Adlay (Coix lachryma-jobi L. var. ma-yuen Stapf.) Hull Extract and Active Compounds Inhibit Proliferation of Primary Human Leiomyoma Cells and Protect against Sexual Hormone-Induced Mice Smooth Muscle Hyperproliferation




Abstract
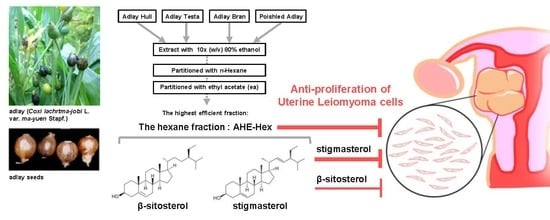
1. Introduction
2. Results
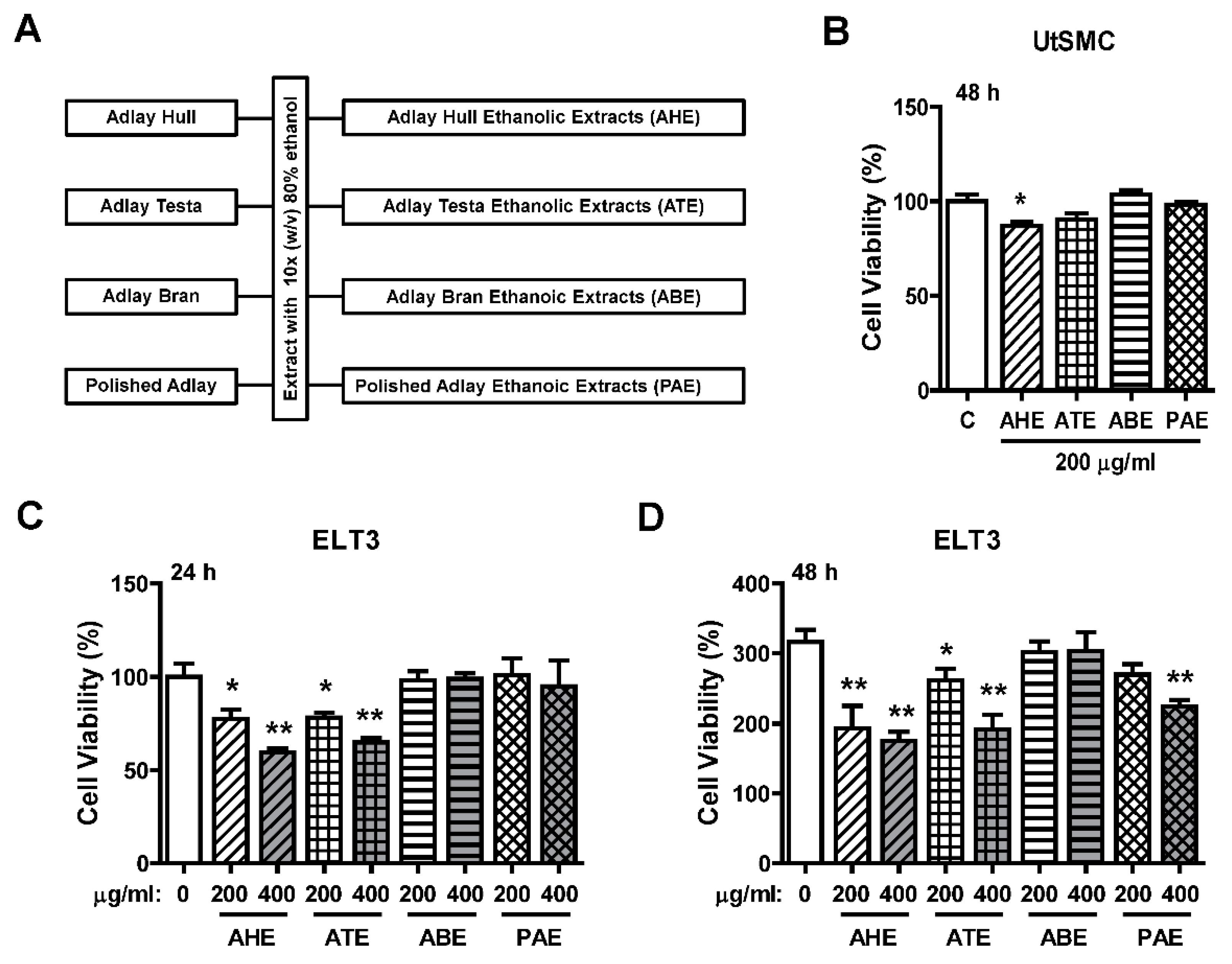
2.1. Effects of the Ethanolic Extracts from Four Parts of Adlay Seed on ELT3 Cell Viability
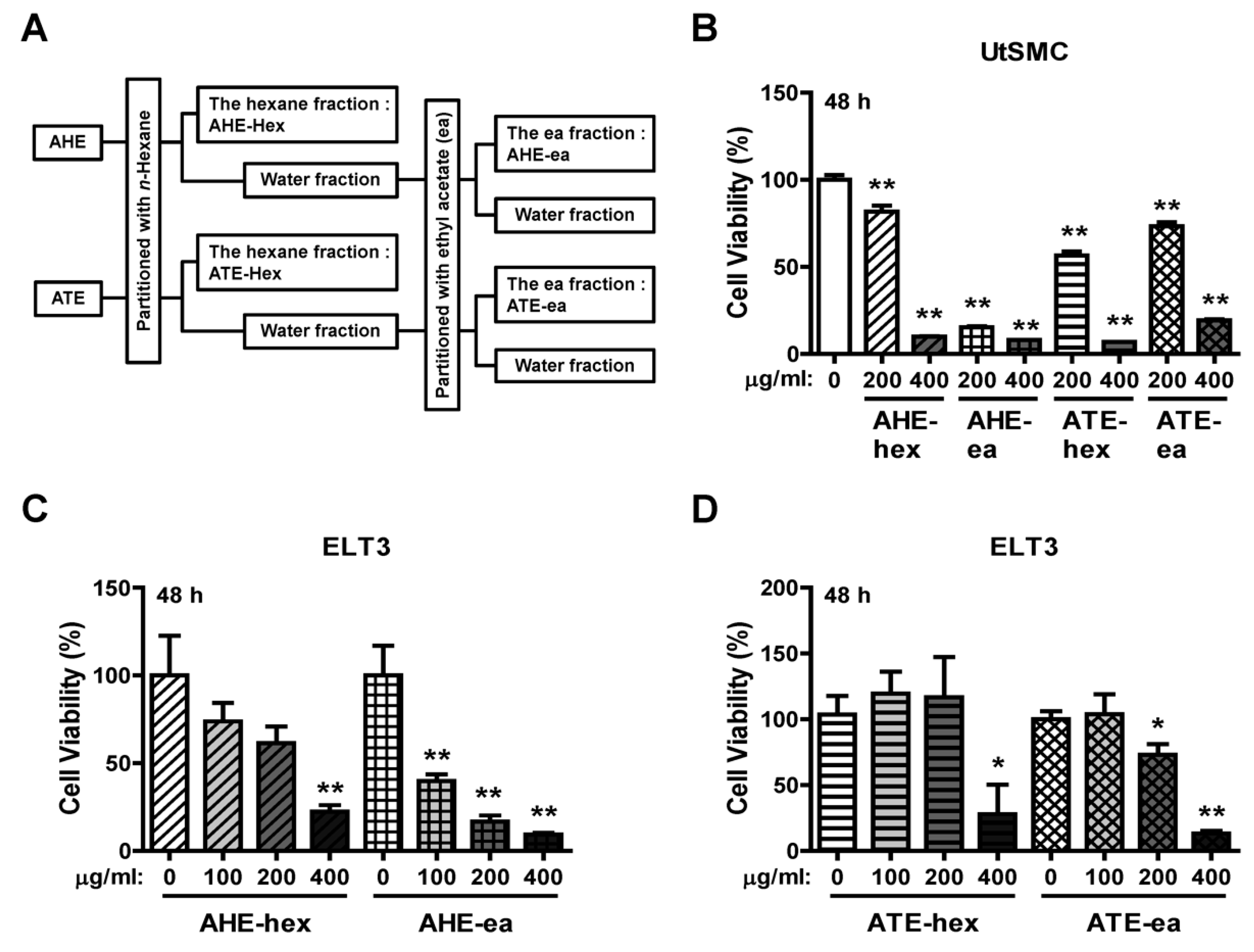
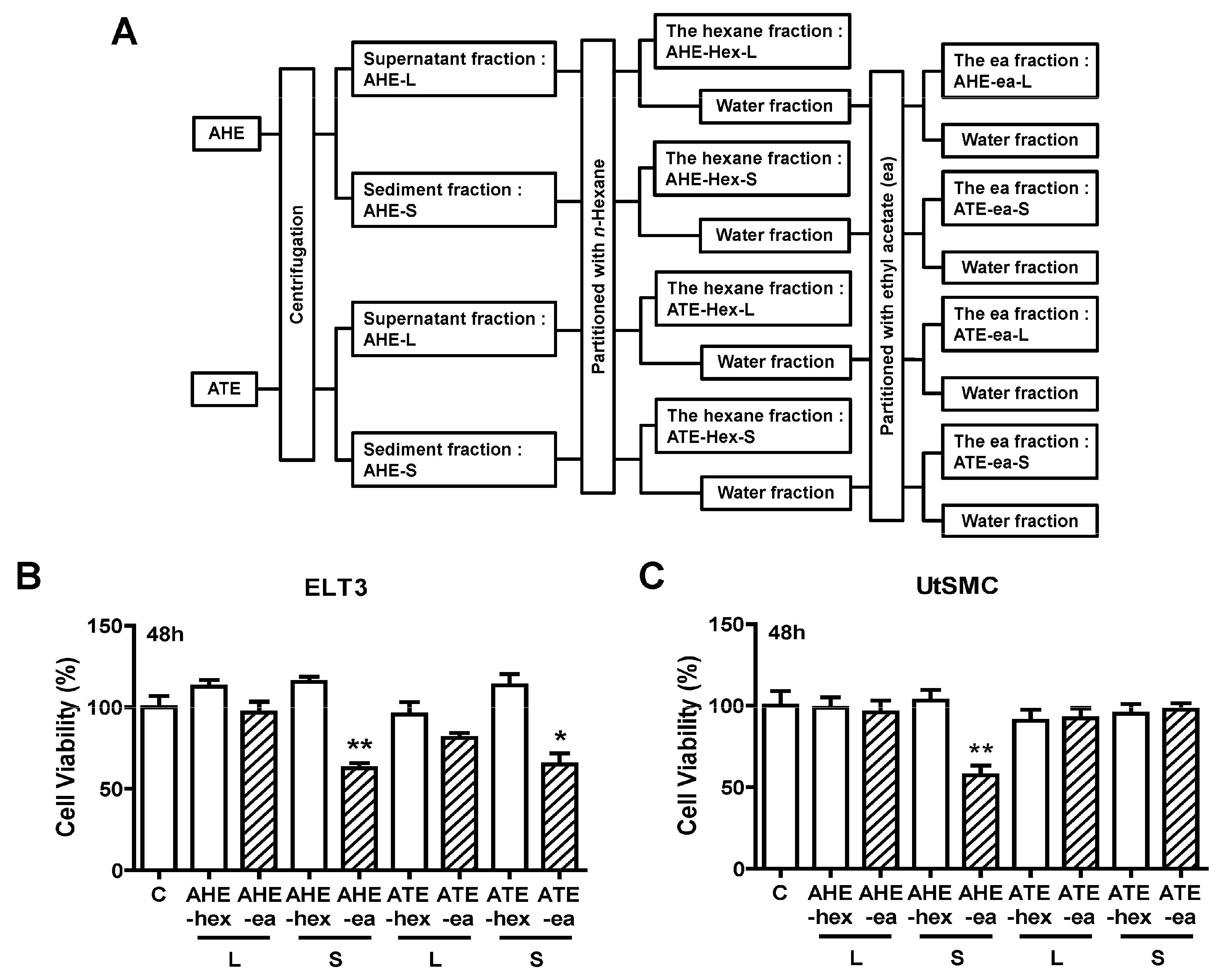
2.2. Effects of Each Fraction of AHE and ATE on ELT3 Cell Viability
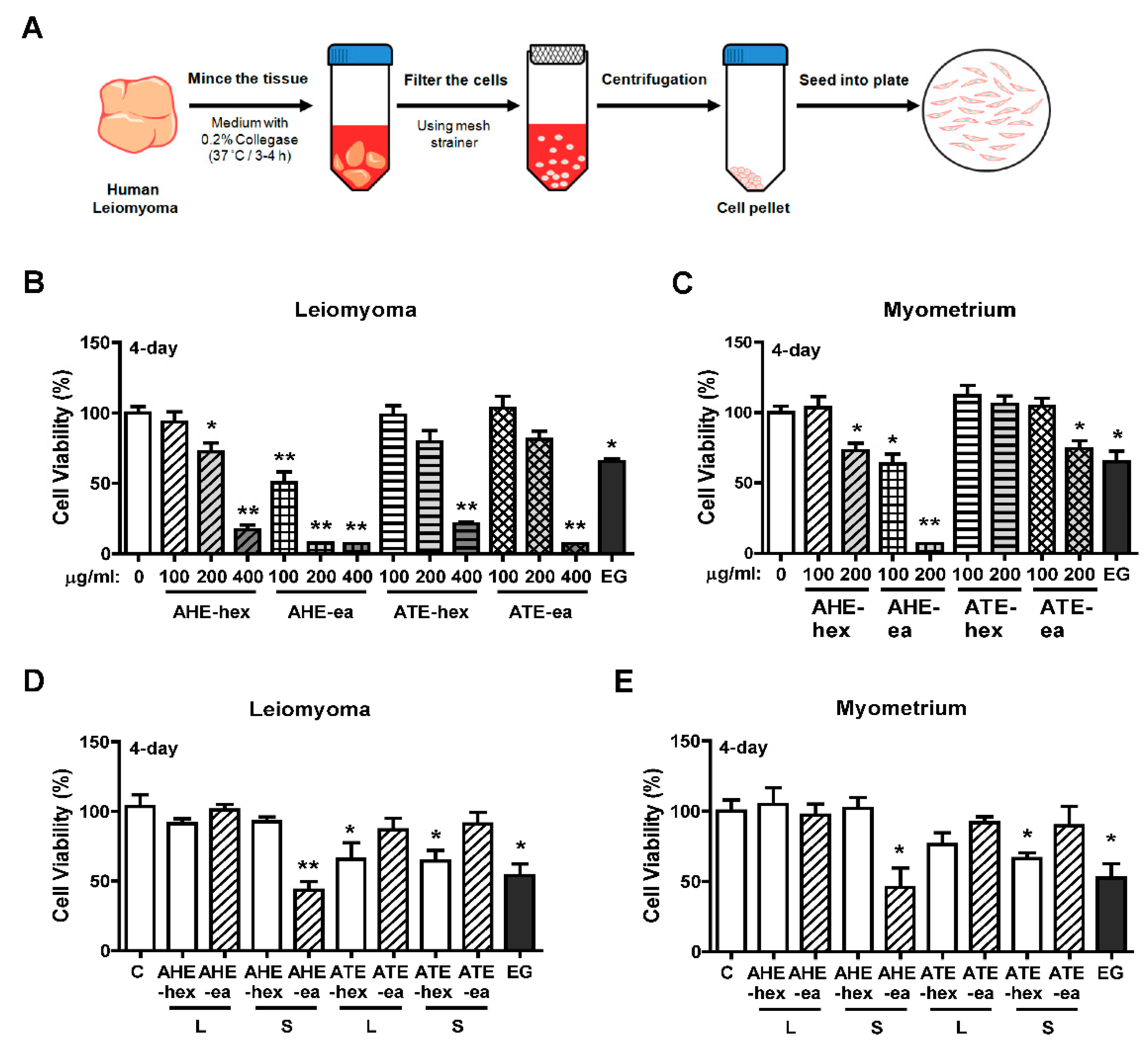
2.3. Effects of Each Fraction of AHE and ATE on Primary Human Uterine Leiomyoma (hUL) Cells
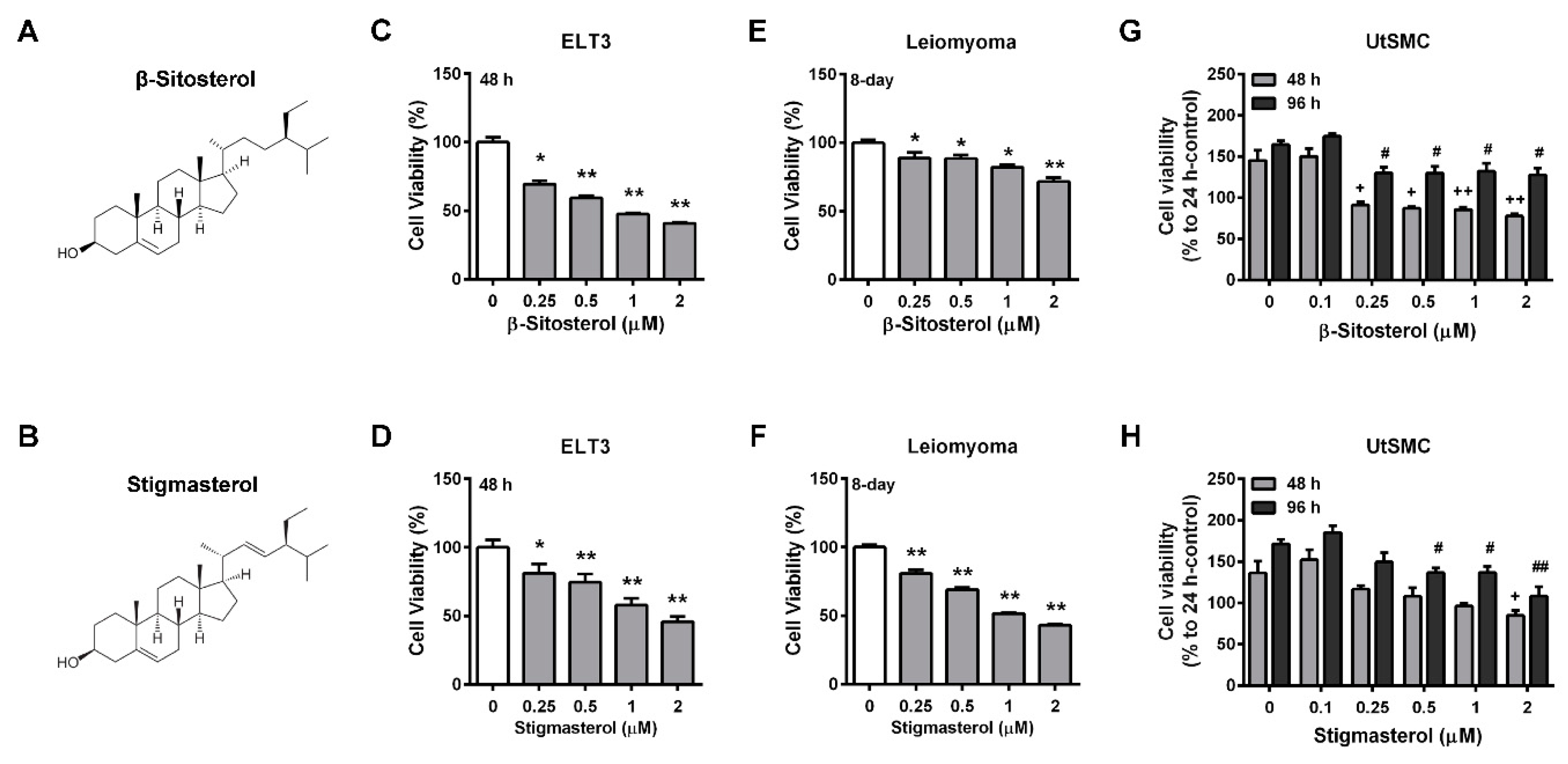
2.4. Identification of Potential Effective Compounds in the Ethyl Acetate Fraction of AHE (AHE-ea)
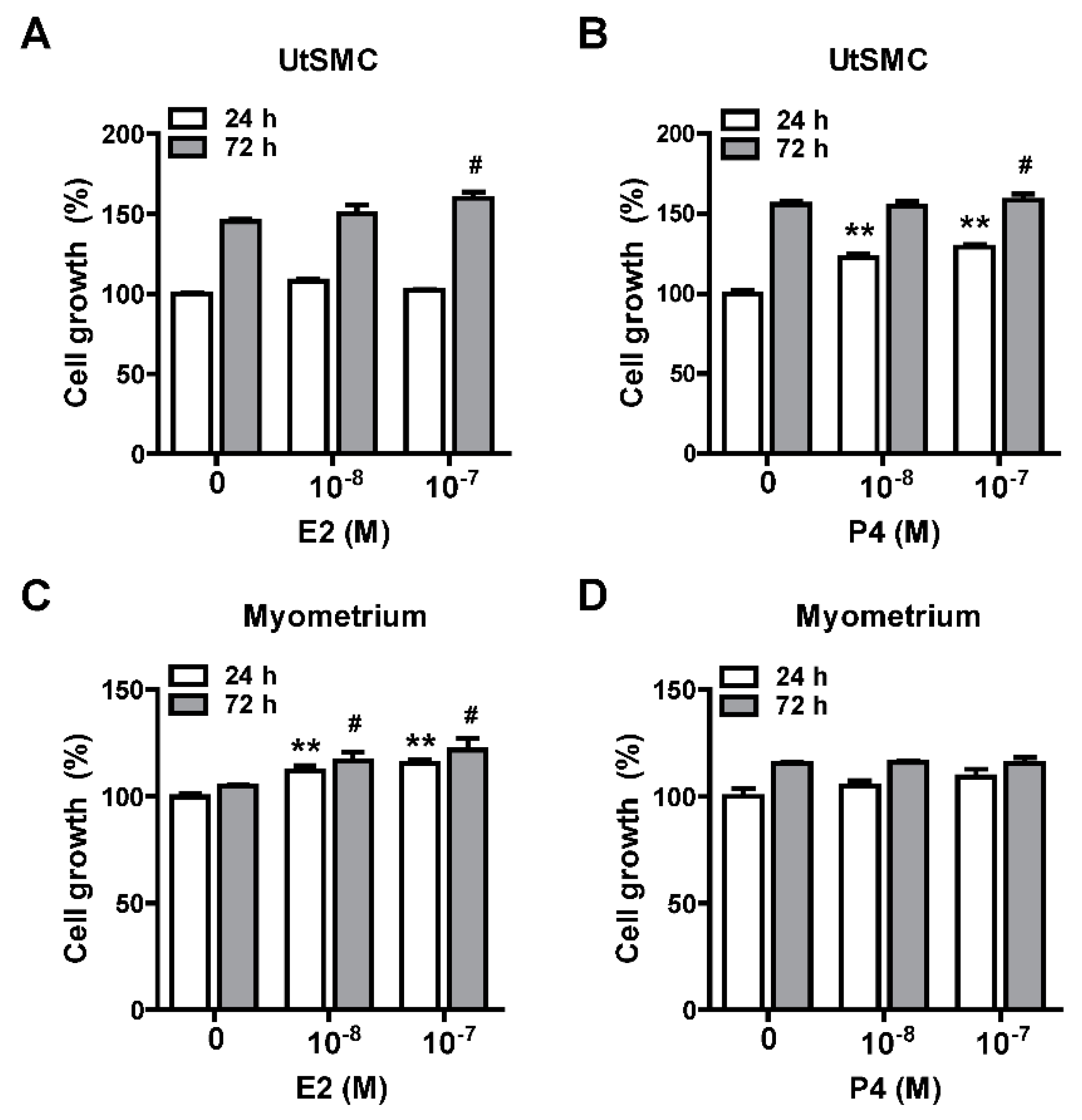
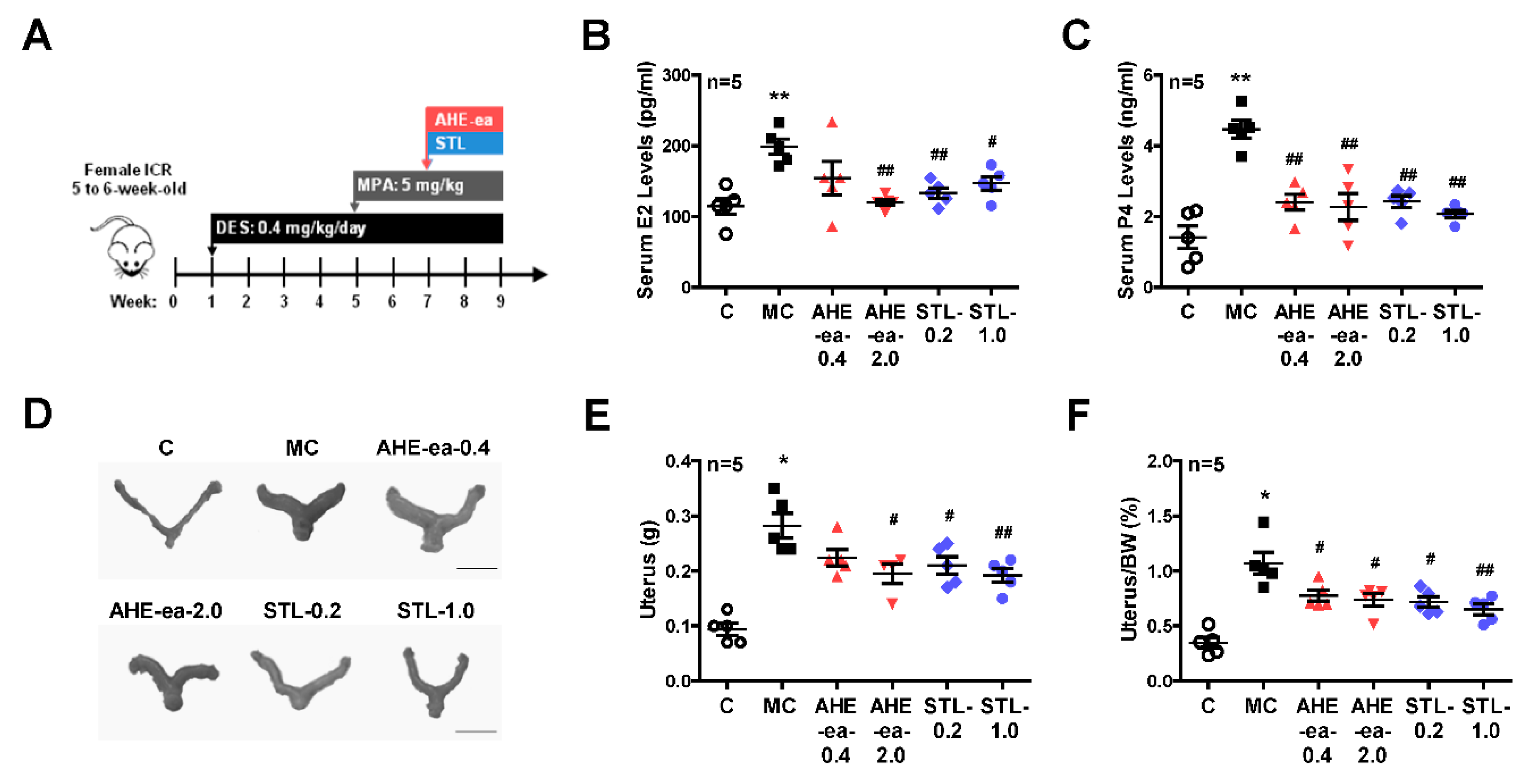
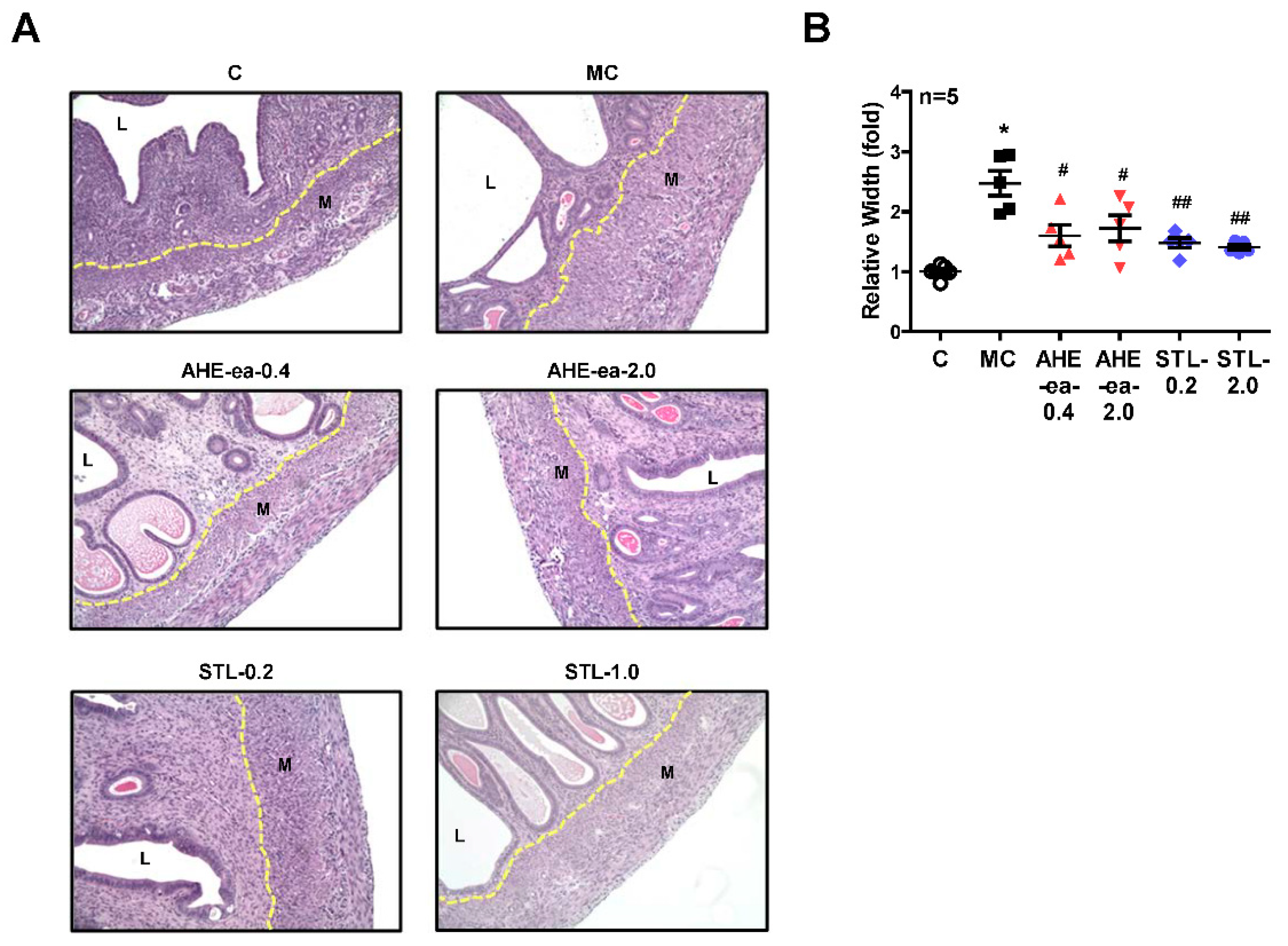
2.5. Inhibition of Uterine Myometrial Growth by AHE-ea Fraction In Vivo
3. Discussion
4. Materials and Methods
4.1. Preparation of Ethanolic Extracts from Each Part of Adlay Seed
4.2. Preparation of Various Fractions from Ethanolic Extracts of Adlay Hull (AHE) and Testa (ATE).
4.3. Cell Lines and Culture Conditions
4.4. Preparation of Primary Human Uterine Leiomyoma (hUL) Cells and Human Uterine Myometrial (hUM) Cells
4.5. Cell Survival Assay
4.6. High-Performance Liquid Chromatography (HPLC) Analysis
4.7. Animals
4.8. Enzyme-Linked Immunosorbent Assay (ELISA) of Serum E2 and P4
4.9. H&E Staining
4.10. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Stewart, E.A. Uterine fibroids. Lancet 2001, 357, 293–298. [Google Scholar] [CrossRef]
- Stewart, E.A.; Laughlin-Tommaso, S.K.; Catherino, W.H.; Lalitkumar, S.; Gupta, D.; Vollenhoven, B. Uterine fibroids. Nat. Rev. Dis. Primers 2016, 2, 16043. [Google Scholar] [CrossRef]
- Vilos, G.A.; Allaire, C.; Laberge, P.-Y.; Leyland, N.; Vilos, A.G.; Murji, A.; Chen, I. The Management of Uterine Leiomyomas. J. Obstet. Gynaecol. Can. 2015, 37, 157–178. [Google Scholar] [CrossRef]
- Stewart, E.A. Clinical practice. Uterine fibroids. N. Engl. J. Med. 2015, 372, 1646–1655. [Google Scholar] [CrossRef]
- Stewart, E.A.; Cookson, C.L.; Gandolfo, R.A.; Schulze-Rath, R. Epidemiology of uterine fibroids: A systematic review. BJOG 2017, 124, 1501–1512. [Google Scholar] [CrossRef] [PubMed]
- Fuldeore, M.J.; Soliman, A.M. Patient-reported prevalence and symptomatic burden of uterine fibroids among women in the United States: Findings from a cross-sectional survey analysis. Int. J. Women’s Health 2017, 9, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, A.; Bernuit, D.; Gerlinger, C.; Schaefers, M.; Geppert, K. Prevalence, symptoms and management of uterine fibroids: An international internet-based survey of 21,746 women. BMC Women’s Health 2012, 12, 6. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.K.; Leppert, P.C. Treatment for Uterine Fibroids: Searching for Effective Drug Therapies. Drug Discov. Today Ther. Strateg. 2012, 9, e41–e49. [Google Scholar] [CrossRef] [PubMed]
- Lethaby, A.; Vollenhoven, B. Fibroids (uterine myomatosis, leiomyomas). BMJ Clin. Evid. 2015, 2015. [Google Scholar]
- Greenwell, M.; Rahman, P. Medicinal Plants: Their Use in Anticancer Treatment. Int. J. Pharm. Sci. Res. 2015, 6, 4103–4112. [Google Scholar]
- Tsai, W.H.; Yang, C.C.; Li, P.C.; Chen, W.C.; Chien, C.T. Therapeutic potential of traditional chinese medicine on inflammatory diseases. J. Tradit. Complement. Med. 2013, 3, 142–151. [Google Scholar] [CrossRef]
- Yen, H.R.; Chen, Y.Y.; Huang, T.P.; Chang, T.T.; Tsao, J.Y.; Chen, B.C.; Sun, M.F. Prescription patterns of Chinese herbal products for patients with uterine fibroid in Taiwan: A nationwide population-based study. J. Ethnopharmacol. 2015, 171, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.-C.; Chen, H.-H.; Chiang, W. Adlay (yì yĭ; “soft-shelled job’s tears”; the seeds of Coix lachryma-jobi L. var. ma-yuen Stapf) is a Potential Cancer Chemopreventive Agent toward Multistage Carcinogenesis Processes. J. Tradit. Complement. Med. 2012, 2, 267–275. [Google Scholar] [CrossRef]
- Wang, L.; Chen, J.; Xie, H.; Ju, X.; Liu, R.H. Phytochemical profiles and antioxidant activity of adlay varieties. J. Agric. Food Chem. 2013, 61, 5103–5113. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Chen, L.; Ali, B.; Yang, N.; Chen, Y.; Wu, F.; Jin, Z.; Xu, X. Impact of germination on nutritional and physicochemical properties of adlay seed (Coixlachryma-jobi L.). Food Chem. 2017, 229, 312–318. [Google Scholar] [CrossRef]
- Yao, Y.; Zhu, Y.; Gao, Y.; Ren, G. Effect of ultrasonic treatment on immunological activities of polysaccharides from adlay. Int. J. Biol. Macromol. 2015, 80, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Kato, M.; Ayugase, J. Anti-diabetic Effects of Adlay Protein in Type 2 Diabetic Mice. Food Sci. Technol. Res. 2012, 18, 383–390. [Google Scholar] [CrossRef]
- Yu, F.; Gao, J.; Zeng, Y.; Liu, C.X. Effects of adlay seed oil on blood lipids and antioxidant capacity in hyperlipidemic rats. J. Sci. Food Agric. 2011, 91, 1843–1848. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.J.; Lo, Y.C.; Chiang, W. Inhibitory effects of adlay bran (Coix lachryma-jobi L. var. ma-yuen Stapf) on chemical mediator release and cytokine production in rat basophilic leukemia cells. J. Ethnopharmacol. 2012, 141, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.P.; Hsu, C.Y.; Lin, J.H.; Kuo, Y.H.; Chiang, W.; Lin, Y.L. Antiproliferative lactams and spiroenone from adlay bran in human breast cancer cell lines. J. Agric. Food Chem. 2011, 59, 1185–1194. [Google Scholar] [CrossRef]
- Lu, X.; Liu, W.; Wu, J.; Li, M.; Wang, J.; Wu, J.; Luo, C. A polysaccharide fraction of adlay seed (Coixlachryma-jobi L.) induces apoptosis in human non-small cell lung cancer A549 cells. Biochem. Biophys. Res. Commun. 2013, 430, 846–851. [Google Scholar] [CrossRef] [PubMed]
- Li, S.C.; Chen, C.M.; Lin, S.H.; Chiang, W.; Shih, C.K. Effects of adlay bran and its ethanolic extract and residue on preneoplastic lesions of the colon in rats. J. Sci. Food Agric. 2011, 91, 547–552. [Google Scholar] [CrossRef]
- Shi, G.; Zheng, X.; Zhang, S.; Wu, X.; Yu, F.; Wang, Y.; Xing, F. Kanglaite inhibits EMT caused by TNF-α via NF-κB inhibition in colorectal cancer cells. Oncotarget 2017, 9, 6771–6779. [Google Scholar] [PubMed]
- Wang, Y.; Zhang, C.; Zhang, S.; Zhao, Z.; Wang, J.; Song, J.; Wang, Y.; Liu, J.; Hou, S. Author Correction: Kanglaite sensitizes colorectal cancer cells to Taxol via NF-kappaBeta inhibition and connexin 43 upregulation. Sci. Rep. 2018, 7, 1280. [Google Scholar] [CrossRef]
- Schwartzberg, L.S.; Arena, F.P.; Bienvenu, B.J.; Kaplan, E.H.; Camacho, L.H.; Campos, L.T.; Waymack, J.P.; Tagliaferri, M.A.; Chen, M.M.; Li, D. A Randomized, Open-Label, Safety and Exploratory Efficacy Study of Kanglaite Injection (KLTi) plus Gemcitabine versus Gemcitabine in Patients with Advanced Pancreatic Cancer. J. Cancer 2017, 8, 1872–1883. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yang, Q.; Xi, Y.; Yu, K.; Wang, W.; Zhao, X.; Kou, X. Kanglaite injection combined with chemotherapy versus chemotherapy alone in the treatment of advanced non-small cell lung carcinoma. J. Cancer Res. Ther. 2014, 10, 46–51. [Google Scholar] [PubMed]
- Ding, H. Clinical application effect evaluation of Kanglaite combined with DCF chemotherapy program in patients with gastric cancer. Proc. Anticancer Res. 2017, 1, 7–12. [Google Scholar] [CrossRef][Green Version]
- Li, S. Ben Cao Gang Mu (Systematic Pharmacopeia); Chinese Medical Technology Press: Beijing, China, 2001. [Google Scholar]
- Tzeng, H.P.; Chiang, W.; Ueng, T.H.; Liu, S.H. The abortifacient effects from the seeds of Coix lachryma-jobi L. var. ma-yuen Stapf. J. Toxicol. Environ. Health A 2005, 68, 1557–1565. [Google Scholar] [CrossRef]
- Hsia, S.M.; Kuo, Y.H.; Chiang, W.; Wang, P.S. Effects of adlay hull extracts on uterine contraction and Ca2+ mobilization in the rat. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E719–E726. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Huang, L.H.; Chiang, W.; Hsia, S.M. Hexane fraction of adlay (Coix lachryma-jobi L.) testa ethanolic extract inhibits human uterine sarcoma cancer cells growth and chemosensitizes human uterine sarcoma cells to doxorubicin. Phytomedicine 2018, 47, 69–80. [Google Scholar] [CrossRef]
- Cheung CS, C.L. Preliminary report of 136 cases of uterine myoma treated by gong zheng tang. J. Am. Coll. Tradit. Chin. Med. 1982, 1, 64–66. [Google Scholar]
- Zhang, D.; Al-Hendy, M.; Richard-Davis, G.; Montgomery-Rice, V.; Rajaratnam, V.; Al-Hendy, A. Antiproliferative and Proapoptotic Effects of Epigallocatechin Gallate on Human Leiomyoma Cells. Fertil. Steril. 2010, 94, 1887–1893. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Al-Hendy, M.; Richard-Davis, G.; Montgomery-Rice, V.; Sharan, C.; Rajaratnam, V.; Khurana, A.; Al-Hendy, A. Green tea extract inhibits proliferation of uterine leiomyoma cells in vitro and in nude mice. Am. J. Obstet. Gynecol. 2010, 202, 289 e1-9. [Google Scholar] [CrossRef] [PubMed]
- Bulun, S.E. Uterine fibroids. N. Engl. J. Med. 2013, 369, 1344–1355. [Google Scholar] [CrossRef]
- Newcomb, P.M.; Cramer, S.F.; Leppert, P.C. Myometrial hyperplasia mimics the clinical presentation of uterine fibroids: A report of 3 cases. Int. J. Gynecol. Pathol. 2013, 32, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Buttram, V.C., Jr.; Reiter, R.C. Uterine leiomyomata: Etiology, symptomatology, and management. Fertil. Steril. 1981, 36, 433–445. [Google Scholar] [PubMed]
- Islam, M.S.; Protic, O.; Giannubilo, S.R.; Toti, P.; Tranquilli, A.L.; Petraglia, F.; Castellucci, M.; Ciarmela, P. Uterine leiomyoma: Available medical treatments and new possible therapeutic options. J. Clin. Endocrinol. Metab. 2013, 98, 921–934. [Google Scholar] [CrossRef]
- Kim, D.C.; Ramachandran, S.; Baek, S.H.; Kwon, S.H.; Kwon, K.Y.; Cha, S.D.; Bae, I.; Cho, C.H. Induction of growth inhibition and apoptosis in human uterine leiomyoma cells by isoliquiritigenin. Reprod. Sci. 2008, 15, 552–558. [Google Scholar] [CrossRef]
- Kim, C.-W.; Hwang, K.-A.; Choi, K.-C. Anti-metastatic potential of resveratrol and its metabolites by the inhibition of epithelial-mesenchymal transition, migration, and invasion of malignant cancer cells. Phytomedicine 2016, 23, 1787–1796. [Google Scholar] [CrossRef]
- Wu, C.-H.; Shieh, T.-M.; Wei, L.-H.; Cheng, T.-F.; Chen, H.-Y.; Huang, T.-C.; Wang, K.-L.; Hsia, S.-M. Resveratrol inhibits proliferation of myometrial and leiomyoma cells and decreases extracellular matrix-associated protein expression. J. Funct. Foods 2016, 23, 241–252. [Google Scholar] [CrossRef]
- Hsia, S.M.; Lin, K.H.; Chiang, W.; Wu, C.H.; Shieh, T.M.; Huang, T.C.; Chen, H.Y.; Lin, L.C. Effects of Adlay Hull and Testa Ethanolic Extracts on the Growth of Uterine Leiomyoma Cells. In Adaptive Medicine; Airiti Press: Taipei, Taiwan, 2017; Volume 9, pp. 85–96. [Google Scholar]
- Wong, J.Y.; Gold, E.B.; Johnson, W.O.; Lee, J.S. Circulating Sex Hormones and Risk of Uterine Fibroids: Study of Women’s Health Across the Nation (SWAN). J. Clin. Endocrinol. Metab. 2016, 101, 123–130. [Google Scholar] [CrossRef]
- Ishikawa, H.; Ishi, K.; Serna, V.A.; Kakazu, R.; Bulun, S.E.; Kurita, T. Progesterone is essential for maintenance and growth of uterine leiomyoma. Endocrinology 2010, 151, 2433–2442. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Sefton, E.C. The role of progesterone signaling in the pathogenesis of uterine leiomyoma. Mol. Cell. Endocrinol. 2012, 358, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Templeman, C.; Marshall, S.F.; Clarke, C.A.; Henderson, K.D.; Largent, J.; Neuhausen, S.; Reynolds, P.; Ursin, G.; Bernstein, L. Risk factors for surgically removed fibroids in a large cohort of teachers. Fertil. Steril. 2009, 92, 1436–1446. [Google Scholar] [CrossRef] [PubMed]
- Blake, R.E. Leiomyomata uteri: Hormonal and molecular determinants of growth. J. Natl. Med. Assoc. 2007, 99, 1170–1184. [Google Scholar] [PubMed]
- Hsia, S.M.; Yeh, C.L.; Kuo, Y.H.; Wang, P.S.; Chiang, W. Effects of adlay (Coix lachryma-jobi L. var. ma-yuen Stapf.) hull extracts on the secretion of progesterone and estradiol in vivo and in vitro. Exp. Biol. Med. (Maywood) 2007, 232, 1181–1194. [Google Scholar] [CrossRef] [PubMed]
- Kangsamaksin, T.; Chaithongyot, S.; Wootthichairangsan, C.; Hanchaina, R.; Tangshewinsirikul, C.; Svasti, J. Lupeol and stigmasterol suppress tumor angiogenesis and inhibit cholangiocarcinoma growth in mice via downregulation of tumor necrosis factor-α. PLoS ONE 2017, 12, e0189628. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Dixit, S.; Ali, D.; Alqahtani, S.M.; Alkahtani, S.; Alarifi, S. Isolation and evaluation of anticancer efficacy of stigmasterol in a mouse model of DMBA-induced skin carcinoma. Drug Des. Dev. Ther. 2015, 9, 2793–2800. [Google Scholar] [CrossRef]
- Hyder, S.M.; Huang, J.C.; Nawaz, Z.; Boettger-Tong, H.; Makela, S.; Chiappetta, C.; Stancel, G.M. Regulation of vascular endothelial growth factor expression by estrogens and progestins. Environ. Health Perspect. 2000, 108, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Tal, R.; Segars, J.H. The role of angiogenic factors in fibroid pathogenesis: Potential implications for future therapy. Hum. Reprod. Update 2014, 20, 194–216. [Google Scholar] [CrossRef]
- Wei, J.J.; Zhang, X.M.; Chiriboga, L.; Yee, H.; Perle, M.A.; Mittal, K. Spatial differences in biologic activity of large uterine leiomyomata. Fertil. Steril. 2006, 85, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Hsia, S.M.; Tseng, Y.W.; Wang, S.W.; Kuo, Y.H.; Huang, D.W.; Wang, P.S.; Chiang, W. Effect of adlay (Coix lachryma-jobi L. var. ma-yuen Stapf.) hull extracts on testosterone release from rat Leydig cells. Phytother. Res. 2009, 23, 687–695. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds isoliquiritigenin, β-sitosterol and stigmasterol are available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Weight (g) | Yield (%) |
|---|---|---|
| AHE-hex | 26.7 | 0.19 |
| AHE-ea | 18.1 | 0.12 |
| ATE-hex | 90.9 | 1.31 |
| ATE-ea | 50.6 | 0.73 |
| Compound | Weight (g) | Yield (%) |
|---|---|---|
| AHE-hex-L | 7.5 | 0.04 |
| AHE-ea-L | 18.9 | 0.09 |
| AHE-hex-S | 23.9 | 0.12 |
| AHE-ea-S | 10.2 | 0.05 |
| ATE-hex-L | 19.5 | 0.24 |
| ATE-ea-L | 9.9 | 0.12 |
| ATE-hex-S | 88.7 | 1.11 |
| ATE-ea-S | 34.3 | 0.43 |
| Sample | β-Sitosterol | Stigmasterol | Campesterol |
|---|---|---|---|
| AHE-ea | 522.80 | 216.24 | Trace |
| AHE-ea-L | 21.61 | 23.88 | Trace |
| AHE-ea-S | 2243.29 | 1417.84 | 588.74 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, P.-H.; Shih, C.-K.; Yen, Y.-T.; Chiang, W.; Hsia, S.-M. Adlay (Coix lachryma-jobi L. var. ma-yuen Stapf.) Hull Extract and Active Compounds Inhibit Proliferation of Primary Human Leiomyoma Cells and Protect against Sexual Hormone-Induced Mice Smooth Muscle Hyperproliferation. Molecules 2019, 24, 1556. https://doi.org/10.3390/molecules24081556
Lin P-H, Shih C-K, Yen Y-T, Chiang W, Hsia S-M. Adlay (Coix lachryma-jobi L. var. ma-yuen Stapf.) Hull Extract and Active Compounds Inhibit Proliferation of Primary Human Leiomyoma Cells and Protect against Sexual Hormone-Induced Mice Smooth Muscle Hyperproliferation. Molecules. 2019; 24(8):1556. https://doi.org/10.3390/molecules24081556
Chicago/Turabian StyleLin, Po-Han, Chun-Kuang Shih, Yu-Ting Yen, Wenchang Chiang, and Shih-Min Hsia. 2019. "Adlay (Coix lachryma-jobi L. var. ma-yuen Stapf.) Hull Extract and Active Compounds Inhibit Proliferation of Primary Human Leiomyoma Cells and Protect against Sexual Hormone-Induced Mice Smooth Muscle Hyperproliferation" Molecules 24, no. 8: 1556. https://doi.org/10.3390/molecules24081556
APA StyleLin, P.-H., Shih, C.-K., Yen, Y.-T., Chiang, W., & Hsia, S.-M. (2019). Adlay (Coix lachryma-jobi L. var. ma-yuen Stapf.) Hull Extract and Active Compounds Inhibit Proliferation of Primary Human Leiomyoma Cells and Protect against Sexual Hormone-Induced Mice Smooth Muscle Hyperproliferation. Molecules, 24(8), 1556. https://doi.org/10.3390/molecules24081556