
Chronological and Carbohydrate-Dependent Transformation of Fatty Acids in the Larvae of Black Soldier Fly Following Food Waste Treatment

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
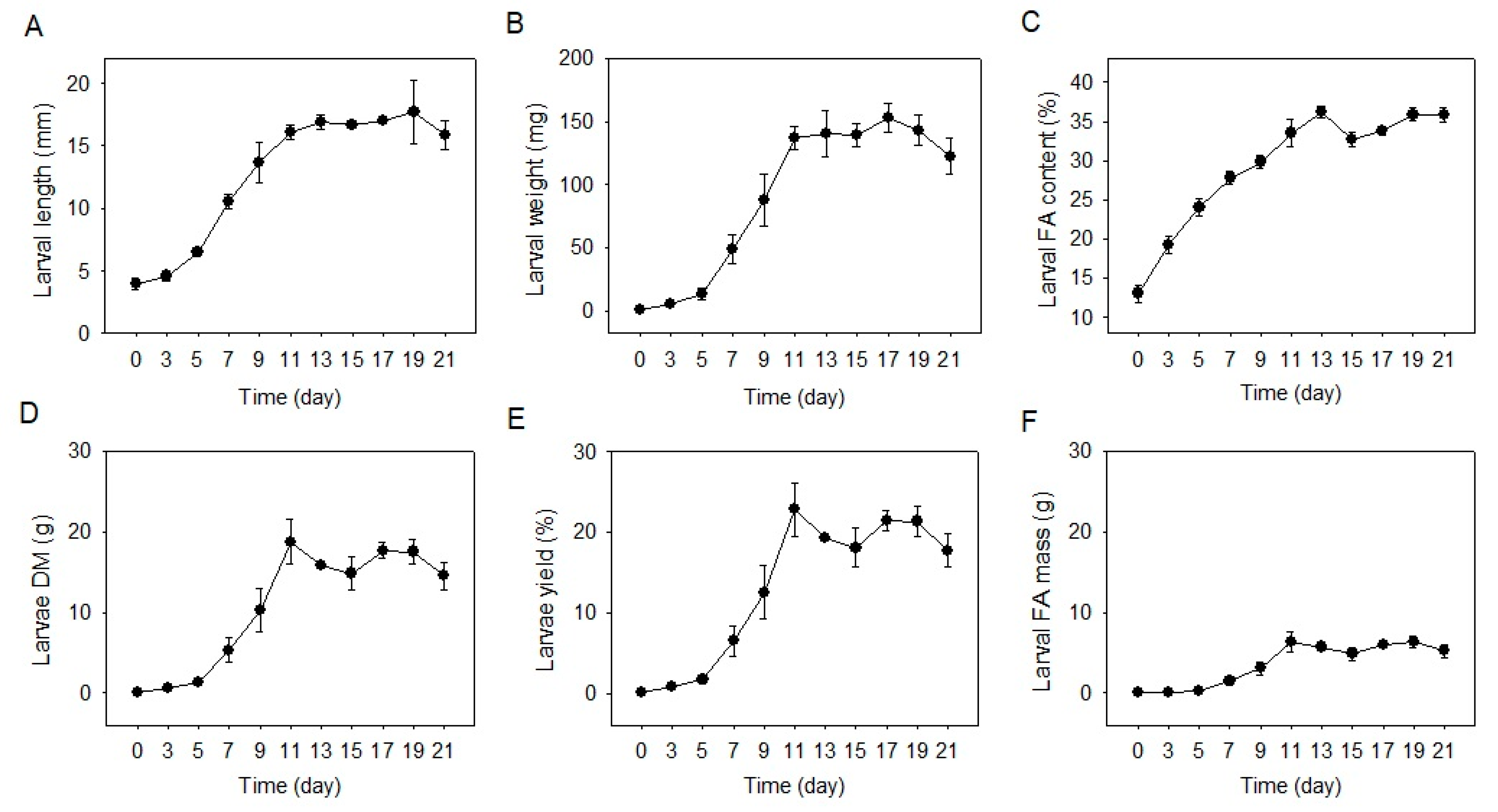
2.1. Larvae Development
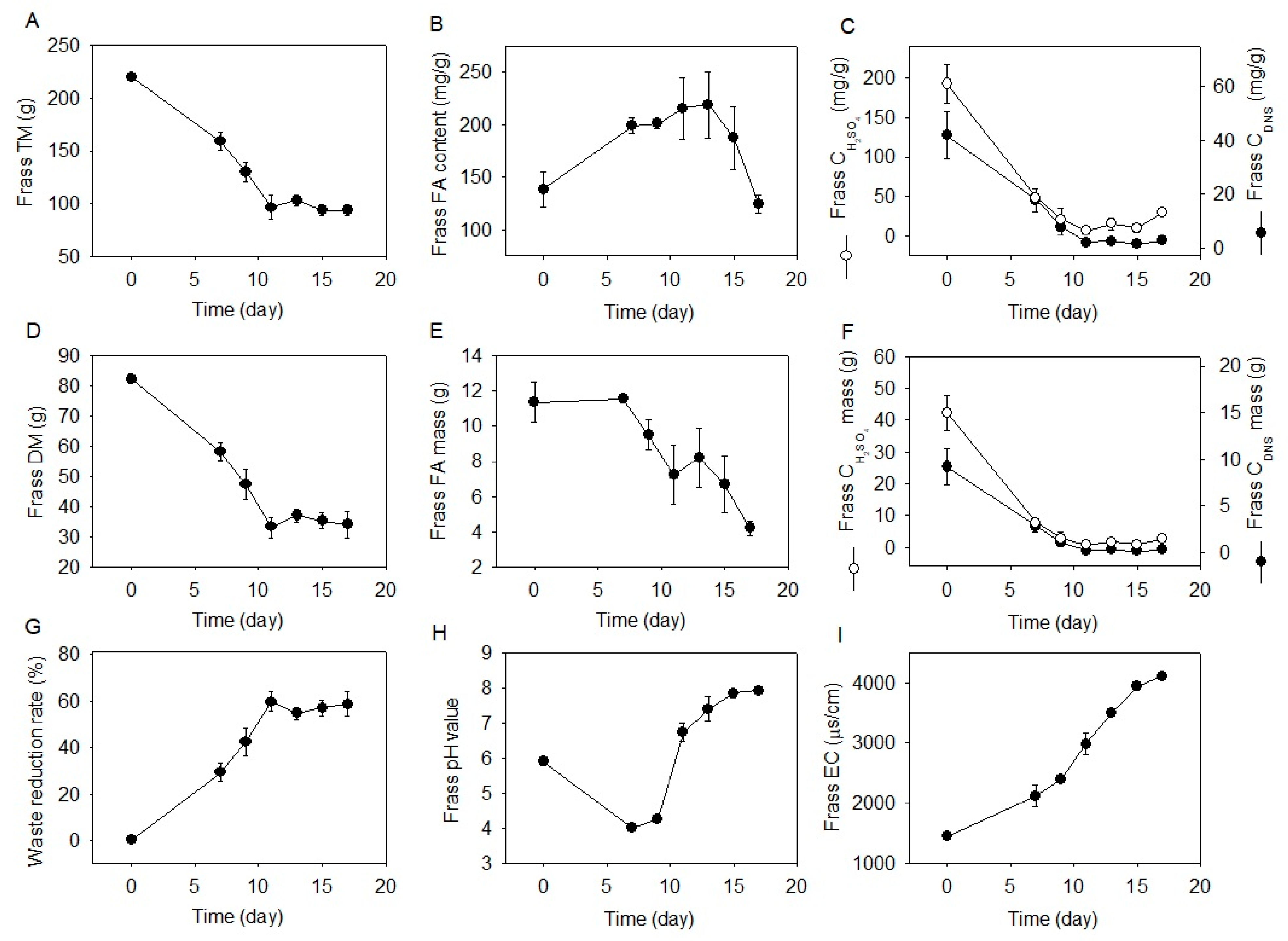
2.2. Waste Reduction and Frass Properties
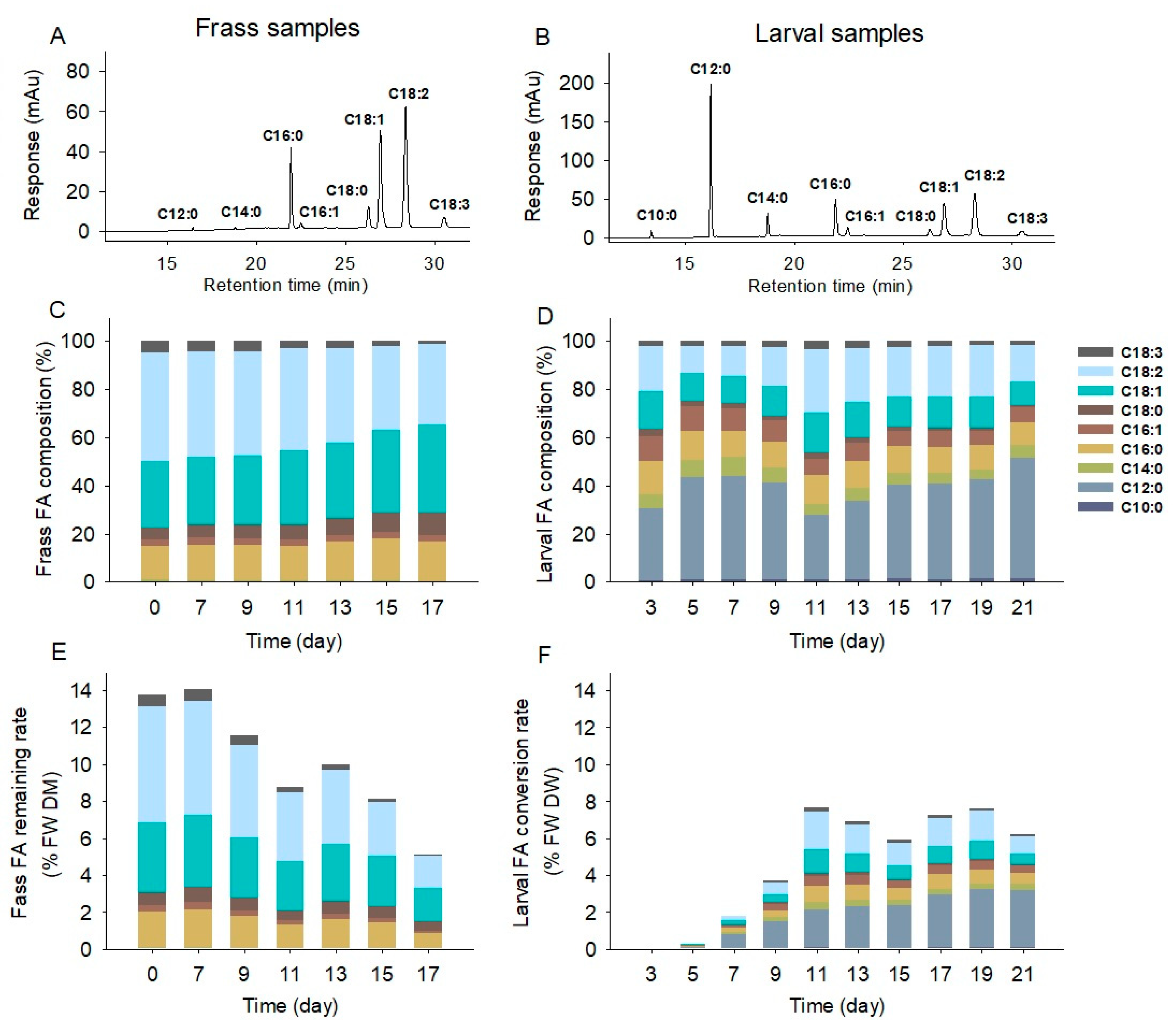
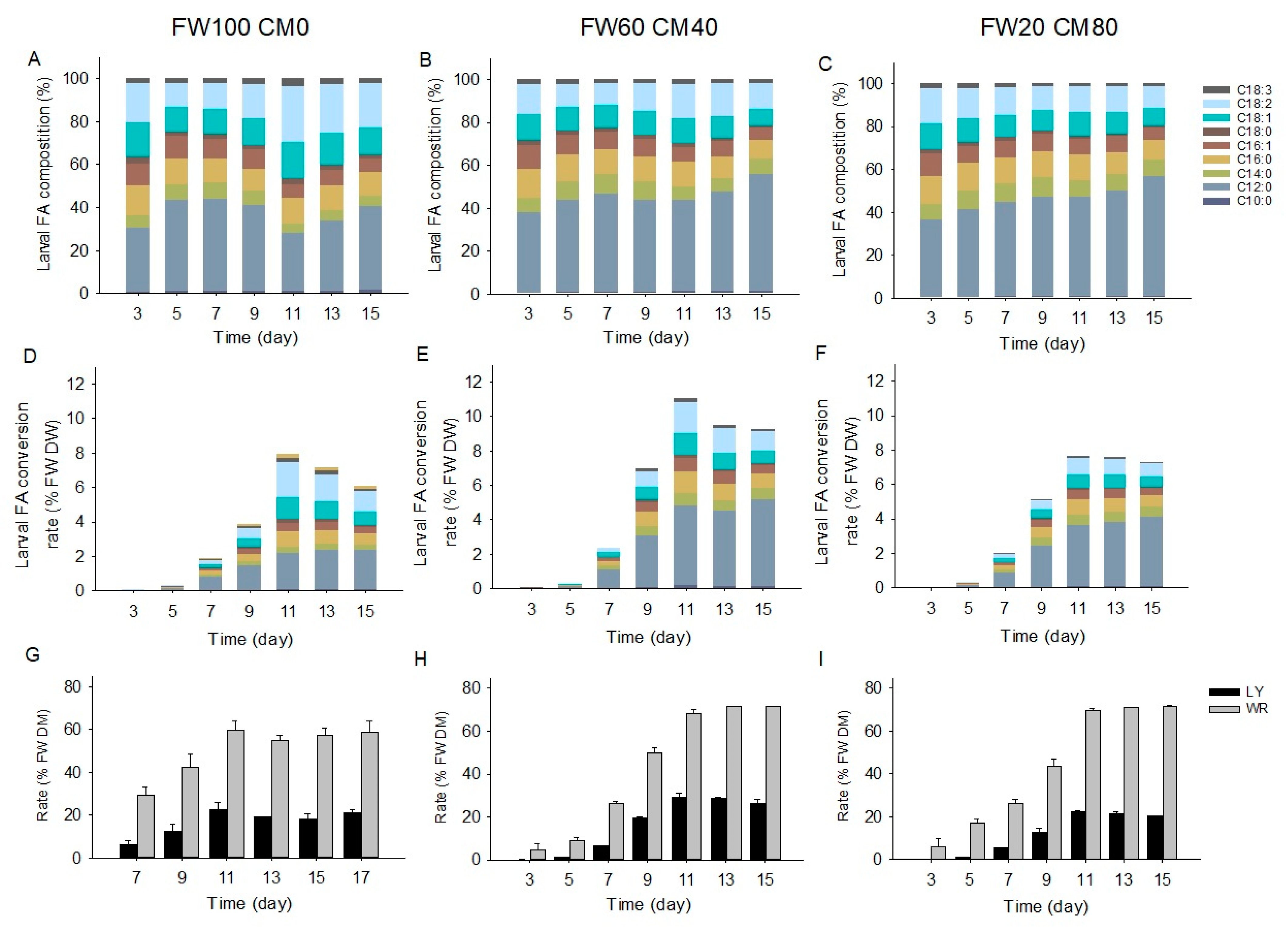
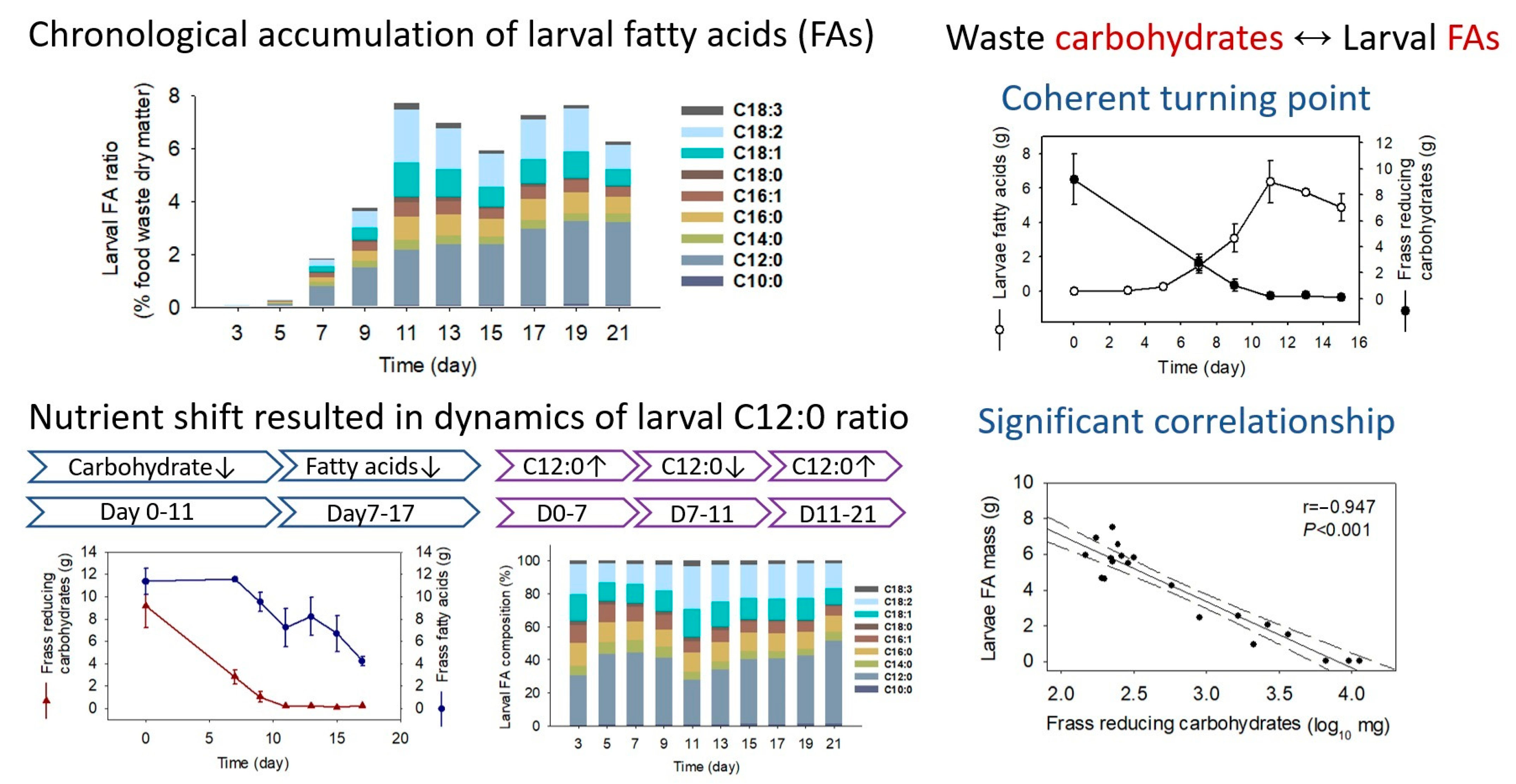
2.3. Fatty Acid Composition and Transformation
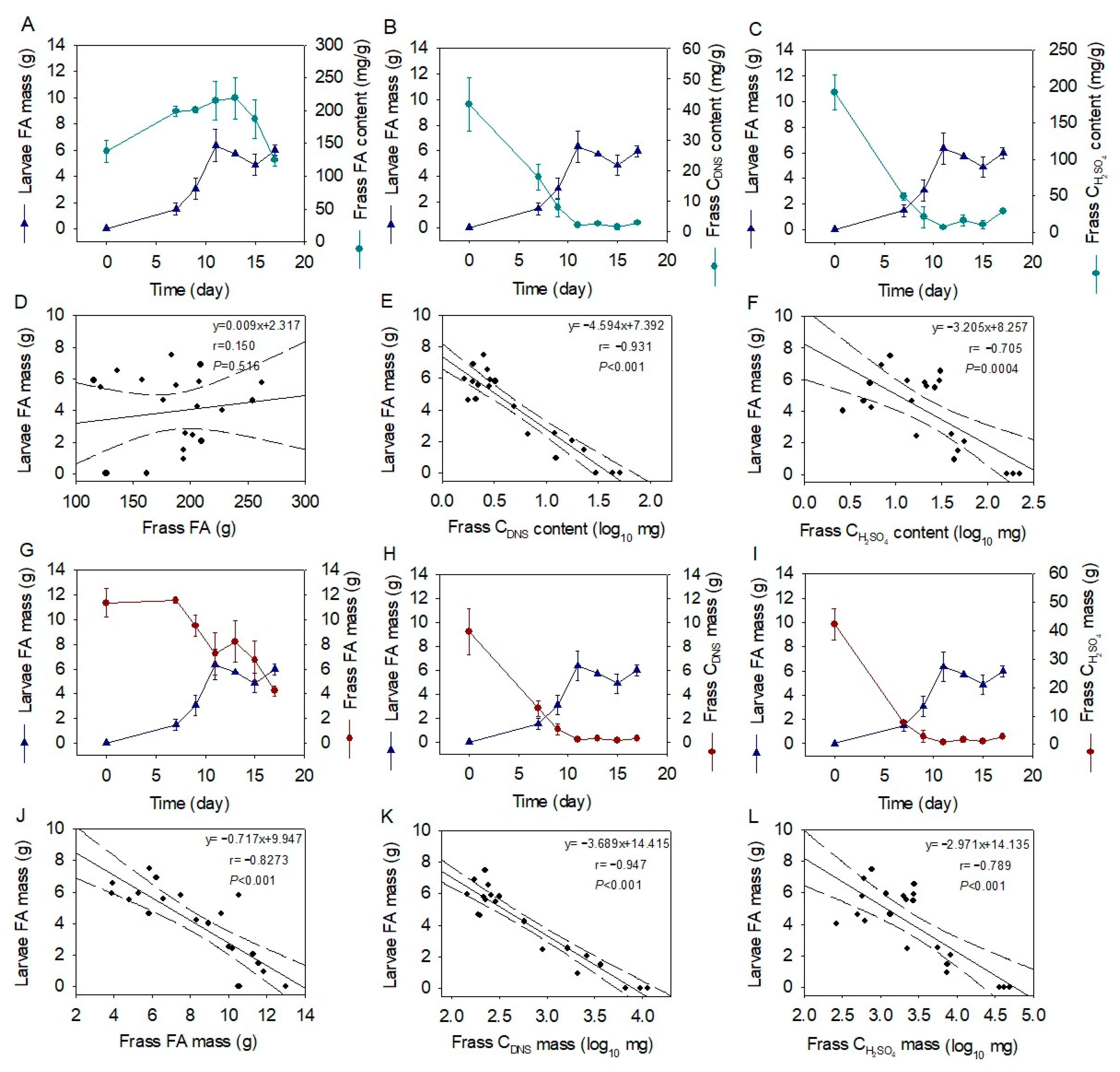
2.4. Correlations of Larval Fatty Acid and Frass Macronutrient
2.5. Fatty Acid Transformation of Adjusted Substrate
3. Materials and Methods
3.1. Materials and Reagents
3.2. Food Waste Treatment
3.3. Fatty Acid Analysis
3.4. Physiochemical Property Analysis
3.5. Conversion Efficiency Estimates
3.6. Statistical Analyses
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- United Nations Environmental Programme. Food Waste Index Report 2021. Nairobi. Available online: https://www.unep.org/resources/report/unep-food-waste-index-report-2021/ (accessed on 15 November 2022).
- National Bureau of Statistics. China Statistical Yearbook 2021. Available online: https://www.stats.gov.cn/tjsj/ndsj/ (accessed on 15 November 2022).
- Amrul, N.F.; Ahmad, I.K.; Basri, N.E.A.; Suja, F.; Jalil, N.A.A.; Azman, N.A. A Review of Organic Waste Treatment Using Black Soldier Fly (Hermetia illucens). Sustainability 2022, 14, 4565. [Google Scholar] [CrossRef]
- Fu, S.F.; Wang, D.H.; Xie, Z.; Zou, H.; Zheng, Y. Producing insect protein from food waste digestate via black soldier fly larvae cultivation: A promising choice for digestate disposal. Sci. Total Environ. 2022, 830, 154654. [Google Scholar] [CrossRef] [PubMed]
- Astuti, D.A.; Wiryawan, K.G. Black soldier fly as feed ingredient for ruminants. Anim. Biosci. 2022, 35, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Arabzadeh, G.; Delisle-Houde, M.; Tweddell, R.J.; Deschamps, M.H.; Dorais, M.; Lebeuf, Y.; Derome, N.; Vandenberg, G. Diet Composition Influences Growth Performance, Bioconversion of Black Soldier Fly Larvae: Agronomic Value and In Vitro Biofungicidal Activity of Derived Frass. Agronomy 2022, 12, 1765. [Google Scholar] [CrossRef]
- Addo, P.; Oduro-Kwarteng, S.; Gyasi, S.F.; Awuah, E. Bioconversion of municipal organic solid waste in to compost using Black Soldier Fly (Hermetia Illucens). Int. J. Recycl. Org. 2022, 11, 515–526. [Google Scholar]
- Basri, N.E.A.; Azman, N.A.; Ahmad, I.K.; Suja, F.; Jalil, N.A.A.; Amrul, N.F. Potential Applications of Frass Derived from Black Soldier Fly Larvae Treatment of Food Waste: A Review. Foods 2022, 11, 2664. [Google Scholar] [CrossRef]
- Sayed, W.A.A.; Alm-Eldin, M.M.S.; Hassan, R.S.; Sileem, T.M.; Rumpold, B.A. Recycling of Mediterranean Fruit Fly Rearing Waste by Black Soldier Fly, Hermetia illucens. Waste Biomass Valorization 2022, 14, 93–104. [Google Scholar] [CrossRef]
- Park, J.Y.; Jung, S.; Na, Y.G.; Jeon, C.H.; Cheon, H.Y.; Yun, E.Y.; Lee, S.H.; Kwon, E.E.; Kim, J.K. Biodiesel production from the black soldier fly larvae grown on food waste and its fuel property characterization as a potential transportation fuel. Environ. Eng. Res. 2022, 27, 200704. [Google Scholar] [CrossRef]
- Chemello, G.; Zarantoniello, M.; Randazzo, B.; Gioacchini, G.; Truzzi, C.; Cardinaletti, G.; Riolo, P.; Olivotto, I. Effects of black soldier fly (Hermetia illucens) enriched with Schizochytrium sp. on zebrafish (Danio rerio) reproductive performances. Aquaculture 2022, 550, 737853. [Google Scholar] [CrossRef]
- Franco, A.; Salvia, R.; Scieuzo, C.; Schmitt, E.; Russo, A.; Falabella, P. Lipids from Insects in Cosmetics and for Personal Care Products. Insects 2022, 13, 41. [Google Scholar] [CrossRef]
- Liu, X.; Chen, X.; Wang, H.; Yang, Q.Q.; Rehman, K.U.; Li, W.; Cai, M.M.; Li, Q.; Mazza, L.; Zhang, J.B.; et al. Dynamic changes of nutrient composition throughout the entire life cycle of black soldier fly. PLoS ONE 2017, 12, e0182601. [Google Scholar] [CrossRef]
- Zhu, Z.; Rehman, K.U.; Yu, Y.; Liu, X.; Wang, H.; Tomberlin, J.K.; Sze, S.H.; Cai, M.; Zhang, J.; Yu, Z.; et al. De novo transcriptome sequencing and analysis revealed the molecular basis of rapid fat accumulation by black soldier fly (Hermetia illucens, L.) for development of insectival biodiesel. Biotechnol. Biofuels 2019, 12, 194. [Google Scholar] [CrossRef]
- Li, W.; Li, M.S.; Zheng, L.Y.; Liu, Y.S.; Zhang, Y.L.; Yu, Z.N.; Ma, Z.H.; Li, Q. Simultaneous utilization of glucose and xylose for lipid accumulation in black soldier fly. Biotechnol. Biofuels 2015, 8, 117. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.B.; Kim, D.H.; Jeong, S.B.; Lee, J.W.; Kim, T.H.; Lee, H.G.; Lee, K.W. Black soldier fly larvae oil as an alternative fat source in broiler nutrition. Poult. Sci. 2020, 99, 3133–3143. [Google Scholar] [CrossRef] [PubMed]
- Abu Bakar, N.H.; Razak, S.A.; Taufek, N.M.; Alias, Z. Evaluation of black soldier fly (Hermetia illucens) prepupae oil as meal supplementation in diets for red hybrid tilapia (Oreochromis sp.). Int. J. Trop. Insect Sci. 2021, 41, 2093–2102. [Google Scholar] [CrossRef]
- Agbohessou, P.S.; Mandiki, S.N.M.; Gougbedji, A.; Megido, R.C.; Lima, L.M.W.; Cornet, V.; Lambert, J.; Purcaro, G.; Francis, F.; Laleye, P.A.; et al. Efficiency of fatty acid-enriched dipteran-based meal on husbandry, digestive activity and immunological responses of Nile tilapia Oreochromis niloticus juveniles. Aquaculture 2021, 545, 737193. [Google Scholar] [CrossRef]
- Kim, B.; Bang, H.T.; Jeong, J.Y.; Kim, M.; Kim, K.H.; Chun, J.L.; Ji, S.Y. Effects of Dietary Supplementation of Black Soldier Fly (Hermetia illucens) Larvae Oil on Broiler Health. J. Poult. Sci. 2021, 58, 222–229. [Google Scholar] [CrossRef]
- Mai, H.C.; Dao, N.D.; Lam, T.D.; Nguyen, B.V.; Nguyen, D.C.; Bach, L.G. Purification Process, Physicochemical Properties, and Fatty Acid Composition of Black Soldier Fly (Hermetia illucens Linnaeus) Larvae Oil. J. Am. Oil Chem. Soc. 2019, 96, 1303–1311. [Google Scholar] [CrossRef]
- Hoc, B.; Genva, M.; Fauconnier, M.L.; Lognay, G.; Francis, F.; Caparros Megido, R. About lipid metabolism in Hermetia illucens (L. 1758): On the origin of fatty acids in prepupae. Sci. Rep. 2020, 10, 11916. [Google Scholar] [CrossRef] [PubMed]
- Skrivanova, E.; Marounek, M.; Benda, V.; Brezina, P. Susceptibility of Escherichia coli, Salmonella sp and Clostridium perfringens to organic acids and monolaurin. Vet. Med. 2006, 51, 81–88. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Kao, M.C.; Fang, J.Y.; Zouboulis, C.C.; Zhang, L.F.; Gallo, R.L.; Huang, C.M. Antimicrobial Property of Lauric Acid Against Propionibacterium Acnes: Its Therapeutic Potential for Inflammatory Acne Vulgaris. J. Investig. Dermatol. 2009, 129, 2480–2488. [Google Scholar] [CrossRef] [PubMed]
- Park, K.M. Hydrophilic and lipophilic characteristics of non-fatty acid moieties: Significant factors affecting antibacterial activity of lauric acid esters. Food Sci. Biotechnol. 2018, 27, 1245. [Google Scholar] [CrossRef] [PubMed]
- Zeiger, K.; Popp, J.; Becker, A.; Hankel, J.; Visscher, C.; Klein, G.; Meemken, D. Lauric acid as feed additive—An approach to reducing Campylobacter spp. in broiler meat. PLoS ONE 2017, 12, e0175693. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Hu, Y.; Xu, D.W.; Cai, K.Y. Surface functionalization of titanium substrates with chitosan-lauric acid conjugate to enhance osteoblasts functions and inhibit bacteria adhesion. Colloids Surf. B-Biointerfaces 2014, 119, 115–125. [Google Scholar] [CrossRef]
- Saadoun, J.H.; Montevecchi, G.; Zanasi, L.; Bortolini, S.; Macavei, L.I.; Masino, F.; Maistrello, L.; Antonelli, A. Lipid profile and growth of black soldier flies (Hermetia illucens, Stratiomyidae) reared on by-products from different food chains. J. Sci. Food Agric. 2020, 100, 3648–3657. [Google Scholar] [CrossRef]
- Romano, N.; Fischer, H.; Kumar, V.; Francis, S.A.; Sinha, A.K. Productivity, conversion ability, and biochemical composition of black soldier fly (Hermetia illucens) larvae fed with sweet potato, spent coffee or dough. Int. J. Trop. Insect Sci. 2022, 42, 183–190. [Google Scholar] [CrossRef]
- Ewald, N.; Vidakovic, A.; Langeland, M.; Kiessling, A.; Sampels, S.; Lalander, C. Fatty acid composition of black soldier fly larvae (Hermetia illucens)—Possibilities and limitations for modification through diet. Waste Manag. 2020, 102, 40–47. [Google Scholar] [CrossRef]
- Cohn, Z.; Latty, T.; Abbas, A. Understanding dietary carbohydrates in black soldier fly larvae treatment of organic waste in the circular economy. Waste Manag. 2022, 137, 9–19. [Google Scholar] [CrossRef]
- Klammsteiner, T.; Walter, A.; Bogataj, T.; Heussler, C.D.; Stres, B.; Steiner, F.M.; Schlick-Steiner, B.C.; Insam, H. Impact of Processed Food (Canteen and Oil Wastes) on the Development of Black Soldier Fly (Hermetia illucens) Larvae and Their Gut Microbiome Functions. Front. Microbiol. 2021, 12, 619112. [Google Scholar] [CrossRef]
- Oonincx, D.; Laurent, S.; Veenenbos, M.E.; van Loon, J.J.A. Dietary enrichment of edible insects with omega 3 fatty acids. Insect Sci. 2020, 27, 500–509. [Google Scholar] [CrossRef]
- Li, X.C.; Dong, Y.W.; Sun, Q.X.; Tan, X.H.; You, C.H.; Huang, Y.H.; Zhou, M. Growth and Fatty Acid Composition of Black Soldier Fly Hermetia illucens (Diptera: Stratiomyidae) Larvae Are Influenced by Dietary Fat Sources and Levels. Animals 2022, 12, 486. [Google Scholar] [CrossRef] [PubMed]
- Lozica, L.; Starcevic, K.; Gavrilovic, A.; Masek, T. The influence of feeding media enriched with different oils on the fatty acid composition of the Black soldier fly (Hermetia illucens). Vet. Arh. 2022, 92, 291–300. [Google Scholar]
- Opare, L.O.; Holm, S.; Esperk, T. Temperature-modified density effects in the black soldier fly: Low larval density leads to large size, short development time and high fat content. J. Insects Food Feed. 2022, 8, 783–802. [Google Scholar] [CrossRef]
- Lalander, C.; Diener, S.; Zurbrugg, C.; Vinneras, B. Effects of feedstock on larval development and process efficiency in waste treatment with black soldier fly (Hermetia illucens). J. Clean. Prod. 2019, 208, 211–219. [Google Scholar] [CrossRef]
- Galassi, G.; Jucker, C.; Parma, P.; Lupi, D.; Crovetto, G.M.; Savoldelli, S.; Colombini, S. Impact of Agro-industrial Byproducts on Bioconversion, Chemical Composition, in vitro Digestibility, and Microbiota of the Black Soldier Fly (Diptera: Stratiomyidae) Larvae. J. Insect Sci. 2021, 21, 8. [Google Scholar] [CrossRef]
- Fischer, H.; Romano, N.; Sinha, A.K. Conversion of Spent Coffee and Donuts by Black Soldier Fly (Hermetia illucens) Larvae into Potential Resources for Animal and Plant Farming. Insects 2021, 12, 332. [Google Scholar] [CrossRef]
- Singh, A.; Srikanth, B.H.; Kumari, K. Determining the Black Soldier fly larvae performance for plant-based food waste reduction and the effect on Biomass yield (vol 130, pg 147, 2021). Waste Manag. 2022, 144, 122. [Google Scholar] [CrossRef]
- Diener, S.; Solano, N.M.S.; Gutierrez, F.R.; Zurbrugg, C.; Tockner, K. Biological Treatment of Municipal Organic Waste using Black Soldier Fly Larvae. Waste Biomass Valorization 2011, 2, 357–363. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, M.; Yang, X.; Ren, M.; Zhang, S.; Jin, N.; Liu, Y.; Lu, H.; Xu, J.; Xu, W. Effects of calcium mineral and microbial inoculum on the lipid conversion efficiency during food waste treatment by black soldier fly larvae. Acta Sci. Circumstantiae 2021, 41, 4973–4985. [Google Scholar]
- Montalva-Medel, M.; Ledger, T.; Ruz, G.A.; Goles, E. Lac Operon Boolean Models: Dynamical Robustness and Alternative Improvements. Mathematics 2021, 9, 600. [Google Scholar] [CrossRef]
- Li, Q.; Zheng, L.; Cai, H.; Garza, E.; Yu, Z.; Zhou, S. From organic waste to biodiesel: Black soldier fly, Hermetia illucens, makes it feasible. Fuel 2011, 90, 1545–1548. [Google Scholar] [CrossRef]
- Jin, N.; Liu, Y.; Zhang, S.; Sun, S.; Wu, M.; Dong, X.; Tong, H.; Xu, J.; Zhou, H.; Guan, S.; et al. C/N-Dependent Element Bioconversion Efficiency and Antimicrobial Protein Expression in Food Waste Treatment by Black Soldier Fly Larvae. Int. J. Mol. Sci. 2022, 23, 5036. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhou, X.; Yang, Q.; Zhang, C.; He, X.; Zhou, N. Content Comparative Analysis of Total Sugar, Soluble Polysaccharide and Reducing Sugarin Lycopus lucidus var. hirtus Regel from Different Habitats. Chin. Wild Plant Resour. 2020, 39, 23–27. [Google Scholar]
- Feng, Y.; Ding, Z.; Yang, X. The Detection of Total Sugar and Reducing Sugar Content in Traditional Chiffon Cakes. J. Xinjiang Vocat. Univ. 2020, 28, 78–80. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Liu, J.; He, J.; Lu, H.; Sun, S.; Ji, F.; Dong, X.; Bao, Y.; Xu, J.; He, G.; et al. Chronological and Carbohydrate-Dependent Transformation of Fatty Acids in the Larvae of Black Soldier Fly Following Food Waste Treatment. Molecules 2023, 28, 1903. https://doi.org/10.3390/molecules28041903
Liu Y, Liu J, He J, Lu H, Sun S, Ji F, Dong X, Bao Y, Xu J, He G, et al. Chronological and Carbohydrate-Dependent Transformation of Fatty Acids in the Larvae of Black Soldier Fly Following Food Waste Treatment. Molecules. 2023; 28(4):1903. https://doi.org/10.3390/molecules28041903
Chicago/Turabian StyleLiu, Yanxia, Junliang Liu, Jinwen He, Hongxu Lu, Shibo Sun, Fengyun Ji, Xiaoying Dong, Yongming Bao, Jianqiang Xu, Gaohong He, and et al. 2023. "Chronological and Carbohydrate-Dependent Transformation of Fatty Acids in the Larvae of Black Soldier Fly Following Food Waste Treatment" Molecules 28, no. 4: 1903. https://doi.org/10.3390/molecules28041903
APA StyleLiu, Y., Liu, J., He, J., Lu, H., Sun, S., Ji, F., Dong, X., Bao, Y., Xu, J., He, G., & Xu, W. (2023). Chronological and Carbohydrate-Dependent Transformation of Fatty Acids in the Larvae of Black Soldier Fly Following Food Waste Treatment. Molecules, 28(4), 1903. https://doi.org/10.3390/molecules28041903