
Illuminating the Tiny World: A Navigation Guide for Proper Raman Studies on Microorganisms

 , , ,
, , ,  , and
, and 
Abstract
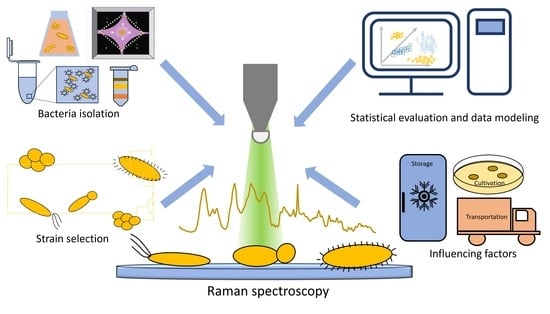
1. Introduction
2. Raman Spectroscopy
3. Strain Selection
4. Principal Factors Influencing Raman Measurements
4.1. Storage and Transport Conditions
4.2. Cultivation Conditions
4.3. Time
4.4. Atmospheric Gases
4.5. Media Composition
4.6. Matrix Simulation
4.7. Influence of the Raman Setup
5. Isolation of Bacteria
5.1. Sample Collection
5.2. Transportation of Samples and Storage Conditions
5.3. Sample Processing: Isolation from Matrix and Isolation Strategies
6. Statistical Evaluation and Data Modeling
6.1. Design of Experiment
6.2. Pre-Treatment Methods
6.3. Pre-Processing Methods
6.4. Modeling
7. Summary and Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Makarewicz, O.; Stein, C.; Pfister, W.; Löffler, B.; Pletz, M. Identification Methods—An Overview. In Modern Techniques for Pathogen Detection; Popp, J., Bauer, M., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2015; pp. 19–53. [Google Scholar]
- Bhunia, A.K.; Singh, A.K.; Parker, K.; Applegate, B.M. Petri-plate, bacteria, and laser optical scattering sensor. Front. Cell. Infect. Microbiol. 2022, 12, 1087074. [Google Scholar] [CrossRef]
- Lee, K.S.; Landry, Z.; Pereira, F.C.; Wagner, M.; Berry, D.; Huang, W.E.; Taylor, G.T.; Kneipp, J.; Popp, J.; Zhang, M.; et al. Raman microspectroscopy for microbiology. Nat. Rev. Methods Primers 2021, 1, 80. [Google Scholar] [CrossRef]
- Stöckel, S.; Kirchhoff, J.; Neugebauer, U.; Rösch, P.; Popp, J. The application of Raman spectroscopy for the detection and identification of microorganisms. J. Raman Spectrosc. 2016, 47, 89–109. [Google Scholar] [CrossRef]
- Senger, R.S.; Scherr, D. Resolving complex phenotypes with Raman spectroscopy and chemometrics. Curr. Opin. Biotechnol. 2020, 66, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Pahlow, S.; Meisel, S.; Cialla-May, D.; Weber, K.; Rösch, P.; Popp, J. Isolation and identification of bacteria by means of Raman spectroscopy. Adv. Drug Del. Rev. 2015, 89, 105–120. [Google Scholar] [CrossRef] [PubMed]
- Petersen, M.; Yu, Z.L.; Lu, X.N. Application of Raman Spectroscopic Methods in Food Safety: A Review. Biosensors 2021, 11, 187. [Google Scholar] [CrossRef] [PubMed]
- Pistiki, A.; Salbreiter, M.; Sultan, S.; Rösch, P.; Popp, J. Application of Raman spectroscopy in the hospital environment. Transl. Biophotonics 2022, 4, e202200011. [Google Scholar] [CrossRef]
- Wang, L.; Liu, W.; Tang, J.W.; Wang, J.J.; Liu, Q.H.; Wen, P.B.; Wang, M.M.; Pan, Y.C.; Gu, B.; Zhang, X. Applications of Raman Spectroscopy in Bacterial Infections: Principles, Advantages, and Shortcomings. Front. Microbiol. 2021, 12, 683580. [Google Scholar] [CrossRef]
- Cialla-May, D.; Krafft, C.; Rösch, P.; Deckert-Gaudig, T.; Frosch, T.; Jahn, I.J.; Pahlow, S.; Stiebing, C.; Meyer-Zedler, T.; Bocklitz, T.; et al. Raman Spectroscopy and Imaging in Bioanalytics. Anal. Chem. 2022, 94, 86–119. [Google Scholar] [CrossRef]
- Cialla-May, D.; Rösch, P.; Popp, J. Raman-Spektroskopie in der Infektionsforschung. BIOspektrum 2022, 28, 597–599. [Google Scholar] [CrossRef]
- Hong, J.K.; Kim, S.B.; Lyou, E.S.; Lee, T.K. Microbial phenomics linking the phenotype to function: The potential of Raman spectroscopy. J. Microbiol. 2021, 59, 249–258. [Google Scholar] [CrossRef]
- Rebrošová, K.; Samek, O.; Kizovsky, M.; Bernatova, S.; Hola, V.; Ruzicka, F. Raman Spectroscopy-A Novel Method for Identification and Characterization of Microbes on a Single-Cell Level in Clinical Settings. Front. Cell. Infect. Microbiol. 2022, 12, 866463. [Google Scholar] [CrossRef]
- Lorenz, B.; Wichmann, C.; Stöckel, S.; Rösch, P.; Popp, J. Cultivation-free Raman Spectroscopic Investigations on Bacteria. Trends Microbiol. 2017, 25, 413–424. [Google Scholar] [CrossRef]
- Pezzotti, G. Raman spectroscopy in cell biology and microbiology. J. Raman Spectrosc. 2021, 52, 2348–2443. [Google Scholar] [CrossRef]
- Rodriguez, L.; Zhang, Z.Y.; Wang, D.H. Recent advances of Raman spectroscopy for the analysis of bacteria. Anal. Sci. Adv. 2023, 4, 81–95. [Google Scholar] [CrossRef]
- Wang, P.C.; Sun, H.; Yang, W.; Fang, Y.M. Optical Methods for Label-Free Detection of Bacteria. Biosensors 2022, 12, 1171. [Google Scholar] [CrossRef]
- Zhan, L.J.; Liu, H.M.; Hou, Z.H.; Gao, Y.W.; Chu, B.Y.; Huang, G.M. Recent advances in single bacterium metabolic analysis techniques. TrAC—Trends Anal. Chem. 2023, 163, 117076. [Google Scholar] [CrossRef]
- Ramzan, M.; Raza, A.; Nisa, Z.U.; Musharraf, S.G. Recent studies on advance spectroscopic techniques for the identification of microorganisms: A review. Arab. J. Chem. 2023, 16, 104521. [Google Scholar] [CrossRef]
- Salbreiter, M.; Pistiki, A.; Cialla-May, D.; Rösch, P.; Popp, J. Raman Spectroscopy for Infection Diagnosis. In Raman Spectroscopy in Human Health and Biomedicine; Sato, H., Popp, J., Wood, B.R., Ozaki, Y., Eds.; World Scientific: Singapore, 2023; pp. 337–410. [Google Scholar]
- Azemtsop Matanfack, G.; Rüger, J.; Stiebing, C.; Schmitt, M.; Popp, J. Imaging the Invisible-Bioorthogonal Raman Probes for Imaging of Cells and Tissues. J. Biophotonics 2020, 13, e202000129. [Google Scholar] [CrossRef]
- Daniel, F.; Kesterson, D.; Lei, K.; Hord, C.; Patel, A.; Kaffenes, A.; Congivaram, H.; Prakash, S. Application of Microfluidics for Bacterial Identification. Pharmaceuticals 2022, 15, 1531. [Google Scholar] [CrossRef]
- Kumar, B.N.V.; Kampe, B.; Rösch, P.; Popp, J. Characterization of carotenoids in soil bacteria and investigation of their photostability towards UVA radiation via resonance Raman spectroscopy. Analyst 2015, 140, 4584–4593. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.N.V.; Kampe, B.; Rösch, P.; Popp, J. Classification and identification of pigmented cocci bacteria relevant to the soil environment via Raman spectroscopy. Environ. Sci. Pollut. Res. 2015, 22, 19317–19325. [Google Scholar] [CrossRef] [PubMed]
- Kniggendorf, A.-K.; Gaul, T.W.; Meinhardt-Wollweber, M. Effects of Ethanol, Formaldehyde, and Gentle Heat Fixation in Confocal Resonance Raman Microscopy of Purple NonSulfur Bacteria. Microsc. Res. Tech. 2011, 74, 177–183. [Google Scholar] [CrossRef]
- Mukherjee, R.; Verma, T.; Nandi, D.; Umapathy, S. Identification of a resonance Raman marker for cytochrome to monitor stress responses in Escherichia coli. Anal. Bioanal. Chem. 2020, 412, 5379–5388. [Google Scholar] [CrossRef]
- Liu, C.; Guo, Z.; Zhang, H.; Li, J.; Zhu, C.; Zhu, G. Single-cell Raman spectra reveals the cytochrome c-mediated electron transfer in nanoscale zero-valent iron coupled denitrification process. Chem. Eng. J. 2023, 454, 140241. [Google Scholar] [CrossRef]
- Samek, O.; Bernatova, S.; Dohnal, F. The potential of SERS as an AST methodology in clinical settings. Nanophotonics 2021, 10, 2537–2561. [Google Scholar] [CrossRef]
- Beeram, R.; Vepa, K.R.; Soma, V.R. Recent Trends in SERS-Based Plasmonic Sensors for Disease Diagnostics, Biomolecules Detection, and Machine Learning Techniques. Biosensors 2023, 13, 328. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.L.; Ma, W.R.; Wang, X.; Li, S.B. Recent Progress of Surface-Enhanced Raman Spectroscopy for Bacteria Detection. Biosensors 2023, 13, 350. [Google Scholar] [CrossRef]
- Usman, M.; Tang, J.W.; Li, F.; Lai, J.X.; Liu, Q.H.; Liu, W.; Wang, L. Recent advances in surface enhanced Raman spectroscopy for bacterial pathogen identifications. J. Adv. Res. 2023, 51, 91–107. [Google Scholar] [CrossRef]
- Zhu, A.F.; Ali, S.; Jiao, T.H.; Wang, Z.; Ouyang, Q.; Chen, Q.S. Advances in surface-enhanced Raman spectroscopy technology for detection of foodborne pathogens. Compr. Rev. Food Sci. Food Saf. 2023, 22, 1466–1494. [Google Scholar] [CrossRef]
- Liu, H.Q.; Gao, X.; Xu, C.; Liu, D.B. SERS Tags for Biomedical Detection and Bioimaging. Theragnostigs 2022, 12, 1870–1903. [Google Scholar] [CrossRef]
- Allen, D.M.; Einarsson, G.G.; Tunney, M.M.; Bell, S.E.J. Characterization of Bacteria Using Surface-Enhanced Raman Spectroscopy (SERS): Influence of Microbiological Factors on the SERS Spectra. Anal. Chem. 2022, 94, 9327–9335. [Google Scholar] [CrossRef]
- Berry, M.E.; Kearns, H.; Graham, D.; Faulds, K. Surface enhanced Raman scattering for the multiplexed detection of pathogenic microorganisms: Towards point-of-use applications. Analyst 2021, 146, 6084–6101. [Google Scholar] [CrossRef]
- Cui, L.; Zhang, D.D.; Yang, K.; Zhang, X.; Zhu, Y.G. Perspective on Surface-Enhanced Raman Spectroscopic Investigation of Microbial World. Anal. Chem. 2019, 91, 15345–15354. [Google Scholar] [CrossRef]
- Al-Shaebi, Z.; Akdeniz, M.; Ahmed, A.O.; Altunbek, M.; Aydin, O. Breakthrough Solution for Antimicrobial Resistance Detection: Surface-Enhanced Raman Spectroscopy-based on Artificial Intelligence. Adv. Mat. Interf. 2023, 2300664. [Google Scholar] [CrossRef]
- Panikar, S.S.; Cialla-May, D.; De la Rosa, E.; Salas, P.; Popp, J. Towards translation of surface-enhanced Raman spectroscopy (SERS) to clinical practice: Progress and trends. TrAC—Trends Anal. Chem. 2021, 134, 116122. [Google Scholar] [CrossRef]
- Hussain, M.; Zou, J.; Zhang, H.; Zhang, R.; Chen, Z.; Tang, Y.J. Recent Progress in Spectroscopic Methods for the Detection of Foodborne Pathogenic Bacteria. Biosensors 2022, 12, 869. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Natarajan, B.; Kannan, P. Nanostructured biosensing platforms for the detection of food- and water-borne pathogenic Escherichia coli. Anal. Bioanal. Chem. 2023, 415, 3111–3129. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Lu, S.S.; Chen, B.; Hu, F.X.; Li, C.M.; Guo, C.X. Machine learning-assisted optical nano-sensor arrays in microorganism analysis. TrAC—Trends Anal. Chem. 2023, 159, 116945. [Google Scholar] [CrossRef]
- Wang, H.Q.; Zhu, L.S.; Liang, Z.D.; Pang, Y.J.; Wang, X.H.; Zhou, L. Recent Advances in Nanomaterials-Based Sensing Platforms for the Determination of Multiple Bacterial Species: A Minireview. Anal. Lett. 2024, 57, 920–939. [Google Scholar] [CrossRef]
- Doolittle, W.F.; Zhaxybayeva, O. On the origin of prokaryotic species. Genome Res. 2009, 19, 744–756. [Google Scholar] [CrossRef] [PubMed]
- OECD. Safety Assessment of Transgenic Organisms in the Environment; OECD: Paris, France, 2016; Volume 5. [Google Scholar] [CrossRef]
- Nakar, A.; Wagenhaus, A.; Rösch, P.; Popp, J. Raman spectroscopy for the differentiation of Enterobacteriaceae: A comparison of two methods. Analyst 2022, 147, 3938–3946. [Google Scholar] [CrossRef] [PubMed]
- Willemse-Erix, D.F.; Scholtes-Timmerman, M.J.; Jachtenberg, J.-W.; van Leeuwen, W.B.; Horst-Kreft, D.; Bakker Schut, T.C.; Deurenberg, R.H.; Puppels, G.J.; van Belkum, A.; Vos, M.C. Optical Fingerprinting in Bacterial Epidemiology: Raman Spectroscopy as a Real-Time Typing Method. J. Clin. Microbiol. 2009, 47, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Chen, L.; Ma, X.; Ma, L.; Chou Keng, C.; Cao, Y.; Khan Izhar, U.H.; Gölz, G.; Lu, X.; Master Emma, R. Arcobacter Identification and Species Determination Using Raman Spectroscopy Combined with Neural Networks. Appl. Environ. Microbiol. 2020, 86, e00924-20. [Google Scholar] [CrossRef]
- Rebrošová, K.; Siler, M.; Samek, O.; Ruzicka, F.; Bernatova, S.; Hola, V.; Ježek, J.; Zemanek, P.; Sokolova, J.; Petras, P. Rapid identification of staphylococci by Raman spectroscopy. Sci. Rep. 2017, 7, 14846. [Google Scholar] [CrossRef]
- Ayala, O.D.; Wakeman, C.A.; Pence, I.J.; Gaddy, J.A.; Slaughter, J.C.; Skaar, E.P.; Mahadevan-Jansen, A. Drug-Resistant Staphylococcus aureus Strains Reveal Distinct Biochemical Features with Raman Microspectroscopy. ACS Infect. Dis. 2018, 4, 1197–1210. [Google Scholar] [CrossRef] [PubMed]
- Lister, A.P.; Highmore, C.J.; Hanrahan, N.; Read, J.; Munro, A.P.S.; Tan, S.; Allan, R.N.; Faust, S.N.; Webb, J.S.; Mahajan, S. Multi-Excitation Raman Spectroscopy for Label-Free, Strain-Level Characterization of Bacterial Pathogens in Artificial Sputum Media. Anal. Chem. 2022, 94, 669–677. [Google Scholar] [CrossRef]
- Pistiki, A.; Monecke, S.; Shen, H.; Ryabchykov, O.; Bocklitz, T.; Rösch, P.; Ehricht, R.; Popp, J. Comparison of Different Label-Free Raman Spectroscopy Approaches for the Discrimination of Isogenic MRSA and MSSA Clinical Isolates. Microbiol. Spectrum 2022, 10, e0076322. [Google Scholar] [CrossRef]
- Novikov, A.; Sayfutdinova, A.; Botchkova, E.; Kopitsyn, D.; Fakhrullin, R. Antibiotic Susceptibility Testing with Raman Biosensing. Anibiotics 2022, 11, 1812. [Google Scholar] [CrossRef]
- Shen, H.; Rösch, P.; Pletz, M.; Popp, J. In Vitro Fiber-probe based Raman Spectroscopic identification on pathogenic biofilms. Anal. Chem. 2022, 94, 5375–5381. [Google Scholar] [CrossRef]
- Azemtsop Matanfack, G.; Pistiki, A.; Rösch, P.; Popp, J. Raman Stable Isotope Labeling of Single Bacteria in Visible and Deep UV-Ranges. Life 2021, 11, 1003. [Google Scholar] [CrossRef]
- Maquelin, K.; Choo-Smith, L.P.; van Vreeswijk, T.; Endtz, H.P.; Smith, B.; Bennett, R.; Bruining, H.A.; Puppels, G.J. Raman Spectroscopic Method for Identification of Clinically Relevant Microorganisms Growing on Solid Culture Medium. Anal. Chem. 2000, 72, 12–19. [Google Scholar] [CrossRef]
- Ghebremedhin, M.; Heitkamp, R.; Yesupriya, S.; Clay, B.; Crane Nicole, J.; Munson, E. Accurate and Rapid Differentiation of Acinetobacter baumannii Strains by Raman Spectroscopy: A Comparative Study. J. Clin. Microbiol. 2017, 55, 2480–2490. [Google Scholar] [CrossRef]
- Verma, T.; Annappa, H.; Singh, S.; Umapathy, S.; Nandi, D. Profiling antibiotic resistance in Escherichia coli strains displaying differential antibiotic susceptibilities using Raman spectroscopy. J. Biophotonics 2021, 14, e202000231. [Google Scholar] [CrossRef]
- Rusciano, G.; Capriglione, P.; Pesce, G.; Abete, P.; Carnovale, V.; Sasso, A. Raman spectroscopy as a new tool for early detection of bacteria in patients with cystic fibrosis. Laser Phys. Lett. 2013, 10, 075603. [Google Scholar] [CrossRef]
- Lorenz, B.; Rösch, P.; Popp, J. Isolation matters-processing blood for Raman microspectroscopic identification of bacteria. Anal. Bioanal. Chem. 2019, 11, 5445–5454. [Google Scholar] [CrossRef]
- Dekter, H.E.; Orelio, C.C.; Morsink, M.C.; Tektas, S.; Vis, B.; te Witt, R.; van Leeuwen, W.B. Antimicrobial susceptibility testing of Gram-positive and -negative bacterial isolates directly from spiked blood culture media with Raman spectroscopy. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Kloß, S.; Kampe, B.; Sachse, S.; Rösch, P.; Straube, E.; Pfister, W.; Kiehntopf, M.; Popp, J. Culture Independent Raman Spectroscopic Identification of Urinary Tract Infection Pathogens: A Proof of Principle Study. Anal. Chem. 2013, 85, 9610–9616. [Google Scholar] [CrossRef] [PubMed]
- Walter, A.; Reinicke, M.; Bocklitz, T.; Schumacher, W.; Rösch, P.; Kothe, E.; Popp, J. Raman spectroscopic detection of physiology changes in plasmid-bearing Escherichia coli with and without antibiotic treatment. Anal. Bioanal. Chem. 2011, 400, 2763–2773. [Google Scholar] [CrossRef] [PubMed]
- Germond, A.; Ichimura, T.; Horinouchi, T.; Fujita, H.; Furusawa, C.; Watanabe, T.M. Raman spectral signature reflects transcriptomic features of antibiotic resistance in Escherichia coli. Commun. Biol. 2018, 1, 85. [Google Scholar] [CrossRef] [PubMed]
- Saikia, D.; Jadhav, P.; Hole, A.R.; Krishna, C.M.; Singh, S.P. Unraveling the Secrets of Colistin Resistance with Label-Free Raman Spectroscopy. Biosensors 2022, 12, 747. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Xu, F.; Zhu, L.; Li, H.; Sun, Q.; Yan, A.; Ren, B.; Zhu, Y.G.; Cui, L. An Isotope-Labeled Single-Cell Raman Spectroscopy Approach for Tracking the Physiological Evolution Trajectory of Bacteria toward Antibiotic Resistance. Angew. Chem. Int. Ed. 2023, 62, e202217412. [Google Scholar] [CrossRef] [PubMed]
- Maquelin, K.; Kirschner, C.; Choo-Smith, L.-P.; Ngo-Thi, N.; Van Vreeswijk, T.; Stammler, M.; Endtz, H.; Bruining, H.; Naumann, D.; Puppels, G. Prospective Study of the Performance of Vibrational Spectroscopies for Rapid Identification of Bacterial and Fungal Pathogens Recovered from Blood Cultures. J. Clin. Microbiol. 2003, 41, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Read, D.S.; Whiteley, A.S. Chemical fixation methods for Raman spectroscopy-based analysis of bacteria. J. Microbiol. Meth. 2015, 109, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Wichmann, C.; Chhallani, M.; Bocklitz, T.; Rösch, P.; Popp, J. Simulation of Transportation and Storage and Their Influence on Raman Spectra to Bacteria. Anal. Chem. 2019, 91, 13688–13694. [Google Scholar] [CrossRef]
- Wang, W.; Rahman, A.; Kang, S.; Vikesland, P.J. Investigation of the Influence of Stress on Label-Free Bacterial Surface-Enhanced Raman Spectra. Anal. Chem. 2023, 95, 3675–3683. [Google Scholar] [CrossRef]
- Mlynáriková, K.; Samek, O.; Bernatová, S.; Růžička, F.; Ježek, J.; Hároniková, A.; Šiler, M.; Zemánek, P.; Holá, V. Influence of Culture Media on Microbial Fingerprints Using Raman Spectroscopy. Sensors 2015, 15, 29635–29647. [Google Scholar] [CrossRef]
- Harz, M.; Rösch, P.; Peschke, K.D.; Ronneberger, O.; Burkhardt, H.; Popp, J. Micro-Raman spectroscopic identification of bacterial cells of the genus Staphylococcus and dependence on their cultivation conditions. Analyst 2005, 130, 1543–1550. [Google Scholar] [CrossRef]
- Schuster, K.C.; Urlaub, E.; Gapes, J.R. Single-cell analysis of bacteria by Raman microscopy: Spectral information on the chemical composition of cells and on the heterogeneity in a culture. J. Microbiol. Meth. 2000, 42, 29–38. [Google Scholar] [CrossRef]
- Kniggendorf, A.K.; Nogueira, R.; Nasiri Bahmanabad, S.; Pommerening-Röser, A.; Roth, B.W. Small Sample Stress: Probing Oxygen-Deprived Ammonia-Oxidizing Bacteria with Raman Spectroscopy In Vivo. Microorganisms 2020, 8, 432. [Google Scholar] [CrossRef] [PubMed]
- Wichmann, C.; Bocklitz, T.; Rösch, P.; Popp, J. Bacterial phenotype dependency from CO2 measured by Raman spectroscopy. Spectrochim. Acta A 2021, 248, 119170. [Google Scholar] [CrossRef]
- Keiner, R.; Frosch, T.; Hanf, S.; Rusznyak, A.; Akob, D.M.; Küsel, K.; Popp, J. Raman Spectroscopy-An Innovative and Versatile Tool to Follow the Respirational Activity and Carbonate Biomineralization of Important Cave Bacteria. Anal. Chem. 2013, 85, 8708–8714. [Google Scholar] [CrossRef]
- Li, M.; Canniffe, D.P.; Jackson, P.J.; Davison, P.A.; FitzGerald, S.; Dickman, M.J.; Burgess, J.G.; Hunter, C.N.; Huang, W.E. Rapid resonance Raman microspectroscopy to probe carbon dioxide fixation by single cells in microbial communities. ISME J. 2012, 6, 875–885. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.; Gong, Y.; Xu, T.; Davison, P.A.; MacGregor-Chatwin, C.; Hunter, C.N.; Xu, L.; Meng, Y.; Ji, Y.; Ma, B.; et al. Revealing CO2-Fixing SAR11 Bacteria in the Ocean by Raman-Based Single-Cell Metabolic Profiling and Genomics. BioDes. Res. 2022, 2022, 9782712. [Google Scholar] [CrossRef]
- Hanson, C.; Bishop, M.M.; Barney, J.T.; Vargis, E. Effect of growth media and phase on Raman spectra and discrimination of mycobacteria. J. Biophotonics 2019, 12, e201900150. [Google Scholar] [CrossRef]
- Kunapareddy, N.; Grun, J.; Lunsford, R.; Nikitin, S.; Wang, Z.; Gillis, D. Multiwavelength Resonance Raman Characterization of the Effect of Growth Phase and Culture Medium on Bacteria. Appl. Spectrosc. 2015, 69, 966–971. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Matange, N.; Umapathy, S.; Visweswariah, S.S. Linking carbon metabolism to carotenoid production in mycobacteria using Raman spectroscopy. FEMS Microbiol. Lett. 2015, 362, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Huayhongthong, S.; Khuntayaporn, P.; Thirapanmethee, K.; Wanapaisan, P.; Chomnawang, M.T. Raman spectroscopic analysis of food-borne microorganisms. LWT 2019, 114, 108419. [Google Scholar] [CrossRef]
- Daniels, J.K.; Caldwell, T.P.; Christensen, K.A.; Chumanov, G. Monitoring the Kinetics of Bacillus subtilis Endospore Germination via Surface-Enhanced Raman Scattering Spectroscopy. Anal. Chem. 2006, 78, 1724–1729. [Google Scholar] [CrossRef]
- Madigan, M.T.; Bender, K.S.; Buckley, D.H.; Sattley, W.M.; Stahl, D.A. Brock Biology of Microorganisms; Pearson Education Limited: London, UK, 2020. [Google Scholar]
- Yamamoto, T.; Taylor, J.N.; Koseki, S.; Koyama, K. Prediction of growth/no growth status of previously unseen bacterial strain using Raman spectroscopy and machine learning. LWT 2023, 174, 114449. [Google Scholar] [CrossRef]
- Hutsebaut, D.; Maquelin, K.; De Vos, P.; Vandenabeele, P.; Moens, L.; Puppels, G. Effect of Culture Conditions on the Achievable Taxonomic Resolution of Raman Spectroscopy Disclosed by Three Bacillus Species. Anal. Chem. 2004, 76, 6274–6281. [Google Scholar] [CrossRef]
- van der Mei, H.; Naumann, D.; Busscher, H. Grouping of Streptococcus mitis strains grown on different growth media by FT-IR. Infrared Phys. Technol. 1996, 37, 561–564. [Google Scholar] [CrossRef]
- Stöckel, S.; Stanca, A.S.; Helbig, J.; Rösch, P.; Popp, J. Raman spectroscopic monitoring of the growth of pigmented and non-pigmented Mycobacteria. Anal. Bioanal. Chem. 2015, 407, 8919–8923. [Google Scholar] [CrossRef]
- Wang, H.; Hollywood, K.; Jarvis, R.M.; Lloyd, J.R.; Goodacre, R. Phenotypic Characterization of Shewanella oneidensis MR-1 under Aerobic and Anaerobic Growth Conditions by Using Fourier Transform Infrared Spectroscopy and High-Performance Liquid Chromatography Analyses. Appl. Environ. Microbiol. 2010, 76, 6266–6276. [Google Scholar] [CrossRef]
- Silge, A.; Abdou, E.; Schneider, K.; Meisel, S.; Bocklitz, T.; Lu-Walther, H.W.; Heintzmann, R.; Rösch, P.; Popp, J. Shedding light on host niches: Label-free in situ detection of Mycobacterium gordonae via carotenoids in macrophages by Raman microspectroscopy. Cell. Microbiol. 2015, 17, 832–842. [Google Scholar] [CrossRef]
- Azzam, Z.S.; Sharabi, K.; Guetta, J.; Bank, E.M.; Gruenbaum, Y. The physiological and molecular effects of elevated CO2 levels. Cell Cycle 2010, 9, 1528–1532. [Google Scholar] [CrossRef]
- Tanniche, I.; Collakova, E.; Denbow, C.; Senger, R.S. Characterizing metabolic stress-induced phenotypes of Synechocystis PCC6803 with Raman spectroscopy. PeerJ 2020, 8, e8535. [Google Scholar] [CrossRef]
- Ullah, U.; Tahir, Z.; Qazi, O.; Mirza, S.; Cheema, M.I. Raman spectroscopy and machine learning-based optical probe for tuberculosis diagnosis via sputum. Tuberculosis 2022, 136, 102251. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Gopinathan, R.; Chandra, G.K.; Umapathy, S.; Saini, D.K. Rapid detection of bacterial infection and viability assessment with high specificity and sensitivity using Raman microspectroscopy. Anal. Bioanal. Chem. 2020, 412, 2505–2516. [Google Scholar] [CrossRef]
- Kloß, S.; Rösch, P.; Pfister, W.; Kiehntopf, M.; Popp, J. Towards Culture-Free Raman Spectroscopic Identification of Pathogens in Ascitic Fluid. Anal. Chem. 2015, 87, 937–943. [Google Scholar] [CrossRef] [PubMed]
- Farrugia, M.A.; Le Corvec, M.; Renou, C.; Nousbaum, J.B.; Ouizeman, D.J.; Sire, O.; Loreal, O.; Tariel, H.; Bernard, J.; Piche, T.; et al. Mid-Infrared Spectroscopy as a New Tool for Ruling Out Spontaneous Bacterial Peritonitis: A Proof-of-Concept Study. Biomedicines 2023, 11, 838. [Google Scholar] [CrossRef]
- Meisel, S.; Stöckel, S.; Rösch, P.; Popp, J. Identification of meat associated pathogens via Raman microspectroscopy. Food Microbiol. 2014, 38, 36–43. [Google Scholar] [CrossRef]
- Alexandrakis, D.; Downey, G.; Scannell, A.G.M. Detection and Identification of Bacteria in an Isolated System with Near-Infrared Spectroscopy and Multivariate Analysis. J. Agric. Food Chem. 2008, 56, 3431–3437. [Google Scholar] [CrossRef]
- Amamcharla, J.K.; Panigrahi, S.; Logue, C.M.; Marchello, M.; Sherwood, J.S. Application of vapour-phase Fourier transform infrared spectroscopy (FTIR) and statistical feature selection methods for identifying Salmonella enterica typhimurium contamination in beef. Biosyst. Eng. 2010, 107, 1–9. [Google Scholar] [CrossRef]
- Ammor, M.S.; Argyri, A.; Nychas, G.J. Rapid monitoring of the spoilage of minced beef stored under conventionally and active packaging conditions using Fourier transform infrared spectroscopy in tandem with chemometrics. Meat Sci. 2009, 81, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Argyri, A.; Panagou, E.; Tarantilis, P.; Polysiou, M.; Nychas, G.-J. Rapid Qualitative and Quantitative Detection of Beef Fillets Spoilage Based on Fourier Transform Infrared Spectroscopy Data and Artificial Neural Networks. Sens. Actuators B Chem. 2010, 145, 146–154. [Google Scholar] [CrossRef]
- Davis, R.; Burgula, Y.; Deering, A.; Irudayaraj, J.; Reuhs, B.L.; Mauer, L.J. Detection and differentiation of live and heat-treated Salmonella enterica serovars inoculated onto chicken breast using Fourier transform infrared (FT-IR) spectroscopy. J. Appl. Microbiol. 2010, 109, 2019–2031. [Google Scholar] [CrossRef]
- Ellis, D.I.; Broadhurst, D.; Kell, D.B.; Rowland, J.J.; Goodacre, R. Rapid and Quantitative Detection of the Microbial Spoilage of Meat by Fourier Transform Infrared Spectroscopy and Machine Learning. Appl. Environ. Microbiol. 2002, 68, 2822–2828. [Google Scholar] [CrossRef]
- Gaus, K.; Rösch, P.; Petry, R.; Peschke, K.D.; Ronneberger, O.; Burkhardt, H.; Baumann, K.; Popp, J. Classification of Lactic Acid Bacteria with UV-Resonance Raman Spectroscopy. Biopolymers 2006, 82, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Maquelin, K.; Kirschner, C.; Choo-Smith, L.P.; van den Braak, N.; Endtz, H.P.; Naumann, D.; Puppels, G.J. Identification of medically relevant microorganisms by vibrational spectroscopy. J. Microbiol. Meth. 2002, 51, 255–271. [Google Scholar] [CrossRef]
- Wichmann, C.; Rösch, P.; Popp, J. Isolation of bacteria from artificial bronchoalveolar lavage fluid using density gradient centrifugation and their accessibility by Raman spectroscopy. Anal. Bioanal. Chem. 2021, 413, 5193–5200. [Google Scholar] [CrossRef]
- Kusić, D.; Kampe, B.; Rösch, P.; Popp, J. Identification of water pathogens by Raman microspectroscopy. Water Res. 2014, 48, 179–189. [Google Scholar] [CrossRef]
- Arslan, A.H.; Ciloglu, F.U.; Yilmaz, U.; Simsek, E.; Aydin, O. Discrimination of waterborne pathogens, Cryptosporidium parvum oocysts and bacteria using surface-enhanced Raman spectroscopy coupled with principal component analysis and hierarchical clustering. Spectrochim. Acta A 2022, 267, 120475. [Google Scholar] [CrossRef]
- Dhankhar, D.; Nagpal, A.; Li, R.; Chen, J.; Cesario, T.C.; Rentzepis, P.M. Resonance Raman Spectra for the In Situ Identification of Bacteria Strains and Their Inactivation Mechanism. Appl. Spectrosc. 2021, 75, 1146–1154. [Google Scholar] [CrossRef] [PubMed]
- Aubrechtová Dragounová, K.; Ryabchykov, O.; Steinbach, D.; Recla, V.; Lindig, N.; González Vázquez, M.J.; Foller, S.; Bauer, M.; Bocklitz, T.W.; Popp, J.; et al. Identification of bacteria in mixed infection from urinary tract of patient’s samples using Raman analysis of dried droplets. Analyst 2023, 148, 3806–3816. [Google Scholar] [CrossRef] [PubMed]
- Tien, N.; Lin, T.H.; Hung, Z.C.; Lin, H.S.; Wang, I.K.; Chen, H.C.; Chang, C.T. Diagnosis of Bacterial Pathogens in the Urine of Urinary-Tract-Infection Patients Using Surface-Enhanced Raman Spectroscopy. Molecules 2018, 23, 3374. [Google Scholar] [CrossRef] [PubMed]
- Hadjigeorgiou, K.; Kastanos, E.; Kyriakides, A.; Pitris, C. Raman spectroscopy for UTI Diagnosis and Antibiogram. In Proceedings of the 2009 9th International Conference on Information Technology and Applications in Biomedicine, Larnaka, Cyprus, 4–7 November 2009; pp. 1–4. [Google Scholar]
- Jarvis, R.M.; Goodacre, R. Ultra-violet resonance Raman spectroscopy for the rapid discrimination of urinary tract infection bacteria. FEMS Microbiol. Lett. 2004, 232, 127–132. [Google Scholar] [CrossRef][Green Version]
- Rebrošová, K.; Bernatová, S.; Šiler, M.; Uhlirova, M.; Samek, O.; Ježek, J.; Holá, V.; Růžička, F.; Zemanek, P. Raman spectroscopy—A tool for rapid differentiation among microbes causing urinary tract infections. Anal. Chim. Acta 2022, 1191, 339292. [Google Scholar] [CrossRef]
- Dryden, S.D.; Anastasova, S.; Satta, G.; Thompson, A.J.; Leff, D.R.; Darzi, A. Rapid uropathogen identification using surface enhanced Raman spectroscopy active filters. Sci. Rep. 2021, 11, 8802. [Google Scholar] [CrossRef]
- Wang, J.; Kong, K.; Guo, C.; Yin, G.; Meng, S.; Lan, L.; Luo, L.; Song, Y. Cultureless enumeration of live bacteria in urinary tract infection by single-cell Raman spectroscopy. Front. Microbiol. 2023, 14, 1144607. [Google Scholar] [CrossRef]
- Rebrošová, K.; Bernatová, S.; Šiler, M.; Mašek, J.; Samek, O.; Ježek, J.; Kizovsky, M.; Holá, V.; Zemanek, P.; Růžička, F. Rapid Identification of Pathogens Causing Bloodstream Infections by Raman Spectroscopy and Raman Tweezers. Microbiol. Spectrum 2023, 11, e00028-23. [Google Scholar] [CrossRef]
- Kotanen, C.N.; Martinez, L.; Alvarez, R.; Simecek, J.W. Surface enhanced Raman scattering spectroscopy for detection and identification of microbial pathogens isolated from human serum. Sens. Bio-Sens. Res. 2016, 8, 20–26. [Google Scholar] [CrossRef]
- Safir, F.; Vu, N.; Tadesse, L.F.; Firouzi, K.; Banaei, N.; Jeffrey, S.S.; Saleh, A.A.E.; Khuri-Yakub, B.P.T.; Dionne, J.A. Combining Acoustic Bioprinting with AI-Assisted Raman Spectroscopy for High-Throughput Identification of Bacteria in Blood. Nano Lett. 2023, 23, 2065–2073. [Google Scholar] [CrossRef]
- Han, Y.Y.; Lin, Y.C.; Cheng, W.C.; Lin, Y.T.; Teng, L.J.; Wang, J.K.; Wang, Y.L. Rapid antibiotic susceptibility testing of bacteria from patients’ blood via assaying bacterial metabolic response with surface-enhanced Raman spectroscopy. Sci. Rep. 2020, 10, 12538. [Google Scholar] [CrossRef]
- Nicolaou, N.; Xu, Y.; Goodacre, R. Fourier Transform Infrared and Raman Spectroscopies for the Rapid Detection, Enumeration, and Growth Interaction of the Bacteria Staphylococcus aureus and Lactococcus lactis ssp. cremoris in Milk. Anal. Chem. 2011, 83, 5681–5687. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Huang, H.; Peng, Y.; Wang, J.; Gao, Z. Rapid determination of Staphylococcus aureus enterotoxin B in milk using Raman spectroscopy and chemometric methods. J. Raman Spectrosc. 2022, 53, 709–714. [Google Scholar] [CrossRef]
- Jarvis, R.M.; Goodacre, R. Discrimination of Bacteria Using Surface-Enhanced Raman Spectroscopy. Anal. Chem. 2004, 76, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Lima, C.; Muhamadali, H.; Goodacre, R. The Role of Raman Spectroscopy within Quantitative Metabolomics. Annu. Rev. Anal. Chem. 2021, 14, 323–345. [Google Scholar] [CrossRef]
- Azemtsop Matanfack, G.; Pistiki, A.; Rösch, P.; Popp, J. Raman 18O-Labeling of Bacteria in Visible and Deep UV-Ranges. J. Biophotonics 2021, 14, e202100013. [Google Scholar] [CrossRef]
- Stöckel, S.; Meisel, S.; Elschner, M.; Melzer, F.; Rösch, P.; Popp, J. Raman spectroscopic detection and identification of Burkholderia mallei and Burkholderia pseudomallei in feedstuff. Anal. Bioanal. Chem. 2015, 407, 787–794. [Google Scholar] [CrossRef]
- Lu, X.; Huang, Q.; Miller, W.G.; Aston, D.E.; Xu, J.; Xue, F.; Zhang, H.; Rasco, B.A.; Wang, S.; Konkel, M.E. Comprehensive Detection and Discrimination of Campylobacter Species by Use of Confocal Micro-Raman Spectroscopy and Multilocus Sequence Typing. J. Clin. Microbiol. 2012, 50, 2932–2946. [Google Scholar] [CrossRef]
- Rösch, P.; Harz, M.; Schmitt, M.; Peschke, K.-D.; Ronneberger, O.; Burkhardt, H.; Motzkus, H.-W.; Lankers, M.; Hofer, S.; Thiele, H.; et al. Chemotaxonomic Identification of Single Bacteria by Micro-Raman Spectroscopy: Application to Clean-Room-Relevant Biological Contaminations. Appl. Environ. Microbiol. 2005, 71, 1626–1637. [Google Scholar] [CrossRef]
- Rousseau, A.N.; Faure, N.; Rol, F.; Sedaghat, Z.; Le Galudec, J.; Mallard, F.; Josso, Q. Fast Antibiotic Susceptibility Testing via Raman Microspectrometry on Single Bacteria: An MRSA Case Study. ACS Omega 2021, 6, 16273–16279. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Timermans, C.; Rubbens, P.; Kerckhof, F.M.; Buysschaert, B.; Khalenkow, D.; Waegeman, W.; Skirtach, A.G.; Boon, N. Label-free Raman characterization of bacteria calls for standardized procedures. J. Microbiol. Meth. 2018, 151, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Nakar, A.; Pistiki, A.; Ryabchykov, O.; Bocklitz, T.; Rösch, P.; Popp, J. Label-free differentiation of clinical E. coli and Klebsiella isolates with Raman Spectroscopy. J. Biophotonics 2022, 15, e202200005. [Google Scholar] [CrossRef] [PubMed]
- Lima, C.; Ahmed, S.; Xu, Y.; Muhamadali, H.; Parry, C.; McGalliard, R.J.; Carrol, E.D.; Goodacre, R. Simultaneous Raman and infrared spectroscopy: A novel combination for studying bacterial infections at the single cell level. Chem. Sci. 2022, 13, 8171–8179. [Google Scholar] [CrossRef]
- Nakar, A.; Pistiki, A.; Ryabchykov, O.; Bocklitz, T.; Rösch, P.; Popp, J. Detection of multi-resistant clinical strains of E. coli with Raman spectroscopy. Anal. Bioanal. Chem. 2022, 414, 1481–1492. [Google Scholar] [CrossRef]
- Kirschner, C.; Maquelin, K.; Pina, P.; Thi, N.A.N.; Choo-Smith, L.-P.; Sockalingum, G.D.; Sandt, C.; Ami, D.; Orsini, F.; Doglia, S.M.; et al. Classification and Identification of Enterococci: A Comparative Phenotypic, Genotypic, and Vibrational Spectroscopic Study. J. Clin. Microbiol. 2001, 39, 1763–1770. [Google Scholar] [CrossRef]
- Assmann, C.; Kirchhoff, J.; Beleites, C.; Hey, J.; Kostudis, S.; Pfister, W.; Schlattmann, P.; Popp, J.; Neugebauer, U. Identification of vancomycin interaction with Enterococcus faecalis within 30 min of interaction time using Raman spectroscopy. Anal. Bioanal. Chem. 2015, 407, 8343–8352. [Google Scholar] [CrossRef]
- Kloß, S.; Lorenz, B.; Dees, S.; Labugger, I.; Rösch, P.; Popp, J. Destruction-free procedure for the isolation of bacteria from sputum samples for Raman spectroscopic analysis. Anal. Bioanal. Chem. 2015, 407, 8333–8341. [Google Scholar] [CrossRef]
- Harz, M.; Claus, R.A.; Bockmeyer, C.L.; Baum, M.; Rösch, P.; Kentouche, K.; Deigner, H.-P.; Popp, J. UV-Resonance Raman Spectroscopic Study of Human Plasma of Healthy Donors and Patients with Thrombotic Microangiopathy. Biopolymers 2006, 82, 317–324. [Google Scholar] [CrossRef]
- Silge, A.; Heinke, R.; Bocklitz, T.; Wiegand, C.; Hipler, U.-C.; Rösch, P.; Popp, J. The application of UV resonance Raman spectroscopy for the differentiation of clinically relevant Candida species. Anal. Bioanal. Chem. 2018, 410, 5839–5847. [Google Scholar] [CrossRef]
- López-Díez, E.C.; Goodacre, R. Characterization of Microorganisms Using UV Resonance Raman Spectroscopy and Chemometrics. Anal. Chem. 2004, 76, 585–591. [Google Scholar] [CrossRef] [PubMed]
- López-Díez, E.C.; Winder, C.L.; Ashton, L.; Currie, F.; Goodacre, R. Monitoring the Mode of Action of Antibiotics Using Raman Spectroscopy: Investigating Subinhibitory Effects of Amikacin on Pseudomonas aeruginosa. Anal. Chem. 2005, 77, 2901–2906. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Hamilton, T.; Nelson, W.H.; Elliott, S.; Sperry, J.F.; Wu, M. UV Raman Spectral Intensities of E. coli and Other Bacteria Excited at 228.9, 244.0, and 248.2 nm. Anal. Chem. 2001, 73, 3432–3440. [Google Scholar] [CrossRef] [PubMed]
- Manoharan, R.; Ghiamati, E.; Dalterio, R.A.; Britton, K.A.; Nelson, W.H.; Sperry, J.F. UV resonance Raman spectra of bacteria, bacterial spores, protoplasts and calcium dipicolinate. J. Microbiol. Meth. 1990, 11, 1–15. [Google Scholar] [CrossRef]
- Nelson, W.H.; Manoharan, R.; Sperry, J.F. UV Resonance Raman Studies of Bacteria. Appl. Spectrosc. Rev. 1992, 27, 67–124. [Google Scholar] [CrossRef]
- Manoharan, R.; Ghiamati, E.; Chadha, S.; Nelson, W.H.; Sperry, J.F. Effect of Cultural Conditions on Deep UV Resonance Raman Spectra of Bacteria. Appl. Spectrosc. 1993, 47, 2145–2150. [Google Scholar] [CrossRef]
- Buijtels, P.C.A.M.; Willemse-Erix, H.F.M.; Petit, P.L.C.; Endtz, H.P.; Puppels, G.J.; Verbrugh, H.A.; van Belkum, A.; van Soolingen, D.; Maquelin, K. Rapid Identification of Mycobacteria by Raman Spectroscopy. J. Clin. Microbiol. 2008, 46, 961–965. [Google Scholar] [CrossRef]
- Willemse-Erix, H.F.; Jachtenberg, J.; Barutçi, H.; Puppels, G.J.; van Belkum, A.; Vos, M.C.; Maquelin, K. Proof of Pprinciple for Successful Characterization of Methicillin-Resistant Coagulase-Negative Staphylococci Isolated from Skin by Use of Raman Spectroscopy and Pulsed-Field Gel Electrophoresis. J. Clin. Microbiol. 2010, 48, 736–740. [Google Scholar] [CrossRef]
- Tang, M.; McEwen, G.D.; Wu, Y.; Miller, C.D.; Zhou, A. Characterization and analysis of mycobacteria and Gram-negative bacteria and co-culture mixtures by Raman microspectroscopy, FTIR, and atomic force microscopy. Anal. Bioanal. Chem. 2013, 405, 1577–1591. [Google Scholar] [CrossRef]
- Muhamadali, H.; Weaver, D.; Subaihi, A.; AlMasoud, N.; Trivedi, D.K.; Ellis, D.I.; Linton, D.; Goodacre, R. Chicken, beams, and Campylobacter: Rapid differentiation of foodborne bacteria via vibrational spectroscopy and MALDI-mass spectrometry. Analyst 2016, 141, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Oust, A.; Møretrø, T.; Naterstad, K.; Sockalingum, G.D.; Adt, I.; Manfait, M.; Kohler, A. Fourier Transform Infrared and Raman Spectroscopy for Characterization of Listeria monocytogenes Strains. Appl. Environ. Microbiol. 2006, 72, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Rösch, P.; Schmitt, M.; Kiefer, W.; Popp, J. The identification of microorganisms by micro-Raman spectroscopy. J. Mol. Struct. 2003, 661–662, 363–369. [Google Scholar] [CrossRef]
- Ryabchykov, O.; Bocklitz, T.; Ramoji, A.; Neugebauer, U.; Foerster, M.; Kroegel, C.; Bauer, M.; Kiehntopf, M.; Popp, J. Automatization of spike correction in Raman spectra of biological samples. Chemometr. Intell. Lab. 2016, 155, 1–6. [Google Scholar] [CrossRef]
- Almarashi, J.F.; Kapel, N.; Wilkinson, T.S.; Telle, H.H. Raman Spectroscopy of Bacterial Species and Strains Cultivated Under Reproducible Conditions. Spectrosc. Int. J. 2012, 27, 361–365. [Google Scholar] [CrossRef]
- Assaf, A.; Cordella, C.B.; Thouand, G. Raman spectroscopy applied to the horizontal methods ISO 6579:2002 to identify Salmonella spp. in the food industry. Anal. Bioanal. Chem. 2014, 406, 4899–4910. [Google Scholar] [CrossRef][Green Version]
- Shen, H.; Rösch, P.; Popp, J. Fiber Probe-Based Raman Spectroscopic Identification of Pathogenic Infection Microorganisms on Agar Plates. Anal. Chem. 2022, 94, 4635–4642. [Google Scholar] [CrossRef]
- Ellis, D.I.; Cowcher, D.P.; Ashton, L.; O’Hagan, S.; Goodacre, R. Illuminating disease and enlightening biomedicine: Raman spectroscopy as a diagnostic tool. Analyst 2013, 138, 3871–3884. [Google Scholar] [CrossRef]
- Meisel, S.; Stöckel, S.; Elschner, M.; Melzer, F.; Rösch, P.; Popp, J. Raman Spectroscopy as a Potential Tool for Detection of Brucella spp. in Milk. Appl. Environ. Microbiol. 2012, 78, 5575–5583. [Google Scholar] [CrossRef]
- Li, Y.-S.; Church, J.S. Raman spectroscopy in the analysis of food and pharmaceutical nanomaterials. J. Food Drug Anal. 2014, 22, 29–48. [Google Scholar] [CrossRef]
- Pahlow, S.; Hentschel, S.; Horbert, P.; Romero, C.; Lehniger, L.; Wagner, S.; Popp, J.; Weber, K. Isolation of pathogenic bacteria from sputum samples using a 3D-printed cartridge system. Anal. Meth. 2021, 13, 4884–4895. [Google Scholar] [CrossRef]
- Stöckel, S.; Meisel, S.; Lorenz, B.; Kloß, S.; Henk, S.; Dees, S.; Richter, E.; Andres, S.; Merker, M.; Labugger, I.; et al. Raman spectroscopic identification of Mycobacterium tuberculosis. J. Biophotonics 2017, 10, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.N.V.; Guo, S.; Bocklitz, T.; Rösch, P.; Popp, J. Demonstration of Carbon Catabolite Repression in Naphthalene Degrading Soil Bacteria via Raman Spectroscopy Based Stable Isotope Probing. Anal. Chem. 2016, 88, 7574–7582. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, O.M.; Abd El Kareem, H.; Selim, N. Nitrate modulation of Bacillus sp. biofilm components: A proposed model for sustainable bioremediation. Biotechnol. Lett. 2021, 43, 2185–2197. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, J.; Das, S. Application of spectroscopic techniques for monitoring microbial diversity and bioremediation. Appl. Spectrosc. Rev. 2016, 52, 1–38. [Google Scholar] [CrossRef]
- Olaniyi, O.O.; Yang, K.; Zhu, Y.G.; Cui, L. Heavy water-labeled Raman spectroscopy reveals carboxymethylcellulose-degrading bacteria and degradation activity at the single-cell level. Appl. Microbiol. Biotechnol. 2019, 103, 1455–1464. [Google Scholar] [CrossRef] [PubMed]
- Shokrekhodaei, M.; Quinones, S. Review of Non-invasive Glucose Sensing Techniques: Optical, Electrical and Breath Acetone. Sensors 2020, 20, 1251. [Google Scholar] [CrossRef] [PubMed]
- Das, R.S.; Agrawal, Y.K. Raman spectroscopy: Recent advancements, techniques and applications. Vib. Spectrosc. 2011, 57, 163–176. [Google Scholar] [CrossRef]
- Esmonde-White, K.A.; Cuellar, M.; Uerpmann, C.; Lenain, B.; Lewis, I.R. Raman spectroscopy as a process analytical technology for pharmaceutical manufacturing and bioprocessing. Anal. Bioanal. Chem. 2017, 409, 637–649. [Google Scholar] [CrossRef]
- Bergholt, M.S.; Serio, A.; Albro, M.B. Raman Spectroscopy: Guiding Light for the Extracellular Matrix. Front. Bioeng. Biotechnol. 2019, 7, 303. [Google Scholar] [CrossRef]
- Sivakesava, S.; Irudayaraj, J.; Demirci, A. Monitoring a bioprocess for ethanol production using FT-MIR and FT-Raman spectroscopy. J. Ind. Microbiol. Biotechnol. 2001, 26, 185–190. [Google Scholar] [CrossRef]
- Baron, E.J.; Miller, J.M.; Weinstein, M.P.; Richter, S.S.; Gilligan, P.H.; Thomson, R.B., Jr.; Bourbeau, P.; Carroll, K.C.; Kehl, S.C.; Dunne, W.M.; et al. A Guide to Utilization of the Microbiology Laboratory for Diagnosis of Infectious Diseases: 2013 Recommendations by the Infectious Diseases Society of America (IDSA) and the American Society for Microbiology (ASM). Clin. Infect. Dis. 2013, 57, e22–e121. [Google Scholar] [CrossRef]
- Sánchez-Romero, M.I.; Moya, J.M.G.-L.; López, J.J.G.; Mira, N.O. Collection, transport and general processing of clinical specimens in Microbiology laboratory. Enfermedades Infect. Microbiol. Clin. 2019, 37, 127–134. [Google Scholar] [CrossRef]
- Wilson, M.L. General Principles of Specimen Collection and Transport. Clin. Infect. Dis. 1996, 22, 766–777. [Google Scholar] [CrossRef] [PubMed]
- Escoriza, M.F.; Vanbriesen, J.M.; Stewart, S.; Maier, J. Studying Bacterial Metabolic States Using Raman Spectroscopy. Appl. Spectrosc. 2006, 60, 971–976. [Google Scholar] [CrossRef]
- Moritz, T.J.; Taylor, D.S.; Polage, C.R.; Krol, D.M.; Lane, S.M.; Chan, J.W. Effect of Cefazolin Treatment on the Nonresonant Raman Signatures of the Metabolic State of Individual Escherichia coli Cells. Anal. Chem. 2010, 82, 2703–2710. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.E.; Griffiths, R.I.; Thompson, I.P.; Bailey, M.J.; Whiteley, A.S. Raman Microscopic Analysis of Single Microbial Cells. Anal. Chem. 2004, 76, 4452–4458. [Google Scholar] [CrossRef]
- Ciobotă, V.; Burkhardt, E.-M.; Rösch, P.; Küsel, K.; Popp, J. The influence of intracellular storage material on bacterial identification by means of Raman spectroscopy. Anal. Bioanal. Chem. 2010, 397, 2929–2937. [Google Scholar] [CrossRef]
- WHO. Global Antimicrobial Resistance Surveillance System (GLASS): Molecular Methods for Antimicrobial Resistance (AMR) Diagnostics to Enhance the Global Antimicrobial Resistance Surveillance System; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Mauch, H. Importance of short storage and transportation time of samples for timely microbiological diagnosis considering patients safety and economic issues. J. Lab. Med. 2009, 33, 188–193. [Google Scholar]
- Hogardt, M. MIQ 24: Atemwegsinfektionen bei Mukoviszidose: Qualitätsstandards in der Mikrobiologisch-Infektiologischen Diagnostik; Elsevier Health Sciences: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Washington, J.A. Rapid Antimicrobial Susceptibility Testing: Technical and Clinical Considerations. Clin. Microbiol. Newslett. 1993, 15, 153–156. [Google Scholar] [CrossRef]
- Lynne, S.; Garcia, H.D.I. Specimen Collection, Transport, and Acceptability. In Clinical Microbiology Procedures Handbook, 3rd ed.; Wiley: Hoboken, NJ, USA, 2010. [Google Scholar]
- Pepper, I.L.; Gerba, C.P. Environmental sample collection and processing. In Environmental Microbiology; Academic Press: Cambridge, MA, USA, 2015; pp. 157–175. [Google Scholar]
- Alam, M.; Sadique, A.; Nur, A.H.; Bhuiyan, N.A.; Nair, G.B.; Siddique, A.K.; Sack, D.A.; Ahsan, S.; Huq, A.; Sack, R.B.; et al. Effect of Transport at Ambient Temperature on Detection and Isolation of Vibrio cholerae from Environmental Samples. Appl. Environ. Microbiol. 2006, 72, 2185–2190. [Google Scholar] [CrossRef]
- Rubin, B.E.; Gibbons, S.M.; Kennedy, S.; Hampton-Marcell, J.; Owens, S.; Gilbert, J.A. Investigating the Impact of Storage Conditions on Microbial Community Composition in Soil Samples. PLoS ONE 2013, 8, e70460. [Google Scholar] [CrossRef]
- Roelofsen, E.; Van Leeuwen, M.; Meijer-Severs, G.; Wilkinson, M.; Degener, J. Evaluation of the Effects of Storage in two Different Swab Fabrics and Under Three Different Transport Conditions on Recovery of Aerobic and Anaerobic Bacteria. J. Clin. Microbiol. 1999, 37, 3041–3043. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, H.; Dalton, H.P.; Escobar, M.R.; Allison, M.J. Transportation Delay and the Microbiological Quality of Clinical Specimens. Am. J. Clin. Pathol. 1975, 64, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Kanematsu, H.; Kudara, H.; Kanesaki, S.; Kogo, T.; Ikegai, H.; Ogawa, A.; Hirai, N. Application of a Loop-Type Laboratory Biofilm Reactor to the Evaluation of Biofilm for Some Metallic Materials and Polymers such as Urinary Stents and Catheters. Materials 2016, 9, 824. [Google Scholar] [CrossRef] [PubMed]
- Meisel, S.; Stöckel, S.; Elschner, M.; Rösch, P.; Popp, J. Assessment of two isolation techniques for bacteria in milk towards their compatibility to Raman spectroscopy. Analyst 2011, 136, 4997–5005. [Google Scholar] [CrossRef] [PubMed]
- Pahlow, S.; Kloß, S.; Blättel, V.; Kirsch, K.; Hübner, U.; Cialla, D.; Rösch, P.; Weber, K.; Popp, J. Isolation and Enrichment of Pathogens with a Surface-Modified Aluminium Chip for Raman Spectroscopic Applications. ChemPhysChem 2013, 14, 3600–3605. [Google Scholar] [CrossRef] [PubMed]
- Schröder, U.-C.; Bokeloh, F.; O’Sullivan, M.; Glaser, U.; Wolf, K.; Pfister, W.; Popp, J.; Ducrée, J.; Neugebauer, U. Rapid, culture-independent, optical diagnostics of centrifugally captured bacteria from urine samples. Biomicrofluidics 2015, 9, 044118. [Google Scholar] [CrossRef] [PubMed]
- Schröder, U.-C.; Ramoji, A.; Glaser, U.; Sachse, S.; Leiterer, C.; Csaki, A.; Hübner, U.; Fritzsche, W.; Pfister, W.; Bauer, M.; et al. Combined Dielectrophoresis–Raman Setup for the Classification of Pathogens Recovered from the Urinary Tract. Anal. Chem. 2013, 85, 10717–10724. [Google Scholar] [CrossRef] [PubMed]
- Schröder, U.C.; Kirchhoff, J.; Hübner, U.; Mayer, G.; Glaser, U.; Henkel, T.; Pfister, W.; Fritzsche, W.; Popp, J.; Neugebauer, U. On-chip spectroscopic assessment of microbial susceptibility to antibiotics within 3.5 hours. J. Biophotonics 2017, 10, 1547–1557. [Google Scholar] [CrossRef]
- Hahn, M.W.; Koll, U.; Schmidt, J. Isolation and Cultivation of Bacteria. In The Structure and Function of Aquatic Microbial Communities; Springer: Berlin/Heidelberg, Germany, 2019; pp. 313–351. [Google Scholar]
- Choo-Smith, L.-P.; Maquelin, K.; Van Vreeswijk, T.; Bruining, H.; Puppels, G.; Thi, N.N.; Kirschner, C.; Naumann, D.; Ami, D.; Villa, A. Investigating Microbial (Micro)colony Heterogeneity by Vibrational Spectroscopy. Appl. Environ. Microbiol. 2001, 67, 1461–1469. [Google Scholar] [CrossRef]
- Mathey, R.; Dupoy, M.; Espagnon, I.; Leroux, D.; Mallard, F.; Novelli-Rousseau, A. Viability of 3 h grown bacterial micro-colonies after direct Raman identification. J. Microbiol. Meth. 2015, 109, 67–73. [Google Scholar] [CrossRef]
- Ramoji, A.; Galler, K.; Glaser, U.; Henkel, T.; Mayer, G.; Dellith, J.; Bauer, M.; Popp, J.; Neugebauer, U. Characterization of different substrates for Raman spectroscopic imaging of eukaryotic cells. J. Raman Spectrosc. 2016, 47, 773–786. [Google Scholar] [CrossRef]
- Mikoliunaite, L.; Rodriguez, R.D.; Sheremet, E.; Kolchuzhin, V.; Mehner, J.; Ramanavicius, A.; Zahn, D.R.T. The substrate matters in the Raman spectroscopy analysis of cells. Sci. Rep. 2015, 5, 13150. [Google Scholar] [CrossRef]
- Bonnet, M.; Lagier, J.C.; Raoult, D.; Khelaifia, S. Bacterial culture through selective and non-selective conditions: The evolution of culture media in clinical microbiology. New Microbe New Infect. 2020, 34, 100622. [Google Scholar] [CrossRef]
- Stöckel, S.; Meisel, S.; Böhme, R.; Elschner, M.; Rösch, P.; Popp, J. Effect of supplementary manganese on the sporulation of Bacillus endospores analysed by Raman spectroscopy. J. Raman Spectrosc. 2009, 40, 1469–1477. [Google Scholar] [CrossRef]
- Stöckel, S.; Schumacher, W.; Meisel, S.; Elschner, M.; Rösch, P.; Popp, J. Raman Spectroscopy-Compatible Inactivation Method for Pathogenic Endospores. Appl. Environ. Microbiol. 2010, 76, 2895–2907. [Google Scholar] [CrossRef]
- Mariani, M.M.; Lampen, P.; Popp, J.; Wood, B.R.; Deckert, V. Impact of fixation on in vitro cell culture lines monitored with Raman spectroscopy. Analyst 2009, 134, 1154–1161. [Google Scholar] [CrossRef]
- Pistiki, A.; Ryabchykov, O.; Bocklitz, T.; Rösch, P.; Popp, J. Calibration for UV resonance Raman spectroscopy. Spectrochim. Acta A 2023, 287, 122062. [Google Scholar] [CrossRef]
- Amann, R.I.; Ludwig, W.; Schleifer, K.H. Phylogenetic Identification and In Situ Detection of Individual Microbial Cells without Cultivation. Microbiol. Rev. 1995, 59, 143–169. [Google Scholar] [CrossRef]
- Hofer, U. The majority is uncultured. Nat. Rev. Microbiol. 2018, 16, 716–717. [Google Scholar] [CrossRef]
- Hugenholtz, P.; Goebel, B.M.; Pace, N.R. Impact of Culture-Independent Studies on the Emerging Phylogenetic View of Bacterial Diversity. J. Bacteriol. 1998, 180, 4765–4774. [Google Scholar] [CrossRef]
- Hahn, M.W.; Stadler, P.; Wu, Q.L.; Pockl, M. The filtration-acclimatization method for isolation of an important fraction of the not readily cultivable bacteria. J. Microbiol. Meth. 2004, 57, 379–390. [Google Scholar] [CrossRef]
- Chon, J.W.; Kim, D.H.; Bae, D.; Song, K.Y.; Kim, H.; Sung, K.; Seo, K.H. Comparison of Direct Syringe Filtration and Membrane Filtration for the Selective Isolation of Campylobacter jejuni from Ready-to-Eat Sprouts. Foodborne Pathog. Dis. 2019, 16, 371–375. [Google Scholar] [CrossRef]
- Stevens, K.A.; Jaykus, L.A. Bacterial separation and concentration from complex sample matrices: A review. Crit. Rev. Microbiol. 2004, 30, 7–24. [Google Scholar] [CrossRef]
- Chen, J.; Wu, X.; Huang, Y.-W.; Zhao, Y. Detection of E. coli using SERS active filters with silver nanorod array. Sens. Actuators B 2014, 191, 485–490. [Google Scholar] [CrossRef]
- Ravindranath, S.P.; Wang, Y.; Irudayaraj, J. SERS driven cross-platform based multiplex pathogen detection. Sens. Actuators B 2011, 152, 183–190. [Google Scholar] [CrossRef]
- Sharpe, A. Separation and Concentration of Pathogens from Foods; Marcel Dekker: New York, NY, USA, 1997. [Google Scholar]
- Thomas, D.S. Electropositively charged filters for the recovery of yeasts and bacteria from beverages. J. Appl. Bacteriol. 1988, 65, 35–41. [Google Scholar] [CrossRef]
- Mortimer, M.; Petersen, E.J.; Buchholz, B.A.; Holden, P.A. Separation of Bacteria, Protozoa and Carbon Nanotubes by Density Gradient Centrifugation. Nanomaterials 2016, 6, 181. [Google Scholar] [CrossRef] [PubMed]
- Stöckel, S.; Meisel, S.; Elschner, M.; Rösch, P.; Popp, J. Identification of Bacillus anthracis via Raman Spectroscopy and Chemometric Approaches. Anal. Chem. 2012, 84, 9873–9880. [Google Scholar] [CrossRef] [PubMed]
- Stöckel, S.; Meisel, S.; Elschner, M.; Rösch, P.; Popp, J. Raman Spectroscopic Detection of Anthrax Endospores in Powder Samples. Angew. Chem. Int. Ed. 2012, 51, 5339–5342. [Google Scholar] [CrossRef]
- Kovacovicova, K.; Vinciguerra, M. Isolation of senescent cells by iodixanol (OptiPrep) density gradient-based separation. Cell Prolif. 2019, 52, e12674. [Google Scholar] [CrossRef]
- Kumar, R.K.; Lykke, A.W. Cell separation: A review. Pathology 1984, 16, 53–62. [Google Scholar] [CrossRef]
- Bell, C.H. The Effects of Centrifugation and Filtration as Pre-Treatement in Bacterial Retention Studies. J. Young Investig. 2005. Available online: https://api.semanticscholar.org/CorpusID:105389251 (accessed on 16 February 2024).
- Peterson, B.W.; Sharma, P.K.; van der Mei, H.C.; Busscher, H.J. Bacterial Cell Surface Damage Due to Centrifugal Compaction. Appl. Environ. Microbiol. 2012, 78, 120–125. [Google Scholar] [CrossRef]
- Gilbert, P.; Coplan, F.; Brown, M.R. Centrifugation injury of Gram-negative bacteria. J. Antimicrob. Chemother. 1991, 27, 550–551. [Google Scholar] [CrossRef] [PubMed]
- Pembrey, R.S.; Marshall, K.C.; Schneider, R.P. Cell Surface Analysis Techniques: What Do Cell Preparation Protocols Do to Cell Surface Properties? Appl. Environ. Microbiol. 1999, 65, 2877–2894. [Google Scholar] [CrossRef]
- Tsuneda, S.; Jung, J.; Hayashi, H.; Aikawa, H.; Hirata, A.; Sasaki, H. Influence of extracellular polymers on electrokinetic properties of heterotrophic bacterial cells examined by soft particle electrophoresis theory. Colloid Surf. B 2003, 29, 181–188. [Google Scholar] [CrossRef]
- Pohl, H.A.; Crane, J.S. Dielectrophoresis of Cells. Biophys. J. 1971, 11, 711–727. [Google Scholar] [CrossRef]
- Pohl, H.A. Dielectrophoresis: The Behavior of Neutral Matter in Nonuniform Electric Fields. 1978. Available online: https://api.semanticscholar.org/CorpusID:117876476 (accessed on 16 February 2024).
- Brown, A.P.; Betts, W.B.; Harrison, A.B.; O’Neill, J.G. Evaluation of a dielectrophoretic bacterial counting technique. Biosens. Bioelectron. 1999, 14, 341–351. [Google Scholar] [CrossRef]
- Pethig, R. Review Article-Dielectrophoresis: Status of the Theory, Technology, and Applications. Biomicrofluidics 2010, 4, 022811. [Google Scholar] [CrossRef]
- Lorenz, B.; Ali, N.; Bocklitz, T.; Rösch, P.; Popp, J. Discrimination between pathogenic and non-pathogenic E. coli strains by means of Raman microspectroscopy. Anal. Bioanal. Chem. 2020, 412, 8241–8247. [Google Scholar] [CrossRef]
- Voldman, J. Electrical Forces for Microscale Cell Manipulation. Annu. Rev. Biomed. Eng. 2006, 8, 425–454. [Google Scholar] [CrossRef]
- Huang, Y.; Pethig, R. Electrode design for negative dielectrophoresis. Measur. Sci. Technol. 1991, 2, 1142. [Google Scholar] [CrossRef]
- Barzan, G.; Sacco, A.; Mandrile, L.; Giovannozzi, A.M.; Brown, J.; Portesi, C.; Alexander, M.R.; Williams, P.; Hardie, K.R.; Rossi, A.M. New frontiers against antibiotic resistance: A Raman-based approach for rapid detection of bacterial susceptibility and biocide-induced antibiotic cross-tolerance. Sens. Actuators B Chem. 2020, 309, 127774. [Google Scholar] [CrossRef]
- Páez-Avilés, C.; Juanola-Feliu, E.; Punter-Villagrasa, J.; Del Moral Zamora, B.; Homs-Corbera, A.; Colomer-Farrarons, J.; Miribel-Catala, P.L.; Samitier, J. Combined Dielectrophoresis and Impedance Systems for Bacteria Analysis in Microfluidic On-Chip Platforms. Sensors 2016, 16, 1514. [Google Scholar] [CrossRef]
- Hanson, C.; Barney, J.T.; Bishop, M.M.; Vargis, E. Simultaneous isolation and label-free identification of bacteria using contactless dielectrophoresis and Raman spectroscopy. Electrophoresis 2019, 40, 1446–1456. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Cao, Z.; Yuan, Y.J. Study on non-bioparticles and Staphylococcus aureus by dielectrophoresis. RSC Adv. 2020, 10, 2598–2614. [Google Scholar] [CrossRef] [PubMed]
- Zellner, P.; Agah, M. Silicon insulator-based dielectrophoresis devices for minimized heating effects. Electrophoresis 2012, 33, 2498–2507. [Google Scholar] [CrossRef] [PubMed]
- Kusić, D.; Rösch, P.; Popp, J. Fast label-free detection of Legionella spp. in biofilms by applying immunomagnetic beads and Raman spectroscopy. Syst. Appl. Microbiol. 2016, 39, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Ji, S.; Xiang, Y.; Han, D.; Liu, C.; Du, Y.; Peng, Y.; Li, S.; Ren, S.; Qin, K.; Wang, Y. Functionalized magnetic nanobeads for SERS-based detection of Staphylococcus aureus. Anal. Meth. 2023, 15, 3393–3403. [Google Scholar] [CrossRef]
- Rochelle, P.A.; De Leon, R.; Johnson, A.; Stewart, M.H.; Wolfe, R.L. Evaluation of Immunomagnetic Separation for Recovery of Infectious Cryptosporidium parvum Oocysts from Environmental Samples. Appl. Environ. Microbiol. 1999, 65, 841–845. [Google Scholar] [CrossRef] [PubMed]
- Gee, A.P. Immunomagnetic cell separation using antibodies and superparamagnetic microspheres. In Cell Separation Methods and Applications; CRC Press: Boca Raton, FL, USA, 1998. [Google Scholar]
- Wang, Z.; Cai, R.; Gao, Z.; Yuan, Y.; Yue, T. Immunomagnetic separation: An effective pretreatment technology for isolation and enrichment in food microorganisms detection. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3802–3824. [Google Scholar] [CrossRef]
- Skottrup, P.D.; Nicolaisen, M.; Justesen, A.F. Towards on-site pathogen detection using antibody-based sensors. Biosens. Bioelectron. 2008, 24, 339–348. [Google Scholar] [CrossRef]
- Li, S.; Marquardt, R.R.; Abramson, D. Immunochemical Detection of Molds: A Review. J. Food Prot. 2000, 63, 281–291. [Google Scholar] [CrossRef]
- Mircescu, N.E.; Zhou, H.; Leopold, N.; Chiş, V.; Ivleva, N.P.; Niessner, R.; Wieser, A.; Haisch, C. Towards a receptor-free immobilization and SERS detection of urinary tract infections causative pathogens. Anal. Bioanal. Chem. 2014, 406, 3051–3058. [Google Scholar] [CrossRef]
- Liu, B.; Liu, K.; Wang, N.; Ta, K.; Liang, P.; Yin, H.; Li, B. Laser tweezers Raman spectroscopy combined with deep learning to classify marine bacteria. Talanta 2022, 244, 123383. [Google Scholar] [CrossRef]
- Ai, M.; Liu, J.-X.; Huang, S.-S.; Wang, G.-W.; Chen, X.-L.; Chen, Z.-C.; Yao, H.-L. Application and Progress of Raman Tweezers in Single Cells. Chin. J. Anal. Chem. 2009, 37, 758–763. [Google Scholar] [CrossRef]
- Guo, H.; Li, Z. Optical tweezers technique and its applications. Sci. China-Phys. Mech. Astron. 2013, 56, 2351–2360. [Google Scholar] [CrossRef]
- Yan, S.; Qiu, J.; Guo, L.; Li, D.; Xu, D.; Liu, Q. Development overview of Raman-activated cell sorting devoted to bacterial detection at single-cell level. Appl. Microbiol. Biotechnol. 2021, 105, 1315–1331. [Google Scholar] [CrossRef]
- Bernatová, S.; Rebrosová, K.; Pilát, Z.; Sery, M.; Gjevik, A.; Samek, O.; Jezek, J.; Siler, M.; Kizovsky, M.; Klementová, T.; et al. Rapid detection of antibiotic sensitivity of Staphylococcus aureus by Raman tweezers. Eur. Phys. J. Plus 2021, 136, 233. [Google Scholar] [CrossRef]
- Du, F.S.; He, L.; Lu, X.X.; Li, Y.Q.; Yuan, Y.F. Accurate identification of living Bacillus spores using laser tweezers Raman spectroscopy and deep learning. Spectrochim. Acta A 2023, 289, 122216. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.S.; Dong, X.S.; Liu, Y.M.H.; Wright, K.M.; Humphris, S.N.; Dupuy, L.X.; MacDonald, M.P. In situ control of root-bacteria interactions using optical trapping in transparent soil. J. Exp. Bot. 2023, 74, 787–799. [Google Scholar] [CrossRef] [PubMed]
- Vaculík, O.; Bernatová, S.; Rebrošová, K.; Samek, O.; Šilhan, L.; Růžička, F.; Šery, M.; Šiler, M.; Ježek, J.; Zemanek, P. Rapid identification of pathogens in blood serum via Raman tweezers in combination with advanced processing methods. Biomed. Opt. Exp. 2023, 14, 6410–6421. [Google Scholar] [CrossRef]
- Nilsson, D.P.; Jonsmoen, U.L.; Malyshev, D.; Öberg, R.; Wiklund, K.; Andersson, M. Physico-chemical characterization of single bacteria and spores using optical tweezers. Res. Microbiol. 2023, 174, 104060. [Google Scholar] [CrossRef] [PubMed]
- Snook, R.D.; Harvey, T.J.; Correia Faria, E.; Gardner, P. Raman tweezers and their application to the study of singly trapped eukaryotic cells. Integr. Biol. 2009, 1, 43–52. [Google Scholar] [CrossRef]
- Wang, Y.H.; Chen, J.; Su, G.F.; Mei, J.X.; Li, J.Y. A Review of Single-Cell Microrobots: Classification, Driving Methods and Applications. Micromachines 2023, 14, 1710. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Sun, L.Y.; Fang, T.; Li, H.X.; Zhang, R.; Ye, A.P. Rapid and accurate identification of pathogenic bacteria at the single-cell level using laser tweezers Raman spectroscopy and deep learning. J. Biophotonics 2022, 15, e202100312. [Google Scholar] [CrossRef]
- Huang, W.E.; Ward, A.D.; Whiteley, A.S. Raman tweezers sorting of single microbial cells. Environ. Microbiol. Rep. 2009, 1, 44–49. [Google Scholar] [CrossRef]
- Lee, K.S.; Pereira, F.C.; Palatinszky, M.; Behrendt, L.; Alcolombri, U.; Berry, D.; Wagner, M.; Stocker, R. Optofluidic Raman-activated cell sorting for targeted genome retrieval or cultivation of microbial cells with specific functions. Nat. Prot. 2021, 16, 45. [Google Scholar] [CrossRef]
- Pesce, G.; Jones, P.H.; Maragò, O.M.; Volpe, G. Optical tweezers: Theory and practice. Eur. Phys. J. Plus 2020, 135, 949. [Google Scholar] [CrossRef]
- Malyshev, D.; Öberg, R.; Dahlberg, T.; Wiklund, K.; Landström, L.; Andersson, P.O.; Andersson, M. Laser induced degradation of bacterial spores during micro-Raman spectroscopy. Spectrochim. Acta A 2022, 265, 120381. [Google Scholar] [CrossRef]
- Bhat, A.V.; Soma, V.R.; Ananthamurthy, S. Probing the Onset of the Bacterial Membrane Photodamage in Time using a Raman Optical Tweezer. Vib. Spectrosc. 2023, 129, 103611. [Google Scholar] [CrossRef]
- Hu, S.; Ye, J.-Y.; Zhao, Y.; Zhu, C.-L. Advanced optical tweezers on cell manipulation and analysis. Eur. Phys. J. Plus 2022, 137, 1024. [Google Scholar] [CrossRef]
- Ryabchykov, O.; Guo, S.; Bocklitz, T. Analyzing Raman spectroscopic data. Phys. Sci. Rev. 2018, 4, 20170043. [Google Scholar]
- Guo, S.X.; Popp, J.; Bocklitz, T. Chemometric analysis in Raman spectroscopy from experimental design to machine learning-based modeling. Nat. Protoc. 2021, 16, 5426–5459. [Google Scholar] [CrossRef] [PubMed]
- Ryabchykov, O.; Guo, S.; Bocklitz, T. 4. Analyzing Raman spectroscopic data. In Micro-Raman Spectroscopy; Jürgen, P., Thomas, M., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2020; pp. 81–106. [Google Scholar]
- Guo, S.; Ryabchykov, O.; Ali, N.; Houhou, R.; Bocklitz, T. Comprehensive chemometrics. In Comprehensive Chemometrics-Chemical and Biochemical Data Analysis, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 333–359. [Google Scholar]
- Smith, E.; Dent, G. Modern Raman Spectroscopy: A Practical Approach; John Wiley & Sons: Hoboken, NJ, USA, 2019. [Google Scholar]
- Butler, H.J.; Ashton, L.; Bird, B.; Cinque, G.; Curtis, K.; Dorney, J.; Esmonde-White, K.; Fullwood, N.J.; Gardner, B.; Martin-Hirsch, P.L. Using Raman spectroscopy to characterize biological materials. Nat. Protoc. 2016, 11, 664–687. [Google Scholar] [CrossRef]
- Ali, N.; Girnus, S.; Rösch, P.; Popp, J.; Bocklitz, T. Sample-Size Planning for Multivariate Data: A Raman-Spectroscopy-Based Example. Anal. Chem. 2018, 90, 12485–12492. [Google Scholar] [CrossRef]
- Hajian-Tilaki, K. Sample size estimation in diagnostic test studies of biomedical informatics. J. Biomed. Inform. 2014, 48, 193–204. [Google Scholar] [CrossRef]
- Saccenti, E.; Timmerman, M.E. Approaches to Sample Size Determination for Multivariate Data: Applications to PCA and PLS-DA of Omics Data. J. Proteome Res. 2016, 15, 2379–2393. [Google Scholar] [CrossRef]
- Morais, C.L.; Lima, K.M.; Singh, M.; Martin, F.L. Tutorial: Multivariate classification for vibrational spectroscopy in biological samples. Nat. Protoc. 2020, 15, 2143–2162. [Google Scholar] [CrossRef] [PubMed]
- Barton, S.J.; Hennelly, B.M. An Algorithm for the Removal of Cosmic Ray Artifacts in Spectral Data Sets. Appl. Spectrosc. 2019, 73, 893–901. [Google Scholar] [CrossRef] [PubMed]
- Anthony, S.M.; Timlin, J.A. Removing Cosmic Spikes Using a Hyperspectral Upper-Bound Spectrum Method. Appl. Spectrosc. 2017, 71, 507–519. [Google Scholar] [CrossRef]
- Maury, A.; Revilla, R.I. Autocorrelation Analysis Combined with a Wavelet Transform Method to Detect and Remove Cosmic Rays in a Single Raman Spectrum. Appl. Spectrosc. 2015, 69, 984–992. [Google Scholar] [CrossRef]
- Sharan, T.; Sharma, S.; Sharma, N. Denoising and spike removal from Raman spectra using double density dual-tree complex wavelet transform. J. Appl. Spectrosc. 2021, 88, 117–124. [Google Scholar] [CrossRef]
- Whitaker, D.A.; Hayes, K. A simple algorithm for despiking Raman spectra. Chemometr. Intell. Lab. 2018, 179, 82–84. [Google Scholar] [CrossRef]
- Yamamoto, T.; Taylor, J.N.; Koseki, S.; Koyama, K. Classification of food spoilage bacterial species and their sodium chloride, sodium acetate and glycine tolerance using chemometrics analysis and Raman spectroscopy. J. Microbiol. Meth. 2021, 190, 106326. [Google Scholar] [CrossRef]
- Bocklitz, T.W.; Dörfer, T.; Heinke, R.; Schmitt, M.; Popp, J. Spectrometer calibration protocol for Raman spectra recorded with different excitation wavelengths. Spectrochim. Acta A 2015, 149, 544–549. [Google Scholar] [CrossRef]
- Mostafapour, S.; Dörfer, T.; Heinke, R.; Rösch, P.; Popp, J.; Bocklitz, T. Investigating the effect of different pre-treatment methods on Raman spectra recorded with different excitation wavelengths. Spectrochim. Acta A 2023, 302, 123100. [Google Scholar] [CrossRef] [PubMed]
- Raj, A.; Kato, C.; Witek, H.A.; Hamaguchi, H.O. Toward standardization of Raman spectroscopy: Accurate wavenumber and intensity calibration using rotational Raman spectra of H2, HD, D2, and vibration–rotation spectrum of O2. J. Raman Spectrosc. 2020, 51, 2066–2082. [Google Scholar] [CrossRef]
- Okajima, H.; Hamaguchi, H.O. Accurate intensity calibration for low wavenumber (−150 to 150 cm−1) Raman spectroscopy using the pure rotational spectrum of N2. J. Raman Spectrosc. 2015, 46, 1140–1144. [Google Scholar] [CrossRef]
- Mishra, P.; Biancolillo, A.; Roger, J.M.; Marini, F.; Rutledge, D.N. New data preprocessing trends based on ensemble of multiple preprocessing techniques. TrAC—Trends Anal. Chem. 2020, 132, 116045. [Google Scholar] [CrossRef]
- Rinnan, Å. Pre-processing in vibrational spectroscopy—When, why and how. Anal. Meth. 2014, 6, 7124–7129. [Google Scholar] [CrossRef]
- Helin, R.; Indahl, U.G.; Tomic, O.; Liland, K.H. On the possible benefits of deep learning for spectral preprocessing. J. Chemom. 2022, 36, e3374. [Google Scholar] [CrossRef]
- Kazemzadeh, M.; Hisey, C.L.; Martinez-Calderon, M.; Chamley, L.W.; Xu, P.; Broderick, N. Deep learning as an improved method of preprocessing biomedical Raman spectroscopy data. TechRxiv 2022. [Google Scholar] [CrossRef]
- Korinth, F.; Shaik, T.A.; Popp, J.; Krafft, C. Assessment of shifted excitation Raman difference spectroscopy in highly fluorescent biological samples. Analyst 2021, 146, 6760–6767. [Google Scholar] [CrossRef]
- Guo, Y.; Jin, W.; Wang, W.; He, Y.; Qiu, S. Baseline correction for Raman spectra using a spectral estimation-based asymmetrically reweighted penalized least squares method. Appl. Opt. 2023, 62, 4766–4776. [Google Scholar] [CrossRef]
- Chen, K.; Zhang, H.; Wei, H.; Li, Y. Improved Savitzky–Golay-method-based fluorescence subtraction algorithm for rapid recovery of Raman spectra. Appl. Opt. 2014, 53, 5559–5569. [Google Scholar] [CrossRef] [PubMed]
- Barton, B.; Thomson, J.; Diz, E.L.; Portela, R. Chemometrics for Raman Spectroscopy Harmonization. Appl. Spectrosc. 2022, 76, 1021–1041. [Google Scholar] [CrossRef] [PubMed]
- Sadergaski, L.R.; Hager, T.J.; Andrews, H.B. Design of Experiments, Chemometrics, and Raman Spectroscopy for the Quantification of Hydroxylammonium, Nitrate, and Nitric Acid. ACS Omega 2022, 7, 7287–7296. [Google Scholar] [CrossRef]
- Storey, E.E.; Helmy, A.S. Optimized preprocessing and machine learning for quantitative Raman spectroscopy in biology. J. Raman Spectrosc. 2019, 50, 958–968. [Google Scholar] [CrossRef]
- Wang, H.-P.; Chen, P.; Dai, J.-W.; Liu, D.; Li, J.-Y.; Xu, Y.-P.; Chu, X.-L. Recent advances of chemometric calibration methods in modern spectroscopy: Algorithms, strategy, and related issues. TrAC—Trends Anal. Chem. 2022, 153, 116648. [Google Scholar] [CrossRef]
- Salem, N.; Hussein, S. Data dimensional reduction and principal components analysis. Procedia Comput. Sci. 2019, 163, 292–299. [Google Scholar] [CrossRef]
- Gracia, A.; González, S.; Robles, V.; Menasalvas, E. A methodology to compare dimensionality reduction algorithms in terms of loss of quality. Inform. Sci. 2014, 270, 1–27. [Google Scholar] [CrossRef]
- Yang, J.; Xu, J.; Zhang, X.; Wu, C.; Lin, T.; Ying, Y. Deep learning for vibrational spectral analysis: Recent progress and a practical guide. Anal. Chim. Acta 2019, 1081, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.-S.; Jean, N.; Hogan, C.A.; Blackmon, L.; Jeffrey, S.S.; Holodniy, M.; Banaei, N.; Saleh, A.A.E.; Ermon, S.; Dionne, J. Rapid identification of pathogenic bacteria using Raman spectroscopy and deep learning. Nat. Commun. 2019, 10, 4927. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Li, X.; Lu, W.; Li, H.; Fu, Y.V.; Liu, F. Analysis of Raman Spectra by Using Deep Learning Methods in the Identification of Marine Pathogens. Anal. Chem. 2021, 93, 11089–11098. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Han, D.; Liu, S.; Sun, X.; Ning, B.; Han, T.; Wang, J.; Gao, Z. Raman spectroscopy-based adversarial network combined with SVM for detection of foodborne pathogenic bacteria. Talanta 2022, 237, 122901. [Google Scholar] [CrossRef]
- Zhang, L.; Li, C.; Peng, D.; Yi, X.; He, S.; Liu, F.; Zheng, X.; Huang, W.E.; Zhao, L.; Huang, X. Raman spectroscopy and machine learning for the classification of breast cancers. Spectrochim. Acta A 2022, 264, 120300. [Google Scholar] [CrossRef]
- Klein, D.; Breuch, R.; von der Mark, S.; Wickleder, C.; Kaul, P. Detection of spoilage associated bacteria using Raman-microspectroscopy combined with multivariate statistical analysis. Talanta 2019, 196, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Wang, S.; Qiu, J.; Li, M.; Li, D.; Xu, D.; Li, D.; Liu, Q. Raman spectroscopy combined with machine learning for rapid detection of food-borne pathogens at the single-cell level. Talanta 2021, 226, 122195. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Xu, X.; Feng, S.; Zhang, H.; Xu, L.; Jiang, H.; Sun, B.; Meng, Y.; Chen, W. Rapid identification of Salmonella serovars by using Raman spectroscopy and machine learning algorithm. Talanta 2023, 253, 123807. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, J.; Ding, J.; Lin, Q.; Young, G.M.; Jiang, C. Rapid identification of live and dead Salmonella by surface-enhanced Raman spectroscopy combined with convolutional neural network. Vib. Spectrosc. 2022, 118, 103332. [Google Scholar] [CrossRef]
- Tang, J.W.; Li, J.Q.; Yin, X.C.; Xu, W.W.; Pan, Y.C.; Liu, Q.H.; Gu, B.; Zhang, X.; Wang, L. Rapid Discrimination of Clinically Important Pathogens Through Machine Learning Analysis of Surface Enhanced Raman Spectra. Front. Microbiol. 2022, 13, 843417. [Google Scholar] [CrossRef] [PubMed]
- Tewes, T.J.; Kerst, M.; Platte, F.; Bockmuhl, D.P. Raman Microscopic Identification of Microorganisms on Metal Surfaces via Support Vector Machines. Microorganisms 2022, 10, 556. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.; Li, H.; Cao, S.; Li, X.; Yin, H.; Li, Y.; Dong, X.; Zhang, X. Discrimination of periodontal pathogens using Raman spectroscopy combined with machine learning algorithms. J. Innov. Opt. Health Sci. 2022, 15, 2240001. [Google Scholar] [CrossRef]
- Sil, S.; Mukherjee, R.; Kumbhar, D.; Reghu, D.; Shrungar, D.; Kumar, N.S.; Singh, U.K.; Umapathy, S. Raman spectroscopy and artificial intelligence open up accurate detection of pathogens from DNA-based sub-species level classification. J. Raman Spectrosc. 2021, 52, 2648–2659. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Excitation Wavelengths | Application | Advantages | Disadvantages |
|---|---|---|---|
| UV (244 nm or 257 nm) | Bulk analysis |
|
|
| VIS (mainly 532 nm and 633 nm) | Single-cell analysis |
|
|
| NIR (785 nm or 1064 nm) | Bulk analysis |
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frempong, S.B.; Salbreiter, M.; Mostafapour, S.; Pistiki, A.; Bocklitz, T.W.; Rösch, P.; Popp, J. Illuminating the Tiny World: A Navigation Guide for Proper Raman Studies on Microorganisms. Molecules 2024, 29, 1077. https://doi.org/10.3390/molecules29051077
Frempong SB, Salbreiter M, Mostafapour S, Pistiki A, Bocklitz TW, Rösch P, Popp J. Illuminating the Tiny World: A Navigation Guide for Proper Raman Studies on Microorganisms. Molecules. 2024; 29(5):1077. https://doi.org/10.3390/molecules29051077
Chicago/Turabian StyleFrempong, Sandra Baaba, Markus Salbreiter, Sara Mostafapour, Aikaterini Pistiki, Thomas W. Bocklitz, Petra Rösch, and Jürgen Popp. 2024. "Illuminating the Tiny World: A Navigation Guide for Proper Raman Studies on Microorganisms" Molecules 29, no. 5: 1077. https://doi.org/10.3390/molecules29051077
APA StyleFrempong, S. B., Salbreiter, M., Mostafapour, S., Pistiki, A., Bocklitz, T. W., Rösch, P., & Popp, J. (2024). Illuminating the Tiny World: A Navigation Guide for Proper Raman Studies on Microorganisms. Molecules, 29(5), 1077. https://doi.org/10.3390/molecules29051077