Abstract
Styrylquinolines are heterocyclic compounds that are known for their antifungal and antimicrobial activity. Metal complexation through hydroxyl groups has been claimed to be a plausible mechanism of action for these types of compounds. A series of novel structures with protected hydroxyl groups have been designed and synthesized to verify the literature data. Their antifungal activity against wild-type Candida albicans strain and mutants with silenced efflux pumps activity has been determined. Combinations with fluconazole revealed synergistic interactions that were dependent on the substitution pattern. These results open a new route for designing active antifungal agents on a styrylquinoline scaffold.
1. Introduction
Invasive mycoses are an important public health problem, especially for immunosuppressed patient. Candida albicans is the one that is most often associated with serious invasive fungal infections, and it is responsible for 8–10% of the cases with a high mortality rate (30–80% depending on the patient) [1]. Treatment of fungal infections is markedly limited by the problems of drug safety and fungal resistance. Among the drugs used in antifungal treatment, echinocandin is used for invasive candidiasis while voriconazole and isavuconazole are the drugs of choice for aspergillosis. Other drugs from the conazoles group, like fluconazole (FLC), have rather limited use due to the fact of resistance. Another drug with a wide spectrum of activity is amphotericin B (AmB). Amphotericin B belongs to a group of polyene antibiotics that act by forming an oligomeric pore structure within the fungal plasma membrane by interacting with the membrane sterols. This leads to a flux of cations, membrane depolarization, and cell death [2]. Amphotericin B can also bind to cholesterol in mammalian cells and can cause severe cellular toxicity [3,4]. The mode of action of the azole derivatives is based on inhibiting the ergosterol biosynthesis pathway at different stages [5]. Ergosterol is the major component of the fungal cell membrane; hence, it is one of the targets for antifungal drugs [6]. However, the growing resistance of fungi to azoles is a problem. Candida albicans has developed many drug resistance mechanisms. One example is the overexpression of the drug transporters (e.g., ATP binding cassette (ABC) pumps—Candida drug resistance (Cdr1p and Cdr2p). This mechanism is not very specific; ABC transporters have the ability to export many structurally dissimilar compounds, for instance, azoles, among others [7].
Our team has been investigating the quinoline derivatives for their biological activity for several years. For example, anticancer [8,9], antibacterial [10,11] and antifungal [12,13,14] quinolines have been described during our research. In general, the idea of small-molecule quinolines that have an antifungal activity has been known from the works of Gershon [15,16,17,18]. At the beginning, compounds that were described were based on small quinolines that were substituted with halogens and preferably with the 8-hydroxy group (Figure 1(1)). Their activity was non-specific and covered a broad range of fungal strains. These initial examples were later developed into more sophisticated molecules that had a more specific mechanism of action.
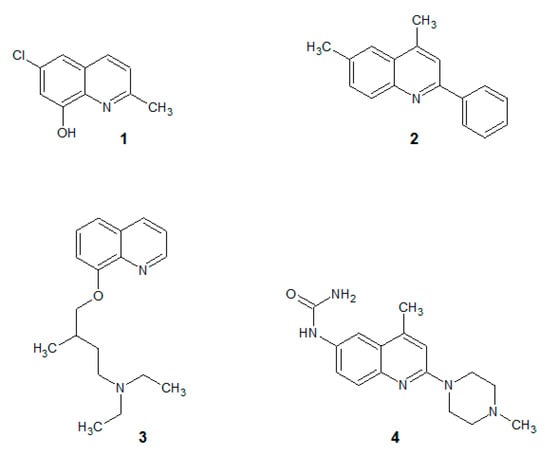
Figure 1.
Antifungal quinolines. (1): oxine derivative; (2): 2-phenylquinoline; (3): alkylated oxine; (4): 2-morpholine derivative.
For example, several quinolines were found to be inhibitors of the fungal cell wall synthesis (Figure 1(2)) [19]. They are lytic for microbial cells (Figure 1(3)) [20] including the Candida strains [21,22] and cause an accumulation of endogenous reactive oxygen species in C. albicans biofilms (Figure 1(4)) [23].
In this study, we present styrylquinolines (SQLs) that have promising, novel antifungal properties. Styrylquinolines are interesting lipophilic compounds that were initially obtained as analogues of allylamines [19], although their spectrum of activity is wider, covering also anticancer [24], antibacterial [10], and antiviral activity [25,26]. Interestingly, favorable patterns of substitution vary among these activities. The 8-hydroxyquinoline core prevails in most of those applications but other positions are more specific and vary among types of activity. For example, for antiviral activity, SQLs should have electron-donating groups in the styryl part of the molecule and particularly effective are 3,4-dihydroxyl or alkoxyl groups [25,27]. The contrary is beneficial for anticancer activity where resonance positions (2-, 4-) in the styryl moiety should be occupied by electron-withdrawing groups such as cyano or nitro [9]. In antifungal activity, particularly high activities may be achieved by additional electron-accepting groups in quinoline moiety such as in 5,7-dichloro-8-hydroxyquinoline [10]. This may be associated with ability to chelate metal ions, that was primarily established as the mechanism of action of small molecule quinoline antifungal agents [28,29]. Otherwise, the structure–antifungal activity relationship for substitution in styryl rings is not yet fully explored. According to our previous works as well as the literature data, the halogen and hydroxyl (but not alkoxyl) substituents at position C4 seems to be effective [10,30]. Therefore, we decided to investigate whether the free hydroxyl group is essential for this activity (Figure 2).
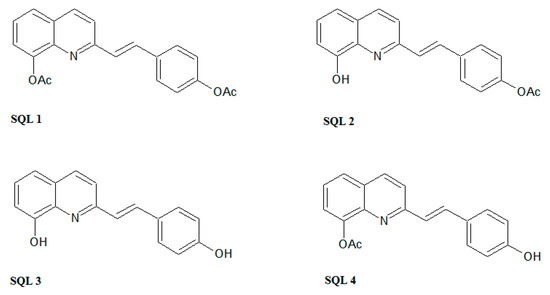
Figure 2.
The styrylquinolines (SQLs) that were synthesized in this study.
2. Results
In order to produce all of the combinations of acetyl/hydroxyl derivatives, we used selective hydrolysis and the modified methods that are typically used for styrylquinoline synthesis as is depicted in Figure 3.
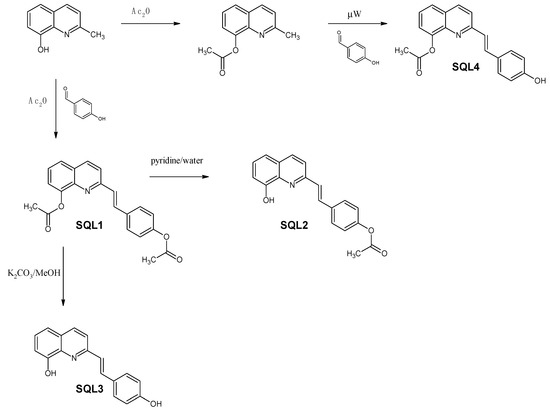
Figure 3.
Synthesis of the styrylquinolines.
Commercially available 8-hydroxy-2-methylquinoline was reacted with acetic anhydride to acetylate the hydroxyl group. The resulting 8-acetoxyquinaldine was converted into SQL 4 under microwave irradiation according to the modified method that was reported for other SQLs [31]. Similarly, 8-hydroxy-2-methylquinoline combined with 4-hydroxybenzaldehyde was refluxed in an acetic anhydride/acetic acid mixture for 18 h to produce SQL 1. Both hydroxyl groups were acetylated in this reaction environment. Then, the acetic anhydride was evaporated under a vacuum. A pyridine/water mixture was added to the residue and refluxed for 3 h, which resulted in selectively deacetylated SQL 2, while the hydrolysis of SQL 1 in methanolic K2CO3 produced SQL 3.
2.1. Antifungal Activity
To assess the antifungal activity of the styrylquinolines that were investigated, we determined the minimal inhibitory concentration (MIC) values for the wild-type C. albicans strain. Furthermore, we selected single and double cdrΔ mutants (Table 1) in order to gain insight into the chance of resistance.

Table 1.
The minimum inhibitory concentration (mg/L) at which 50% of isolates were inhibited (MIC50) for tested styrylquinoline compounds and amphotericin B (AmB) (n = 3).
Compounds SQL 4, SQL 1, and SQL 2, which did not have or had one hydroxyl group, had a rather weak antifungal activity. It is worth noting that the position of the OH/AcO group seemed to be irrelevant as both of the SQLs that had only one OH group that was unprotected had the same MIC level as SQL 1 (Table 2). The SQL 3 had a strong antifungal effect, especially on the cdr1Δ strain. Control compounds 8-hydroxyquinoline and 8-hydroxyquinaldine had a similar or worse effect on the strains (Table 2).
Table 2.
The fractional inhibitory concentration (FIC) values for styrylquinoline compounds (n = 3) that were tested.
The simultaneous applications of two or more compounds is a good strategy against resistant strains. The synergistic action of the mixtures of drugs can overcome resistance even with apparently inactive substances [32]. To measure interactions of the SQLs with FLC, we determined the fractional inhibitory concentration (FIC) for the drugs that were tested separately and in combinations. Next, we estimated the FIC index for all of the interactions. These indexes can be used to determine interaction types among two or more drugs that are used simultaneously. In general, FICs below 0.5 can be considered as synergistic as presented in Section 4.3. We found a synergism between SQL 3 and FLC against C. albicans (Table 2) but no difference in the FIC between the wild-type strain and the mutants which indicates a non-specific activity of SQL 3 and FLC to the ABC pumps (Table 2). We found a four-fold weaker FIC interaction for SQL 2 for the cdr1Δ and cdr1Δcdr2Δ strains than for the rest of strains. This compound as well as FLC separately were ejected from the Candida cell by the ABC transporters, but when the cells were treated with a combination of drugs, their export decreased due to the competitive inhibition of Cdr2p, hence the low FIC for the cdr1Δ and cdr1Δcdr2Δ mutant. A similar situation was observed in the case of SQL 4 for which the synergy was only apparent in the strains that had one or both of the transporters deleted—when both of the Cdr pumps were expressed, they complemented each other’s activity and no synergistic action was observed.
2.2. The Rhodamine 6G Efflux Assay
In order to determine whether SQLs influence ABC transporters in C. albicans, we investigated the activity of the efflux pumps using an R6G assay. All of the compounds were applied at a concentration that was below their antifungal activity concentration to maintain living cells during the experiment. The SQL 4 had a weak influence on the R6G leakage from the cells (Figure 4). Despite the high synergism of SQL 3 with FLC, we did not observe any inhibition of the pumps removing the R6G from the cells (Figure 4). We also observed differences between the synergisms of SQL 1 and SQL 2 with FLC and their influence on the R6G efflux from the cells (Table 2, Figure 4).
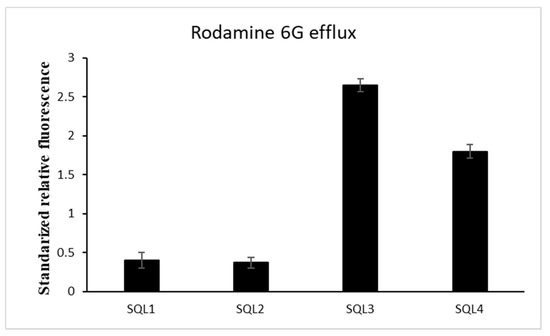
Figure 4.
Styrylquinolines (½MIC) influence on energy-dependent rhodamine 6G efflux 30 min after efflux was induced by adding glucose. (±SD, n = 3).
2.3. The Influence of Styrylquinolines on the Cdr1p Level
The treatment of C. albicans with antifungals and glucose can increase the expression of the ABC transporters [33,34]. Fluorescent microscopy (Figure 5) showed a stronger signal from the green fluorescent protein (GFP)-tagged Cdr1p in the cells that had been incubated with SQLs than in the control without the compounds that were tested. To confirm this observation, we performed Western blot experiments with crude protein extracts that were obtained from cells that were incubated with SQLs (Figure 6). The Cdr1p-GFP protein level in these extracts was also higher in the treated cells than in control.
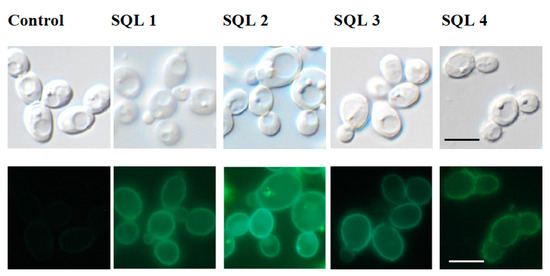
Figure 5.
Microscopic imagining of the Candida albicans cells. Top panels: differential interference contrast; bottom panels: Cdr1p-GFP. Scale bars = 10 μm.

Figure 6.
Cdr1p-GFP protein levels in crude protein extracts of the cells after treatment with styrylquinolines (½MIC) as was evidenced by the anti-GFP Western blot (n = 4); DMSO-treated C. albicans cells were used as the control.
Among them, SQL 2 was the most effective inductor of Cdr1p-GFP protein synthesis followed by SQL 1 and SQL 4. On the other hand, SQL 3 caused reduction in Cdr1p-GFP protein level in the cells.
3. Discussion
Based on our experience with SQLs, we decided to use acetylation as the protection of hydroxyl groups. Although hydrolysis of acetoxy group may be rapid, it depends on structure and condition [35]. The same can be found in enzyme-dependent hydrolysis as in lipases present in Candida spp. [36]. We found that acetylated styrylquinolines are rather resistant to decomposition in solvent and in the cell. The same was reported also in mammalian cells during in vitro examination of anticancer activity [9,37]. Even during prolonged (96 h) incubation, derivatives with hydroxyl groups are more than ten times active than their acetylated representatives. Moreover, low yield of SQL 2 (15%) can also be associated with incomplete hydrolysis of SQL 1 in a pyridine/water environment, and problems with isolation and proper purification of the compound to obtain high purity (multiple crystallization). An alternative method for obtaining styrylquinolines is synthesis in the microwave field, where the reaction environment is excess of aldehyde as in the case of SQL 4. The microwave method significantly reduces the reaction time and resolves the disadvantages associated with the need for hydrolysis. Unfortunately, it is generally only effective when the aldehyde is a liquid or has a low melting point as reported elsewhere [31]. Otherwise, this method leads to a mixture of substrates and product (as in the case of SQL 4, yield 9%), without solving problems with the isolation and purification of final compounds.
Antifungal activity of styrylquinolines that were synthesized was tested against C. albicans strains of different expression of Cdr efflux pumps. The observed activities were only partially in agreement with our other investigations in which the free 8-OH group seems essential for activity. On the other hand, the 4-OH in the styryl part seemed to have an effect that was similar to the 4-OMe and 4-OEt substituents regardless of the quinoline part [10,38]. It is widely documented that the -OH group plays an important role in destroying fungal cells [39,40,41]. Some compounds that contain the phenolic groups can reduce ergosterol biosynthesis and damage the cell membrane of C. albicans [42] or bind to chitin in the cell wall [43]. In this regard, the 8-OH/4-OH derivative expressed some interesting patterns of activity that are worth of further investigation. The same level of activity of SQL 3 against mutants without Cdr1p and both pumps (Table 1) indicated its role as a substrate of Cdr1p. Unlike other tested compounds, AmB showed an MIC50 at the same concentrations for all strains, suggesting that it is not a substrate for ABC transporters.
In our previous investigations, we observed different influences on the Cdr1 transporter when combinations of FLC-beauvericin and diS-C3(3)-beauvericin were applied in which beauvericin was an inhibitor of the ABC transporters and diS-C3(3) was the substrate for these pumps [44]. Similar differences in inhibitory activity were observed for curcumin [45], the modulatory effect of which was restricted to R6G or miconazole, while it had no effect on the efflux of FLC. Larger flat, aromatic and highly lipophilic compounds, such as styrylquinolines, are known to accumulate in the cell membranes. To confirm that the cell membrane of C. albicans was not disrupted by SQL 3 and SQL 4, for which the efflux of rhodamine was higher than the control, the cells that had been treated with styrylquinolines were stained with non-permeable propidium iodide dye [23,46] and observed under a fluorescent microscope. SQL 1, SQL 2, SQL 3, and SQL 4 did not show the red staining that is characteristic of membrane disruption (data not shown). Confirmation of this findings was elaborated by means of measuring expression level of Cdr1p-GFP in crude extracts. Surprisingly, it appeared that the SQL 3 compound reduced the amount of Cdr1p in the cells (Figure 6). Taking into account all of the data that were obtained for this compound renders this observation even more interesting. The SQL3 showed the best antifungal activity of all tested styrylquinolines both alone (Table 1) and in combination with fluconazole (Table 2). SQL1, SQL2, and SQL4 increased both the level of gene expression and the amount of Cdr1 in C. albicans cells (Figure 6) which probably contributed to the high resistance of this fungus to these compounds (Table 1).
To sum up, we designed and synthesized four styrylquinolines in order to verify the hypothesis of the importance of the hydroxyl groups in the quinoline and phenyl moieties. All of the compounds were tested against the C. albicans wild type and mutants that had been deprived of one or two ABC efflux pumps. According to the expectations, the SQL 3 compound with both hydroxyl groups that were unprotected appeared to be the most active against the tested strains. Its activity pattern suggested efflux through the Cdr1 pump. All of the compounds that were obtained were also tested in combination therapy with a known Cdr substrate,FLC. The synergistic interactions and activity were strongly reliant on the substitution pattern. Additional experiments will enable us to determine novel inhibitors of the Cdr1p protein.
4. Materials and Methods
4.1. Compounds
(E)-2-[2-(4-acetoxyphenyl)vinyl]-8-acetoxyquinoline (SQL 1). Yield 67%, mp. 157–159 °C; 1H-NMR (400 MHz, CDCl3-d) δ 8.15 (dd, J = 8.5, 5.1 Hz, 1H), 7.73–7.60 (m, 5H), 7.55–7.37 (m, 3H), 7.19–7.12 (m, 2H), 2.58 (s, 3H), 2.35 (s, 3H); 13C-NMR (101 MHz, CDCl3-d) δ 169.84, 169.33, 155.65, 150.91, 147.38, 140.94, 136.41, 134.25, 133.71, 131.21, 129.10, 128.60, 128.32, 125.75, 125.54, 121.95, 121.68, 120.19, 21.17, 21.03 [47].
(E)-2-[2-(4-acetoxyphenyl)vinyl]quinolin-8-ol (SQL 2). Yield 15%, mp. 147–149 °C; 1H-NMR (400 MHz, DMSO-d6) δ 9.56 (s, 1H), 8.29 (t, J = 10.8 Hz, 1H), 8.14 (d, J = 16.2 Hz, 1H), 7.83–7.72 (m, 3H), 7.53–7.42 (m, 1H), 7.43–7.33 (m, 2H), 7.22 (t, J = 5.6 Hz, 2H), 7.15–7.05 (m, 1H), 2.30 (s, 3H); 13C-NMR (126 MHz, DMSO-d6) δ 169.65, 153.79, 153.40, 151.09, 138.61, 137.00, 134.66, 133.83, 128.65, 128.57, 128.17, 127.55, 122.83, 121.46, 118.05, 111.70, 21.36 [9].
(E)-2-[2-(4-hydroxyphenyl)vinyl]quinolin-8-ol (SQL 3). Yield 63%, mp. 156–157 °C; 1H-NMR (400 MHz, DMSO-d6) δ 11.81 (s, 1H), 10.38 (s, 1H), 8.84 (d, J = 8.9 Hz, 1H), 8.42 (d, J = 8.9 Hz, 1H), 8.22 (d, J = 16.2 Hz, 1H), 7.79 (d, J = 16.3 Hz, 1H), 7.67–7.56 (m, 4H), 7.47 (dd, J = 6.1, 2.6 Hz, 1H), 6.95 (d, J = 8.6 Hz, 2H); 13C-NMR (126 MHz, DMSO-d6) δ 161.19, 153.30, 148.85, 144.14, 143.99, 143.72, 130.85, 129.57, 128.20, 126.55, 119.16, 118.74, 116.83, 116.50 [47].
(E)-2-[2-(4-hydroxyphenyl)vinyl]-8-acetoxyquinoline (SQL 4).
Step 1. 8-Acetoxyquinaldine: The 8-hydroxyquinaldine derivative (2.5 mmol) in 10 mL acetic anhydride was heated for 16 h at 130 °C. Then, the mixture was evaporated to dryness and a solid was crystallized from EtOH to produce a light yellow solid, yield 95%, mp. 67 °C; 1H-NMR (400 MHz, CD2Cl2-d2) δ 8.12 (d, J = 8.5 Hz, 1H), 7.74 (dd, J = 8.2, 1.3 Hz, 1H), 7.50 (t, J = 7.8 Hz, 1H), 7.42 (dd, J = 7.5, 1.3 Hz, 1H), 7.37 (d, J = 8.5 Hz, 1H), 2.74 (s, 3H), 2.48 (s, 3H).
Step 2. (E)-2-[2-(4-hydroxyphenyl)vinyl]-8-acetoxyquinoline: The appropriate quinaldine derivative (1 mmol) was mixed thoroughly with two equivalent aldehyde, put into an open vessel, and exposed to microwave irradiation for 20 min at 180 °C with a maximum power 80 W. Then, the reaction mixture was cooled to 0 °C and the precipitate was filtered off. The solid was recrystallized from EtOH in order to produce a dark yellow solid, yield 9%, mp. 132–136 °C; 1H-NMR (500 MHz, DMSO-d6) δ 9.98 (s, 1H), 8.27 (t, J = 11.4 Hz, 1H), 8.10 (d, J = 16.4 Hz, 1H), 7.79 (d, J = 8.6 Hz, 1H), 7.66–7.59 (m, 1H), 7.56–7.48 (m, 1H), 7.40–7.32 (m, 2H), 7.21–7.13 (m, 1H), 7.13–7.04 (m, 1H), 6.95 (t, J = 8.8 Hz, 1H), 6.88 (t, J = 7.4 Hz, 1H), 1.91 (s, 3H); 13C-NMR (126 MHz, DMSO-d6) δ 169.69, 153.80, 153.38, 151.08, 138.60, 137.01, 134.64, 133.82, 128.66, 128.59, 128.17, 127.56, 122.82, 121.42, 118.07, 116.31, 111.71, 21.35.
4.2. Strains and Growth Media
The C. albicans strains used in this study were a generous gift from Dominique Sanglard (Table 3). All of the strains were kept as frozen stocks in glycerol at −80 °C and routinely grown at 28 °C on a YPD medium with 2% dextrose, 1% Bacto peptone (Diag-med, Warsaw, Poland) and 1% yeast extract (Diag-med, Warsaw, Poland), and a Sabouraud medium (4% dextrose, 1% Bacto peptone (Diag-med, Warsaw, Poland). For all of the experiments, except for a Rhodamine 6G (Sigma–Aldrich, Poznan, Poland) assay, the strains were sub-cultured in w yeast nitrogen base (YNB) liquid medium (0.67% YNB (Diag-med, Warsaw, Poland) and 2% dextrose) at 28 °C and diluted to the desired optical density at 600 nm (OD600). For growth on solid media, 2% agar (Difco) was added.
Table 3.
Collection of C. albicans strains that was used in this study. Strain CAF 2-1 is treated as a wild-type (WT) strain. The deleted genes encoding drug efflux pumps (Cdr1, Cdr2 and Mdr1) are marked delta in the Genotype column.
4.3. Susceptibility Testing
Drug susceptibility testing was performed in microtiter plates with two-fold serial dilutions of the tested compounds according to the EUCAST (EUCAST E.DEF 7.3) specifications with modifications. The cultures were grown for 18 h on Sabouraud agar at 35 °C, and a few colonies were suspended in 0.85% NaCl at 0.5 (1.5 × 106 CFU/mL) in the McFarland scale. Next, the suspension was diluted to a final concentration of 2.5 × 105 CFU/mL and 100 µL was mixed with 100 µL of the tested compound suspended in 2 × YNB or a mixture of FLC (Sigma–Aldrich; Poznan, Poland) and styrylquinoline, each at a starting concentration of MIC in order to obtain the fractional inhibitory concentration (FIC) indexes. Those indexes are determining type of interaction between two drugs, where: synergy: FICI ≤ 0.5, indifference: FICI > 0.5 to 4, antagonism: FICI > 4 [51]. The FIC index was defined as the MIC of drug “a” applied in combination with drug “b”, divided by the MIC of drug “a”, added together with the MIC of drug “b” applied in combination with drug “a”, divided by MIC of drug “b” according to the formula below (Equation (1)):
FICI = MICa+b/MICa + MICb+a/MICb
Microtiter plates were incubated at 28 °C for 24 h and then shaken, and their optical densities were read with a microtiter plate reader at a wavelength of 600 nm. The results are presented as the percentage of growth relative to the control samples, and the MIC values are given as the lowest concentration that inhibited 50% of the growth according to the EUCAST standards. The experiment was performed in three biological replicates with three technical replicates.
4.4. Permeabilization Assays
The yeast cultures were sub-cultured overnight in YNB and diluted to an OD600 of 0.4 in a fresh medium. The styrylquinoline compounds were added and incubated at 28 °C with shaking at 180 rpm. Aliquots of the cultures were taken after 4 h and stained with propidium iodide (manufacturer: Bioshop, distributor: Lab Empire, Rzeszów, Poland) [23,46] for 5 min at room temperature in order to assess the membrane permeability. Observations were made using fluorescent microscopy (Zeiss AxioVision, Poznań, Poland). The experiment was carried out in three biological replicates.
4.5. Western Blotting
The assay was performed according to a previous method [52] with modifications. A crude protein extract was prepared from the cell suspensions after 4 h of induction with the tested compounds. The aliquots of the cell suspensions were pelleted via centrifugation at 2260× g for 5 min and resuspended in 1 mL of deionized water. The cells were lysed by adding 150 μL of 1.85 M NaOH-7.5% β-mercaptoethanol (Sigma–Aldrich, Poznan, Poland) and incubated on ice for 10 min. The proteins were precipitated by adding 150 μL of 50% trichloroacetic acid and incubated on ice for 10 min. Samples were then centrifuged at 10,000× g for 5 min at 4 °C, washed in 1 mL of 1 M Tris-HCl pH 8.0, and then resuspended in 50 μL of the sample buffer (40 mM Tris-HCl, 8 M urea, 5% SDS, 0.1 mM EDTA 1% β-mercaptoethanol, 0.1 mg/mL bromophenol blue). Following incubation at 37 °C for 30 min, the samples were loaded onto 6% sodium dodecyl sulphate-polyacrylamide gel and developed in a Mini-PROTEAN II electrophoresis cell (Bio-Rad, Poznań, Poland). The samples were then transferred onto a nitrocellulose membrane using a Mini-PROTEAN Tetra System electrophoresis cell (Bio-Rad, Poznań, Poland). The membranes were stained with Poncau S to check whether the gels had loaded equally. The Cdr1p was immunodetected using polyclonal mouse anti-GFP antiserum with horseradish peroxidase-conjugated anti-mouse antiserum as a secondary antibody. The signals were detected using an ECL kit from PerkinElmer according to the manufacturer’s instructions. The experiment was performed in four biological replicates.
4.6. Rhodamine 6G Assay
The R6G assay was performed according to Nakamura et al. [53] with modifications. The cell culture was pelleted in the log phase and washed twice in double-distilled water and once in a HEPES buffer (50 mM, pH 7.0). The cells were diluted in a fresh HEPES buffer to an OD600 of 1.0 and incubated for 60 min at 30 °C and 200 rpm with 5 mM 2-deoxy-d-glucose (Sigma–Aldrich, Poznań, Poland). Next, the tested compounds were added in ½MIC concentrations and the cells were incubated for 5 min in the same conditions. Then, 10 µM R6G was added and the cell suspension was incubated for an additional 90 min. The cells were pelleted, washed twice in a HEPES buffer, and suspended in fresh HEPES at an OD600 of 10.0. The cell suspension was incubated for 5 min at 30 °C with shaking, the R6G efflux was initiated by adding 10 mM glucose and the suspensions were incubated for 30 min from which aliquots were removed at 15 min intervals. Aliquots of 400 µL were pelleted and three duplicates of 100 µL of supernatant were added to black microtiter plates. The fluorescence was measured in a Cary Eclipse spectrofluorimeter (Agilent Technologies, Santa Clara, CA, USA) at an excitation wavelength of 529 nm (slit 5) and an emission of 553 nm (slit 10). The experiment was performed in three biological replicates with three technical replicates.
Author Contributions
Conceptualization, A.K. and R.M.; Methodology, W.C. and J.S.; Investigation, W.C. and J.S.; Original draft preparation, A.K.; Review and editing, R.M. and W.C.; Project administration and funding acquisition, R.M. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Polish National Center for Science NCN (grant nos. 2013/09/B/NZ7/00423, 2018/31/B/NZ7/02122) and by The Leading National Research Centre (KNOW) programme for the years 2014–2018.
Acknowledgments
We thank Dominique Sanglard for her kind gift of the C. albicans strains. The technical assistance of Ewelina Spaczynska was greatly appreciated.
Conflicts of Interest
Authors declare no conflicts of interests.
References
- Perlroth, J.; Choi, B.; Spellberg, B. Nosocomial fungal infections: Epidemiology, diagnosis, and treatment. Med. Mycol. 2007, 45, 321–346. [Google Scholar] [CrossRef]
- Akins, R.A. An update on antifungal targets and mechanisms of resistance in Candida albicans. Med. Mycol. 2005, 43, 285–318. [Google Scholar] [CrossRef]
- Ibrahim, F.; Gershkovich, P.; Sivak, O.; Wasan, E.K.; Wasan, K.M. Pharmacokinetics and tissue distribution of amphotericin B following oral administration of three lipid-based formulations to rats. Drug Dev. Ind. Pharm. 2013, 39, 1277–1283. [Google Scholar] [CrossRef]
- Gruszecki, W.I.; Gagos, M.; Kernen, P. Polyene antibiotic amphotericin B in monomolecular layers: Spectrophotometric and scanning force microscopic analysis. FEBS Lett. 2002, 524, 92–96. [Google Scholar] [CrossRef]
- Odds, F. Antifungal agents: Mechanisms of action. Trends Microbiol. 2003, 11, 272–279. [Google Scholar] [CrossRef]
- Suchodolski, J.; Muraszko, J.; Bernat, P.; Krasowska, A. A Crucial Role for Ergosterol in Plasma Membrane Composition, Localisation, and Activity of Cdr1p and H+-ATPase in Candida albicans. Microorganisms 2019, 7, 378. [Google Scholar] [CrossRef]
- Prasad, R.; Rawal, M.K. Efflux pump proteins in antifungal resistance. Front. Pharmacol. 2014, 5, 1–13. [Google Scholar] [CrossRef]
- Mrozek-Wilczkiewicz, A.; Kalinowski, D.S.; Musiol, R.; Finster, J.; Szurko, A.; Serafin, K.; Knas, M.; Kamalapuram, S.K.; Kovacevic, Z.; Jampilek, J.; et al. Investigating the anti-proliferative activity of styrylazanaphthalenes and azanaphthalenediones. Bioorg. Med. Chem. 2010, 18, 2664–2671. [Google Scholar] [CrossRef]
- Mrozek-Wilczkiewicz, A.; Kuczak, M.; Malarz, K.; Cieślik, W.; Spaczyńska, E.; Musiol, R. The synthesis and anticancer activity of 2-styrylquinoline derivatives. A p53 independent mechanism of action. Eur. J. Med. Chem. 2019, 177, 338–349. [Google Scholar] [CrossRef]
- Cieslik, W.; Musiol, R.; Nycz, J.E.; Jampilek, J.; Vejsova, M.; Wolff, M.; Machura, B.; Polanski, J. Contribution to investigation of antimicrobial activity of styrylquinolines. Bioorg. Med. Chem. 2012, 20, 6960–6968. [Google Scholar] [CrossRef]
- Cieslik, W.; Spaczynska, E.; Malarz, K.; Tabak, D.; Nevin, E.; O’Mahony, J.; Coffey, A.; Mrozek-Wilczkiewicz, A.; Jampilek, J.; Musiol, R. Investigation of the Antimycobacterial Activity of 8-Hydroxyquinolines. Med. Chem. 2015, 11, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Musiol, R.; Serda, M.; Hensel-Bielowka, S.; Polanski, J. Quinoline-based antifungals. Curr. Med. Chem. 2010, 17, 1960–1973. [Google Scholar] [CrossRef] [PubMed]
- Musiol, R.; Jampilek, J.; Buchta, V.; Silva, L.; Niedbala, H.; Podeszwa, B.; Palka, A.; Majerz-Maniecka, K.; Oleksyn, B.; Polanski, J. Antifungal properties of new series of quinoline derivatives. Bioorg. Med. Chem. 2006, 14, 3592–3598. [Google Scholar] [CrossRef] [PubMed]
- Szczepaniak, J.; Cieślik, W.; Romanowicz, A.; Musioł, R.; Krasowska, A. Blocking and dislocation of Candida albicans Cdr1p transporter by styrylquinolines. Int. J. Antimicrob. Agents 2017, 50, 171–176. [Google Scholar] [CrossRef]
- Gershon, H.; Clarke, D.D.; Gershon, M. Synergistic Antifungal Action of 8-Quinolinol and Its Bischelate with Copper(Ⅱ) and with Mixed Ligand Chelates Composed of Copper(Ⅱ), 8-Quinolinol, and Aromatic Hydroxy Acids. J. Pharm. Sci. 1989, 78, 975–978. [Google Scholar] [CrossRef]
- Gershon, H.; Clarke, D.D.; Gershon, M. Evidence of Steric Factors in the Fungitoxic Mechanisms of 8-Quinolinol and Its 5- and 7-Halogenated Analogues. J. Pharm. Sci. 1991, 80, 542–544. [Google Scholar] [CrossRef]
- Gershon, H.; Gershon, M. Intramolecular synergism, an explanation for the enhanced fungitoxicity of halo-8-quinolinols. Mon. Chem. Chem. Mon. 1995, 126, 1303–1309. [Google Scholar] [CrossRef]
- Gershon, H.; Clarke, D.D.; McMahon, J.J.; Gershon, M. Preparation and Fungitoxicity of Some Dibromo-8-quinolinols. Mon. Chem. Chem. Mon. 2001, 132, 833–838. [Google Scholar] [CrossRef]
- Urbina, J.M.; Cortes, J.C.G.; Palma, A.; Lopez, S.N.; Zacchino, S.A.; Enriz, R.D.; Ribas, J.C.; Kouznetsov, V.V. Inhibitors of the fungal cell wall. Synthesis of 4-aryl-4-N-arylamine-1-butenes and related compounds with inhibitory activities on β(1–3) glucan and chitin synthases. Bioorg. Med. Chem. 2000, 8, 691–698. [Google Scholar] [CrossRef]
- Ghorab, M.; Abdel-Hamide, S.; Farrag, H. Synthesis of novel quinolines, pyranoquinolines, furoquinolines, thieno-quinoline and their effect on the ultrastructure of some pathogenic microorganisms. Acta Pol Pharm. 2001, 58, 175–184. [Google Scholar]
- Vandekerckhove, S.; Tran, H.G.; Desmet, T.; D’hooghe, M. Evaluation of (4-aminobutyloxy)quinolines as a novel class of antifungal agents. Bioorg. Med. Chem. Lett. 2013, 23, 4641–4643. [Google Scholar] [CrossRef]
- Hazra, A.; Mondal, S.; Maity, A.; Naskar, S.; Saha, P.; Paira, R.; Sahu, K.B.; Paira, P.; Ghosh, S.; Sinha, C.; et al. Amberlite-IRA-402 (OH) ion exchange resin mediated synthesis of indolizines, pyrrolo [1,2-a] quinolines and isoquinolines: Antibacterial and antifungal evaluation of the products. Eur. J. Med. Chem. 2011, 46, 2132–2140. [Google Scholar] [CrossRef]
- Delattin, N.; Bardiot, D.; Marchand, A.; Chaltin, P.; De Brucker, K.; Cammue, B.P.A.; Thevissen, K. Identification of fungicidal 2,6-disubstituted quinolines with activity against candida biofilms. Molecules 2012, 17, 12243–12251. [Google Scholar] [CrossRef]
- Mrozek-Wilczkiewicz, A.; Spaczynska, E.; Malarz, K.; Cieslik, W.; Rams-Baron, M.; Kryštof, V.; Musiol, R. Design, Synthesis and In Vitro Activity of Anticancer Styrylquinolines The p53 Independent Mechanism of Action. PLoS ONE 2015, 10, e0142678. [Google Scholar] [CrossRef]
- Mekouar, K.; Mouscadet, J.F.; Desmaële, D.; Subra, F.; Leh, H.; Savouré, D.; Auclair, C.; d’Angelo, J. Styrylquinoline derivatives: A new class of potent HIV-1 integrase inhibitors that block HIV-1 replication in CEM cells. J. Med. Chem. 1998, 41, 2846–2857. [Google Scholar] [CrossRef]
- Zouhiri, F.; Danet, M.; Bénard, C.; Normand-Bayle, M.; Mouscadet, J.-F.; Leh, H.; Marie Thomas, C.; Mbemba, G.; D’Angelo, J.; Desmaële, D. HIV-1 replication inhibitors of the styrylquinoline class: Introduction of an additional carboxyl group at the C-5 position of the quinoline. Tetrahedron Lett. 2005, 46, 2201–2205. [Google Scholar] [CrossRef]
- Zouhiri, F.; Mouscadet, J.F.; Mekouar, K.; Desmaële, D.; Savouré, D.; Leh, H.; Subra, F.; Le Bret, M.; Auclair, C.; d’Angelo, J. Structure-activity relationships and binding mode of styrylquinolines as potent inhibitors of HIV-1 integrase and replication of HIV-1 in cell culture. J. Med. Chem. 2000, 43, 1533–1540. [Google Scholar] [CrossRef]
- Machura, B.; Wolff, M.; Kowalczyk, W.; Musiol, R. Novel rhenium(V) complexes of 8-hydroxyquinoline derivatives—Synthesis, spectroscopic characterization, X-ray structure and DFT calculations. Polyhedron 2012, 33, 388–395. [Google Scholar] [CrossRef]
- Machura, B.; Wolff, M.; Cieślik, W.; Musioł, R. Novel oxorhenium(V) complexes of 8-hydroxyquinoline derivatives—Synthesis, spectroscopic characterization, X-ray crystal structures and DFT calculations. Polyhedron 2013, 51, 263–274. [Google Scholar] [CrossRef]
- Musiol, R. Styrylquinoline—A versatile scaffold in medicinal chemistry. Med. Chem. 2019, 15. [Google Scholar] [CrossRef]
- Musiol, R.; Podeszwa, B.; Finster, J.; Niedbala, H.; Polanski, J. An Efficient Microwave-Assisted Synthesis of Structurally Diverse Styrylquinolines. Mon. Chem. Chem. Mon. 2006, 137, 1211–1217. [Google Scholar] [CrossRef]
- Musiol, R.; Mrozek-Wilczkiewicz, A.; Polanski, J. Synergy against fungal pathogens: Working together is better than working alone. Curr. Med. Chem. 2014, 21, 870–893. [Google Scholar] [CrossRef]
- Henry, K.W.; Cruz, M.C.; Katiyar, S.K.; Edlind, T.D. Antagonism of azole activity against Candida albicans following induction of multidrug resistance genes by selected antimicrobial agents. Antimicrob. Agents Chemother. 1999, 43, 1968–1974. [Google Scholar] [CrossRef]
- Szczepaniak, J.; Łukaszewicz, M.; Krasowska, A. Estimation of Candida albicans ABC Transporter Behavior in Real-Time via Fluorescence. Front. Microbiol. 2015, 6, 1382. [Google Scholar] [CrossRef]
- Hibbert, F.; Spiers, K.J. Kinetics of hydrolysis of 1-Acetoxy-, 1-Acetoxy-8-hydroxy-, and 1,8-diacetoxy-naphthalenes; intermolecular participation by a hydroxy group. J. Chem. Soc. Perkin Trans. 1988, 20, 571–574. [Google Scholar] [CrossRef]
- Cipiciani, A.; Fringuelli, F.; Scappini, A.M. Enzymatic hydrolyses of acetoxy- and phenethylbenzoates by Candida cylindracea lipase. Tetrahedron 1996, 52, 9869–9876. [Google Scholar] [CrossRef]
- Rams-Baron, M.; Dulski, M.; Mrozek-Wilczkiewicz, A.; Korzec, M.; Cieslik, W.; Spaczyńska, E.; Bartczak, P.; Ratuszna, A.; Polanski, J.; Musiol, R. Synthesis of New Styrylquinoline Cellular Dyes, Fluorescent Properties, Cellular Localization and Cytotoxic Behavior. PLoS ONE 2015, 10, e0131210. [Google Scholar] [CrossRef]
- Musiol, R.; Jampilek, J.; Nycz, J.E.; Pesko, M.; Carroll, J.; Kralova, K.; Vejsova, M.; O’Mahony, J.; Coffey, A.; Mrozek, A.; et al. Investigating the activity spectrum for ring-substituted 8-hydroxyquinolines. Molecules 2010, 15, 288–304. [Google Scholar] [CrossRef]
- Evensen, N.A.; Braun, P.C. The effects of tea polyphenols on Candida albicans: Inhibition of biofilm formation and proteasome inactivation. Can. J. Microbiol. 2009, 55, 1033–1039. [Google Scholar] [CrossRef]
- Shahzad, M.; Sherry, L.; Rajendran, R.; Edwards, C.A.; Combet, E.; Ramage, G. Utilising polyphenols for the clinical management of Candida albicans biofilms. Int. J. Antimicrob. Agents 2014, 44, 269–273. [Google Scholar] [CrossRef]
- Rane, H.S.; Bernardo, S.M.; Howell, A.B.; Lee, S.A. Cranberry-derived proanthocyanidins prevent formation of Candida albicans biofilms in artificial urine through biofilm- and adherence-specific mechanisms. J. Antimicrob. Chemother. 2014, 69, 428–436. [Google Scholar] [CrossRef]
- Ahmad, A.; Khan, A.; Manzoor, N.; Khan, L.A. Evolution of ergosterol biosynthesis inhibitors as fungicidal against Candida. Microb. Pathog. 2010, 48, 35–41. [Google Scholar] [CrossRef]
- Mahata, D.; Mandal, S.M.; Bharti, R.; Gupta, V.K.; Mandal, M.; Nag, A.; Nando, G.B. Self-assembled cardanol azo derivatives as antifungal agent with chitin-binding ability. Int. J. Biol. Macromol. 2014, 69, 5–11. [Google Scholar] [CrossRef]
- Szczepaniak, J.; Łukaszewicz, M.; Krasowska, A. Detection of inhibitors of Candida albicans Cdr transporters using a diS-C3(3) fluorescence. Front. Microbiol. 2015, 6, 176. [Google Scholar] [CrossRef]
- Sharma, M.; Manoharlal, R.; Shukla, S.; Puri, N.; Prasad, T.; Ambudkar, S.V.; Prasad, R. Curcumin modulates efflux mediated by yeast ABC multidrug transporters and is synergistic with antifungals. Antimicrob. Agents Chemother. 2009, 53, 3256–3265. [Google Scholar] [CrossRef]
- Shapiro, H.M. Practical Flow Cytometry; Wiley: Hoboken, NJ, USA, 2003; ISBN 9780471411253. [Google Scholar]
- Dolab, J.; Lima, B.; Spaczynska, E.; Kos, J.; Cano, N.; Feresin, G.; Tapia, A.; Garibotto, F.; Petenatti, E.; Olivella, M.; et al. The Antimicrobial Activity of Annona emarginata (Schltdl.) H. Rainer and Most Active Isolated Compounds against Clinically Important Bacteria. Molecules 2018, 23, 1187. [Google Scholar] [CrossRef]
- Fonzi, W.; Irwin, M. Isogenic strain construction and gene mapping in Candida albicans. Genetics 1993, 134, 717–728. [Google Scholar]
- Sanglard, D.; Ischer, F. Susceptibilities of Candida albicans multidrug transporter mutants to various antifungal agents and other metabolic inhibitors. Antimicrob. Agents 1996, 40, 2300–2305. [Google Scholar] [CrossRef]
- Sanglard, D.; Ischer, F.; Monod, M.; Bille, J. Cloning of Candida albicans genes conferring resistance to azole antifungal agents: Characterization of CDR2, a new multidrug ABC transporter gene. Microbiology 1997, 143, 405–416. [Google Scholar] [CrossRef]
- Hall, M.J.; Middleton, R.F.; Westmacott, D. The fractional inhibitory concentration (FIC) index as a measure of synergy. J. Antimicrob. Chemother. 1983, 11, 427–433. [Google Scholar] [CrossRef]
- Hiller, D.; Sanglard, D.; Morschhäuser, J. Overexpression of the MDR1 gene is sufficient to confer increased resistance to toxic compounds in Candida albicans. Antimicrob. Agents Chemother. 2006, 50, 1365–1371. [Google Scholar] [CrossRef]
- Nakamura, K.; Niimi, M.; Niimi, K.; Holmes, A.R.; Yates, J.E.; Decottignies, A.; Monk, B.C.; Goffeau, A.; Cannon, R.D. Functional expression of Candida albicans drug efflux pump Cdr1p in a Saccharomyces cerevisiae strain deficient in membrane transporters. Antimicrob. Agents Chemother. 2001, 45, 3366. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).