Mulberry Fruit Cultivar ‘Chiang Mai’ Prevents Beta-Amyloid Toxicity in PC12 Neuronal Cells and in a Drosophila Model of Alzheimer’s Disease


 and
and
Abstract
:1. Introduction
2. Results
2.1. Extraction Optimization of Morus cf. nigra ‘Chiang Mai’ (MNCM)
2.2. Antioxidant Activities of MNCM Extract
2.3. Phytochemical Analysis
2.4. MNCM Extract Inhibits Cholinesterase and BACE-1 Activities in Vitro
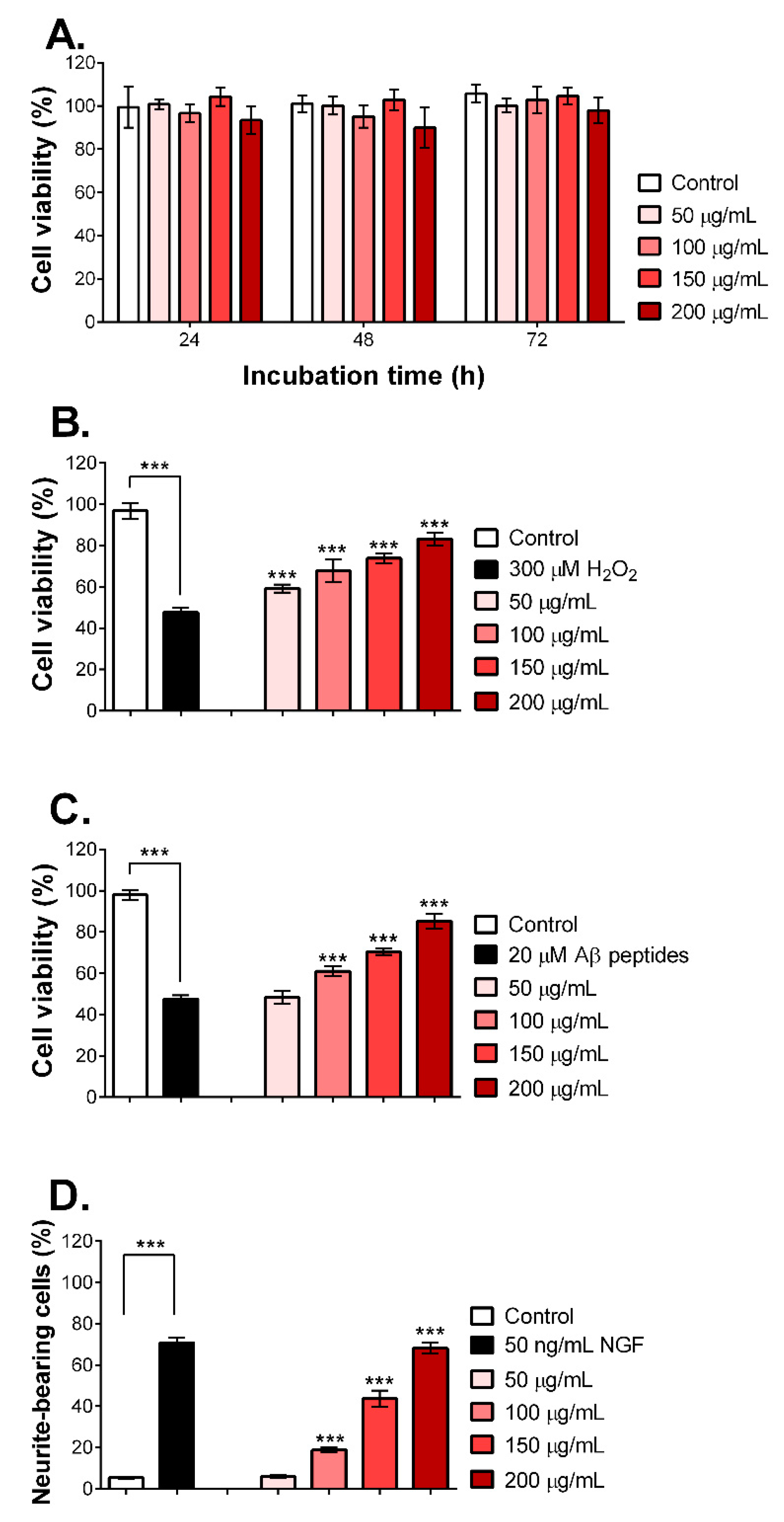
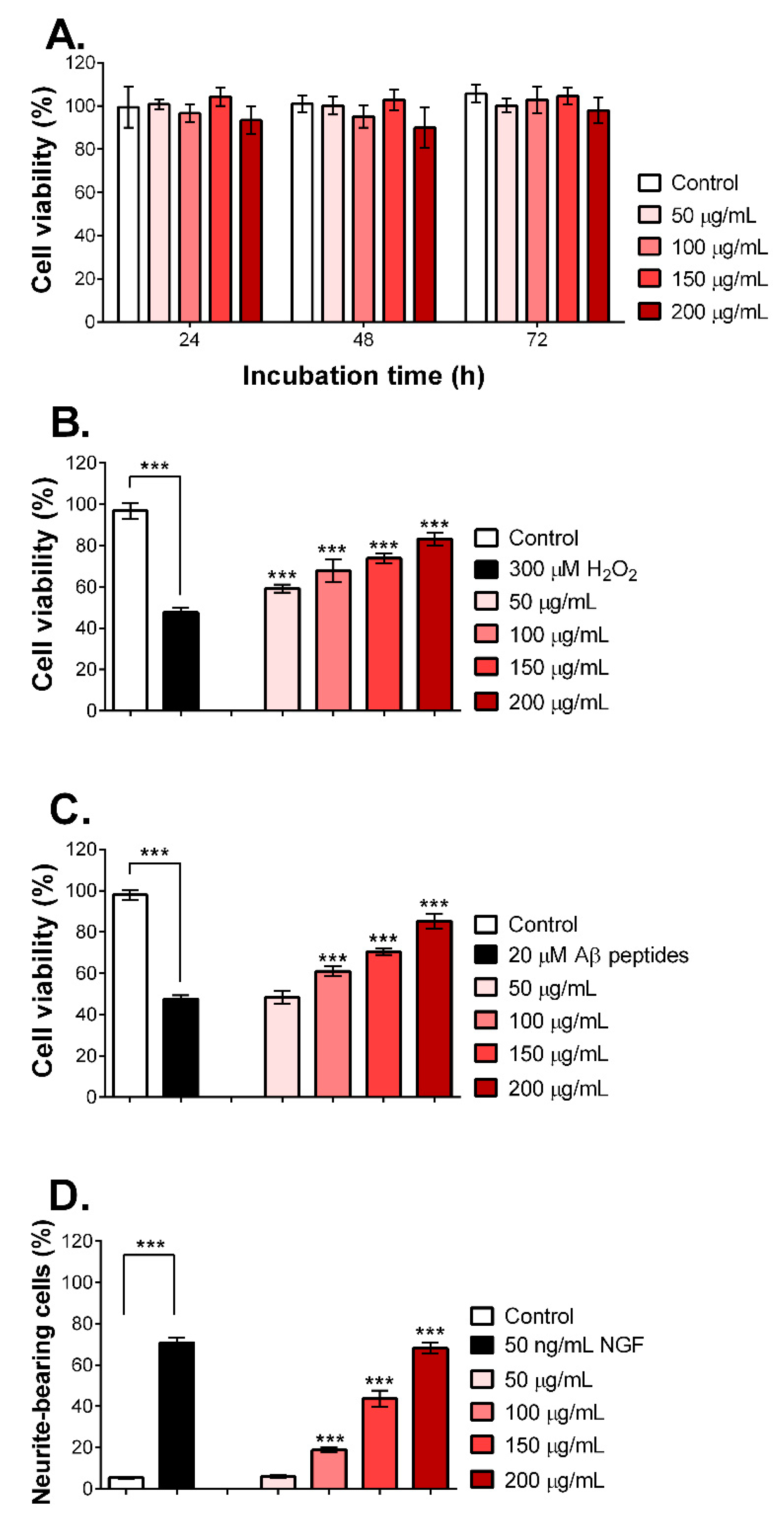
2.5. MNCM Extract Prevents Aβ Peptide-Induced Toxicity and Promotes Neurite Outgrowth
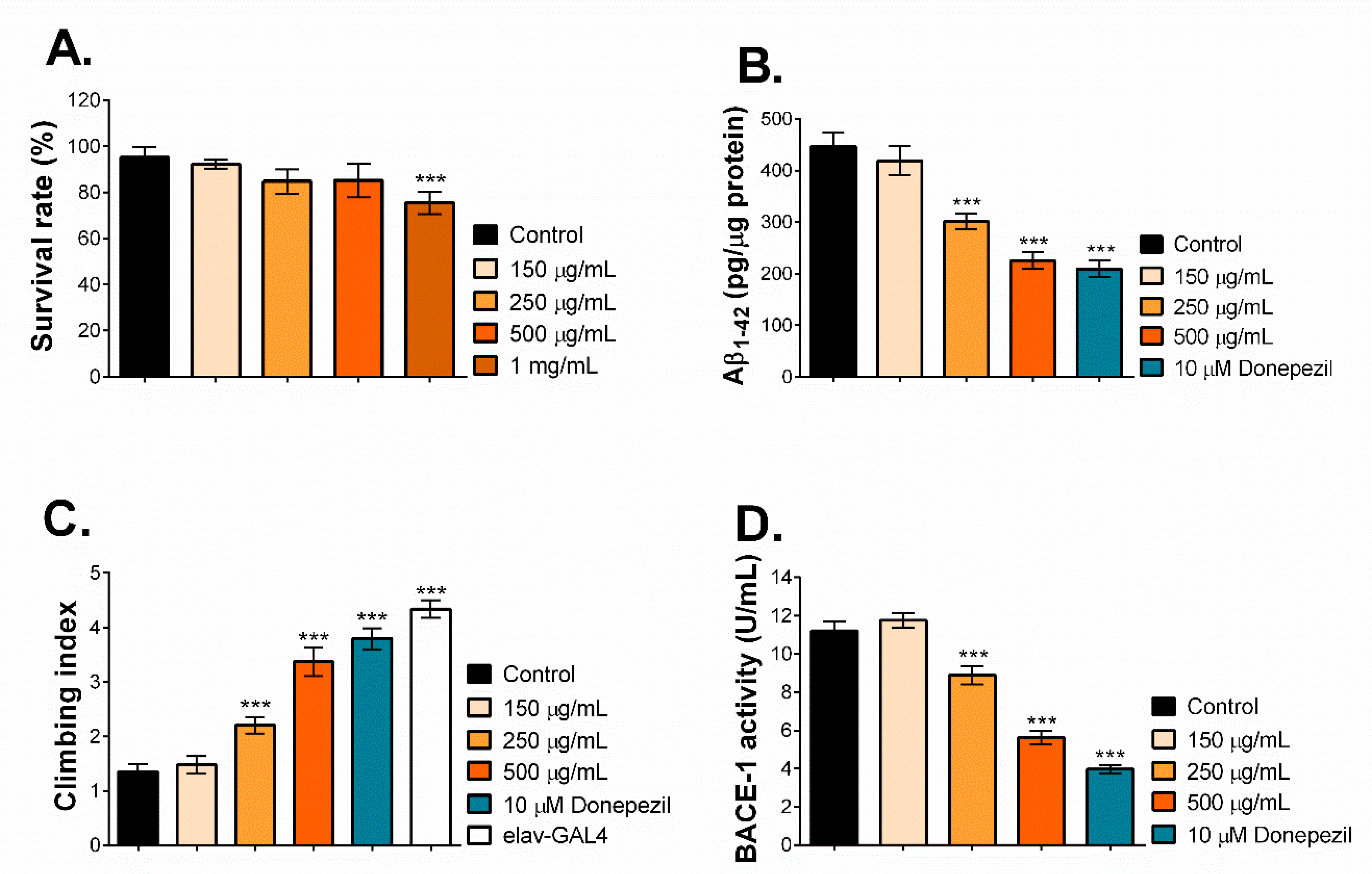
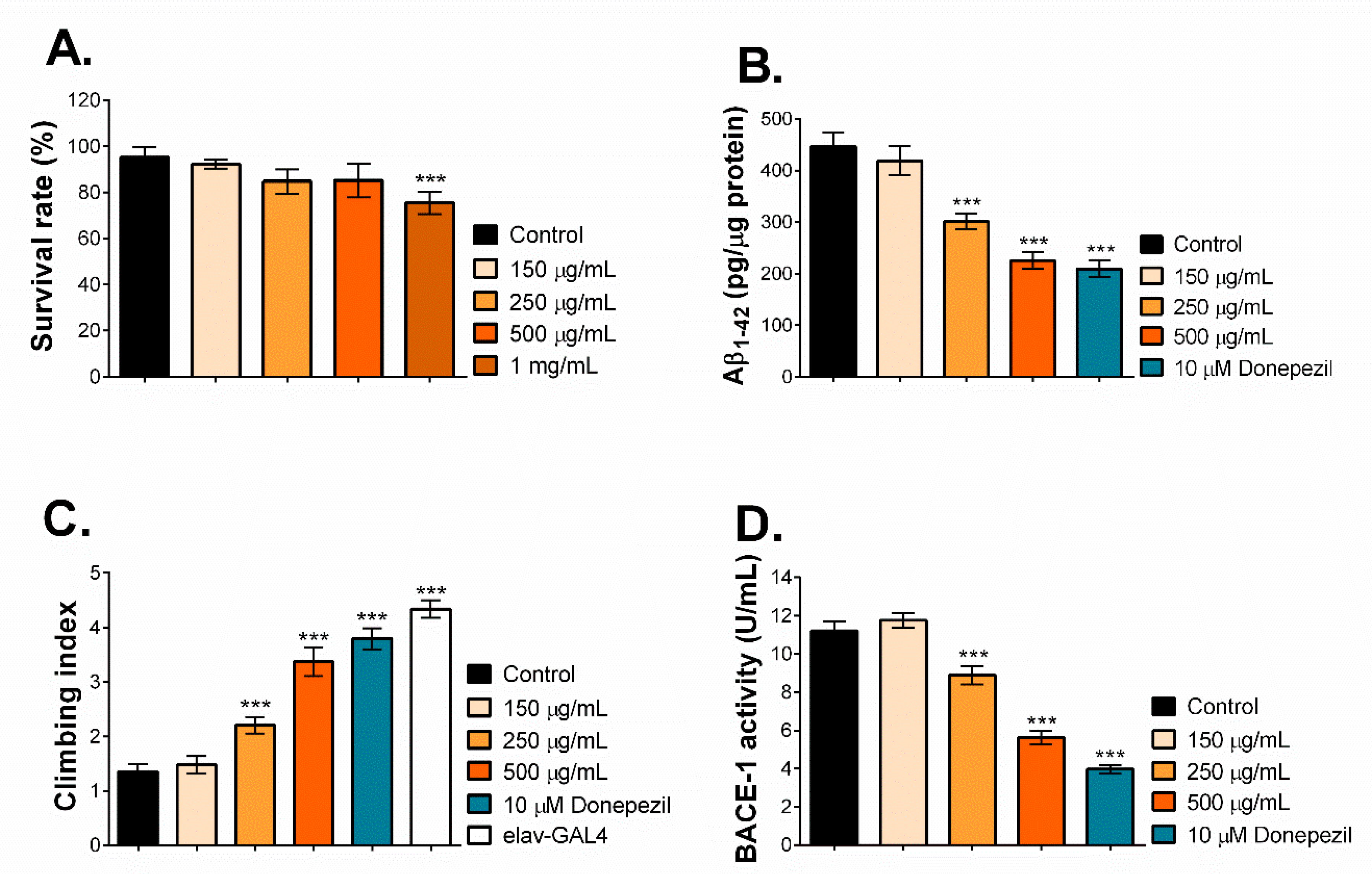
2.6. MNCM Extract Reduces Aβ1-42 by Inhibiting BACE-1 Activity in a Drosophila Model of AD
3. Discussion
4. Materials and Methods
4.1. Mulberry Collection and Preparation
4.2. In Vitro Antioxidant Activity
4.3. Total Phenolic Contents, Anthocyanin, and Anthocyanidin Determination
4.4. Determination of Cholinesterases and Beta-Secretase 1 (BACE-1) Inhibitory Activities
4.5. Cell Culture and Cytotoxicity Analysis by Resazurin Assay
4.6. Prevention of Hydrogen Peroxide (H2O2) and Aβ Peptide-Induced Toxicity
4.7. Determination of Neurite Outgrowth
4.8. Drosophila Stocks and Culture
4.9. Climbing Assay
4.10. Quantification of Aβ Peptide by Enzyme-Linked Immunosorbent Assay (ELISA)
4.11. Determination of BACE-1 Activity in Fly Brain Lysate
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, H.; Naghavi, M.; Allen, C.; Barber, R.M.; A Bhutta, Z.; Carter, A.; Casey, D.C.; Charlson, F.; Chen, A.Z.; Coates, M.M.; et al. Global, regional, and national life expectancy, all-cause mortality, and cause-specific mortality for 249 causes of death, 1980–2015: A systematic analysis for the Global Burden of Disease Study 2015. Lancet 2016, 388, 1459–1544. [Google Scholar] [CrossRef] [Green Version]
- Alzheimer’s association. Alzheimer’s disease facts and figures. Alzheimer’s Dement 2015, 11, 321–387. [Google Scholar]
- Ferreira-Vieira, T.H.; Guimaraes, I.M.; Silva, F.R.; Ribeiro, F. Alzheimer’s Disease: Targeting the Cholinergic System. Curr. Neuropharmacol. 2016, 14, 101–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Citron, M.; Diehl, T.S.; Gordon, G.; Biere, A.L.; Seubert, P.; Selkoe, D.J. Evidence that the 42- and 40-amino acid forms of amyloid β protein are generated from the β-amyloid precursor protein by different protease activities. Proc. Natl. Acad. Sci. USA 1996, 93, 13170–13175. [Google Scholar] [CrossRef] [Green Version]
- Dai, Q.; Borenstein, A.R.; Wu, Y.; Jackson, J.C.; Larson, E.B. Fruit and Vegetable Juices and Alzheimer’s Disease: The Kame Project. Am. J. Med. 2006, 119. [Google Scholar] [CrossRef] [Green Version]
- Loef, M.; Walach, H. Fruit, vegetables and prevention of cognitive decline or dementia: A systematic review of cohort studies. J. Nutr. Health Aging 2012, 16, 626–630. [Google Scholar] [CrossRef]
- Pacheco, S.M.; Soares, M.S.P.; Gutierres, J.M.; Gerzson, M.F.B.; Carvalho, F.B.; Azambuja, J.H.; Schetinger, M.R.C.; Stefanello, F.M.; Spanevello, R.M. Anthocyanins as a potential pharmacological agent to manage memory deficit, oxidative stress and alterations in ion pump activity induced by experimental sporadic dementia of Alzheimer’s type. J. Nutr. Biochem. 2018, 56, 193–204. [Google Scholar] [CrossRef]
- Badshah, H.; Kim, T.H.; Kim, M.O. Protective effects of Anthocyanins against Amyloid beta-induced neurotoxicity in vivo and in vitro. Neurochem. Int. 2015, 80, 51–59. [Google Scholar] [CrossRef]
- Özgen, M.; Serce, S.; Kaya, C. Phytochemical and antioxidant properties of anthocyanin-rich Morus nigra and Morus rubra fruits. Sci. Hortic. 2009, 119, 275–279. [Google Scholar] [CrossRef]
- Chen, H.; Yu, W.; Chen, G.; Meng, S.; Xiang, Z.; He, N. Antinociceptive and Antibacterial Properties of Anthocyanins and Flavonols from Fruits of Black and Non-Black Mulberries. Molecules 2017, 23, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.H.; Choi, C.-I. Pharmacological Properties of Morus nigra L. (Black Mulberry) as A Promising Nutraceutical Resource. Nutrients 2019, 11, 437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyuncu, F.; Cetinbas, M.; Ibrahim, E. Nutritional constituents of wild–grown black mulberry (Morus nigra L.). J. Appl. Bot. Food Qual. 2014, 87, 93–96. [Google Scholar]
- Imran, M.; Khan, H.; Shah, M.; Khan, R.; Khan, F. Chemical composition and antioxidant activity of certain Morus species. J. Zhejiang Univ. Sci. B 2010, 11, 973–980. [Google Scholar] [CrossRef]
- Qiao, A.; Wang, Y.; Zhang, W.; He, X. Neuroprotection of Brain-Targeted Bioactive Dietary Artoindonesianin O (AIO) from Mulberry on Rat Neurons as a Novel Intervention for Alzheimer’s Disease. J. Agric. Food Chem. 2015, 63, 3687–3693. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.-L.; Tang, G.; Guo, Y.-Q.; Xu, Y.-K.; Huang, Z.-S.; Yin, S. Mulberry Diels-Alder-type adducts from Morus alba as multi-targeted agents for Alzheimer’s disease. Phytochemistry 2019, 157, 82–91. [Google Scholar] [CrossRef]
- Chen, Z.; Zhong, C. Oxidative stress in Alzheimer’s disease. Neurosci. Bull. 2014, 30, 271–281. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Boyd-Kimball, D. Oxidative Stress, Amyloid-β Peptide, and Altered Key Molecular Pathways in the Pathogenesis and Progression of Alzheimer’s Disease. J. Alzheimer’s Dis. 2018, 62, 1345–1367. [Google Scholar] [CrossRef] [Green Version]
- Frozza, R.L.; Horn, A.P.; Hoppe, J.B.; Simao, F.; Gerhardt, D.; Comiran, R.A.; Salbego, C.G. A comparative study of beta-amyloid peptides Abeta1–42 and Abeta25–35 toxicity in organotypic hippocampal slice cultures. Neurochem. Res. 2009, 34, 295–303. [Google Scholar] [CrossRef]
- Dowjat, W.K.; Wisniewski, T.; Efthimiopoulos, S.; Wisniewski, H.M. Inhibition of neurite outgrowth by familial Alzheimer’s disease-linked presenilin-1 mutations. Neurosci. Lett. 1999, 267. [Google Scholar] [CrossRef]
- Wang, X.; Kim, J.-R.; Lee, S.-B.; Kim, Y.-J.; Jung, M.Y.; Kwon, H.W.; Ahn, Y.-J. Effects of curcuminoids identified in rhizomes of Curcuma longa on BACE-1 inhibitory and behavioral activity and lifespan of Alzheimer’s disease Drosophila models. BMC Complement. Altern. Med. 2014, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.; Liu, Y.C.; Meng, J.; Zhu, H.; Zhang, X. A microfluidics-based mobility shift assay to identify new inhibitors of beta-secretase for Alzheimer’s disease. Anal. Bioanal. Chem. 2017, 409, 6635–6642. [Google Scholar] [CrossRef]
- Mancini, F.; De Simone, A.; Andrisano, V. Beta-secretase as a target for Alzheimer’s disease drug discovery: An overview of in vitro methods for characterization of inhibitors. Anal. Bioanal. Chem. 2011, 400, 1979–1996. [Google Scholar] [CrossRef]
- Thuphairo, K.; Sornchan, P.; Suttisansanee, U. Bioactive Compounds, Antioxidant Activity and Inhibition of Key Enzymes Relevant to Alzheimer’s Disease from Sweet Pepper (Capsicum annuum) Extracts. Prev. Nutr. Food Sci. 2019, 24, 327–337. [Google Scholar] [CrossRef]
- Bae, S.-H.; Suh, H. Antioxidant activities of five different mulberry cultivars in Korea. LWT 2007, 40, 955–962. [Google Scholar] [CrossRef]
- Oki, T.; Kobayashi, M.; Nakamura, T.; Okuyama, A.; Masuda, M.; Shiratsuchi, H.; Suda, I. Changes in Radical-scavenging Activity and Components of Mulberry Fruit During Maturation. J. Food Sci. 2006, 71, 18–22. [Google Scholar] [CrossRef]
- Natić, M.; Dabić, D.Č; Papetti, A.; Akšić, M.F.; Ognjanov, V.; Ljubojević, M.; Tešić, Ž. Analysis and characterisation of phytochemicals in mulberry (Morus alba L.) fruits grown in Vojvodina, North Serbia. Food Chem. 2015, 171, 128–136. [Google Scholar] [CrossRef]
- Wang, S.Y.; Lin, H.-S. Antioxidant activity in fruits and leaves of blackberry, raspberry, and strawberry varies with cultivar and developmental stage. J. Agric. Food Chem. 2000, 48, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-S. Antioxidant Activities of Selected Berries and Their Free, Esterified, and Insoluble-Bound Phenolic Acid Contents. Prev. Nutr. Food Sci. 2018, 23, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Ştefănuţ, M.N.; Căta, A.; Pop, R.; Mosoarca, C.; Zamfir, A.D. Anthocyanins HPLC-DAD and MS Characterization, Total Phenolics, and Antioxidant Activity of Some Berries Extracts. Anal. Lett. 2011, 44, 2843–2855. [Google Scholar] [CrossRef]
- Pawlowska, A.M.; Oleszek, W.; Braca, A. Quali-quantitative Analyses of Flavonoids of Morus nigra L. and Morus alba L. (Moraceae) Fruits. J. Agric. Food Chem. 2008, 56, 3377–3380. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Li, Y.; Niu, W.; Ding, Y.; Zhang, R.; Shang, X. Analysis and characterisation of anthocyanins in mulberry fruit. Czech. J. Food Sci. 2010, 28, 117–126. [Google Scholar] [CrossRef]
- Polumackanycz, M.; Sledzinski, T.; Goyke, E.; Wesolowski, M.; Viapiana, A. A Comparative Study on the Phenolic Composition and Biological Activities of Morus alba L. Commercial Samples. Molecules 2019, 24, 3082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szwajgier, D. Anticholinesterase Activities of Selected Polyphenols—A Short Report. Pol. J. Food Nutr. Sci. 2014, 64, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Meng, X.; Yan, C.; Wang, C. Effect of Purple Sweet Potato Anthocyanins on β-Amyloid-Mediated PC-12 Cells Death by Inhibition of Oxidative Stress. Neurochem. Res. 2009, 35, 357–365. [Google Scholar] [CrossRef]
- Hong, H.; Liu, G.-Q. Protection against hydrogen peroxide-induced cytotoxicity in PC12 cells by scutellarin. Life Sci. 2004, 74, 2959–2973. [Google Scholar] [CrossRef]
- Song, N.; Yang, H.; Pang, W.; Qie, Z.; Lu, H.; Tan, L.; Li, H.; Sun, S.; Lian, F.; Qin, C.; et al. Mulberry extracts alleviate abeta 25-35-induced injury and change the gene expression profile in PC12 cells. Evid Based Complement. Alternat Med. 2014, 1–9. [Google Scholar] [CrossRef]
- Zheng, Z.-C.; Cho, N.C.; Wang, Y.; Fu, X.-T.; Li, D.-W.; Wang, K.; Wang, X.-Z.; Li, Y.; Sun, B.-L.; Yang, X.-Y. Cyanidin suppresses amyloid beta-induced neurotoxicity by inhibiting reactive oxygen species-mediated DNA damage and apoptosis in PC12 cells. Neural Regen. Res. 2016, 11, 795–800. [Google Scholar] [CrossRef]
- Chen, G.; Bower, K.A.; Xu, M.; Ding, M.; Shi, X.; Ke, Z.; Luo, J. Cyanidin-3-Glucoside Reverses Ethanol-Induced Inhibition of Neurite Outgrowth: Role of Glycogen Synthase Kinase 3 Beta. Neurotox. Res. 2009, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.G.; Oh, M.S. Memory-enhancing effect of Mori Fructus via induction of nerve growth factor. Br. J. Nutr. 2012, 110, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Pandey, U.B.; Nichols, C.D. Human disease models in Drosophila melanogaster and the role of the fly in therapeutic drug discovery. Pharmacol. Rev. 2011, 63. [Google Scholar] [CrossRef] [Green Version]
- Chintamaneni, M.; Bhaskar, M. Biomarkers in Alzheimer’s disease: A review. ISRN Pharmacol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.K.; Gemma, S.; Tang, J. beta-Secretase as a therapeutic target for Alzheimer’s disease. Neurotherapeutics 2008, 5, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Fornasaro, S.; Ziberna, L.; Gasperotti, M.; Tramer, F.; Vrhovšek, U.; Mattivi, F.; Passamonti, S. Determination of cyanidin 3-glucoside in rat brain, liver and kidneys by UPLC/MS-MS and its application to a short-term pharmacokinetic study. Sci. Rep. 2016, 6, 22815. [Google Scholar] [CrossRef] [PubMed]
- Afzal, M.; Redha, A.; AlHasan, R. Anthocyanins Potentially Contribute to Defense against Alzheimer’s Disease. Molecules 2019, 24, 4255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benzie, I.; Strain, J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Fukumoto, L.R.; Mazza, G. Assessing Antioxidant and Prooxidant Activities of Phenolic Compounds. J. Agric. Food Chem. 2000, 48, 3597–3604. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef]
- Sripum, C.; Kukreja, R.K.; Charoenkiatkul, S.; Kriengsinyos, W.; Suttisansanee, U. The effect of extraction conditions on antioxidant activities and total phenolic contents of different processed Thai Jasmine rice. Int. Food Res. J. 2017, 24, 1644–1650. [Google Scholar]
- Jung, H.A.; Min, B.S.; Yokozawa, T.; Lee, J.-H.; Kim, Y.S.; Choi, J.S. Anti-Alzheimer and antioxidant activities of Coptidis Rhizoma alkaloids. Biol. Pharm. Bull. 2009, 32, 1433–1438. [Google Scholar] [CrossRef] [Green Version]
- Nantakornsuttanan, N.; Thuphairo, K.; Kukreja, R.K.; Charoenkiatkul, S.; Suttisansanee, U. Anti-cholinesterase inhibitory activities of different varieties of chili peppers extracts. Int. Food Res. J. 2016, 23, 1953–1959. [Google Scholar]
- Jantrapirom, S.; Piccolo, L.L.; Yoshida, H.; Yamaguchi, M. A new Drosophila model of Ubiquilin knockdown shows the effect of impaired proteostasis on locomotive and learning abilities. Exp. Cell Res. 2018, 362, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Sofola-Adesakin, O.; Khericha, M.; Snoeren, I.; Tsuda, L.; Partridge, L. pGluAbeta increases accumulation of Abeta in vivo and exacerbates its toxicity. Acta Neuropathol. Commun. 2016, 4, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
| Independent Variable (Solvents, % (v/v) Aqueous Ethanol) | Dependent Variable % AChE Inhibition | Controlled Variables |
|---|---|---|
| 0% (v/v) (ultrapure water) | 40.29 ± 1.27 a |
|
| 20% (v/v) | 27.07 ± 1.36 b | |
| 40% (v/v) | ND * | |
| 60% (v/v) | ND * | |
| 80% (v/v) | ND * | |
| 100% (v/v) (absolute ethanol) | ND * |
| Independent Variable (Shaking Time, h) | Dependent Variable % AChE Inhibition | Controlled Variables |
|---|---|---|
| 0.5 | 27.63 ± 0.42 c |
|
| 1 | 33.63 ± 0.97 b | |
| 2 | 43.35 ± 3.32 a | |
| 4 | 30.18 ± 1.15 c | |
| 6 | 28.41 ± 0.58 c |
| Independent Variable (Temperature, °C) | Dependent Variable % AChE Inhibition | Controlled Variables |
|---|---|---|
| 30 | 27.89 ± 1.36 b |
|
| 50 | 32.20 ± 2.67 a | |
| 70 | 23.66 ± 2.16 c | |
| 90 | 11.29 ± 0.18 d |
| Time (min) | Solvent A | Solvent B |
|---|---|---|
| 0 | 88 | 12 |
| 6 | 88 | 12 |
| 8 | 85 | 15 |
| 25 | 85 | 15 |
| 25 | 88 | 12 |
| 30 | 88 | 12 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suttisansanee, U.; Charoenkiatkul, S.; Jongruaysup, B.; Tabtimsri, S.; Siriwan, D.; Temviriyanukul, P. Mulberry Fruit Cultivar ‘Chiang Mai’ Prevents Beta-Amyloid Toxicity in PC12 Neuronal Cells and in a Drosophila Model of Alzheimer’s Disease. Molecules 2020, 25, 1837. https://doi.org/10.3390/molecules25081837
Suttisansanee U, Charoenkiatkul S, Jongruaysup B, Tabtimsri S, Siriwan D, Temviriyanukul P. Mulberry Fruit Cultivar ‘Chiang Mai’ Prevents Beta-Amyloid Toxicity in PC12 Neuronal Cells and in a Drosophila Model of Alzheimer’s Disease. Molecules. 2020; 25(8):1837. https://doi.org/10.3390/molecules25081837
Chicago/Turabian StyleSuttisansanee, Uthaiwan, Somsri Charoenkiatkul, Butsara Jongruaysup, Somying Tabtimsri, Dalad Siriwan, and Piya Temviriyanukul. 2020. "Mulberry Fruit Cultivar ‘Chiang Mai’ Prevents Beta-Amyloid Toxicity in PC12 Neuronal Cells and in a Drosophila Model of Alzheimer’s Disease" Molecules 25, no. 8: 1837. https://doi.org/10.3390/molecules25081837