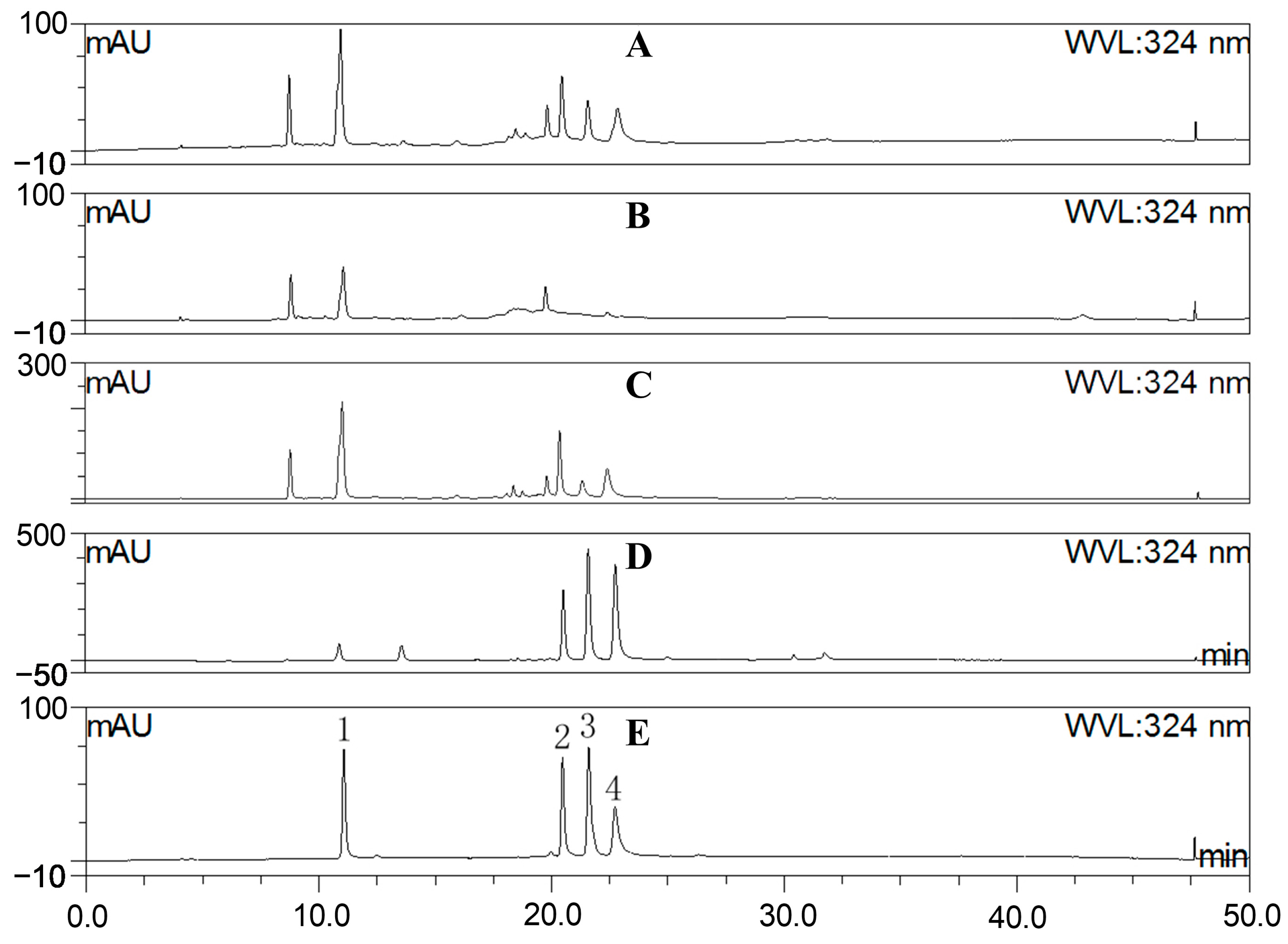
2.2. Fraction Characterization
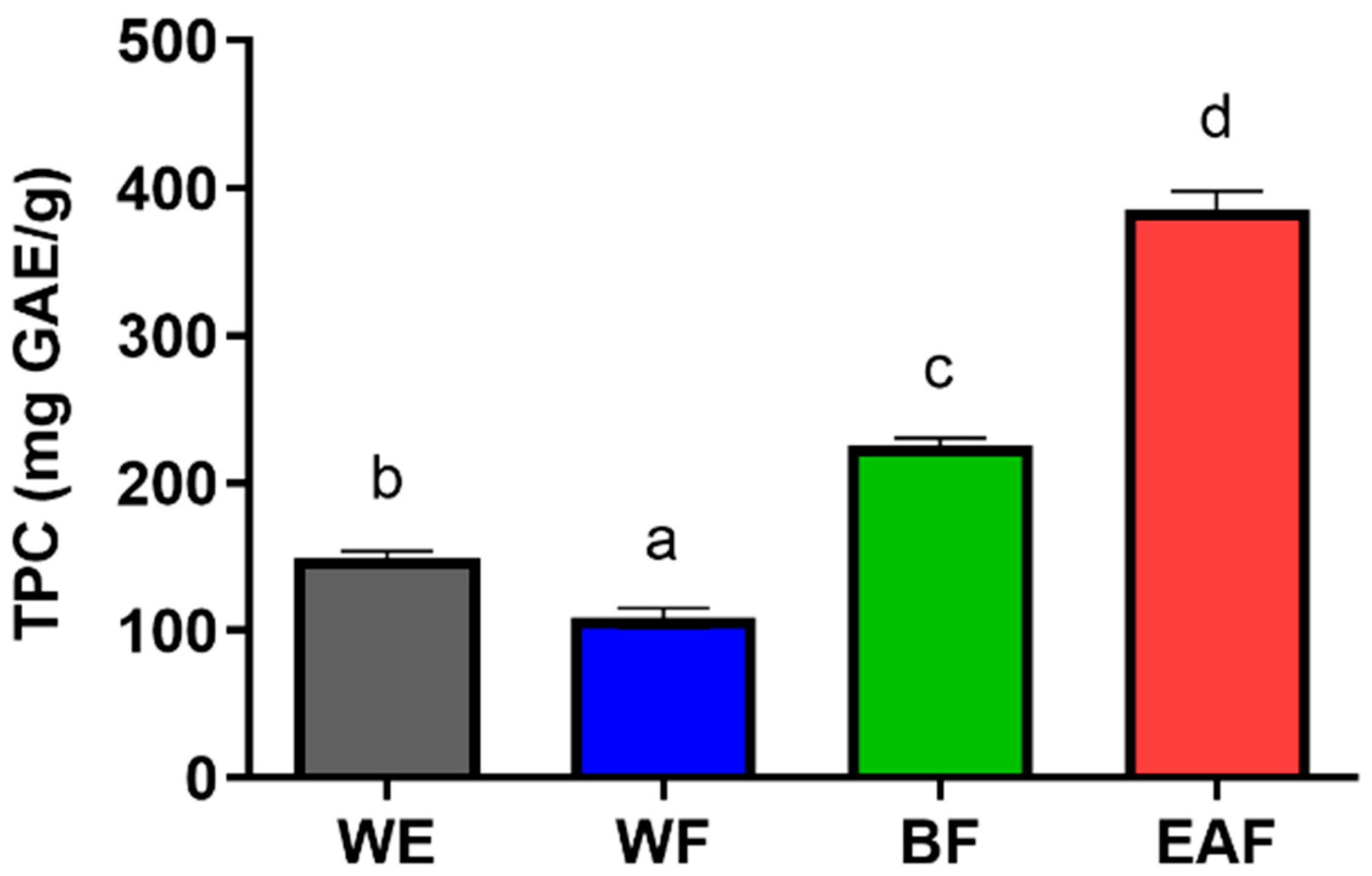
2.3. Total Phenolics
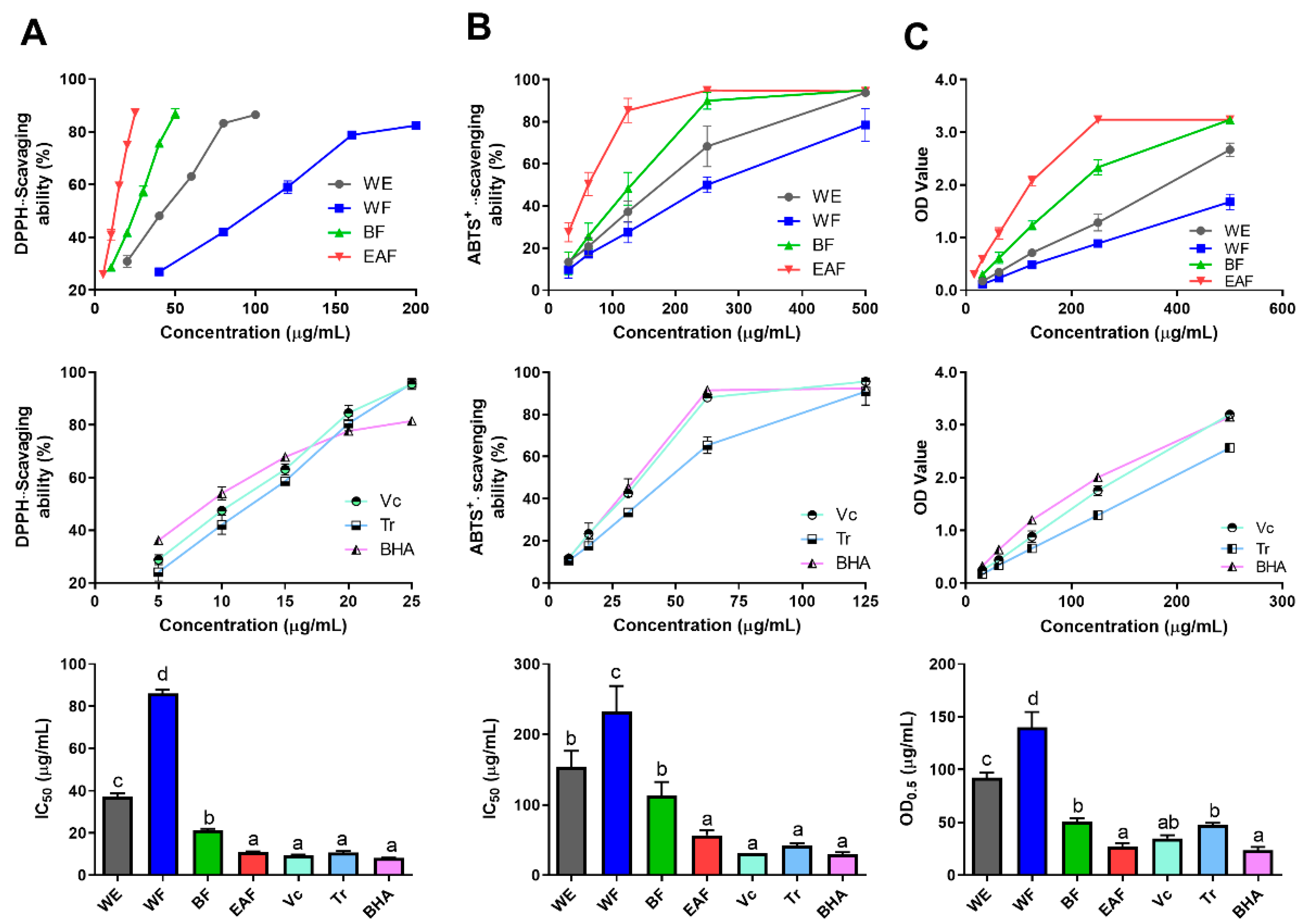
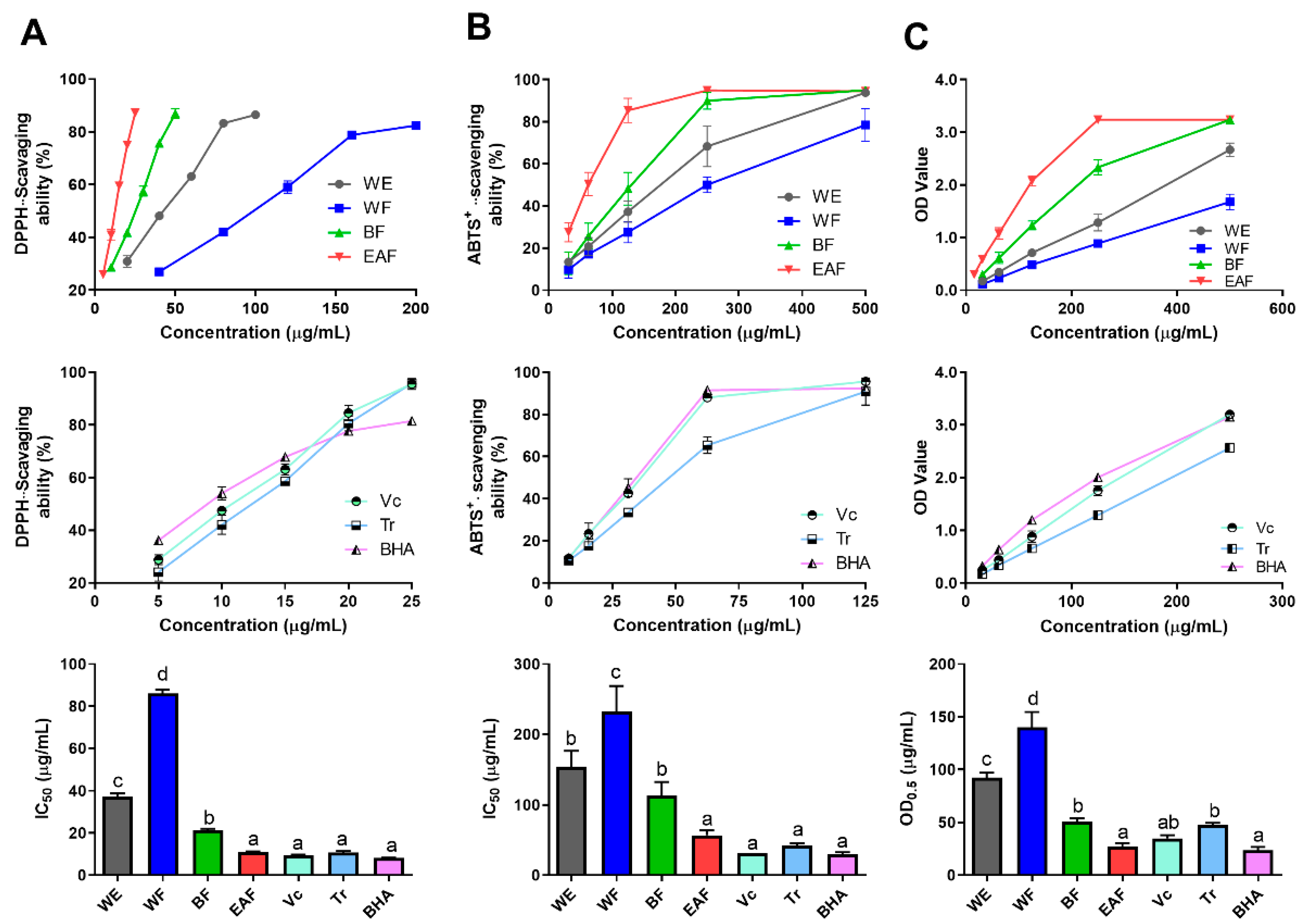
2.4. Comparison of Antioxidant Activity
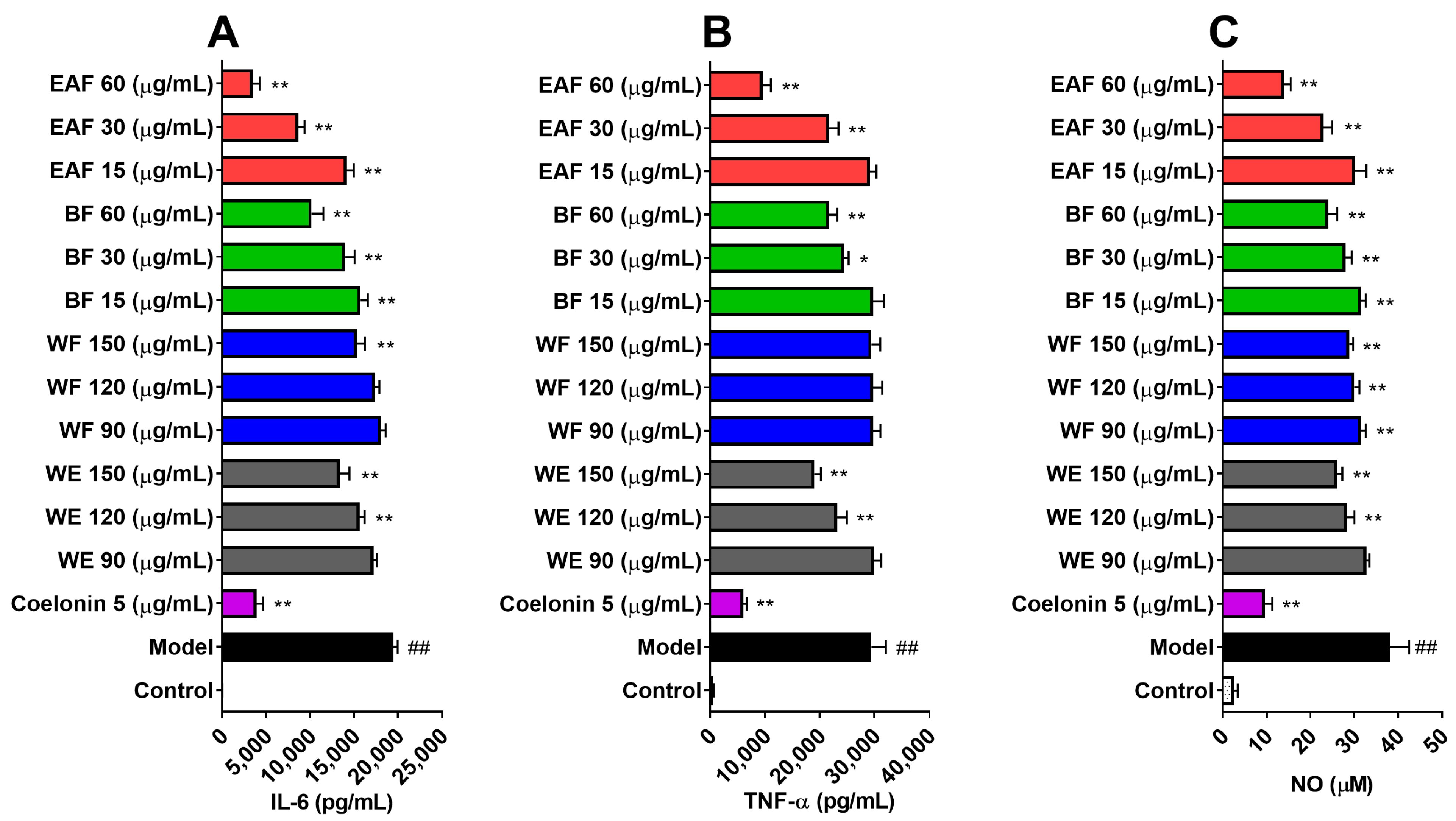
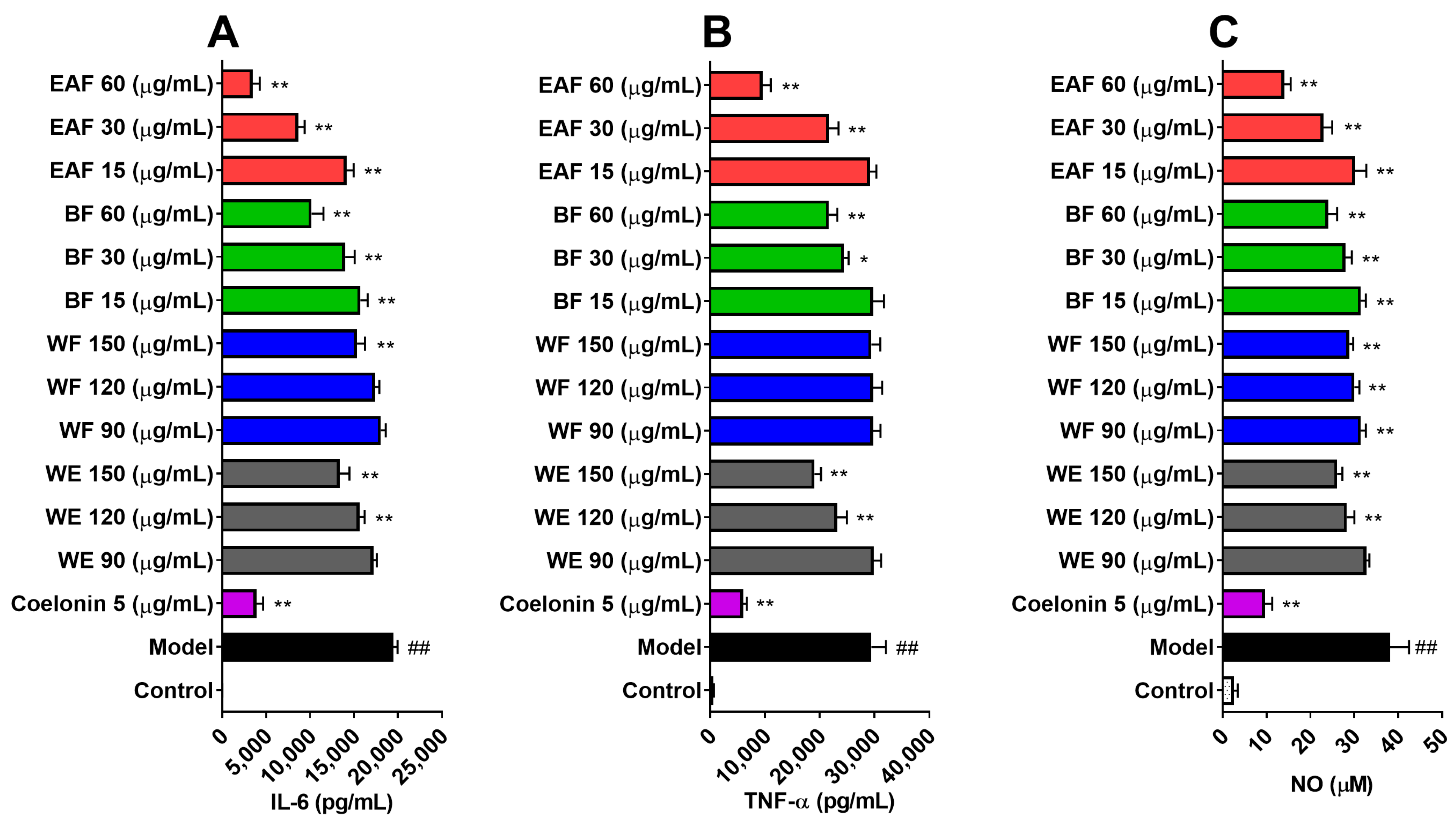
2.5. Anti-Inflammatory Effects on Lipopolysaccharide (LPS)-Induced RAW264.7 Cells
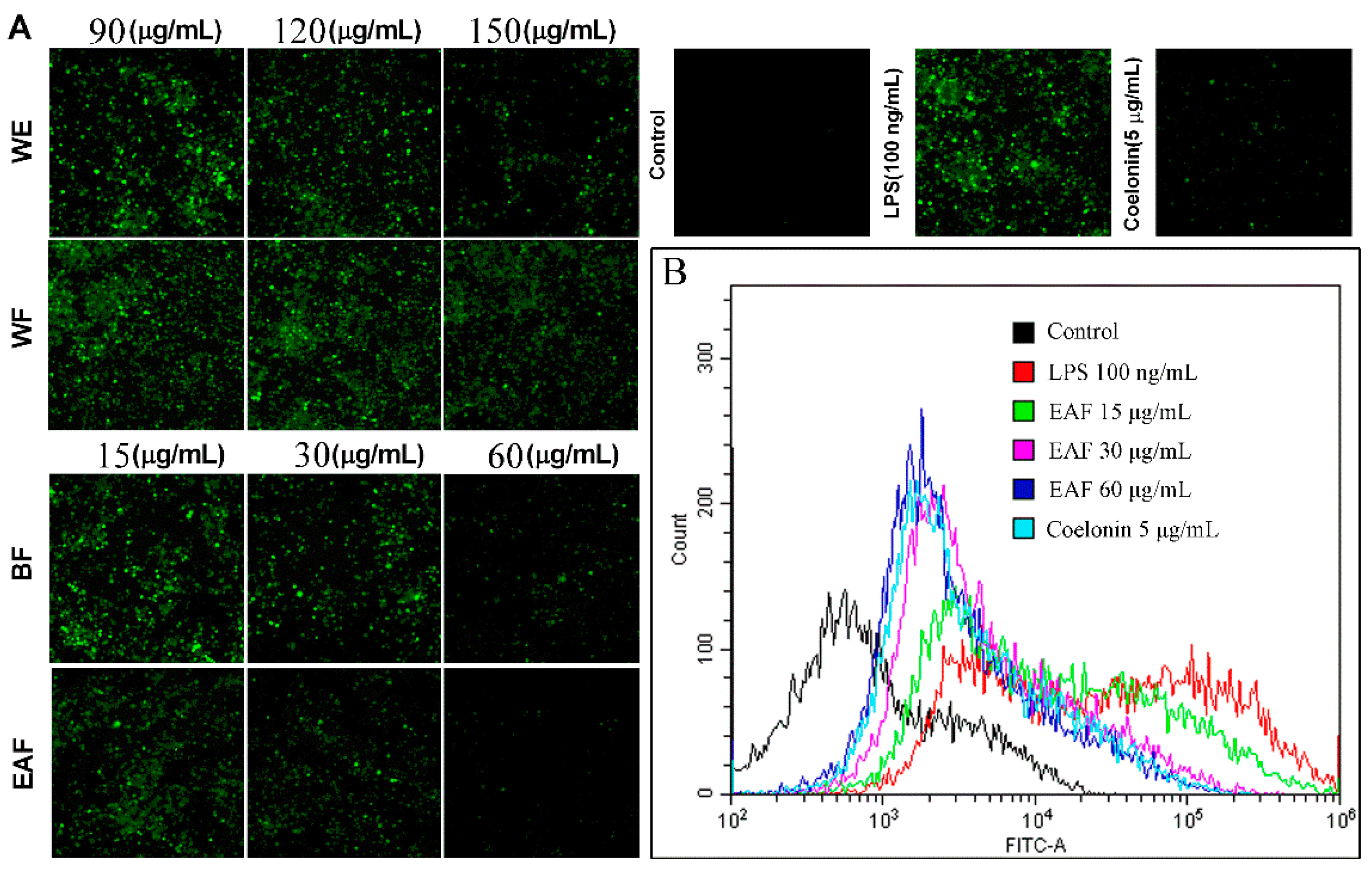
2.6. Effects on NO and Reactive Oxygen Species (ROS) Production on LPS-Induced RAW264.7 Cells
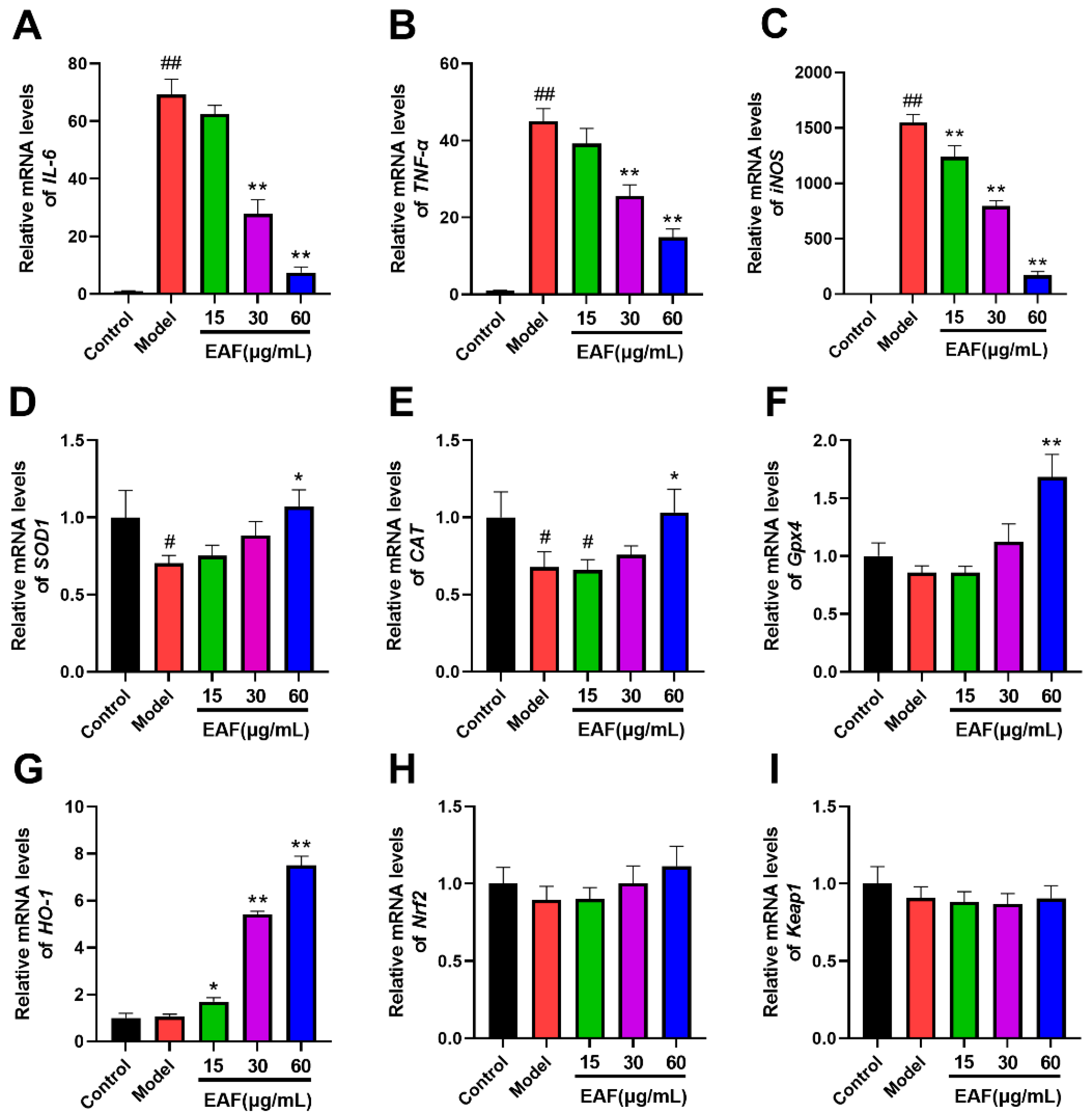
2.7. The Impact of EAF on the Expression of Genes Associated with Inflammation and Oxidative Stress
2.8. EAF Exerts Antioxidant and Anti-Inflammatory Effects through the Regulation of the Nrf2/HO-1 Pathway
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Plant Material
3.3. Sample Preparation
3.4. HPLC Characterization
3.5. UPLC-QTOF-MS2 Conditions
3.6. Determination of Total Phenolic Content (TPC)
3.7. Radical Scanvenging Ability Assays
3.7.1. DPPH• Scanvenging Ability
3.7.2. Reducing Power
3.7.3. ABTS+• Scavenging Activity
3.8. Anti-Inflammatory and Reactive Oxygen Species (ROS) Scavenging Activities on LPS-Induced RAW264.7 Cells
3.8.1. Nitric Oxide (NO) and Cytokines Level in Supernatants
3.8.2. Determination of Intracellular ROS
3.9. Real-Time Fluorescence Quantitative PCR (qPCR) Analysis
3.10. Western Blot Analysis
3.11. Immunofluorescence Analysis
3.12. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abad, M.J.; Bedoya, L.M.; Apaza, L.; Bermejo, P. The Artemisia L. Genus: A review of bioactive essential oils. Molecules 2012, 17, 2542–2566. [Google Scholar] [CrossRef] [PubMed]
- Guetat, A.; Al-Ghamdi, F.A.; Osman, A.K. The genus Artemisia L. In the northern region of Saudi Arabia: Essential oil variability and antibacterial activities. Nat. Prod. Res. 2017, 31, 598–603. [Google Scholar] [CrossRef]
- Koyuncu, I. Evaluation of anticancer, antioxidant activity and phenolic compounds of Artemisia absinthium L. Extract. Cell Mol. Biol. 2018, 64, 25–34. [Google Scholar] [CrossRef]
- Landete, J.M. Updated knowledge about polyphenols: Functions, bioavailability, metabolism, and health. Crit. Rev. Food Sci. Nutr. 2012, 52, 936–948. [Google Scholar] [CrossRef] [PubMed]
- Trendafilova, A.; Moujir, L.M.; Sousa, P.; Seca, A. Research advances on health effects of edible Artemisia species and some sesquiterpene lactones constituents. Foods 2020, 10, 65. [Google Scholar] [CrossRef]
- Huang, T.K.; Ding, Z.D.; Zhao, S.X. Xian Dai Ben Cao Gang Mu: Volume I; Beijing China Medical Science Press: Beijing, China, 2001; p. 1325. ISBN 7-5067-1821-9. [Google Scholar]
- Rashmi, T.R.; Francis, M.S.; Murali, S. Essential oil composition of Artemisia japonica Thunb. from kerala. J. Pharmacogn. Phytochem. 2014, 3, 160–163. [Google Scholar]
- Joshi, R.K. Volatile oil composition of Artemisia japonica Thunb. from western himalaya of uttarakhand. J. Pharmacogn. Phytochem. 2015, 3, 96–97. [Google Scholar]
- Kwon, H.C.; Lee, K.R. Phytochemical constituents of Artemisia japonica ssp. Littoricola. Arch. Pharm. Res. 2001, 24, 194–197. [Google Scholar] [CrossRef]
- Giang, P.M.; Binh, N.T.; Matsunami, K.; Son, P.T. Three new eudesmanes from Artemisia japonica. Nat. Prod. Res. 2014, 28, 631–635. [Google Scholar] [CrossRef]
- Zhang, D.H.; Cheng, P.F.; Ling, L. Antioxidation and genetic toxicity of Artemisia japonica extract. Nat. Prod. RD 2011, 23, 39–42. [Google Scholar]
- Huang, T.H.; Lu, X.M.; Chen, S.L. Pharmacodynamics research and safety evaluation of the folk herb—Artemisia japonica Thumb. J. Chengdu Univ. Tradit. Chin. Med. 2010, 33, 77–79. [Google Scholar]
- Li, N.; Shi, C.; Shi, S.; Wang, H.; Yan, J.; Wang, S. An inulin-type fructan isolated from Artemisia japonica and its anti-arthritic effects. J. Funct. Food 2017, 29, 29–36. [Google Scholar] [CrossRef]
- Zhang, D.H. Study on processing technology for Artemisia japonica tea. Food Ferment. Ind. 2008, 32, 168–171. [Google Scholar]
- Zhang, L.; Tu, Z.C.; Wang, H.; Fu, Z.F.; Wen, Q.H.; Fan, D. Metabolic profiling of antioxidants constituents in Artemisia selengensis leaves. Food Chem. 2015, 186, 123–132. [Google Scholar] [CrossRef]
- Zhang, L.B.; Duan, J.A.; Lv, J.L. Phytochemistry and bioactivities of sesquiterpenoids from the Artemisia species. J. Chin. Pharm. Sci. 2017, 26, 317–334. [Google Scholar] [CrossRef]
- Yu, H.H.; Gao, X.Y. Identification of chemical components in capillary wormwood herb by UPLC-Q-TOF/MS. Cent South Pharm. 2019, 17, 656–661. [Google Scholar]
- Zhang, P.J.; Cao, Y.; Zhang, K.; Song, Y.L.; Li, J.; Tang, L. Chemical profiling of Artemisia rupestris using HPLC-IT-TOF-MS. Chin. Pharm. J. 2020, 45, 4658–4666. [Google Scholar]
- Zhang, L.; Tu, Z.C.; Yuan, T.; Wang, H.; Fu, Z.F.; Wen, Q.H.; Wang, X.Q. Solvent optimization, antioxidant activity, and chemical characterization of extracts from Artemisia selengnesis turcz. Ind. Crop. Prod. 2014, 56, 223–230. [Google Scholar] [CrossRef]
- Gu, D.; Yang, Y.; Abdulla, R.; Aisa, H.A. Characterization and identification of chemical compositions in the extract of Artemisia rupestris L. by liquid chromatography coupled to quadrupole time-of-flight tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2012, 26, 83–100. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical scheme for LC-MSn identification of chlorogenic acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef] [PubMed]
- Fujihara, M.; Muroi, M.; Tanamoto, K.; Suzuki, T.; Azuma, H.; Ikeda, H. Molecular mechanisms of macrophage activation and deactivation by lipopolysaccharide: Roles of the receptor complex. Pharmacol. Ther. 2003, 100, 171–194. [Google Scholar] [CrossRef]
- Jiang, F.S.; Li, M.Y.; Wang, H.Y.; Ding, B.; Zhang, C.C.; Ding, Z.S.; Yu, X.B.; Lv, G.Y. Coelonin, an anti-inflammation active component of Bletilla striata and its potential mechanism. Int. J. Mol. Sci. 2019, 20, 4422. [Google Scholar] [CrossRef]
- Guzik, T.J.; Korbut, R.; Adamek-Guzik, T. Nitric oxide and superoxide in inflammation and immune regulation. J. Physiol. Pharmacol. 2003, 54, 469–487. [Google Scholar]
- Zielonka, J.; Sikora, A.; Joseph, J.; Kalyanaraman, B. Peroxynitrite is the major species formed from different flux ratios of co-generated nitric oxide and superoxide: Direct reaction with boronate-based fluorescent probe. J. Biol. Chem. 2010, 285, 14210–14216. [Google Scholar] [CrossRef]
- Beutler, B.; Du, X.; Poltorak, A. Identification of toll-like receptor 4 (TLR4) as the sole conduit for LPS signal transduction: Genetic and evolutionary studies. J. Endotoxin Res. 2001, 7, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Banskota, S.; Wang, H.; Kwon, Y.H.; Gautam, J.; Haq, S.; Grondin, J.; Steinberg, G.R.; Khan, W.I. Inhibition of nadph oxidase (NOX) 2 mitigates colitis in mice with impaired macrophage AMPK function. Biomedicines 2023, 11, 1443. [Google Scholar] [CrossRef] [PubMed]
- Meng, M.; Jiang, Y.; Wang, Y.; Huo, R.; Ma, N.; Shen, X.; Chang, G. Beta-carotene targets IP3R/GRP75/VDAC1-MCU axis to renovate LPS-induced mitochondrial oxidative damage by regulating STIM1. Free Radic. Biol. Med. 2023, 205, 25–46. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wei, Z.; Wang, X.; Duan, F.; Xiong, L.; Li, J.; Tian, J.; Jia, L.; Gao, H. Premna microphylla Turcz leaf pectin exhibited antioxidant and anti-inflammatory activities in LPS-stimulated RAW264.7 macrophages. Food Chem. 2021, 349, 129164. [Google Scholar] [CrossRef] [PubMed]
- Antar, S.A.; Ashour, N.A.; Marawan, M.E.; Al-Karmalawy, A.A. Fibrosis: Types, effects, markers, mechanisms for disease progression, and its relation with oxidative stress, immunity, and inflammation. Int. J. Mol. Sci. 2023, 24, 4004. [Google Scholar] [CrossRef] [PubMed]
- Luc, K.; Schramm-Luc, A.; Guzik, T.J.; Mikolajczyk, T.P. Oxidative stress and inflammatory markers in prediabetes and diabetes. J. Physiol. Pharmacol. 2019, 70, 809–824. [Google Scholar]
- Neganova, M.; Liu, J.; Aleksandrova, Y.; Klochkov, S.; Fan, R. Therapeutic influence on important targets associated with chronic inflammation and oxidative stress in cancer treatment. Cancers 2021, 13, 6062. [Google Scholar] [CrossRef] [PubMed]
- Andronie-Cioara, F.L.; Ardelean, A.I.; Nistor-Cseppento, C.D.; Jurcau, A.; Jurcau, M.C.; Pascalau, N.; Marcu, F. Molecular mechanisms of neuroinflammation in aging and alzheimer’s disease progression. Int. J. Mol. Sci. 2023, 24, 1869. [Google Scholar] [CrossRef] [PubMed]
- Crimi, E.; Sica, V.; Slutsky, A.S.; Zhang, H.; Williams-Ignarro, S.; Ignarro, L.J.; Napoli, C. Role of oxidative stress in experimental sepsis and multisystem organ dysfunction. Free Radic. Res. 2006, 40, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell. Mol. Life Sci. 2016, 73, 3221–3247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, Y.; Cai, Y.; Li, D.; He, J.; Feng, Z.; Xu, Q. NAT10 regulates the LPS-induced inflammatory response via the NOX2-ROS-NF-kappaB pathway in macrophages. Biochim. Biophys. Acta-Mol. Cell Res. 2023, 1870, 119521. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Jiang, S.; Li, X.; Lin, T.; Qin, T. Astringin protects LPS-induced toxicity by suppressing oxidative stress and inflammation via suppression of PI3K/AKT/NF-kappaB pathway for pediatric acute lung injury. Naunyn-Schmiedebergs Arch. Pharmacol. 2023, 396, 2369–2377. [Google Scholar] [CrossRef] [PubMed]
- Ye, B.; He, D.; Hu, J.; Yang, S.; Gao, X.; Cui, M.; Li, Z.; Wang, H.; Huang, B.; Fu, S.; et al. Notopterol inhibits LPS-induced inflammation in BV-2 cells via AKT/Nrf2/HO-1 signaling axis. Int. Immunopharmacol. 2023, 120, 110334. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Kim, J.Y.; Yang, H.S.; Choe, J.S.; Hwang, I.G. Nepetoidin b from salvia plebeia r. Br. Inhibits inflammation by modulating the NF-kappaB and Nrf2/HO-1 signaling pathways in macrophage cells. Antioxidants 2021, 10, 1208. [Google Scholar] [CrossRef]
- Yang, W.; Zhang, Y.; Lu, D.; Huang, T.; Yan, K.; Wang, W.; Gao, J. Ramelteon protects against human pulmonary microvascular endothelial cell injury induced by lipopolysaccharide (LPS) via activating nuclear factor erythroid 2-related factor 2 (Nrf2)/heme oxygenase-1 (HO-1) pathway. Bioengineered 2022, 13, 1518–1529. [Google Scholar] [CrossRef]
- Lu, H.; Tian, Z.; Cui, Y.; Liu, Z.; Ma, X. Chlorogenic acid: A comprehensive review of the dietary sources, processing effects, bioavailability, beneficial properties, mechanisms of action, and future directions. Compr. Rev. Food. Sci. Food Saf. 2020, 19, 3130–3158. [Google Scholar] [CrossRef]
- Wang, H.N.; Shen, Z.; Liu, Q.; Hou, X.Y.; Cao, Y.; Liu, D.H.; Jiang, H.; Du, H.Z. Isochlorogenic acid (ICGA): Natural medicine with potentials in pharmaceutical developments. Chin. J. Nat. Med. 2020, 18, 860–871. [Google Scholar] [PubMed]
- Krakauer, T. The polyphenol chlorogenic acid inhibits Staphylococcal exotoxin-induced inflammatory cytokines and chemokines. Immunopharmacol. Immunotoxicol. 2002, 24, 113–119. [Google Scholar] [CrossRef]
- Hwang, S.J.; Kim, Y.W.; Park, Y.; Lee, H.J.; Kim, K.W. Anti-inflammatory effects of chlorogenic acid in lipopolysaccharide-stimulated RAW264.7 cells. Inflamm. Res. 2014, 63, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Park, S.Y.; Park, Y.L.; Myung, D.S.; Rew, J.S.; Joo, Y.E. Chlorogenic acid suppresses lipopolysaccharide induced nitric oxide and interleukin1β expression by inhibiting JAK2/STAT3 activation in RAW264.7 cells. Mol. Med. Rep. 2017, 16, 9224–9232. [Google Scholar] [CrossRef]
- Liao, W.C.; Lai, Y.C.; Yuan, M.C.; Hsu, Y.L.; Chan, C.F. Antioxidative activity of water extract of sweet potato leaves in Taiwan. Food Chem. 2011, 127, 1224–1228. [Google Scholar] [CrossRef]
- Brindisi, M.; Bouzidi, C.; Frattaruolo, L.; Loizzo, M.R.; Tundis, R.; Dugay, A.; Deguin, B.; Cappello, A.R.; Cappello, M.S. Chemical profile, antioxidant, anti-inflammatory, and anti-cancer effects of Italian Salvia rosmarinus Spenn. methanol leaves extracts. Antioxidants 2020, 9, 826. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Rt (min) | Exptl. [M-H]− | Theor. [M-H]− | Error (ppm) | Fragments | Chemical Formula | Tentative Identification | Family (Subclass) |
|---|---|---|---|---|---|---|---|---|
| 1 | 0.8136 | 191.0558 | 191.0561 | −1.6994 | 173.04667, 127.03973, 111.04506, 109.02987, 93.03444, 87.00940, 85.02896 | C7H12O6 | Quinic acid | Organic acids |
| 2 | 2.6626 | 353.0890 | 353.0878 | 3.3511 | 191.05583, 179.03461, 135.04426, 127.04007, 85.02972 | C16H18O9 | 1-O-Caffeoylquinic acid | Phenolic acid (Cinnamic acid) |
| 3 | 3.7991 | 353.0879 | 353.0878 | 0.3652 | 215.05802, 191.05548, 179.03509, 135.04519, 127.03939, 85.02859 | C16H18O9 | 5-O-Caffeoylquinic acid 1 | Phenolic acid (Cinnamic acid) |
| 4 | 4.0636 | 353.0879 | 353.0878 | 0.1970 | 191.05580, 179.03444, 173.04576, 135.04410, 127.04087 | C16H18O9 | 4-O-Caffeoylquinic acid | Phenolic acid (Cinnamic acid) |
| 5 | 4.1107 | 593.1528 | 593.1512 | 2.6666 | 463.09090, 408.03010, 239.05638, 163.03970 | C27H30O15 | Kaempferol-3-neohesperidoside | Flavonoid (Flavonol) |
| 6 | 4.1301 | 177.0558 | 177.0557 | 0.4088 | 163.04033, 149.06059 | C10H10O3 | 4-Hydroxy-3-methoxycinnamaldehyde | Phenolics |
| 7 | 4.3013 | 179.0343 | 179.0350 | −3.5685 | 161.02510, 135.04402, 133.03013, 93.03377 | C9H8O4 | Caffeic acid | Phenolic acid (Cinnamic acid) |
| 8 | 4.6835 | 367.1053 | 367.1035 | 4.8973 | 193.05058, 134.03673 | C17H20O9 | 5-O-Feruloylquinic acid | Phenolic acid (Cinnamic acid) |
| 9 | 4.7968 | 353.0889 | 353.0878 | 2.9779 | 191.05601 | C16H18O9 | 3-O-Caffeoylquinic acid | Phenolic acid (Cinnamic acid) |
| 10 | 5.0113 | 337.0945 | 337.0929 | 4.6514 | 191.05673 | C16H18O8 | p-Coumaroylquinic acid | Phenolic acid (Cinnamic acid) |
| 11 | 5.3766 | 593.1537 | 593.1512 | 4.2190 | 515.12411, 353.08952, 191.05683 | C27H30O15 | Luteolin-7-O-rutinoside | Flavonoid (Flavone) |
| 12 | 5.8298 | 357.1207 | 357.1191 | 4.5729 | 195.06719, 151.07656 | C16H22O9 | Sweroside | Terpenoids (Iridoid) |
| 13 | 6.1530 | 367.1042 | 367.1035 | 1.9545 | 193.05224, 173.04529, 134.03753 | C17H20O9 | 4-O-Feruloylquinic acid | Phenolic acid (Cinnamic acid) |
| 14 | 6.2482 | 367.1042 | 367.1035 | 2.0352 | 193.05114, 173.04534, 134.03790 | C17H20O9 | 3-O-Feruloylquinic acid | Phenolic acid (Cinnamic acid) |
| 15 | 6.2988 | 563.1430 | 563.1406 | 4.2293 | 447.0958 | C26H28O14 | Isoshaftoside | Flavonoid (Flavone) |
| 16 | 6.3239 | 609.1474 | 609.1461 | 2.0435 | 563.14221, 447.09580, 271.02368, 161.02540 | C27H30O16 | Kaempferol-3-O-glucosyl (1-2) galactoside | Flavonoid (Flavonol) |
| 17 | 6.5276 | 463.0897 | 463.0882 | 3.3226 | 301.03689 | C21H20O12 | Quercetin-3-O-β-d-glucoside isomer | Flavonoid (Flavonol) |
| 18 | 6.6653 | 341.1250 | 341.1242 | 2.5118 | 193.05047, 149.05998, 133.02946 | C16H22O8 | citrusin D | Phenolics |
| 19 | 6.6808 | 193.0507 | 193.0506 | 0.4165 | 149.05998, 134.03643, 133.02946 | C10H10O4 | Ferulic acid 2 | Phenolic acid (Cinnamic acid) |
| 20 | 6.8748 | 463.0882 | 463.0882 | 0.1086 | 301.03617, 300.02932, 283.02590, 151.00372 | C21H20O12 | Quercetin-7-O-β-d-glucopyranoside | Flavonoid (Flavonol) |
| 21 | 7.2931 | 609.1489 | 609.1461 | 4.6232 | 537.12653, 287.05783, 175.02609, 151.00379, 135.04537 | C27H30O16 | Kaempferol-3-O-glucosyl(1-2)galactoside isomer | Flavonoid (Flavonol) |
| 22 | 7.3235 | 463.0891 | 463.0882 | 2.0099 | 300.03021, 151.00344 | C21H20O12 | Quercetin-3-O-β-d-glucoside isomer | Flavonoid (Flavonol) |
| 23 | 7.4854 | 593.1530 | 593.1512 | 3.0739 | 533.13349 | C27H30O15 | Luteolin-7-rutinoside isomer | Flavonoid (Flavone) |
| 24 | 7.5717 | 463.0893 | 463.0882 | 2.3953 | 301.03636, 255.03164 | C21H20O12 | Quercetin-3-O-β-d-glucoside isomer | Flavonoid (Flavonol) |
| 25 | 7.6157 | 461.0730 | 461.0725 | 0.9859 | 285.04095, 201.02004, 151.00407 | C21H18O12 | Kaempferol-3-O-glucuronide | Flavonoid (Flavonol) |
| 26 | 7.6969 | 447.0950 | 447.0933 | 3.7809 | 323.07978, 285.04160 | C21H20O11 | Kaempferol-7-O-glucoside | Flavonoid (Flavonol) |
| 27 | 7.7526 | 463.1240 | 463.1246 | −1.2866 | 229.01497 | C22H24O11 | Eriodictiol-7-glucoside | Flavonoid (Dihydroflavone) |
| 28 | 7.8486 | 515.1203 | 515.1195 | 1.5494 | 353.08896, 315.05243, 191.05748, 179.03625 | C25H24O12 | Dicaffeoylquinic acid isomer | Phenolic acid (Cinnamic acid) |
| 29 | 8.1879 | 515.1190 | 515.1195 | −0.8976 | 353.08833, 335.07794, 191.05580 | C25H24O12 | 3,4-O-Dicaffeoylquinic acid 1 | Phenolic acid (Cinnamic acid) |
| 30 | 8.3397 | 487.1262 | 487.1246 | 3.2261 | 295.06312, 191.05590 | C24H24O11 | Acacetin-7-O-(6″-O-acetyl)-β-d-glucopyranoside | Flavonoid (Flavone) |
| 31 | 8.4579 | 515.1179 | 515.1195 | −3.0203 | 353.08884, 335.08051, 191.05588, 179.03469, 173.04548, 161.02510, 135.04450 | C25H24O12 | 3,5-O-Dicaffeoylquinic acid 1 | Phenolic acid (Cinnamic acid) |
| 32 | 8.6790 | 353.0888 | 353.0878 | 2.7676 | 181.04996 | C16H18O9 | Scopolin | phenylpropanoid (Coumarin) |
| 33 | 8.7417 | 187.0977 | 187.0976 | 0.5508 | 125.09792 | C9H16O4 | Azelaic acid 2 | Organic acids |
| 34 | 8.8887 | 431.1005 | 431.0984 | 4.8819 | 269.04707, 268.03982 | C21H20O10 | Apigenin-7-O-glucoside | Flavonoid (Flavone) |
| 35 | 8.9063 | 269.0463 | 269.0455 | 2.7163 | 227.03775, 151.00411, 117.03409 | C15H10O5 | Apigenin | Flavonoid (Flavone) |
| 36 | 9.1441 | 515.1187 | 515.1195 | −1.5220 | 353.08859, 335.07898, 191.05625, 179.03476, 173.04525, 161.02518, 135.04435 | C25H24O12 | 4,5-O-Dicaffeoylquinic acid 1 | Phenolic acid (Cinnamic acid) |
| 37 | 9.1444 | 161.0244 | 161.0244 | 0.0043 | 137.02529, 93.03450 | C9H6O3 | 7-Hydroxycoumarin | phenylpropanoid (Coumarin) |
| 38 | 9.7478 | 461.1080 | 461.1089 | −2.1180 | 323.07777 | C22H22O11 | Diosmetin-7-O-β-d-glucopyranosid | Flavonoid (Flavone) |
| 39 | 9.8104 | 549.1991 | 549.1978 | 2.4796 | 387.16709, 207.10416, 161.02516 | C27H34O12 | Vitexnegheteroin M | Phenolic acid (Cinnamic acid) |
| 40 | 10.1438 | 529.1362 | 529.1351 | 2.0388 | 367.10485, 335.08081, 301.03970, 173.04600 | C26H26O12 | Caffeoylferuloylquinic acid isomer | Phenolic acid (Cinnamic acid) |
| 41 | 10.4875 | 529.1360 | 529.1351 | 1.6767 | 499.12753, 367.10461, 135.04617 | C26H26O12 | Caffeoylferuloylquinic acid isomer | Phenolic acid (Cinnamic acid) |
| 42 | 10.5746 | 515.1200 | 515.1195 | 0.9653 | 353.08906, 191.05667, 179.03605, 173.04642 | C25H24O12 | Dicaffeoylquinic acid isomer | Phenolic acid (Cinnamic acid) |
| 43 | 11.0945 | 529.1361 | 529.1351 | 1.7751 | 367.10666, 335.08190, 271.09856, 179.03642, 173.04594, 135.04597 | C26H26O12 | Caffeoylferuloylquinic acid isomer | Phenolic acid (Cinnamic acid) |
| 44 | 11.5266 | 207.0658 | 207.0663 | −2.1425 | 179.03595, 135.04408 | C11H12O4 | Ethyl caffeate | Phenolic acid (Cinnamic acid) |
| 45 | 11.9334 | 677.1515 | 677.1512 | 0.5120 | 555.11800, 515.12006, 353.08866, 335.07949, 191.05651, 179.03505, 173.04614 | C34H30O15 | 3,4,5-Tricaffeoylquinic acid | Phenolic acid (Cinnamic acid) |
| 46 | 12.1084 | 283.0621 | 283.0612 | 3.1644 | 268.03964 | C16H12O5 | Acacetin | Flavonoid (Flavone) |
| 47 | 16.3310 | 293.1760 | 293.1758 | 0.6382 | 236.10563, 221.15397, 220.14706, 192.11523 | C17H26O4 | 7β-Acetoxy-2β-hydroxyoplopenone | Terpenoids (Sesquiterpenoids) |
| 48 | 20.1050 | 233.1552 | 233.1547 | 2.1908 | 221.15524 | C15H22O2 | Artemisic acid 2 | Terpenoids (Sesquiterpenoids) |
| Fractions | Yield (%) | CA (mg/g) | IAA (mg/g) | IAB (mg/g) | IAC (mg/g) |
|---|---|---|---|---|---|
| WE | 25.400 ± 1.600 * | 45.206 ± 2.494 | 14.880 ± 1.363 | 18.610 ±2.459 | 23.048 ± 1.757 |
| WF | 72.719 ± 2.719 | 20.856 ± 3.700 | 0.000 | 0.000 | 1.957 ± 0.099 |
| BF | 21.983 ± 0.715 | 106.814 ± 4.122 | 19.115 ± 0.545 | 53.721 ± 1.790 | 41.916 ± 2.807 |
| EAF | 4.417 ± 0.775 | 28.677 ± 4.714 | 188.537 ± 1.521 | 100.851 ± 2.353 | 201.602 ± 2.866 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, Y.; Li, X.; Chen, M.; Wang, X.; Li, M.; Jiang, F.; Zhang, X.; Zhang, C.; Li, S. The Extracts Derived from Artemisia japonica Thunb. Leaves Mitigate Oxidative Stress and Inflammatory Response Induced by LPS in RAW264.7 Cells through Modulation of the Nrf2/HO-1 Signaling Pathway. Molecules 2024, 29, 1375. https://doi.org/10.3390/molecules29061375
Ye Y, Li X, Chen M, Wang X, Li M, Jiang F, Zhang X, Zhang C, Li S. The Extracts Derived from Artemisia japonica Thunb. Leaves Mitigate Oxidative Stress and Inflammatory Response Induced by LPS in RAW264.7 Cells through Modulation of the Nrf2/HO-1 Signaling Pathway. Molecules. 2024; 29(6):1375. https://doi.org/10.3390/molecules29061375
Chicago/Turabian StyleYe, Yueyu, Xiaomei Li, Man Chen, Xia Wang, Meiya Li, Fusheng Jiang, Xiaobo Zhang, Chunchun Zhang, and Shiqing Li. 2024. "The Extracts Derived from Artemisia japonica Thunb. Leaves Mitigate Oxidative Stress and Inflammatory Response Induced by LPS in RAW264.7 Cells through Modulation of the Nrf2/HO-1 Signaling Pathway" Molecules 29, no. 6: 1375. https://doi.org/10.3390/molecules29061375
APA StyleYe, Y., Li, X., Chen, M., Wang, X., Li, M., Jiang, F., Zhang, X., Zhang, C., & Li, S. (2024). The Extracts Derived from Artemisia japonica Thunb. Leaves Mitigate Oxidative Stress and Inflammatory Response Induced by LPS in RAW264.7 Cells through Modulation of the Nrf2/HO-1 Signaling Pathway. Molecules, 29(6), 1375. https://doi.org/10.3390/molecules29061375