
Nodakenin Ameliorates Ovariectomy-Induced Bone Loss by Regulating Gut Microbiota

 ,
,
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
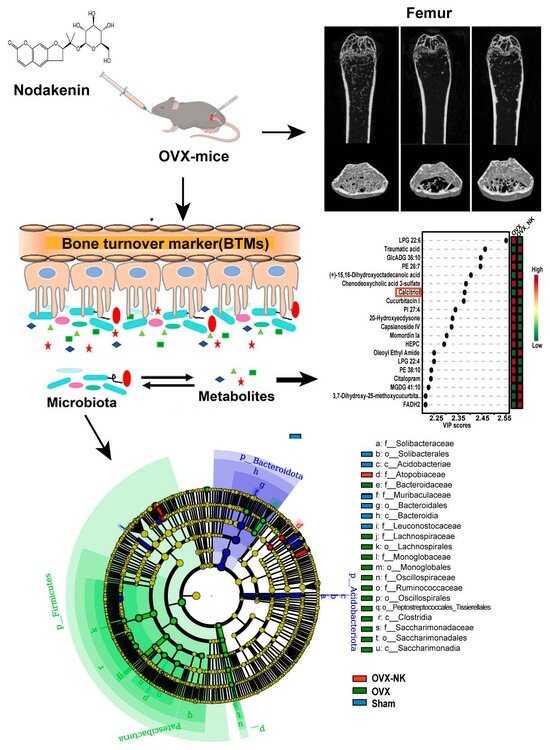
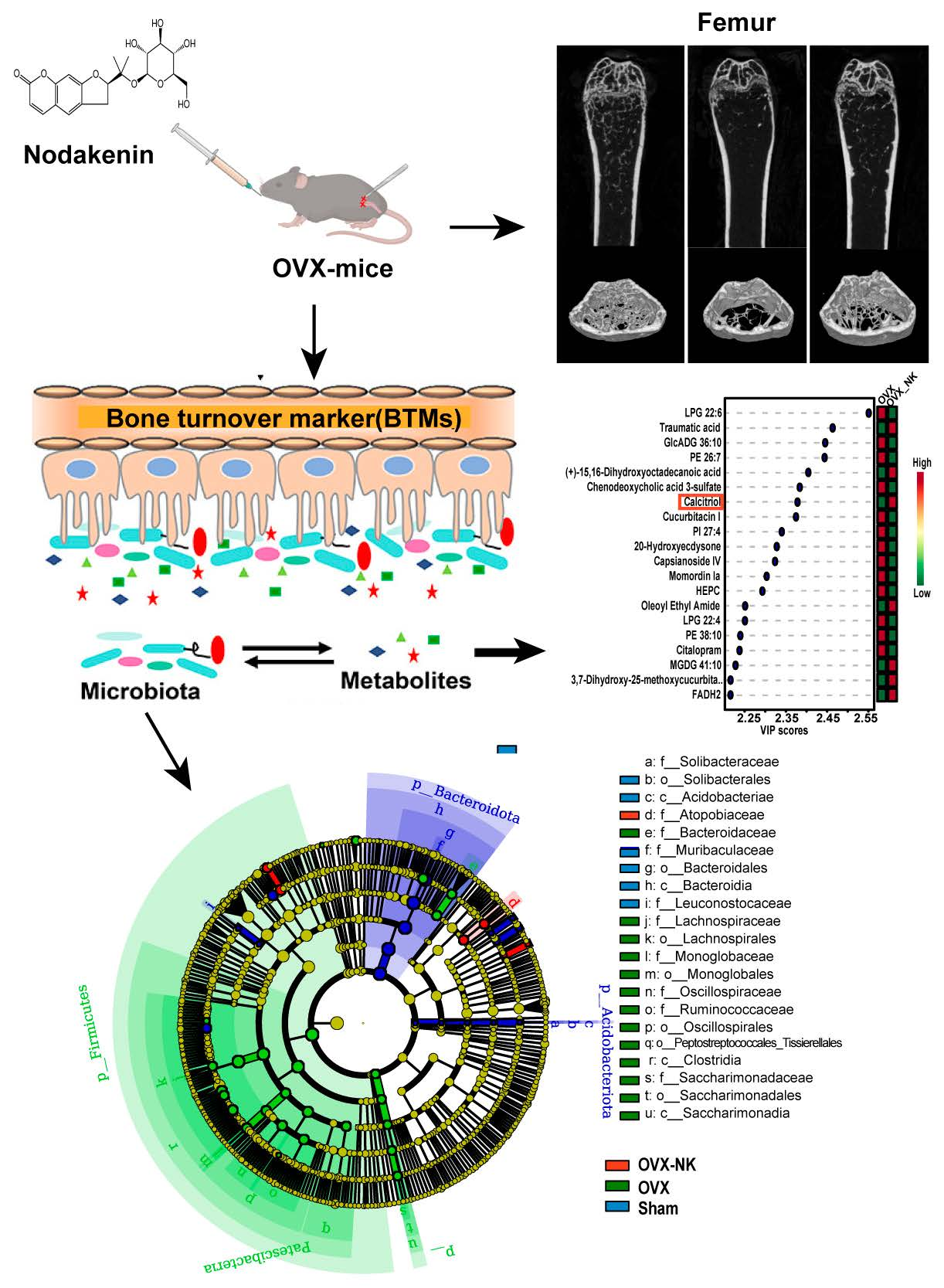
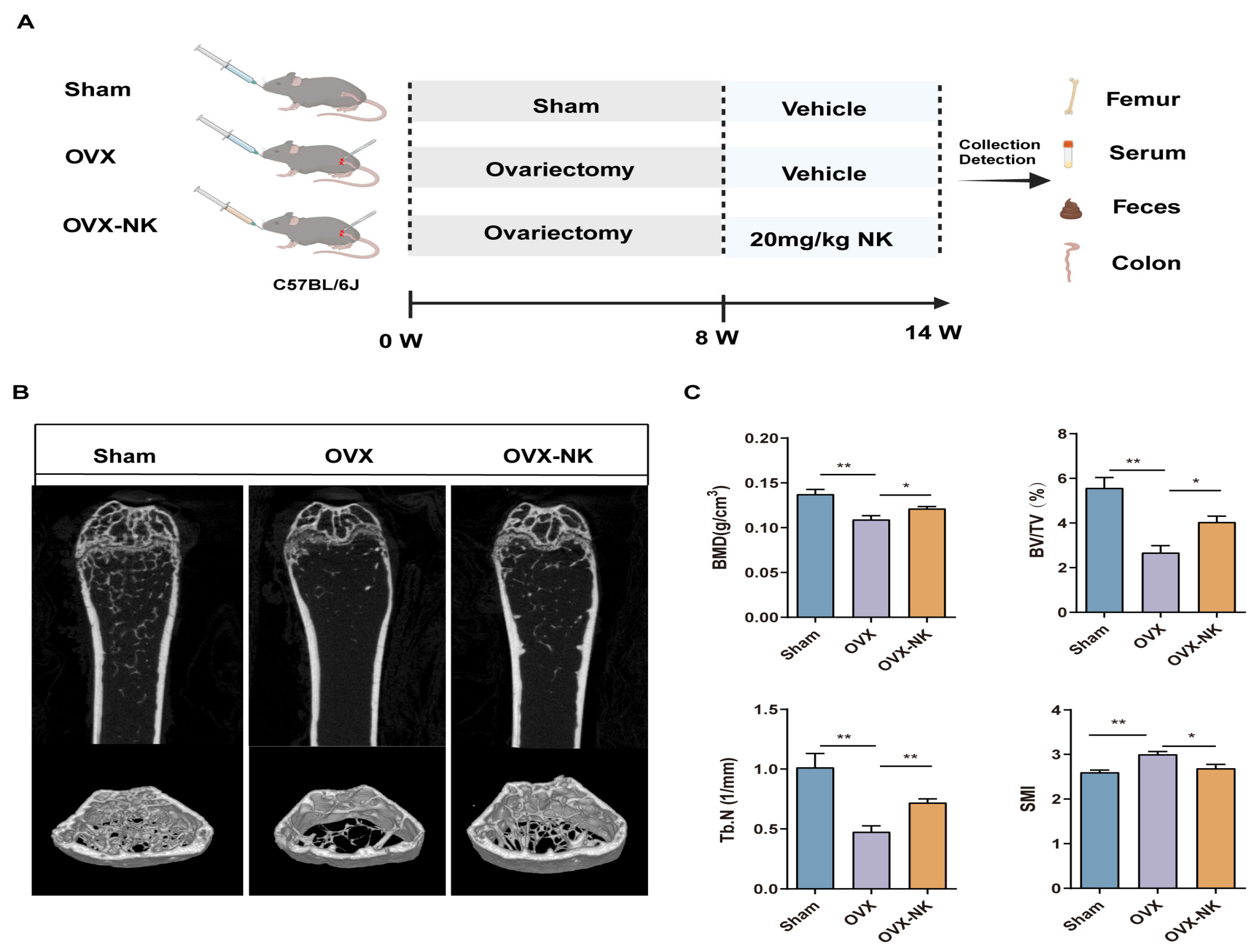
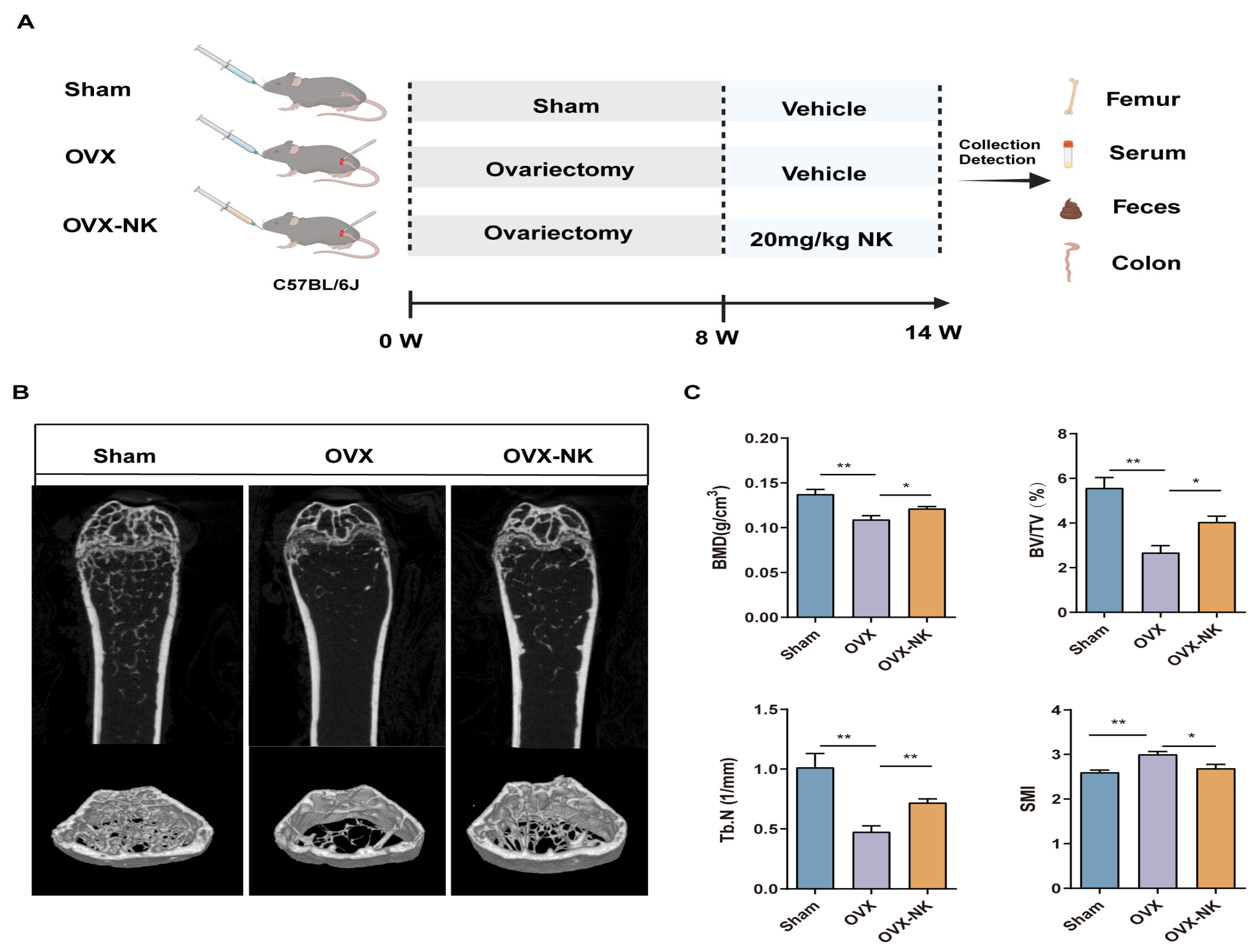
2.1. Nodakenin Alleviates Osteoporosis in OVX Mice
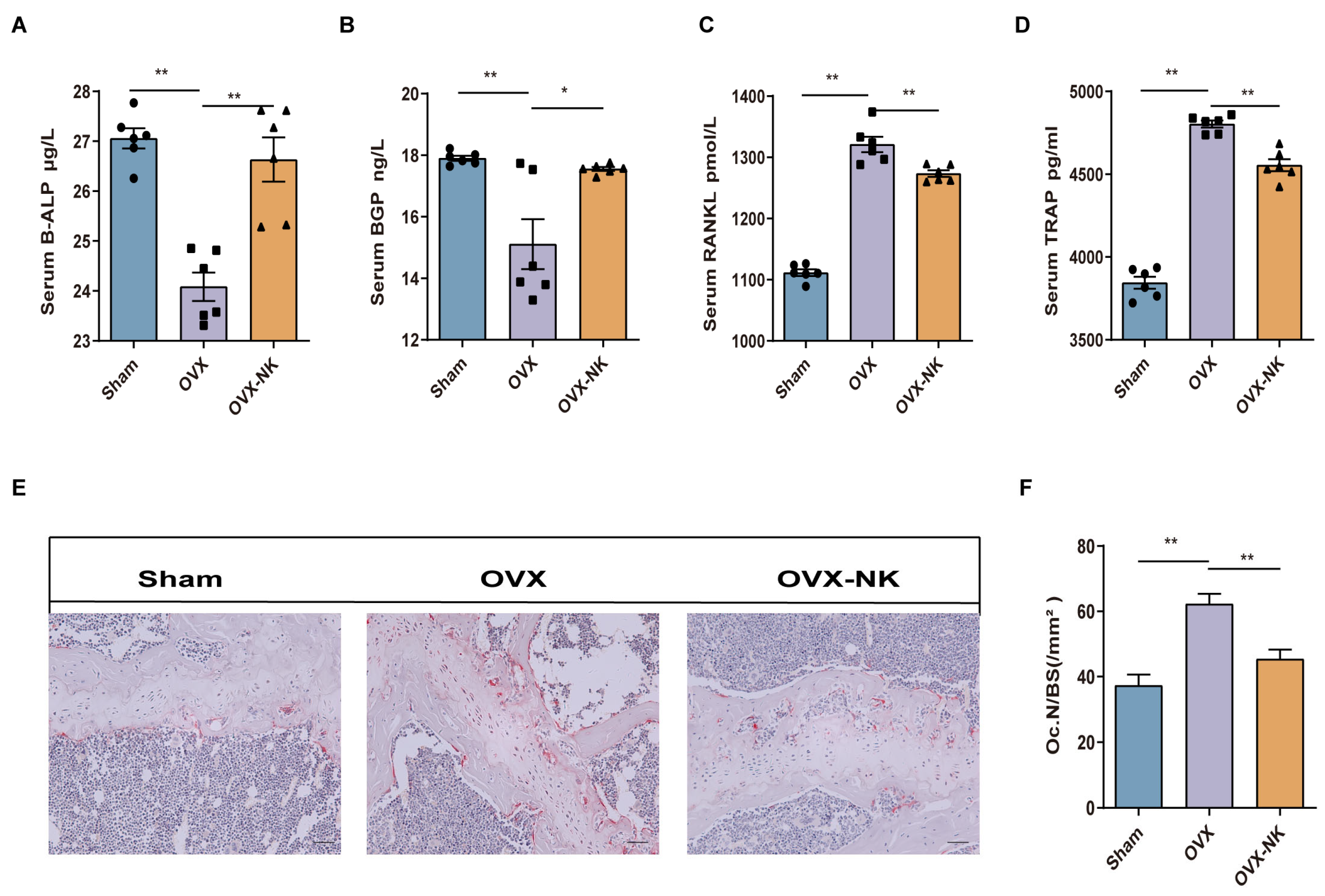
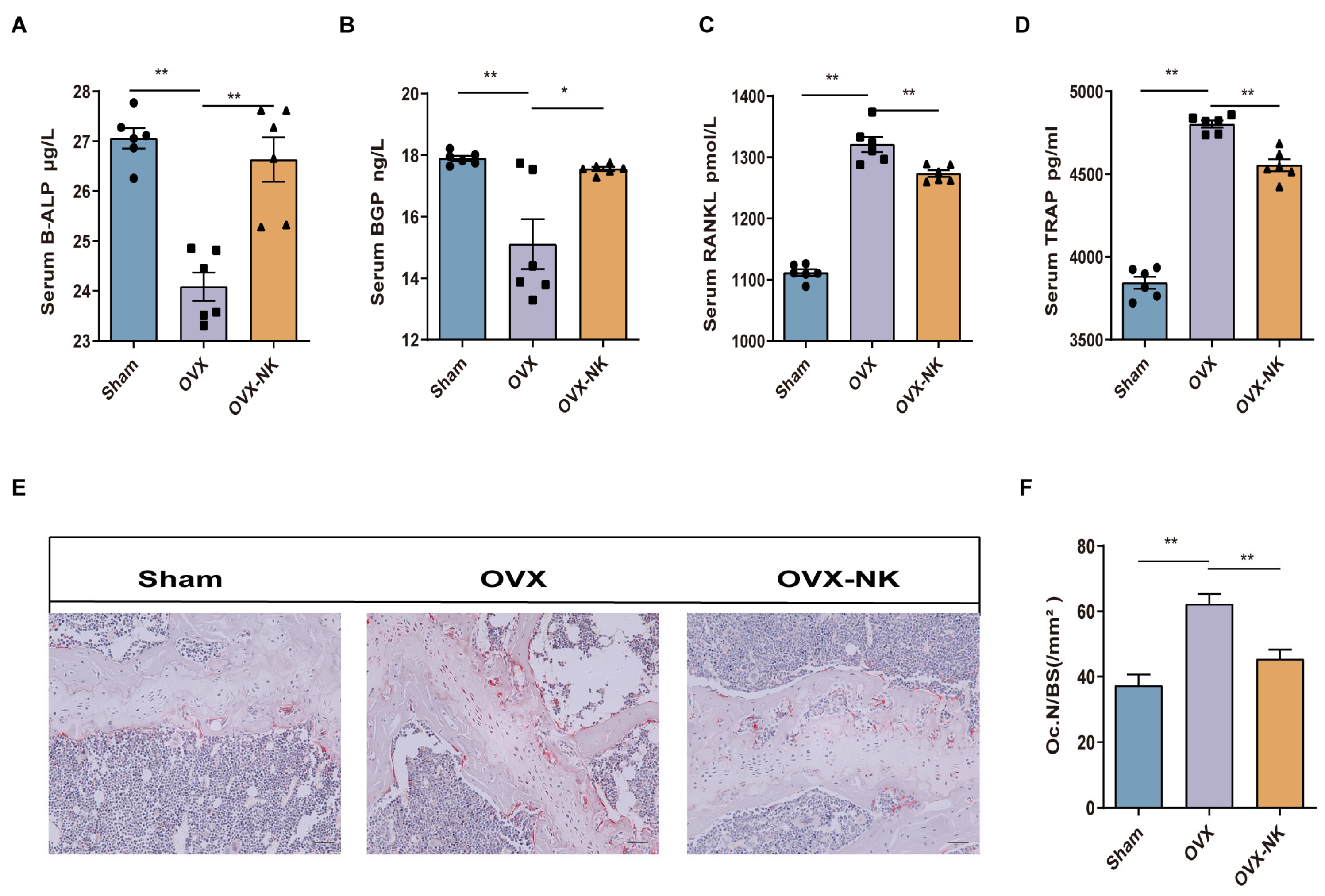
2.2. Nodakenin Regulates Estrogen Deficiency-Induced Bone Turnover
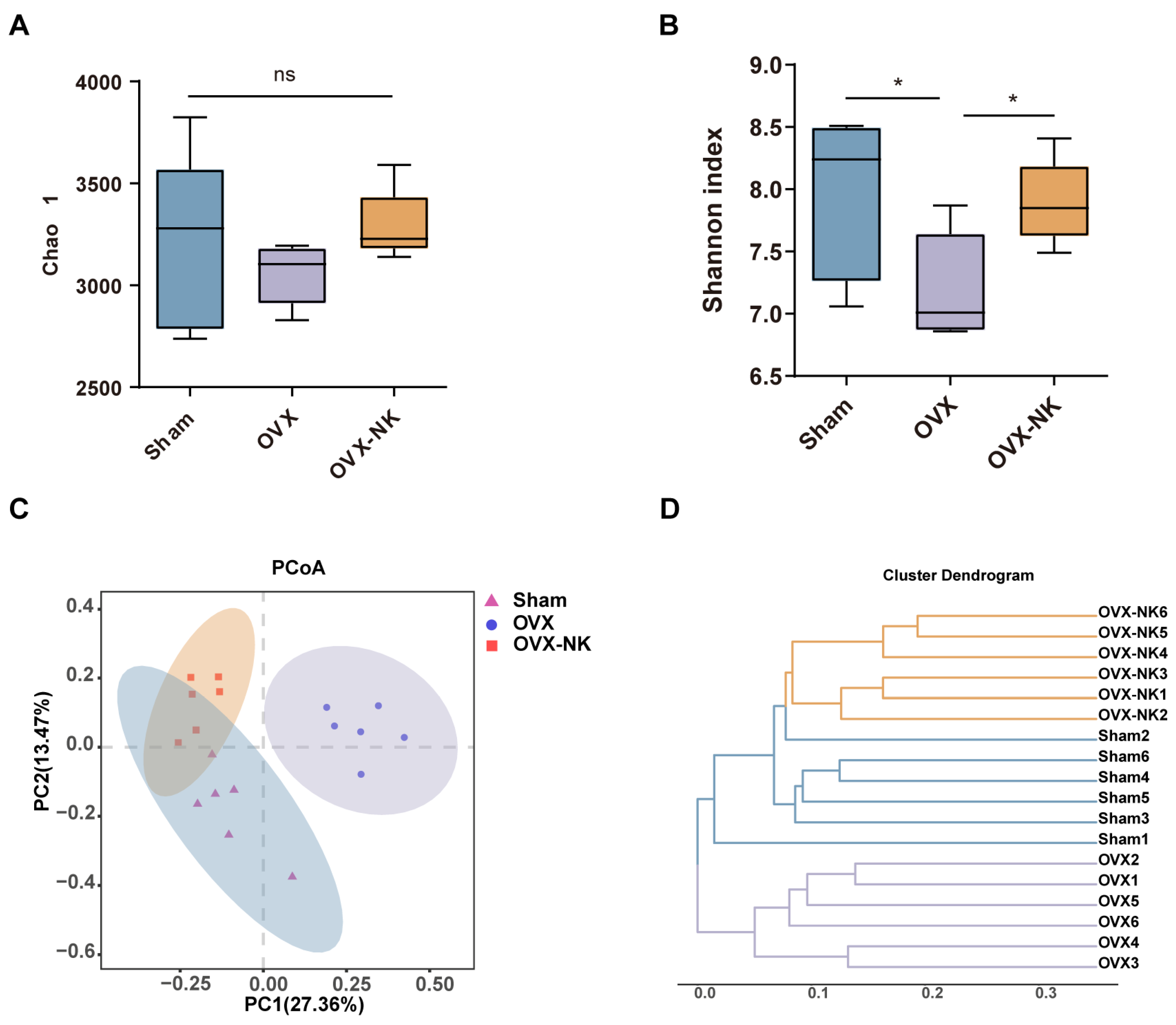
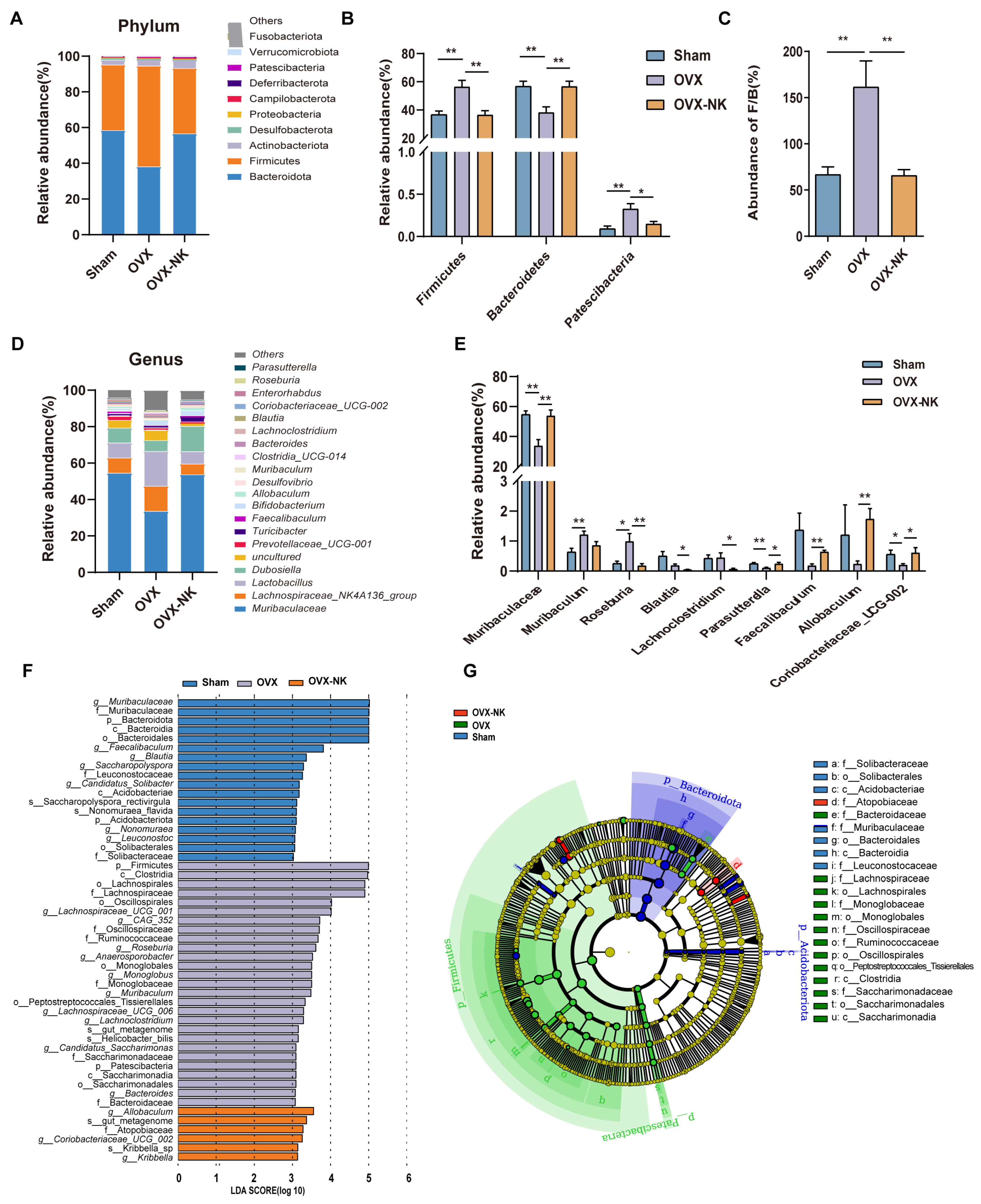
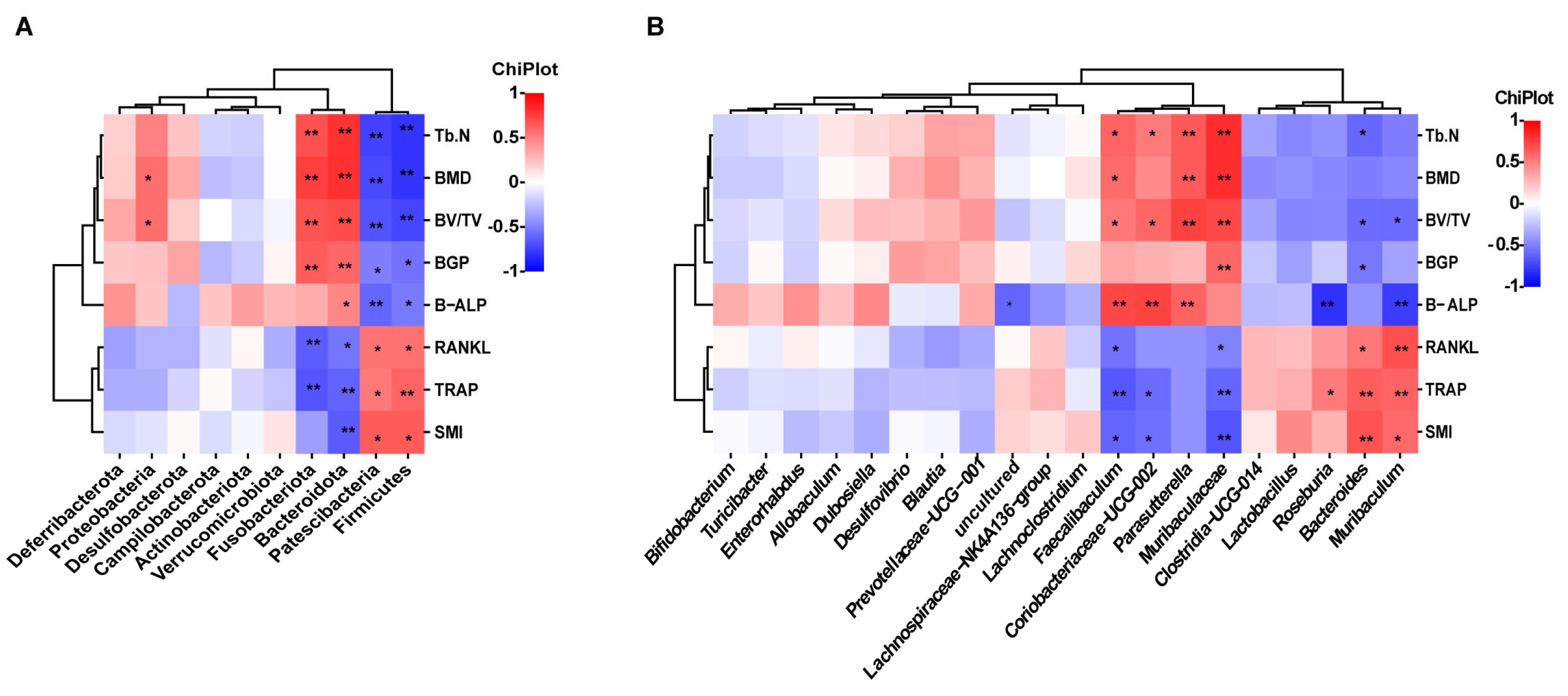
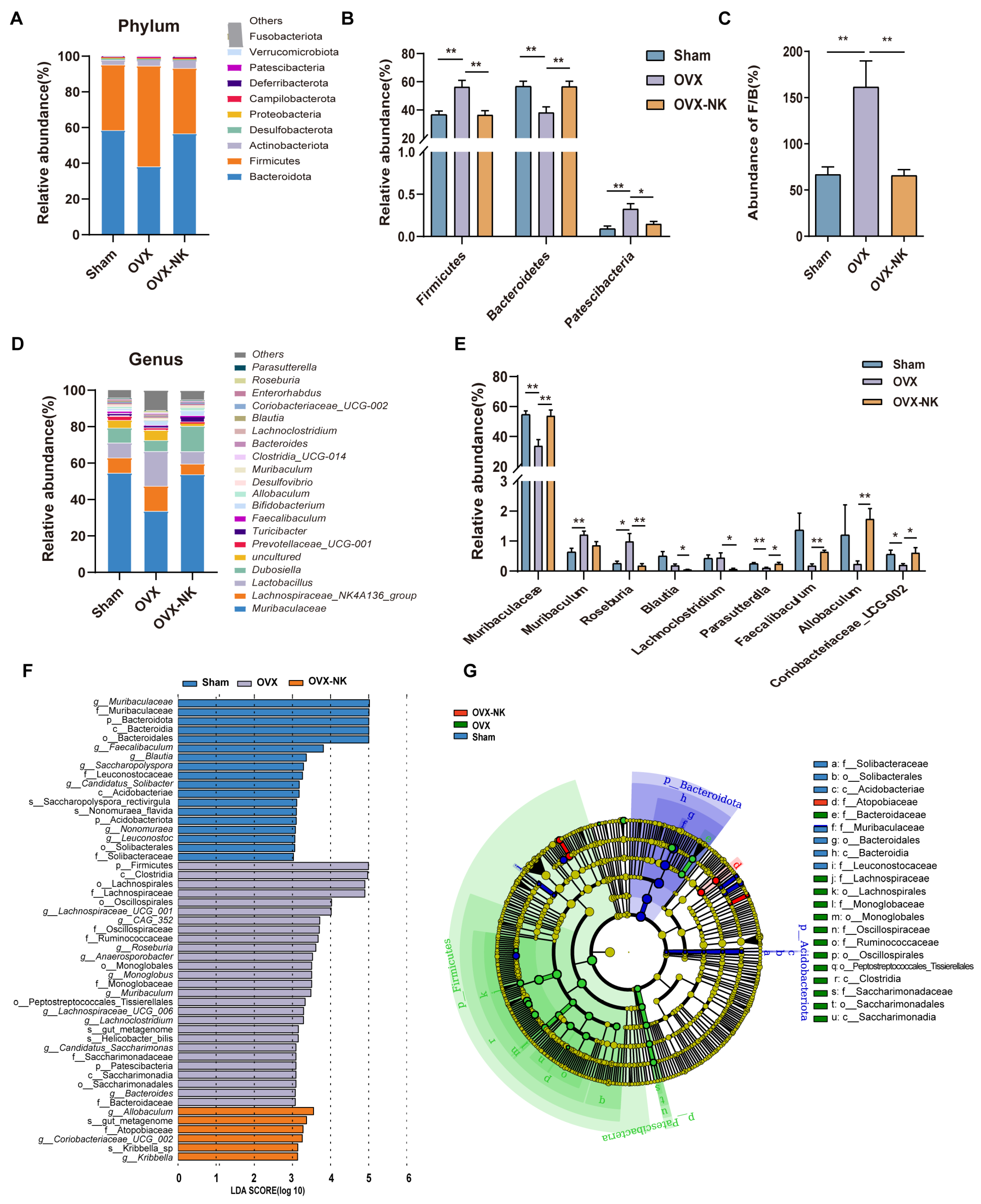
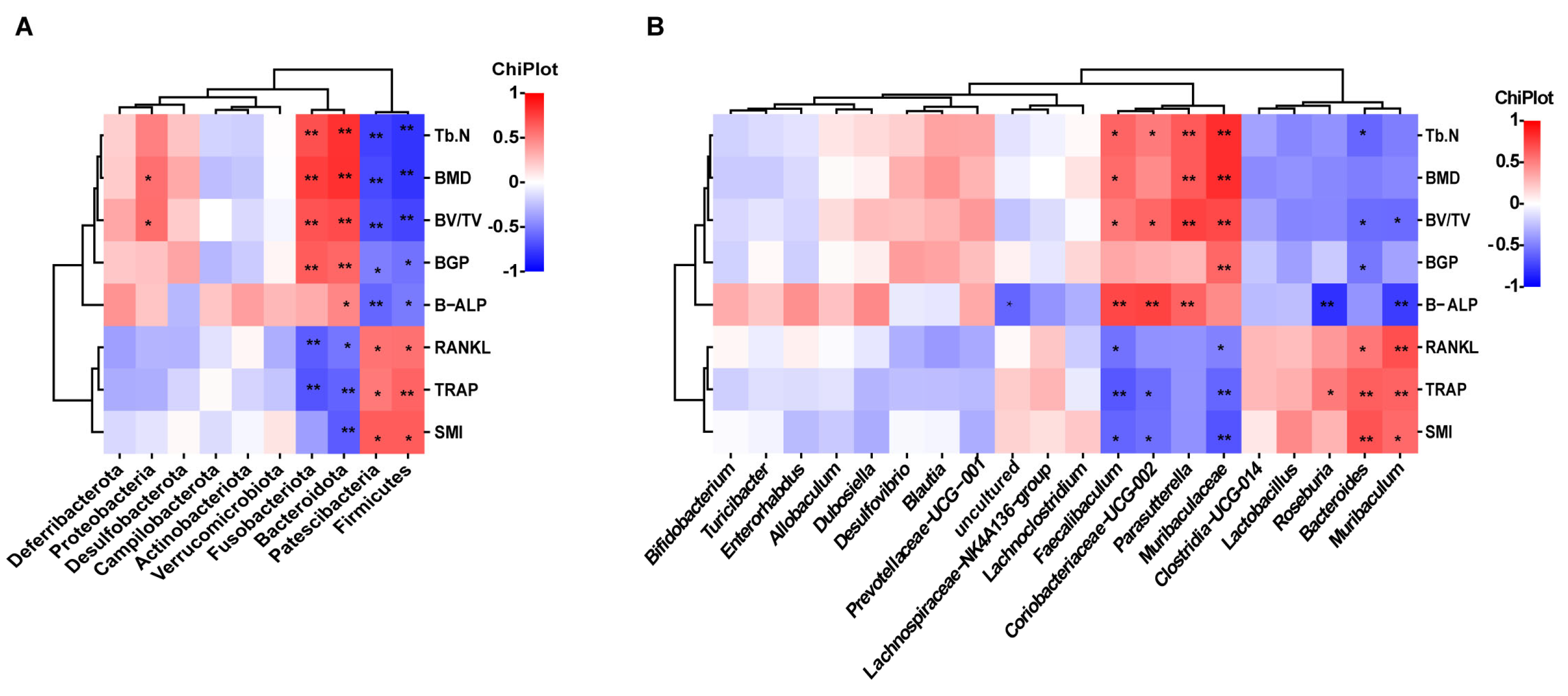
2.3. Nodakenin Alters OVX-Induced Gut Microbiota Imbalance in Mice
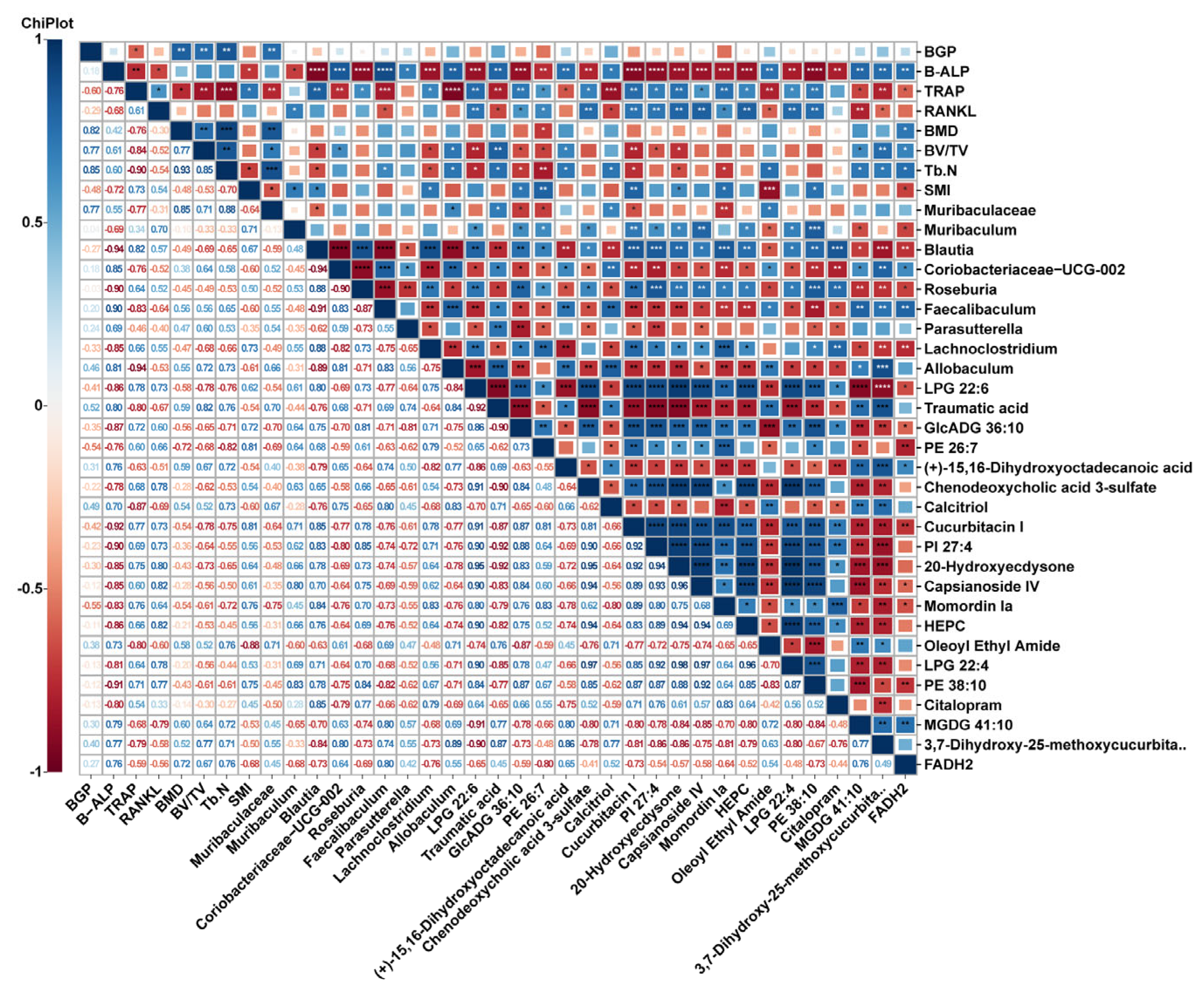
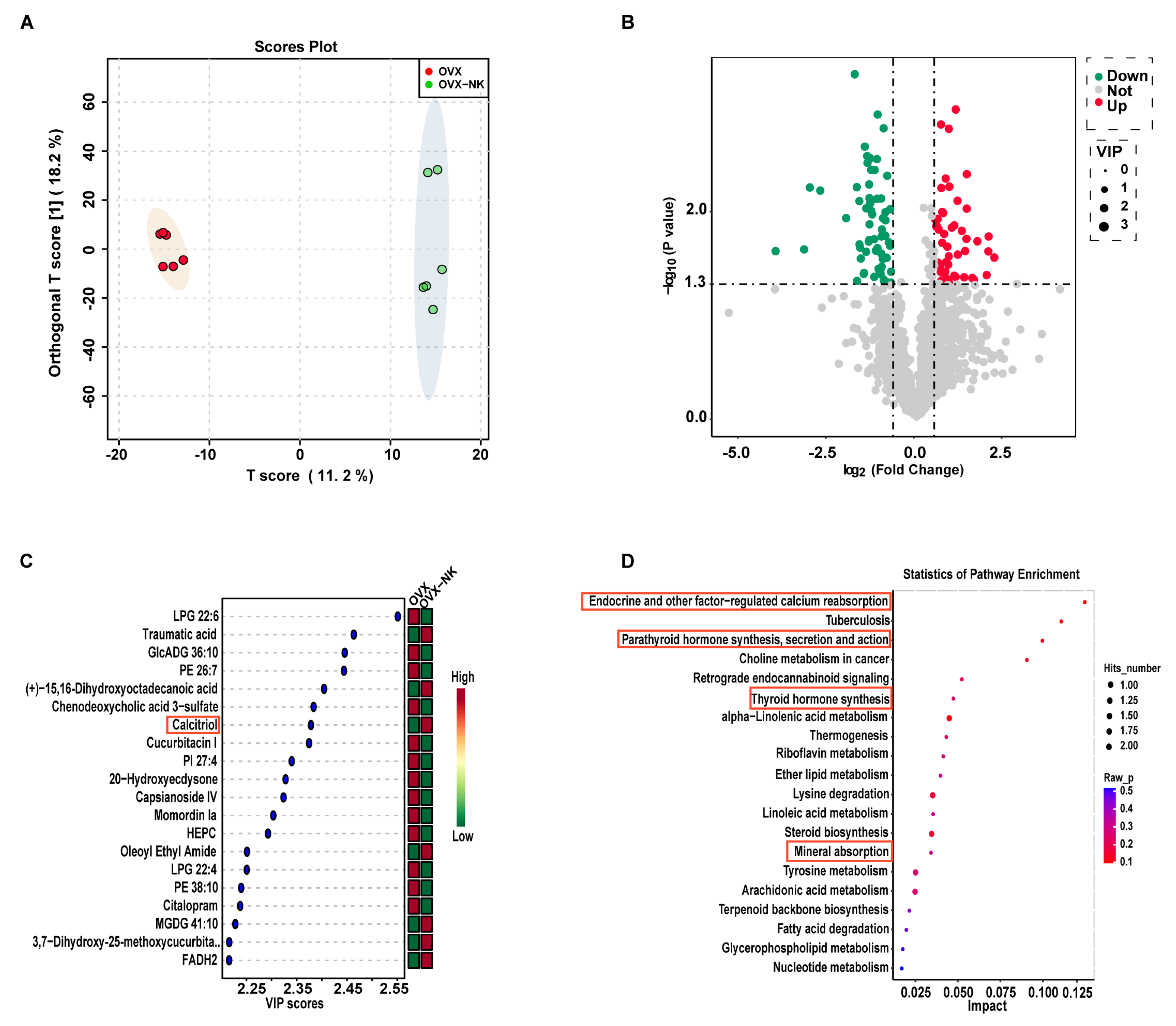
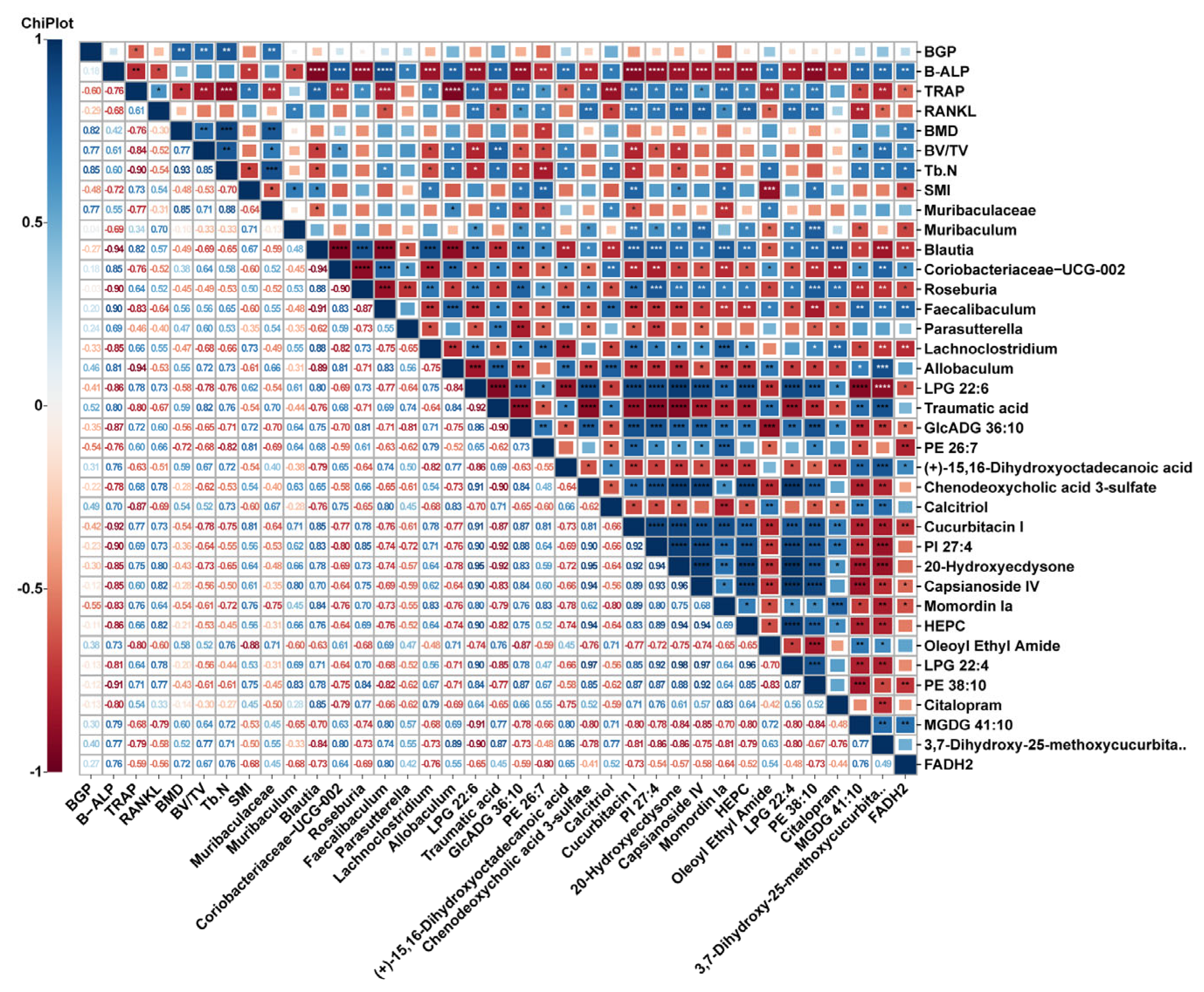
2.4. Metabolite Profiles Change Caused by Nodakenin Treatment
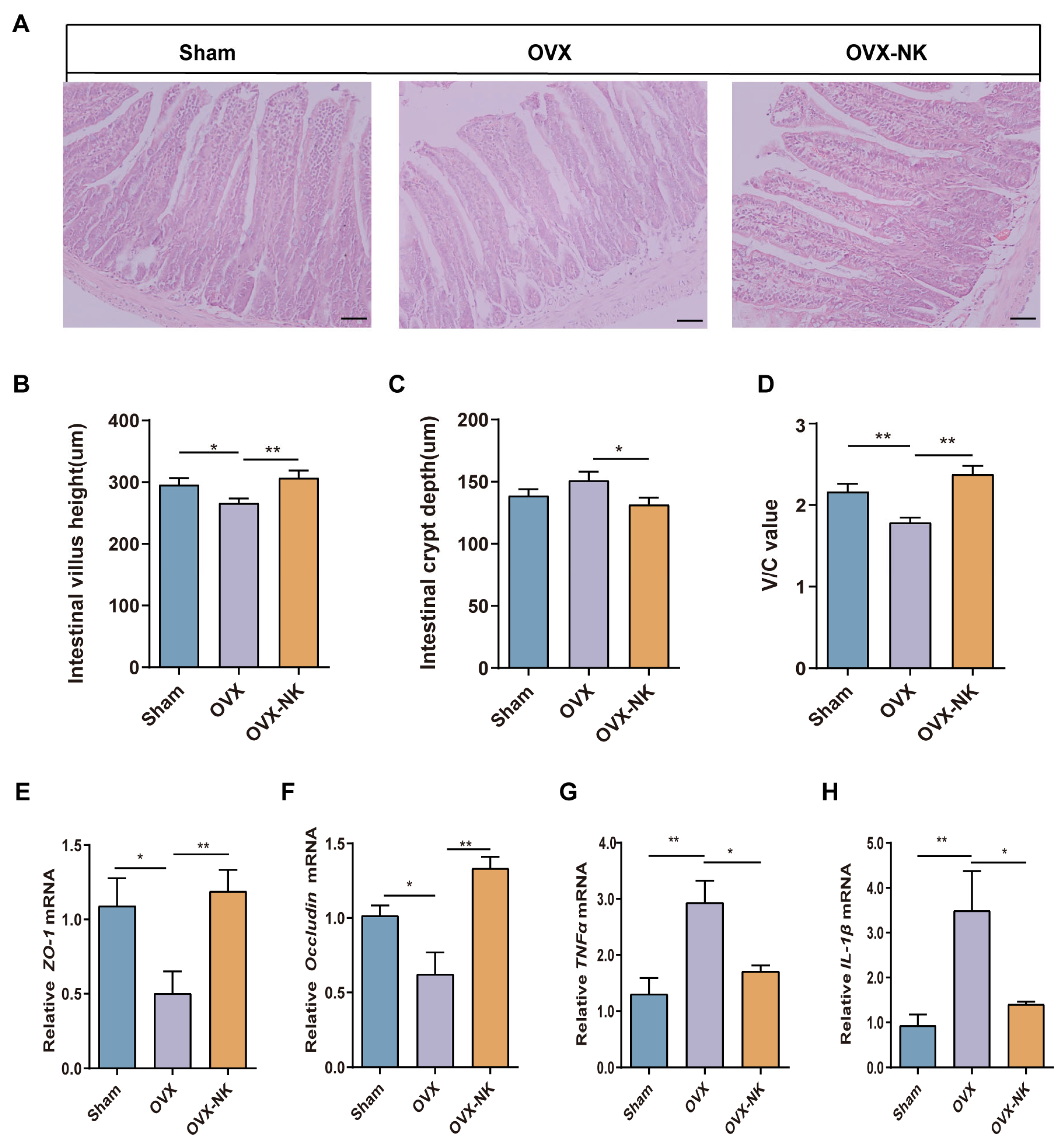
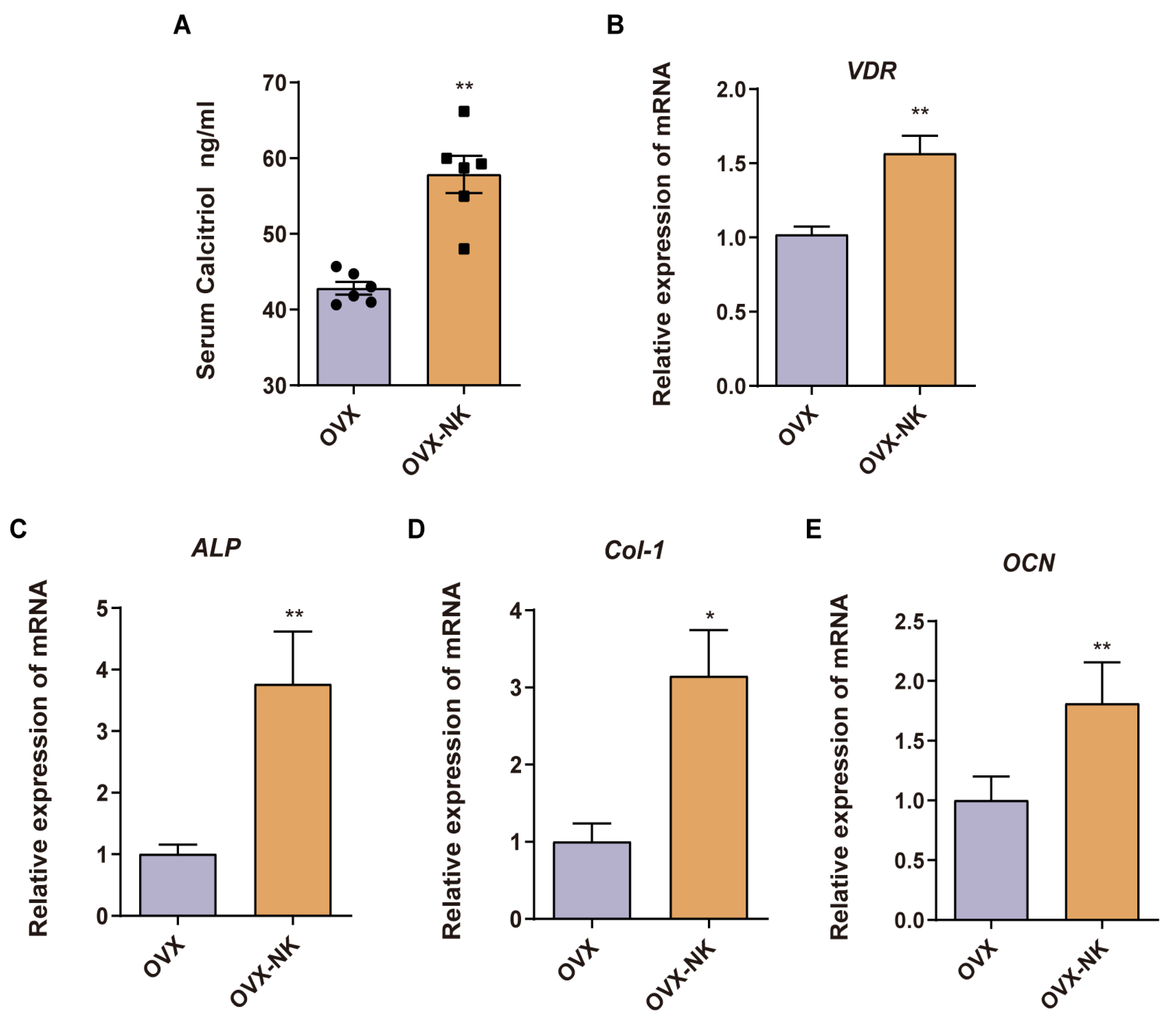
2.5. Nodakenin Ameliorates Intestinal Barrier Impairment in OVX Mice
3. Discussion
4. Materials and Methods
4.1. Animal Experiment and Design
4.2. Micro-CT (μCT) Analysis
4.3. Determination of Biochemical Indicators
4.4. Histomorphometric Analysis
4.5. Quantitative Real-Time PCR
4.6. 16S rRNA Analysis
4.7. Metabolic Analysis of Feces by LC-MS/MS
4.8. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walker, M.D.; Shane, E. Postmenopausal Osteoporosis. N. Engl. J. Med. 2023, 389, 1979–1991. [Google Scholar] [CrossRef]
- Ishikawa, K.; Nagai, T.; Sakamoto, K.; Ohara, K.; Eguro, T.; Ito, H.; Toyoshima, Y.; Kokaze, A.; Toyone, T.; Inagaki, K. High bone turnover elevates the risk of denosumab-induced hypocalcemia in women with postmenopausal osteoporosis. Ther. Clin. Risk Manag. 2016, 12, 1831–1840. [Google Scholar] [CrossRef]
- Zhang, Y.W.; Cao, M.M.; Li, Y.J.; Dai, G.C.; Lu, P.P.; Zhang, M.; Bai, L.Y.; Chen, X.X.; Shi, L.; Zhang, C.; et al. Dietary Protein Intake in Relation to the Risk of Osteoporosis in Middle-Aged and Older Individuals: A Cross-Sectional Study. J. Nutr. Health Aging 2022, 26, 252–258. [Google Scholar] [CrossRef]
- Zhu, Y.; Huang, Z.; Wang, Y.; Xu, W.; Chen, H.; Xu, J.; Luo, S.; Zhang, Y.; Zhao, D.; Hu, J. The efficacy and safety of denosumab in postmenopausal women with osteoporosis previously treated with bisphosphonates: A review. J. Orthop. Transl. 2020, 22, 7–13. [Google Scholar] [CrossRef]
- Arceo-Mendoza, R.M.; Camacho, P.M. Postmenopausal Osteoporosis: Latest Guidelines. Endocrinol. Metab. Clin. N. Am. 2021, 50, 167–178. [Google Scholar] [CrossRef]
- Ke, X.; Walker, A.; Haange, S.-B.; Lagkouvardos, I.; Liu, Y.; Schmitt-Kopplin, P.; von Bergen, M.; Jehmlich, N.; He, X.; Clavel, T.; et al. Synbiotic-driven improvement of metabolic disturbances is associated with changes in the gut microbiome in diet-induced obese mice. Mol. Metab. 2019, 22, 96–109. [Google Scholar] [CrossRef]
- Ejtahed, H.S.; Soroush, A.R.; Angoorani, P.; Larijani, B.; Hasani-Ranjbar, S. Gut Microbiota as a Target in the Pathogenesis of Metabolic Disorders: A New Approach to Novel Therapeutic Agents. Horm. Metab. Res. 2016, 48, 349–358. [Google Scholar] [CrossRef]
- John, G.K.; Mullin, G.E. The Gut Microbiome and Obesity. Curr. Oncol. Rep. 2016, 18, 45. [Google Scholar] [CrossRef]
- Zheng, P.; Li, Z.; Zhou, Z. Gut microbiome in type 1 diabetes: A comprehensive review. Diabetes Metab. Res. Rev. 2018, 34, e3043. [Google Scholar] [CrossRef]
- Cheng, S.; Qi, X.; Ma, M.; Zhang, L.; Cheng, B.; Liang, C.; Liu, L.; Li, P.; Kafle, O.P.; Wen, Y.; et al. Assessing the Relationship Between Gut Microbiota and Bone Mineral Density. Front. Genet. 2020, 11, 6. [Google Scholar] [CrossRef]
- Zhou, T.; Wang, M.; Ma, H.; Li, X.; Heianza, Y.; Qi, L. Dietary Fiber, Genetic Variations of Gut Microbiota-derived Short-chain Fatty Acids, and Bone Health in UK Biobank. J. Clin. Endocrinol. Metab. 2021, 106, 201–210. [Google Scholar] [CrossRef]
- Palacios-González, B.; Ramírez-Salazar, E.G.; Rivera-Paredez, B.; Quiterio, M.; Flores, Y.N.; Macias-Kauffer, L.; Moran-Ramos, S.; Denova-Gutiérrez, E.; Ibarra-González, I.; Vela-Amieva, M.; et al. A Multi-Omic Analysis for Low Bone Mineral Density in Postmenopausal Women Suggests a RELATIONSHIP between Diet, Metabolites, and Microbiota. Microorganisms 2020, 8, 1630. [Google Scholar] [CrossRef]
- Tu, M.-Y.; Han, K.-Y.; Chang, G.R.-L.; Lai, G.-D.; Chang, K.-Y.; Chen, C.-F.; Lai, J.-C.; Lai, C.-Y.; Chen, H.-L.; Chen, C.-M. Kefir Peptides Prevent Estrogen Deficiency-Induced Bone Loss and Modulate the Structure of the Gut Microbiota in Ovariectomized Mice. Nutrients 2020, 12, 3432. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Z.; Hu, Y.; Cui, J.; Zhi, X.; Li, X.; Jiang, H.; Wang, Y.; Gu, Z.; Qiu, Z.; et al. Lactulose Suppresses Osteoclastogenesis and Ameliorates Estrogen Deficiency-Induced Bone Loss in Mice. Aging Dis. 2020, 11, 629–641. [Google Scholar] [CrossRef]
- Li, B.; Liu, M.; Wang, Y.; Gong, S.; Yao, W.; Li, W.; Gao, H.; Wei, M. Puerarin improves the bone micro-environment to inhibit OVX-induced osteoporosis via modulating SCFAs released by the gut microbiota and repairing intestinal mucosal integrity. Biomed. Pharmacother. 2020, 132, 110923. [Google Scholar] [CrossRef]
- Lee, N.Y.; Chung, K.-S.; Jin, J.S.; Lee, Y.-C.; An, H.-J. The Inhibitory Effect of Nodakenin on Mast-Cell-Mediated Allergic Inflammation Via Downregulation of NF-κB and Caspase-1 Activation. J. Cell Biochem. 2017, 118, 3993–4001. [Google Scholar] [CrossRef]
- Lim, J.-Y.; Lee, J.-H.; Yun, D.-H.; Lee, Y.-M.; Kim, D.-K. Inhibitory effects of nodakenin on inflammation and cell death in lipopolysaccharide-induced liver injury mice. Phytomedicine 2021, 81, 153411. [Google Scholar] [CrossRef]
- Li, J.; Wang, L.; Tan, R.; Zhao, S.; Zhong, X.; Wang, L. Nodakenin alleviated obstructive nephropathy through blunting Snail1 induced fibrosis. J. Cell. Mol. Med. 2020, 24, 9752–9763. [Google Scholar] [CrossRef]
- Jin, B.-R.; Lee, M.; An, H.-J. Nodakenin represses obesity and its complications via the inhibition of the VLDLR signalling pathway in vivo and in vitro. Cell Prolif. 2021, 54, e13083. [Google Scholar] [CrossRef]
- Kim, T.W. Nodakenin Induces ROS-Dependent Apoptotic Cell Death and ER Stress in Radioresistant Breast Cancer. Antioxidants 2023, 12, 492. [Google Scholar] [CrossRef]
- Yi, N.; Mi, Y.; Xu, X.; Li, N.; Chen, B.; Yan, K.; Tan, K.; Zhang, B.; Wang, L.; Kuang, G.; et al. Nodakenin attenuates cartilage degradation and inflammatory responses in a mice model of knee osteoarthritis by regulating mitochondrial Drp1/ROS/NLRP3 axis. Int. Immunopharmacol. 2022, 113, 109349. [Google Scholar] [CrossRef]
- Liu, C.; Zhao, M.; Chen, J.; Xu, L.; Wang, K.; Li, G. Nodakenin alleviates ovariectomy-induced osteoporosis by modulating osteoblastogenesis and osteoclastogenesis. Eur. J. Pharmacol. 2023, 960, 176121. [Google Scholar] [CrossRef]
- Zhang, P.; Yang, X.-W. Biotransformation of nodakenin and simultaneous quantification of nodakenin and its aglycone in incubated system of human intestinal bacteria by HPLC method. J. Asian Nat. Prod. Res. 2009, 11, 371–379. [Google Scholar] [CrossRef]
- Feng, W.; Ao, H.; Peng, C.; Yan, D. Gut microbiota, a new frontier to understand traditional Chinese medicines. Pharmacol. Res. 2019, 142, 176–191. [Google Scholar] [CrossRef]
- Kang, D.D.; Li, F.; Kirton, E.; Thomas, A.; Egan, R.; An, H.; Wang, Z. MetaBAT 2: An adaptive binning algorithm for robust and efficient genome reconstruction from metagenome assemblies. PeerJ 2019, 7, e7359. [Google Scholar] [CrossRef]
- Ohlsson, C.; Sjögren, K. Osteomicrobiology: A New Cross-Disciplinary Research Field. Calcif. Tissue Int. 2018, 102, 426–432. [Google Scholar] [CrossRef]
- Reid, I.R. A broader strategy for osteoporosis interventions. Nat. Rev. Endocrinol. 2020, 16, 333–339. [Google Scholar] [CrossRef]
- Behera, J.; Ison, J.; Tyagi, S.C.; Tyagi, N. The role of gut microbiota in bone homeostasis. Bone 2020, 135, 115317. [Google Scholar] [CrossRef]
- Britton, R.A.; Irwin, R.; Quach, D.; Schaefer, L.; Zhang, J.; Lee, T.; Parameswaran, N.; McCabe, L.R.; Probiotic, L. reuteri treatment prevents bone loss in a menopausal ovariectomized mouse model. J. Cell Physiol. 2014, 229, 1822–1830. [Google Scholar] [CrossRef]
- Chen, D.; Zhao, C.-M. The possible existence of a gut-bone axis suggested by studies of genetically manipulated mouse models? Curr. Pharm. Des. 2011, 17, 1552–1555. [Google Scholar] [CrossRef]
- Hao, M.-L.; Wang, G.-Y.; Zuo, X.-Q.; Qu, C.-J.; Yao, B.-C.; Wang, D.-L. Gut microbiota: An overlooked factor that plays a significant role in osteoporosis. J. Int. Med. Res. 2019, 47, 4095–4103. [Google Scholar] [CrossRef]
- Sjögren, K.; Engdahl, C.; Henning, P.; Lerner, U.H.; Tremaroli, V.; Lagerquist, M.K.; Bäckhed, F.; Ohlsson, C. The gut microbiota regulates bone mass in mice. J. Bone Miner. Res. 2012, 27, 1357–1367. [Google Scholar] [CrossRef]
- Yatsonsky Ii, D.; Pan, K.; Shendge, V.B.; Liu, J.; Ebraheim, N.A. Linkage of microbiota and osteoporosis: A mini literature review. World J. Orthop. 2019, 10, 123–127. [Google Scholar] [CrossRef]
- Li, C.; Huang, Q.; Yang, R.; Dai, Y.; Zeng, Y.; Tao, L.; Li, X.; Zeng, J.; Wang, Q. Gut microbiota composition and bone mineral loss-epidemiologic evidence from individuals in Wuhan, China. Osteoporos. Int. 2019, 30, 1003–1013. [Google Scholar] [CrossRef]
- Guo, M.; Liu, H.; Yu, Y.; Zhu, X.; Xie, H.; Wei, C.; Mei, C.; Shi, Y.; Zhou, N.; Qin, K.; et al. Lactobacillus rhamnosus GG ameliorates osteoporosis in ovariectomized rats by regulating the Th17/Treg balance and gut microbiota structure. Gut Microbes 2023, 15, 2190304. [Google Scholar] [CrossRef]
- Weaver, C.M. Diet, gut microbiome, and bone health. Curr. Osteoporos. Rep. 2015, 13, 125–130. [Google Scholar] [CrossRef]
- Jie, L.; Ma, Z.; Gao, Y.; Shi, X.; Yu, L.; Mao, J.; Wang, P. The mechanism of palmatine-mediated intestinal flora and host metabolism intervention in OA-OP comorbidity rats. Front. Med. 2023, 10, 1153360. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, R.; Zhao, R.; Guo, X.; Gou, P.; Bai, H.; Lei, P.; Xue, Y. Intermittent Parathyroid Hormone Alters Gut Microbiota in Ovariectomized Osteoporotic Rats. Orthop. Surg. 2022, 14, 2330–2338. [Google Scholar] [CrossRef]
- Zhou, J.; Cheng, J.; Liu, L.; Luo, J.; Peng, X. Lactobacillus acidophilus (LA) Fermenting Astragalus Polysaccharides (APS) Improves Calcium Absorption and Osteoporosis by Altering Gut Microbiota. Foods 2023, 12, 275. [Google Scholar] [CrossRef]
- He, W.; Xie, Z.; Thøgersen, R.; Rasmussen, M.K.; Zachariassen, L.F.; Jørgensen, N.R.; Nørgaard, J.V.; Andersen, H.J.; Nielsen, D.S.; Hansen, A.K.; et al. Effects of Calcium Source, Inulin, and Lactose on Gut-Bone Associations in an Ovarierectomized Rat Model. Mol. Nutr. Food Res. 2022, 66, e2100883. [Google Scholar] [CrossRef]
- Cao, Y.G.; Bae, S.; Villarreal, J.; Moy, M.; Chun, E.; Michaud, M.; Lang, J.K.; Glickman, J.N.; Lobel, L.; Garrett, W.S. Faecalibaculum rodentium remodels retinoic acid signaling to govern eosinophil-dependent intestinal epithelial homeostasis. Cell Host Microbe 2022, 30, 1295–1310. [Google Scholar] [CrossRef]
- Zagato, E.; Pozzi, C.; Bertocchi, A.; Schioppa, T.; Saccheri, F.; Guglietta, S.; Fosso, B.; Melocchi, L.; Nizzoli, G.; Troisi, J.; et al. Endogenous murine microbiota member Faecalibaculum rodentium and its human homologue protect from intestinal tumour growth. Nat. Microbiol. 2020, 5, 511–524. [Google Scholar] [CrossRef]
- Wu, J.; Guo, W.; Cui, S.; Tang, X.; Zhang, Q.; Lu, W.; Jin, Y.; Zhao, J.; Mao, B.; Chen, W. Broccoli seed extract rich in polysaccharides and glucoraphanin ameliorates DSS-induced colitis via intestinal barrier protection and gut microbiota modulation in mice. J. Sci. Food Agric. 2023, 103, 1749–1760. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond biomarkers and towards mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Tilyard, M.W.; Spears, G.F.; Thomson, J.; Dovey, S. Treatment of postmenopausal osteoporosis with calcitriol or calcium. N. Engl. J. Med. 1992, 326, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Thompson, L.; Wang, S.; Tawfik, O.; Templeton, K.; Tancabelic, J.; Pinson, D.; Anderson, H.C.; Keighley, J.; Garimella, R. Effect of 25-hydroxyvitamin D3 and 1 α,25 dihydroxyvitamin D3 on differentiation and apoptosis of human osteosarcoma cell lines. J. Orthop. Res. 2012, 30, 831–844. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Hu, Z.; Jin, H.; Zheng, G.; Yu, X.; Wu, G.; Liu, H.; Zhu, Z.; Xu, H.; Zhang, C.; et al. Microporous polysaccharide multilayer coated BCP composite scaffolds with immobilised calcitriol promote osteoporotic bone regeneration both in vitro and in vivo. Theranostics 2019, 9, 1125–1143. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Turner, A.G.; Wijenayaka, A.R.; Anderson, P.H.; Morris, H.A.; Atkins, G.J. 1,25-Dihydroxyvitamin D3 and extracellular calcium promote mineral deposition via NPP1 activity in a mature osteoblast cell line MLO-A5. Mol. Cell. Endocrinol. 2015, 412, 140–147. [Google Scholar] [CrossRef]
- Rayalam, S.; Yang, J.-Y.; Della-Fera, M.A.; Baile, C.A. Novel molecular targets for prevention of obesity and osteoporosis. J. Nutr. Biochem. 2011, 22, 1099–1104. [Google Scholar] [CrossRef]
- Mann, S.N.; Pitel, K.S.; Nelson-Holte, M.H.; Iwaniec, U.T.; Turner, R.T.; Sathiaseelan, R.; Kirkland, J.L.; Schneider, A.; Morris, K.T.; Malayannan, S.; et al. 17α-Estradiol prevents ovariectomy-mediated obesity and bone loss. Exp. Gerontol. 2020, 142, 111113. [Google Scholar] [CrossRef]
- Chen, X.; Chen, S.; Ren, Q.; Niu, S.; Yue, L.; Pan, X.; Li, Z.; Zhu, R.; Jia, Z.; Chen, X.; et al. A metabonomics-based renoprotective mechanism analysis of empagliflozin in obese mice. Biochem. Biophys. Res. Commun. 2022, 621, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Zhang, Z.; Dong, X.; Zhao, J.; Peng, Z.; Zhang, L.; Xu, Z.; Xu, L.; Wang, X.; Guo, X. Traumatic acid inhibits ACSL4 associated lipid accumulation in adipocytes to attenuate high-fat diet-induced obesity. FASEB J. 2023, 37, e23278. [Google Scholar] [CrossRef]
- Watson, P.H.; Hanley, D.A. Parathyroid hormone: Regulation of synthesis and secretion. Clin. Invest. Med. 1993, 16, 58–77. [Google Scholar] [PubMed]
- Li, H.; Liu, S.; Miao, C.; Lv, Y.; Hu, Y. Integration of metabolomics and transcriptomics provides insights into enhanced osteogenesis in Ano5Cys360Tyr knock-in mouse model. Front. Endocrinol. 2023, 14, 1117111. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Chen, J.; Zhang, J.; Qin, R.; Wang, R.; Qiu, Y.; Mao, Z.; Goltzman, D.; Miao, D. 1,25-Dihydroxyvitamin D protects against age-related osteoporosis by a novel VDR-Ezh2-p16 signal axis. Aging Cell 2020, 19, e13095. [Google Scholar] [CrossRef] [PubMed]
- Plum, L.A.; DeLuca, H.F. Vitamin D, disease and therapeutic opportunities. Nat. Rev. Drug Discov. 2010, 9, 941–955. [Google Scholar] [CrossRef]
- Fuhrman, B.J.; Feigelson, H.S.; Flores, R.; Gail, M.H.; Xu, X.; Ravel, J.; Goedert, J.J. Associations of the fecal microbiome with urinary estrogens and estrogen metabolites in postmenopausal women. J. Clin. Endocrinol. Metab. 2014, 99, 4632–4640. [Google Scholar] [CrossRef]
- Flores, R.; Shi, J.; Fuhrman, B.; Xu, X.; Veenstra, T.D.; Gail, M.H.; Gajer, P.; Ravel, J.; Goedert, J.J. Fecal microbial determinants of fecal and systemic estrogens and estrogen metabolites: A cross-sectional study. J. Transl. Med. 2012, 10, 253. [Google Scholar] [CrossRef]
- Adeel, S.; Singh, K.; Vydareny, K.H.; Kumari, M.; Shah, E.; Weitzmann, M.N.; Tangpricha, V. Bone loss in surgically ovariectomized premenopausal women is associated with T lymphocyte activation and thymic hypertrophy. J. Investig. Med. 2013, 61, 1178–1183. [Google Scholar] [CrossRef]
- D’Amelio, P.; Grimaldi, A.; Di Bella, S.; Brianza, S.Z.M.; Cristofaro, M.A.; Tamone, C.; Giribaldi, G.; Ulliers, D.; Pescarmona, G.P.; Isaia, G. Estrogen deficiency increases osteoclastogenesis up-regulating T cells activity: A key mechanism in osteoporosis. Bone 2008, 43, 92–100. [Google Scholar] [CrossRef]
- Chen, S.; Zhu, J.; Chen, G.; Zuo, S.; Zhang, J.; Chen, Z.; Wang, X.; Li, J.; Liu, Y.; Wang, P. 1,25-Dihydroxyvitamin D3 preserves intestinal epithelial barrier function from TNF-α induced injury via suppression of NF-kB p65 mediated MLCK-P-MLC signaling pathway. Biochem. Biophys. Res. Commun. 2015, 460, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.; Chen, J.; Wang, Z.; Li, Y.; Zhang, Y.; Li, G. Nodakenin Ameliorates Ovariectomy-Induced Bone Loss by Regulating Gut Microbiota. Molecules 2024, 29, 1240. https://doi.org/10.3390/molecules29061240
Liu C, Chen J, Wang Z, Li Y, Zhang Y, Li G. Nodakenin Ameliorates Ovariectomy-Induced Bone Loss by Regulating Gut Microbiota. Molecules. 2024; 29(6):1240. https://doi.org/10.3390/molecules29061240
Chicago/Turabian StyleLiu, Chunxiao, Jingyue Chen, Zijiao Wang, Yueyao Li, Yuanyuan Zhang, and Guangyu Li. 2024. "Nodakenin Ameliorates Ovariectomy-Induced Bone Loss by Regulating Gut Microbiota" Molecules 29, no. 6: 1240. https://doi.org/10.3390/molecules29061240
APA StyleLiu, C., Chen, J., Wang, Z., Li, Y., Zhang, Y., & Li, G. (2024). Nodakenin Ameliorates Ovariectomy-Induced Bone Loss by Regulating Gut Microbiota. Molecules, 29(6), 1240. https://doi.org/10.3390/molecules29061240