
Within-Plant Variation in Rosmarinus officinalis L. Terpenes and Phenols and Their Antimicrobial Activity against the Rosemary Phytopathogens Alternaria alternata and Pseudomonas viridiflava

 ,
, 


Abstract
1. Introduction
2. Results
2.1. Terpenes
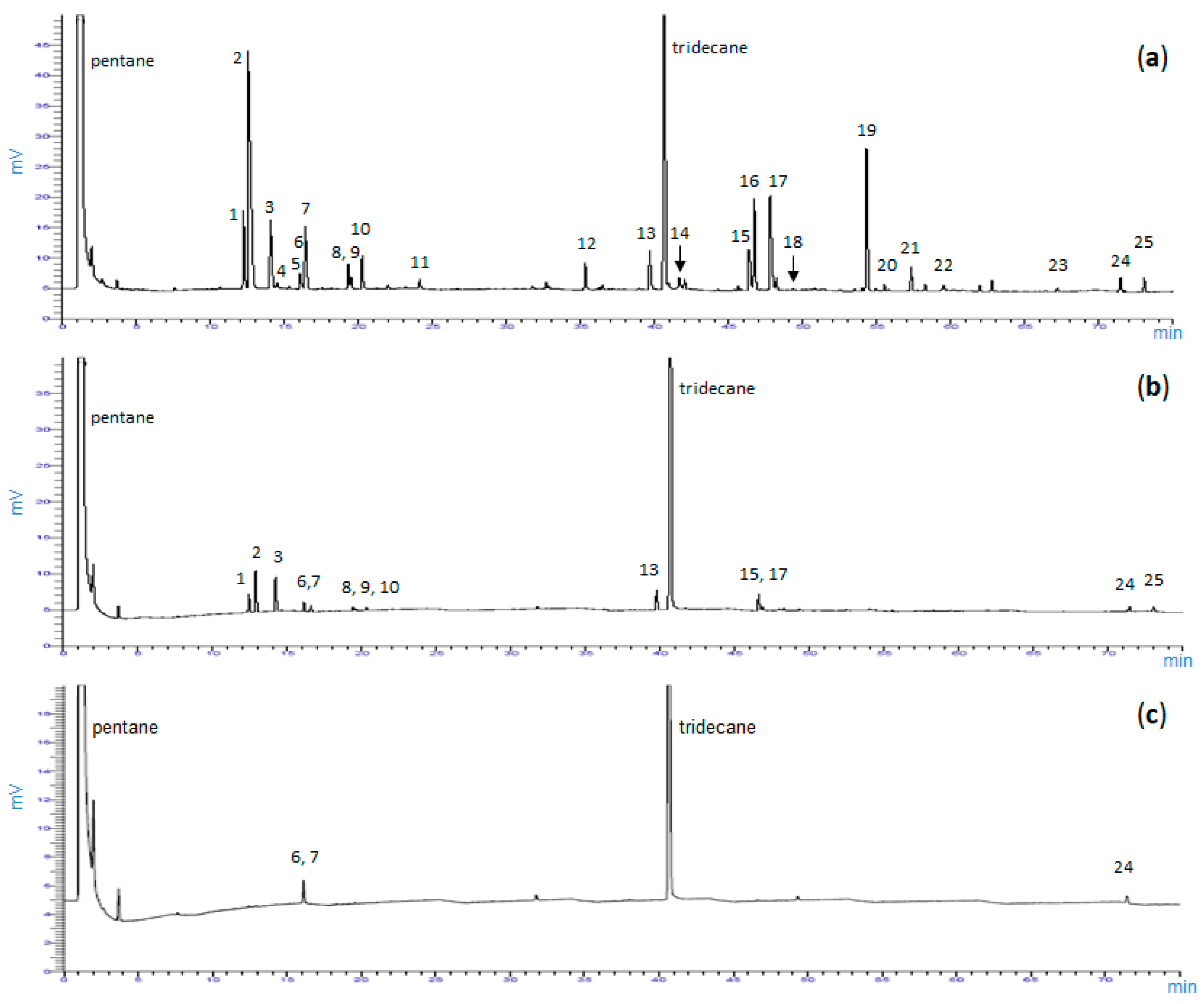
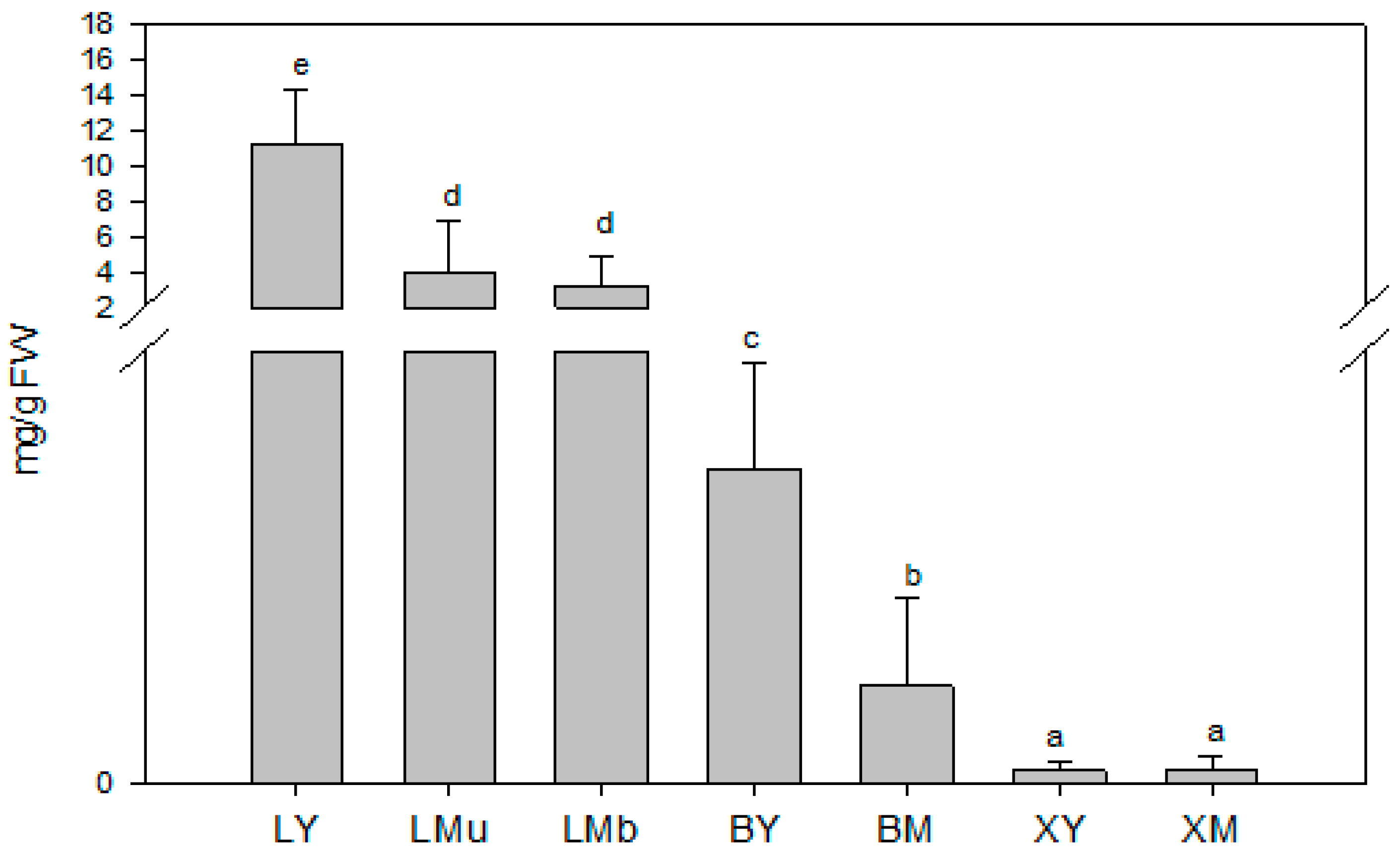
2.1.1. Total Concentrations of Terpenes (TCT)
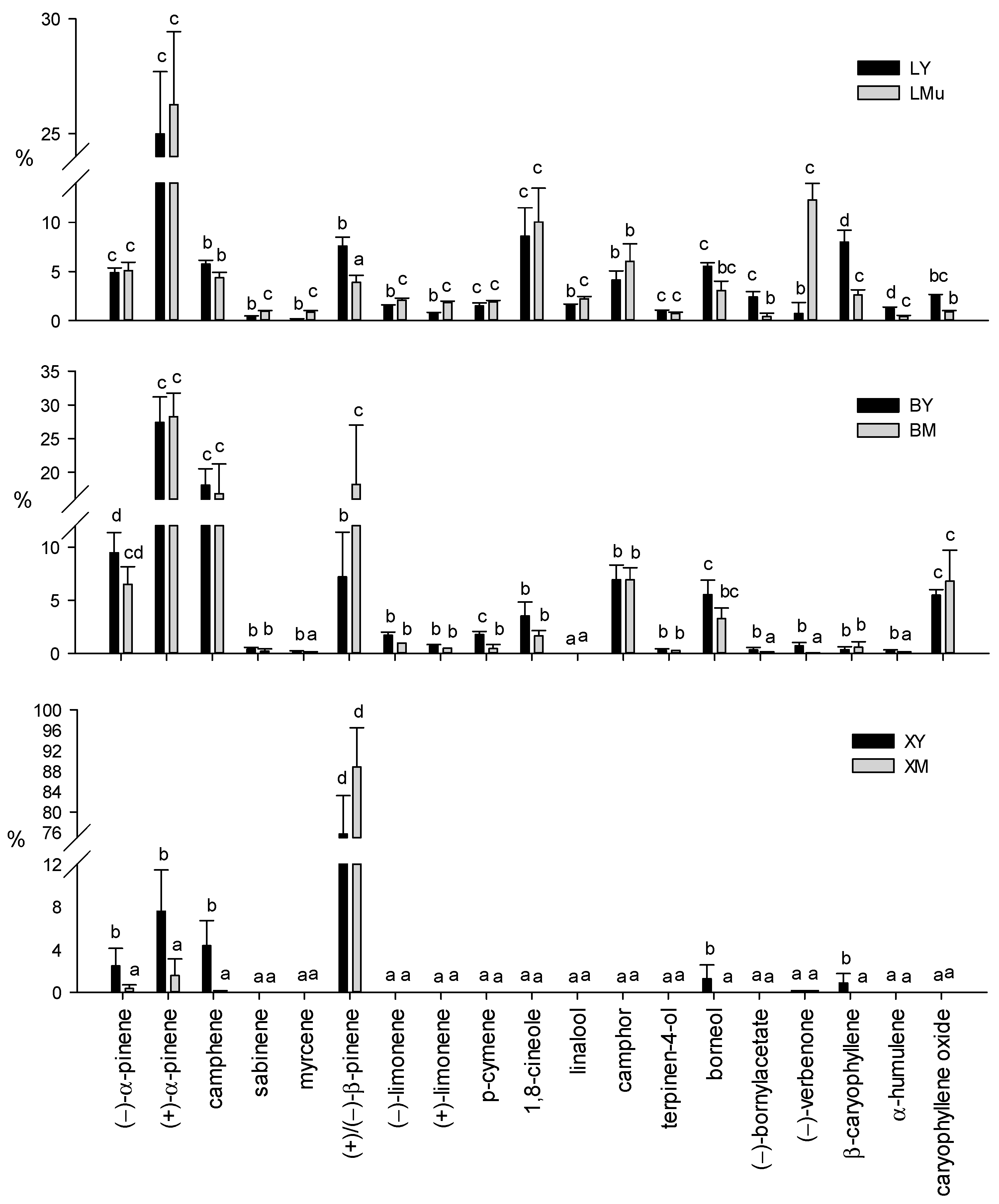
2.1.2. Relative Contents of Terpenes (Terpene Profiles)
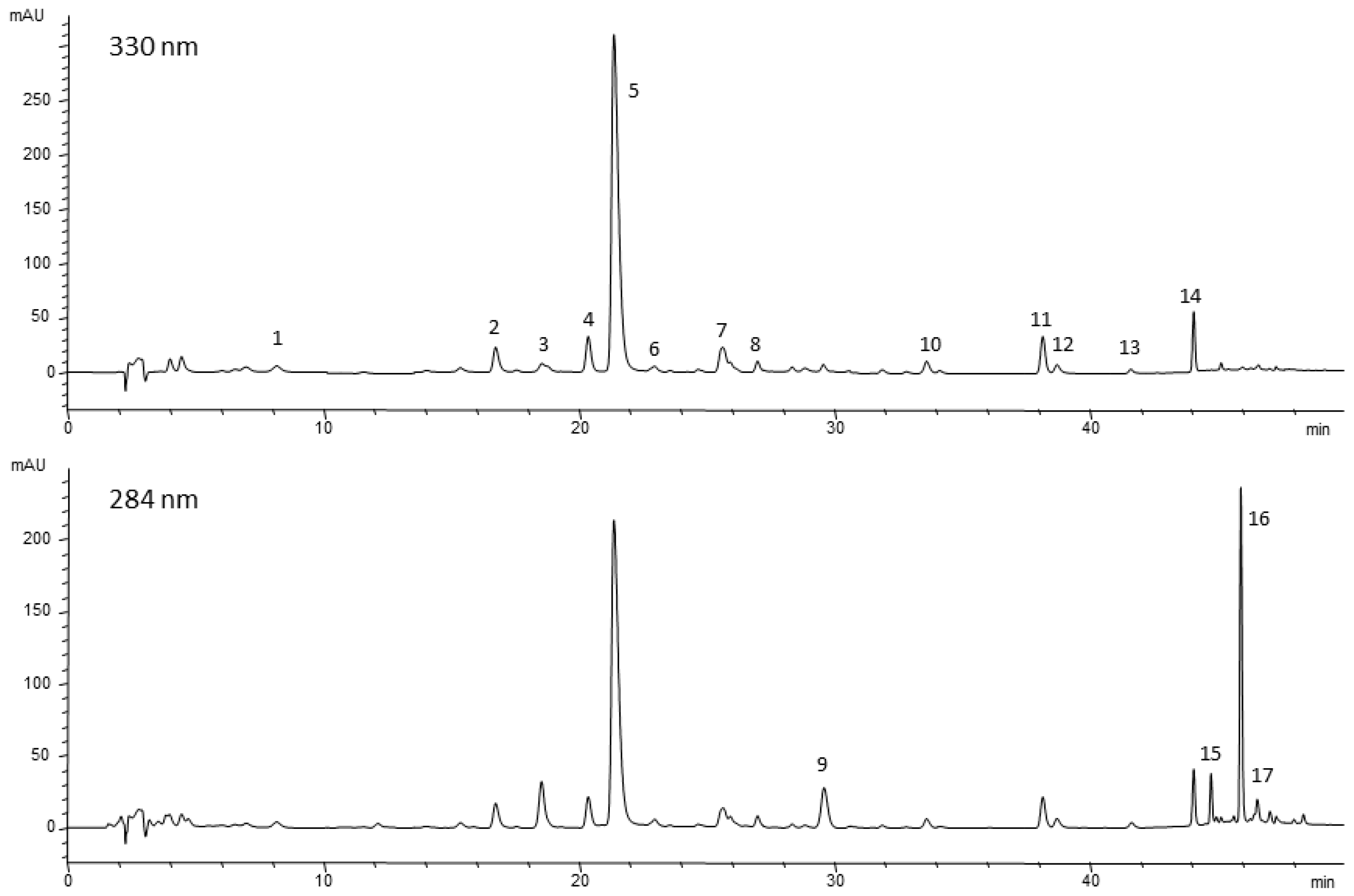
2.2. Phenolic Compounds
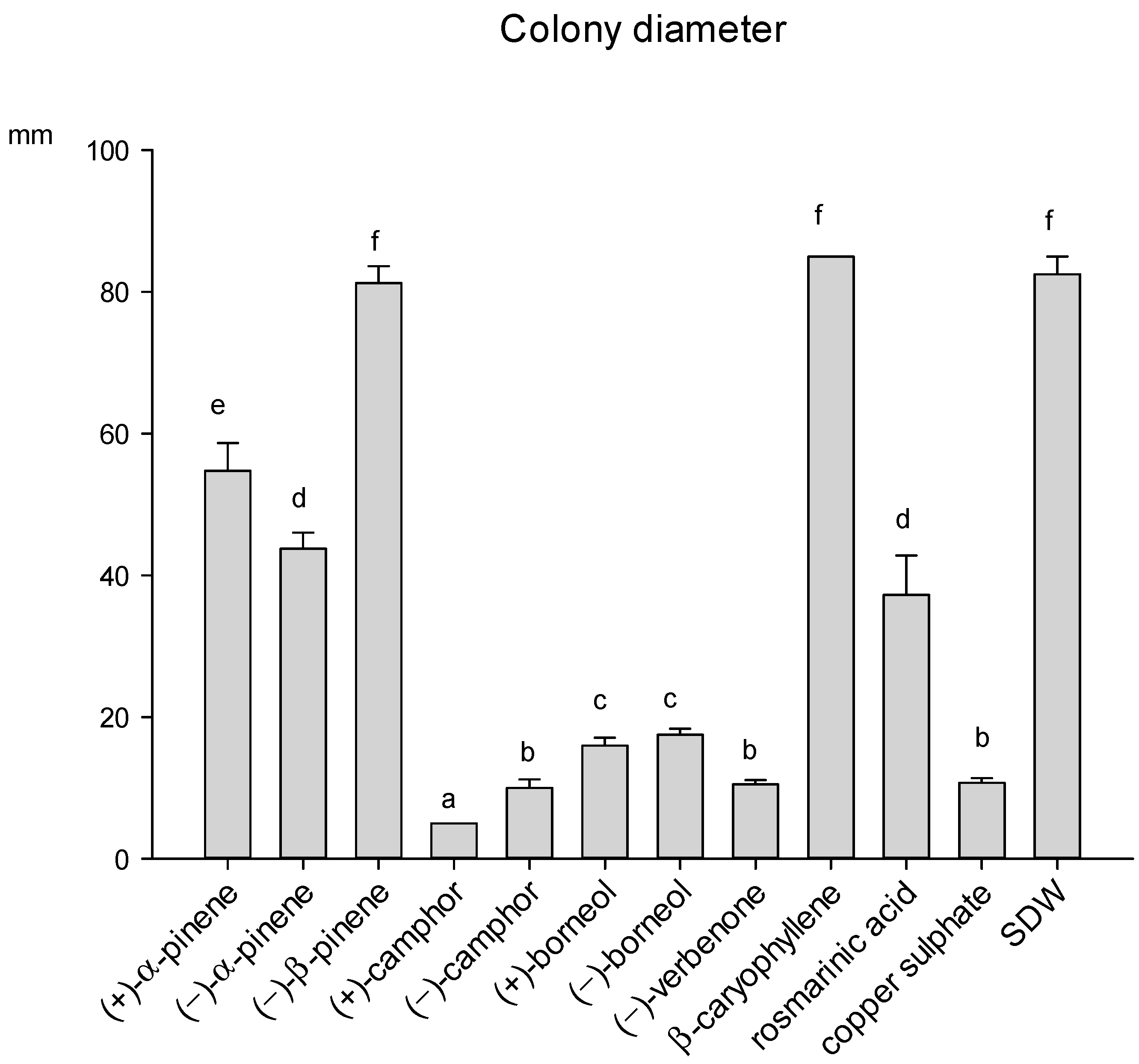
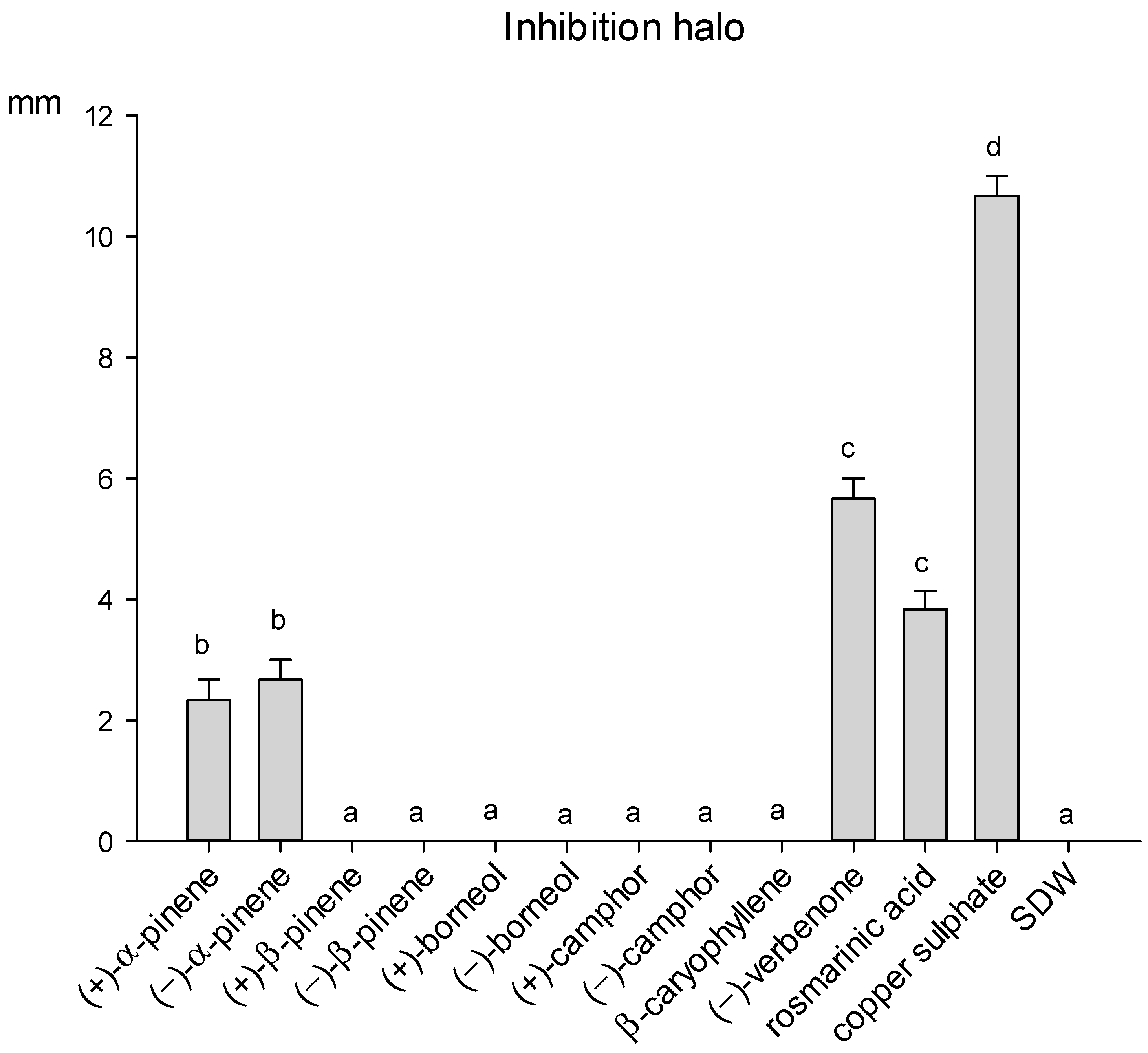
2.3. Effect of Selected Terpenes and Rosmarinic Acid on A. alternata and P. viridiflava
2.4. Determination of the Minimum Inhibitory Concentration (MIC)
3. Discussion
3.1. Terpenes
3.2. Phenolic Compounds
3.3. Antimicrobial Activity against A. alternata and P. viridiflava
4. Materials and Methods
4.1. Plant Material
4.2. Extraction of Terpene Volatile Compounds
4.3. Extraction of Phenolic Compounds
4.4. GC/FID and GC/MS Analyses
4.5. HPLC/DAD/ESI/MS Analyses
4.6. Microbial Strains and Antimicrobial Compounds
4.6.1. Antimicrobial Activity against A. alternata
4.6.2. Antimicrobial Activity against P. viridiflava
4.6.3. Determination of the Minimum Inhibitory Concentration (MIC)
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Moreno, S.; Sana, A.M.O.; Gaya, M.; Barni, M.V.; Castro, O.A.; Baren, C.V. Rosemary compounds as nutraceutical health products. In Food Additives; El-Samragy, Y., Ed.; IntechOpen Science: Rijeka, Croatia, 2012; p. 157. [Google Scholar]
- Ribeiro-Santos, R.; Carvalho-Costa, D.; Cavaleiro, C.; Costa, H.S.; Albuquerque, T.G.; Castilho, M.C.; Ramos, F.; Melo, N.R.; Sanches-Silva, A. A novel insight on an ancient aromatic plant: The rosemary (Rosmarinus officinalis L.). Trends Food Sci. Tech. 2015, 45, 355–368. [Google Scholar] [CrossRef]
- FDA. Subpart 182.10—Spices and other natural seasonings and flavorings. In Food and Drugs; Office of the Federal Register: Washington, DC, USA, 2014. [Google Scholar]
- EFSA. Scientific Opinion of the Panel on Food Additives, Flavourings, Processing Aids and Materials in Contact with Food on a request from the Commission on the use of rosemary extracts as a food additive. EFSA J. 2008, 721, 1–29. [Google Scholar]
- Zuzarte, M.R.; Dinis, A.M.; Cavaleiro, C.; Salgueiro, L.R.; Canhoto, J.M. Trichomes, essential oils and in vitro propagation of Lavandula pedunculata (Lamiaceae). Ind. Crop. Prod. 2010, 32, 580–587. [Google Scholar] [CrossRef]
- Angioni, A.; Barra, A.; Cereti, E.; Barile, D.; Coisson, J.D.; Arlorio, M.; Dessi, S.; Coroneo, V.; Cabras, P. Chemical composition, plant genetic differences, antimicrobial and antifungal activity investigation of the essential oil of Rosmarinus officinalis L. J. Agric. Food Chem. 2004, 52, 3530–3535. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, B.; Marques, A.; Ramos, C.; Neng, N.R.; Nogueira, J.M.F.; Saraiva, J.A.; Nunes, M.L. Chemical composition and antibacterial and antioxidant properties of commercial essential oils. Ind. Crop. Prod. 2013, 43, 587–595. [Google Scholar] [CrossRef]
- Okoh, O.O.; Sadimenko, A.P.; Afolayan, A.J. Comparative evaluation of the antibacterial activities of the essential oils of Rosmarinus officinalis L. obtained by hydrodistillation and solvent free microwave extraction methods. Food Chem. 2010, 120, 308–312. [Google Scholar] [CrossRef]
- Nieto, G.; Ros, G.; Castillo, J. Antioxidant and Antimicrobial Properties of Rosemary (Rosmarinus officinalis, L.): A Review. Medicines 2018, 5, 98. [Google Scholar] [CrossRef]
- Borges, R.S.; Ortiz, B.L.S.; Pereira, A.C.M.; Keita, H.; Carvalho, J.C.T. Rosmarinus officinalis essential oil: A review of its phytochemistry, anti-inflammatory activity, and mechanisms of action involved. J. Ethnopharmacol. 2019, 229, 29–45. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Camargo, A.D.; Herrero, M. Rosemary (Rosmarinus officinalis) as a functional ingredient: Recent scientific evidence. Curr. Opin. Food Sci. 2017, 14, 13–19. [Google Scholar] [CrossRef]
- Arranz, E.; Jaime, L.; Garcia-Risco, M.R.; Fornari, T.; Reglero, G.; Santoyo, S. Anti-inflammatory activity of rosemary extracts obtained by supercritical carbon dioxide enriched in carnosic acid and carnosol. Int. J. Food Sci. Tech. 2015, 50, 674–681. [Google Scholar] [CrossRef]
- Rocha, J.; Eduardo-Figueira, M.; Barateiro, A.; Fernandes, A.; Brites, D.; Bronze, R.; Duarte, C.M.M.; Serra, A.T.; Pinto, R.; Freitas, M.; et al. Anti-inflammatory effect of rosmarinic acid and an extract of rosmarinus officinalis in rat models of local and systemic inflammation. Basic Clin. Pharmacol. 2015, 116, 398–413. [Google Scholar] [CrossRef]
- Gonzalez-Vallinas, M.; Reglero, G.; de Molina, A.R. Rosemary (Rosmarinus officinalis L.) extract as a potential complementary agent in anticancer therapy. Nutr. Cancer 2015, 67, 1221–1229. [Google Scholar] [CrossRef] [PubMed]
- Petiwala, S.M.; Johnson, J.J. Diterpenes from rosemary (Rosmarinus officinalis): Defining their potential for anti-cancer activity. Cancer Lett. 2015, 367, 93–102. [Google Scholar] [CrossRef]
- Kontogianni, V.G.; Tomic, G.; Nikolic, I.; Nerantzaki, A.A.; Sayyad, N.; Stosic-Grujicic, S.; Stojanovic, I.; Gerothanassis, I.P.; Tzakos, A.G. Phytochemical profile of Rosmarinus officinalis and Salvia officinalis extracts and correlation to their antioxidant and anti-proliferative activity. Food Chem. 2013, 136, 120–129. [Google Scholar] [CrossRef]
- Lin, C.Y.; Tsai, C.W. Carnosic acid attenuates 6-Hydroxydopamine-induced neurotoxicity in SH-SY5Y cells by inducing autophagy through an enhanced interaction of parkin and Beclin1. Mol. Neurobiol. 2017, 54, 2813–2822. [Google Scholar] [CrossRef] [PubMed]
- Sedighi, R.; Zhao, Y.T.; Yerke, A.; Sang, S.M. Preventive and protective properties of rosemary (Rosmarinus officinalis L.) in obesity and diabetes mellitus of metabolic disorders: A brief review. Curr. Opin. Food Sci. 2015, 2, 58–70. [Google Scholar] [CrossRef]
- Hassani, F.V.; Shirani, K.; Hosseinzadeh, H. Rosemary (Rosmarinus officinalis) as a potential therapeutic plant in metabolic syndrome: A review. N S Arch. Pharmacol. 2016, 389, 931–949. [Google Scholar] [CrossRef]
- Jordan, M.J.; Lax, V.; Rota, M.C.; Loran, S.; Sotomayor, J.A. Effect of the phenological stage on the chemical composition, and antimicrobial and antioxidant properties of Rosmarinus officinalis L essential oil and its polyphenolic extract. Ind. Crop. Prod. 2013, 48, 144–152. [Google Scholar] [CrossRef]
- Yosr, Z.; Hnia, C.; Rim, T.; Mohamed, B. Changes in essential oil composition and phenolic fraction in Rosmarinus officinalis L. var. typicus Batt. organs during growth and incidence on the antioxidant activity. Ind. Crop. Prod. 2013, 43, 412–419. [Google Scholar] [CrossRef]
- Moore, B.D.; Andrew, R.L.; Kulheim, C.; Foley, W.J. Explaining intraspecific diversity in plant secondary metabolites in an ecological context. New Phytol. 2014, 201, 733–750. [Google Scholar] [CrossRef] [PubMed]
- Langenheim, J.H. Higher-Plant Terpenoids—A Phytocentric Overview of Their Ecological Roles. J. Chem. Ecol. 1994, 20, 1223–1280. [Google Scholar] [CrossRef]
- Gershenzon, J.; Dudareva, N. The function of terpene natural products in the natural world. Nat. Chem. Biol. 2007, 3, 408–414. [Google Scholar] [CrossRef]
- Mahdavi, A.; Moradi, P.; Mastinu, A. Variation in terpene profiles of thymus vulgaris in water deficit stress response. Molecules 2020, 25, 1091. [Google Scholar] [CrossRef]
- Barbero, F.; Maffei, M. Biodiversity and chemotaxonomic significance of specialized metabolites. In Plant Specialized Metabolism: Genomics, Biochemistry, and Biological Functions; Arimura, G.M.M., Ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 23–66. [Google Scholar]
- Larkov, O.; Mayer, A.M.; Ravid, U. Chiral monoterpenes as enantiotaxonomy indicators for the genera Origanum, Salvia, and Rosmarinus. Isr. J. Plant. Sci. 2010, 58, 203–209. [Google Scholar] [CrossRef]
- Bellumori, M.; Michelozzi, M.; Innocenti, M.; Congiu, F.; Cencetti, G.; Mulinacci, N. An innovative approach to the recovery of phenolic compounds and volatile terpenes from the same fresh foliar sample of Rosmarinus officinalis L. Talanta 2015, 131, 81–87. [Google Scholar] [CrossRef]
- Ravid, U.; Putievsky, E.; Katzir, I.; Lewinsohn, E.; Dudai, N. Identication of (1R)(+)-Verbenone in Essential Oils of Rosmarinus officinalis L. Flavour. Frag. J. 1997, 12, 109–112. [Google Scholar] [CrossRef]
- Tomi, K.; Kitao, M.; Konishi, N.; Murakami, H.; Matsumura, Y.; Hayashi, T. Enantioselective GC-MS analysis of volatile components from rosemary (Rosmarinus officinalis L.) essential oils and hydrosols. Biosci. Biotechnol. Biochem. 2016, 80, 840–847. [Google Scholar] [CrossRef] [PubMed]
- Satyal, P.; Jones, T.H.; Lopez, E.M.; McFeeters, R.L.; Ali, N.A.; Mansi, I.; Al-Kaf, A.G.; Setzer, W.N. Chemotypic characterization and biological activity of rosmarinus officinalis. Foods 2017, 6, 20. [Google Scholar] [CrossRef]
- Cutillas, A.B.; Carrasco, A.; Martinez-Gutierrez, R.; Tomas, V.; Tudela, J. Rosmarinus officinalis L. essential oils from Spain: Composition, antioxidant capacity, lipoxygenase and acetylcholinesterase inhibitory capacities, and antimicrobial activities. Plant Biosyst. 2018, 152, 1282–1292. [Google Scholar] [CrossRef]
- Martini, P.; Odasso, M.; Rapetti, S.; Repetto, L.; Rino, E.d.; Gullone, C. Reports of diseases on buttercup, sage and rosemary. Prot. delle Colt. 2009, 2, 109–110. [Google Scholar]
- Perello, A.; Bello, G.M. Foliar necrosis caused by Alternaria alternata on rosemary and Colletotrichum spp. on lavender, sage and marjoram. Investig. Agraria. Prod. Prot. Veg. 1995, 10, 275–281. [Google Scholar]
- Barton, K.E.; Edwards, K.F.; Koricheva, J. Shifts in woody plant defence syndromes during leaf development. Funct. Ecol. 2019, 33, 2095–2104. [Google Scholar] [CrossRef]
- Squillace, A.E. Analyses of monoterpenes of conifers by gas-liquid chromatography. In Modern Methods in Forest Genetics; Springer: Berlin/Heidelberg, Germany, 1976; pp. 120–157. [Google Scholar]
- Michelozzi, M.; Tognetti, R.; Maggino, F.; Radicati, M. Seasonal variations in monoterpene profiles and ecophysiological traits in Mediterranean pine species of group halepensis. Iforest 2008, 1, 65–74. [Google Scholar] [CrossRef]
- Cheng, S.S.; Lin, H.Y.; Chang, S.T. Chemical composition and antifungal activity of essential oils from different tissues of Japanese cedar (Cryptomeria japonica). J. Agric. Food. Chem. 2005, 53, 614–619. [Google Scholar] [CrossRef] [PubMed]
- Napoli, E.M.; Curcuruto, G.; Ruberto, G. Screening of the essential oil composition of wild Sicilian rosemary. Biochem. Syst. Ecol. 2010, 38, 659–670. [Google Scholar] [CrossRef]
- Yassaa, N.; Williams, J. Analysis of enantiomeric and non-enantiomeric monoterpenes in plant emissions using portable dynamic air sampling/solid-phase microextraction (PDAS-SPME) and chiral gas chromatography/mass spectrometry. Atmos. Environ. 2005, 39, 4875–4884. [Google Scholar] [CrossRef]
- Bozin, B.; Mlmica-Dukic, N.; Samojlik, I.; Jovin, E. Antimicrobial and antioxidant properties of rosemary and sage (Rosmarinus officinalis L. and Salvia officinalis L., lamiaceae) essential oils. J. Agric. Food Chem. 2007, 55, 7879–7885. [Google Scholar] [CrossRef]
- Del Campo, J.; Amiot, M.J.; Nguyen-The, C. Antimicrobial effect of rosemary extracts. J. Food Protect. 2000, 63, 1359–1368. [Google Scholar] [CrossRef]
- Hussein, K.A.; Lee, Y.D.; Joo, J.H. Effect of Rosemary Essential Oil and Trichoderma koningiopsis T-403 VOCs on Pathogenic Fungi Responsible for Ginseng Root Rot Disease. J. Microbiol. Biotechn. 2020, 30, 1018–1026. [Google Scholar] [CrossRef] [PubMed]
- van Bruggen, A.H.C.; Finckh, M.R. Plant Diseases and Management Approaches in Organic Farming Systems. Annu. Rev. Phytopathol. 2016, 54, 25–54. [Google Scholar] [CrossRef]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential oils in food preservation: Mode of action, synergies, and interactions with food matrix components. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef]
- Fowler, Z.L.; Baron, C.M.; Panepinto, J.C.; Koffas, M.A.G. Melanization of flavonoids by fungal and bacterial laccases. Yeast 2011, 28, 181–188. [Google Scholar] [CrossRef]
- Mazzeo, P.P.; Carraro, C.; Monica, A.; Capucci, D.; Pelagatti, P.; Bianchi, F.; Agazzi, S.; Careri, M.; Raio, A.; Carta, M.; et al. Designing a palette of cocrystals based on essential oil constituents for agricultural applications. ACS Sustain. Chem. Eng. 2019, 7, 17929–17940. [Google Scholar] [CrossRef]
- Mulinacci, N.; Innocenti, M.; Bellumori, M.; Giaccherini, C.; Martini, V.; Michelozzi, M. Storage method, drying processes and extraction procedures strongly affect the phenolic fraction of rosemary leaves: An HPLC/DAD/MS study. Talanta 2011, 85, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Bellumori, M.; Marzia, I.; Binello, A.; Boffa, L.; Mulinacci, N.; Cravotto, G. Selective recovery of rosmarinic and carnosic acids from rosemary leaves under ultrasound- and microwave-assisted extraction procedures. C. R. Chim. 2016, 19, 699–706. [Google Scholar] [CrossRef]
- Davis, W.W.; Stout, T.R. Disc plate method of microbiological antibiotic assay. I. Factors influencing variability and error. Appl. Microbiol. 1971, 22, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.H.A.; Cutter, C.N. Development and evaluation of pullulan-based composite antimicrobial films (CAF) incorporated with nisin, thymol and lauric arginate to reduce foodborne pathogens associated with muscle foods. Int. J. Food Microbiol. 2020, 320. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LY | LMu | ||||
|---|---|---|---|---|---|
| mg/g FW | SE | mg/g FW | SE | ||
| Isoorientin (luteolin 6-C-glu) | 0.63 b | 0.05 | 0.42 a | 0.01 | * |
| Homoplantaginin (hispidulin 7-O-glu) | 0.62 b | 0.04 | 0.35 a | 0.05 | * |
| Rosmarinic acid | 5.24 | 0.79 | 3.32 | 0.67 | NS |
| Isoscutellarein 7-O-glu | 0.6 | 0.05 | 0.51 | 0.05 | NS |
| Flavonoid 1 $ | 0.48 | 0.02 | 0.51 | 0.04 | NS |
| Cirsimaritin | 0.67 a | 0.04 | 0.98 b | 0.1 | * |
| Flavonoid 2 $ | 0.54 | 0.03 | 0.49 | 0.03 | NS |
| Genkwanin | 0.46 a | 0.02 | 0.92 b | 0.13 | * |
| 4′-methoxytectochrysin | 0.62 | 0.03 | 0.74 | 0.08 | NS |
| Carnosol | 12.87 | 3.78 | 8.27 | 4.01 | NS |
| Carnosic acid | 28.55 | 4.68 | 17.41 | 1.51 | NS |
| Methyl carnosic acid | 2.07 | 0.24 | 1.91 | 0.43 | NS |
| Total flavonoids | 4.62 | 0.24 | 4.92 | 0.34 | NS |
| Total terpenoids | 43.49 | 8.2 | 27.59 | 4.05 | NS |
| Total phenols | 53.35 | 8.82 | 35.83 | 4.82 | NS |
| A. alternata | P. viridiflava | |||
|---|---|---|---|---|
| V.R. | S.R. | V.R. | Resazurin | |
| (+)-α-pinene | 0.625 | 0.625 | 1.25 | 1.25 |
| (−)-β-pinene | 0.313 | 0.313 | n.i. | n.i. |
| (−)-verbenone | 0.625 | 0.625 | 1.25 | 1.25 |
| rosmarinic acid | n.d. | 1.25 | 1.25 | 1.25 |
| copper sulphate | 0.078 | 0.078 | 0.625 | 0.313 |
| Borneol | (+)-Camphor | (−)-Camphor | |
|---|---|---|---|
| SDW | 6.4 d | 6.4 c | 6.4 c |
| 5% | 3.8 a | 4.2 a | 3.7 a |
| 2.5% | 4.5 b | 4.5 ab | 4.9 b |
| 1.25% | 5.2 c | 4.9 b | 5.1 b |
| 0.625% | 6.2 d | 5.0 b | 5.1 b |
| 0.313% | 6.2 d | 6.2 c | 6.6 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellumori, M.; Innocenti, M.; Congiu, F.; Cencetti, G.; Raio, A.; Menicucci, F.; Mulinacci, N.; Michelozzi, M. Within-Plant Variation in Rosmarinus officinalis L. Terpenes and Phenols and Their Antimicrobial Activity against the Rosemary Phytopathogens Alternaria alternata and Pseudomonas viridiflava. Molecules 2021, 26, 3425. https://doi.org/10.3390/molecules26113425
Bellumori M, Innocenti M, Congiu F, Cencetti G, Raio A, Menicucci F, Mulinacci N, Michelozzi M. Within-Plant Variation in Rosmarinus officinalis L. Terpenes and Phenols and Their Antimicrobial Activity against the Rosemary Phytopathogens Alternaria alternata and Pseudomonas viridiflava. Molecules. 2021; 26(11):3425. https://doi.org/10.3390/molecules26113425
Chicago/Turabian StyleBellumori, Maria, Marzia Innocenti, Federica Congiu, Gabriele Cencetti, Aida Raio, Felicia Menicucci, Nadia Mulinacci, and Marco Michelozzi. 2021. "Within-Plant Variation in Rosmarinus officinalis L. Terpenes and Phenols and Their Antimicrobial Activity against the Rosemary Phytopathogens Alternaria alternata and Pseudomonas viridiflava" Molecules 26, no. 11: 3425. https://doi.org/10.3390/molecules26113425
APA StyleBellumori, M., Innocenti, M., Congiu, F., Cencetti, G., Raio, A., Menicucci, F., Mulinacci, N., & Michelozzi, M. (2021). Within-Plant Variation in Rosmarinus officinalis L. Terpenes and Phenols and Their Antimicrobial Activity against the Rosemary Phytopathogens Alternaria alternata and Pseudomonas viridiflava. Molecules, 26(11), 3425. https://doi.org/10.3390/molecules26113425