
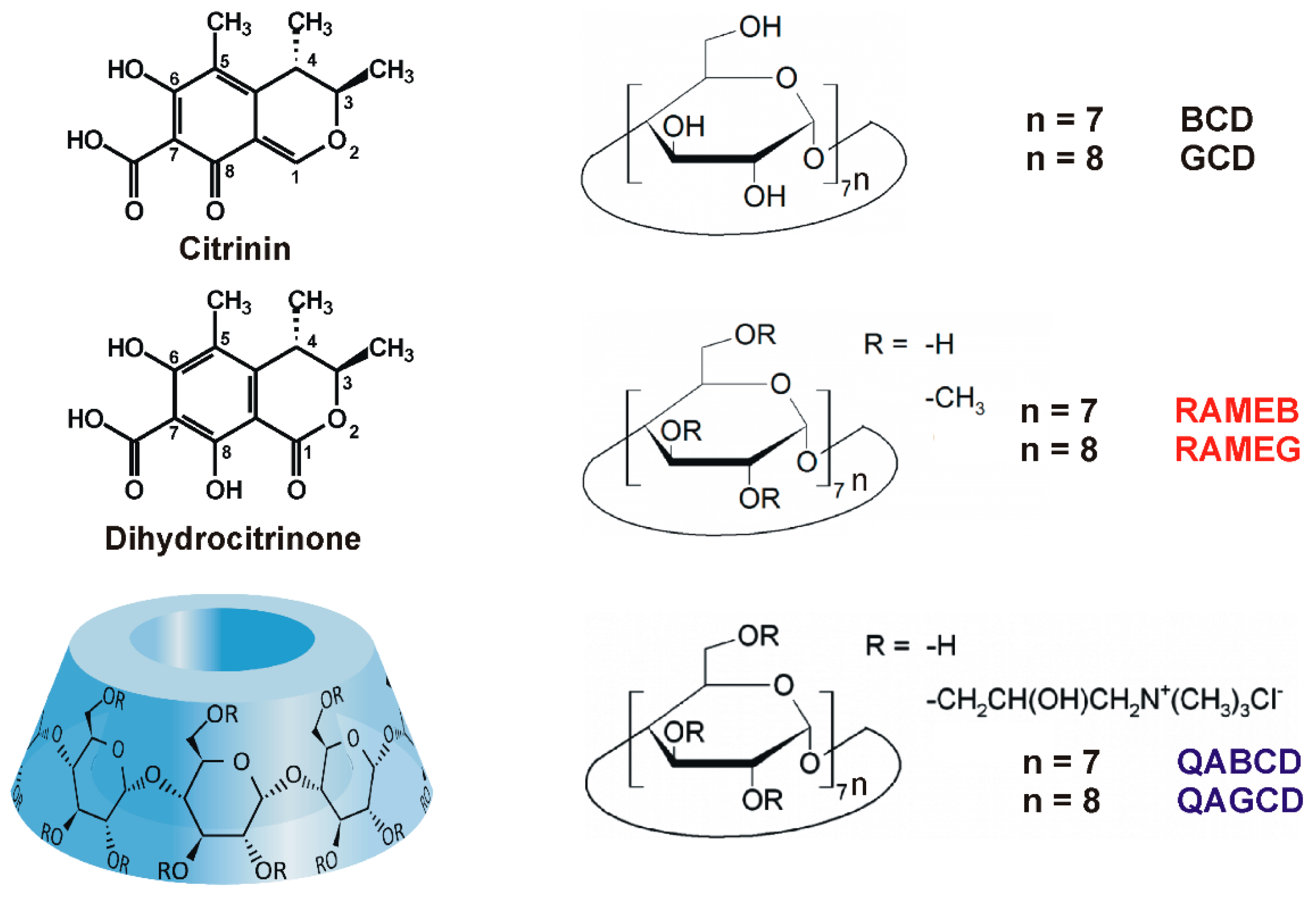
Interaction of Dihydrocitrinone with Native and Chemically Modified Cyclodextrins

 and
and 
Abstract
1. Introduction
2. Results and Discussion
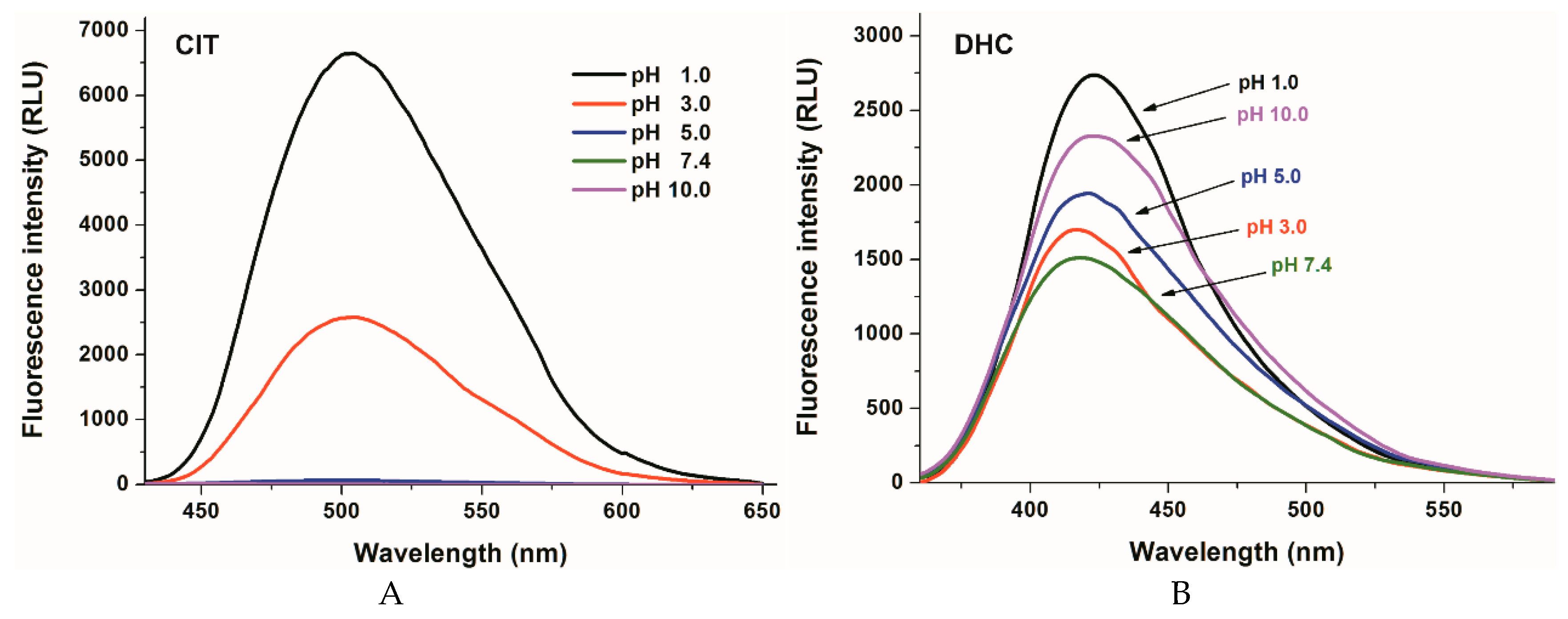
2.1. Fluorescence Properties of DHC and CIT in Different Buffers
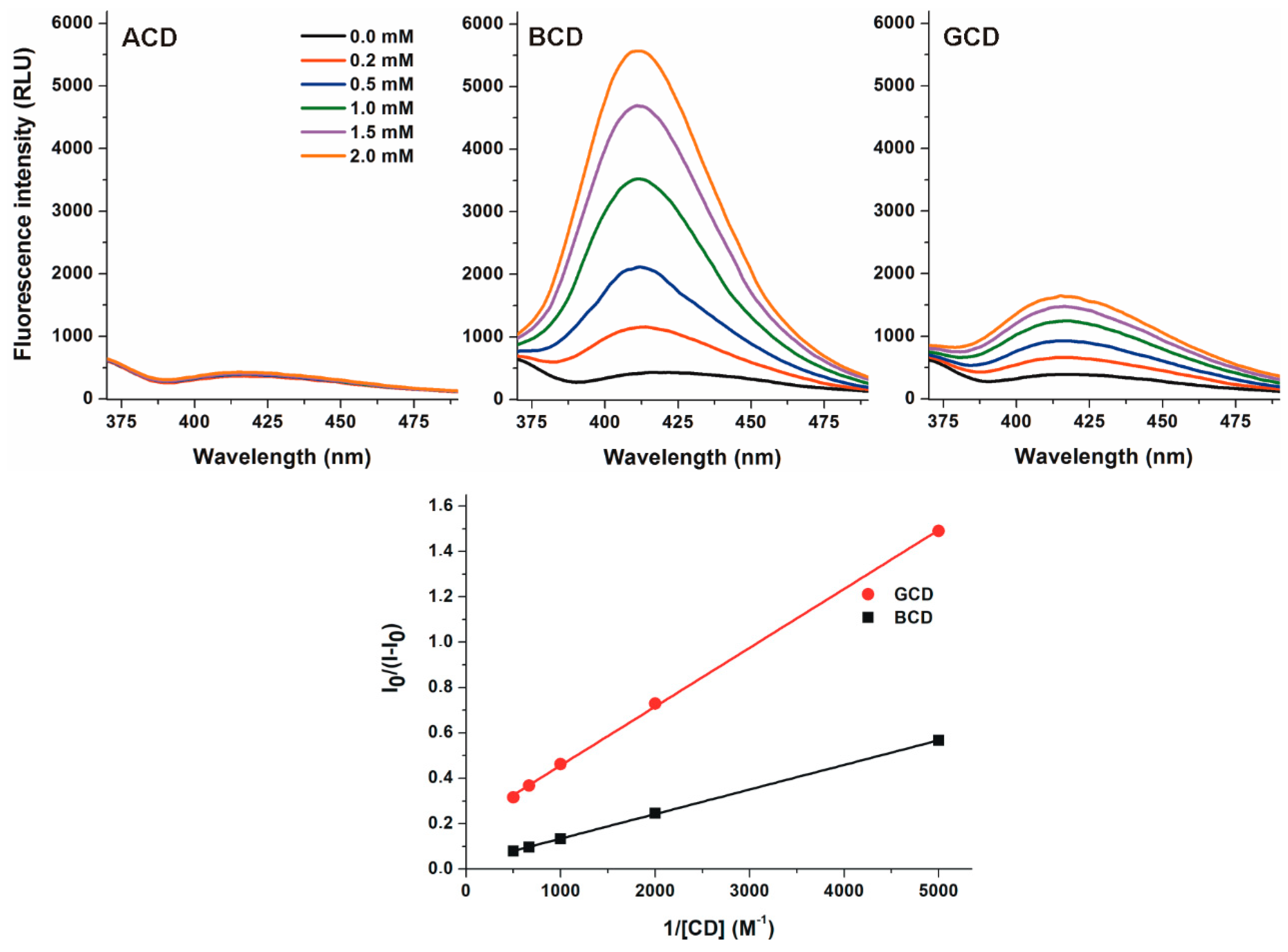
2.2. Effects of Native α-, β-, and γ-CDs on the Fluorescence Signal of DHC
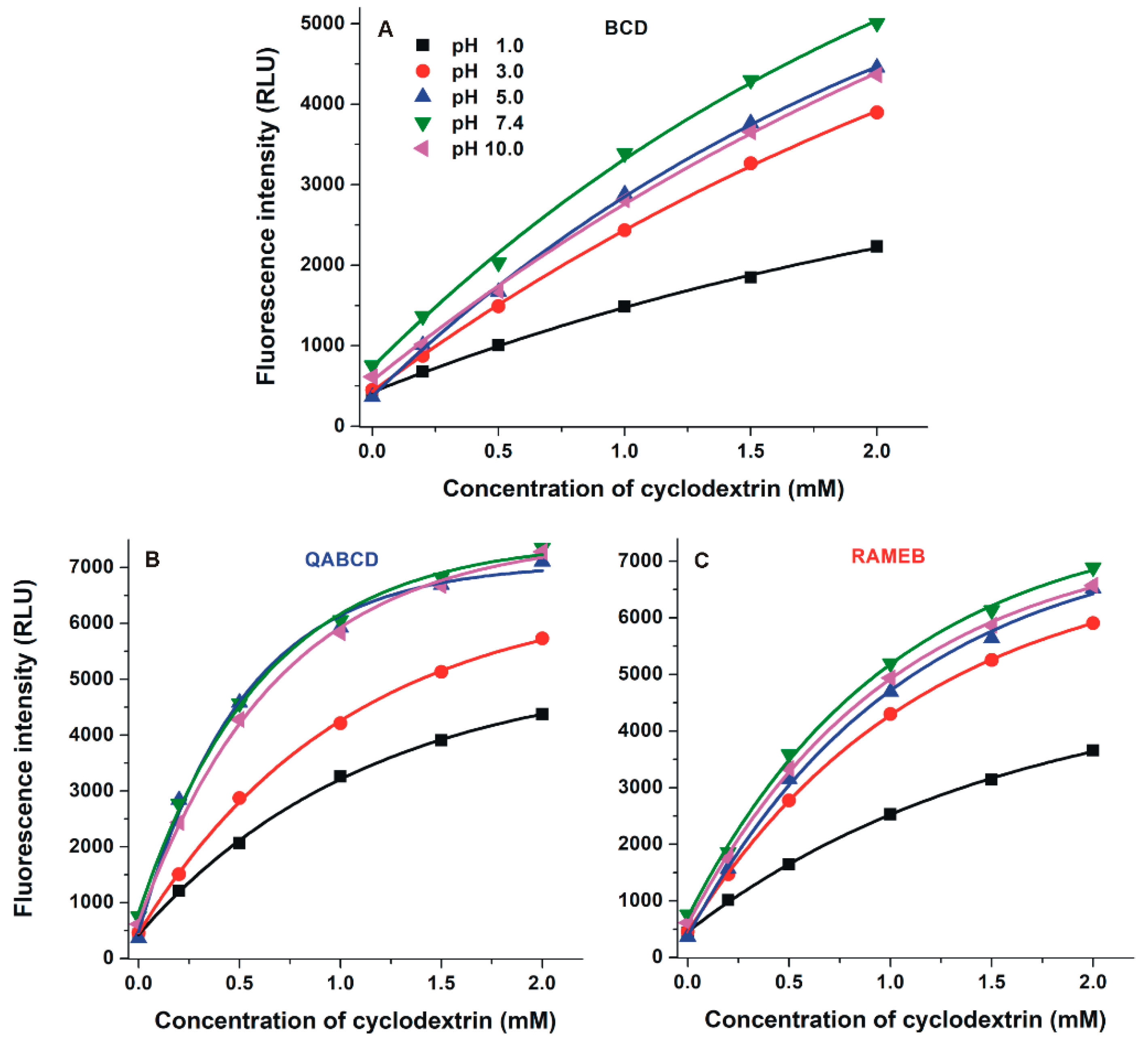
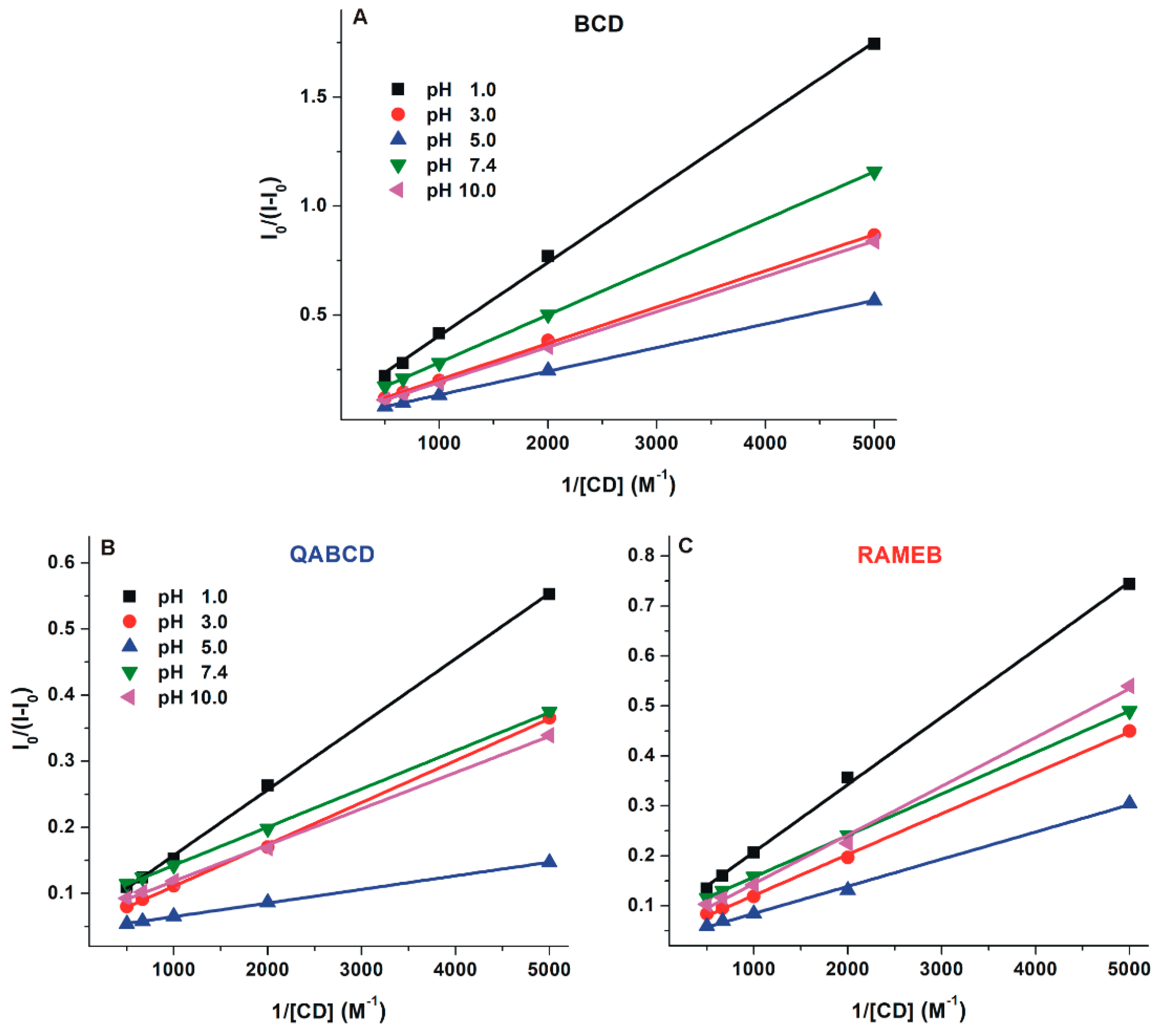
2.3. Interaction of DHC with β-Cyclodextrins
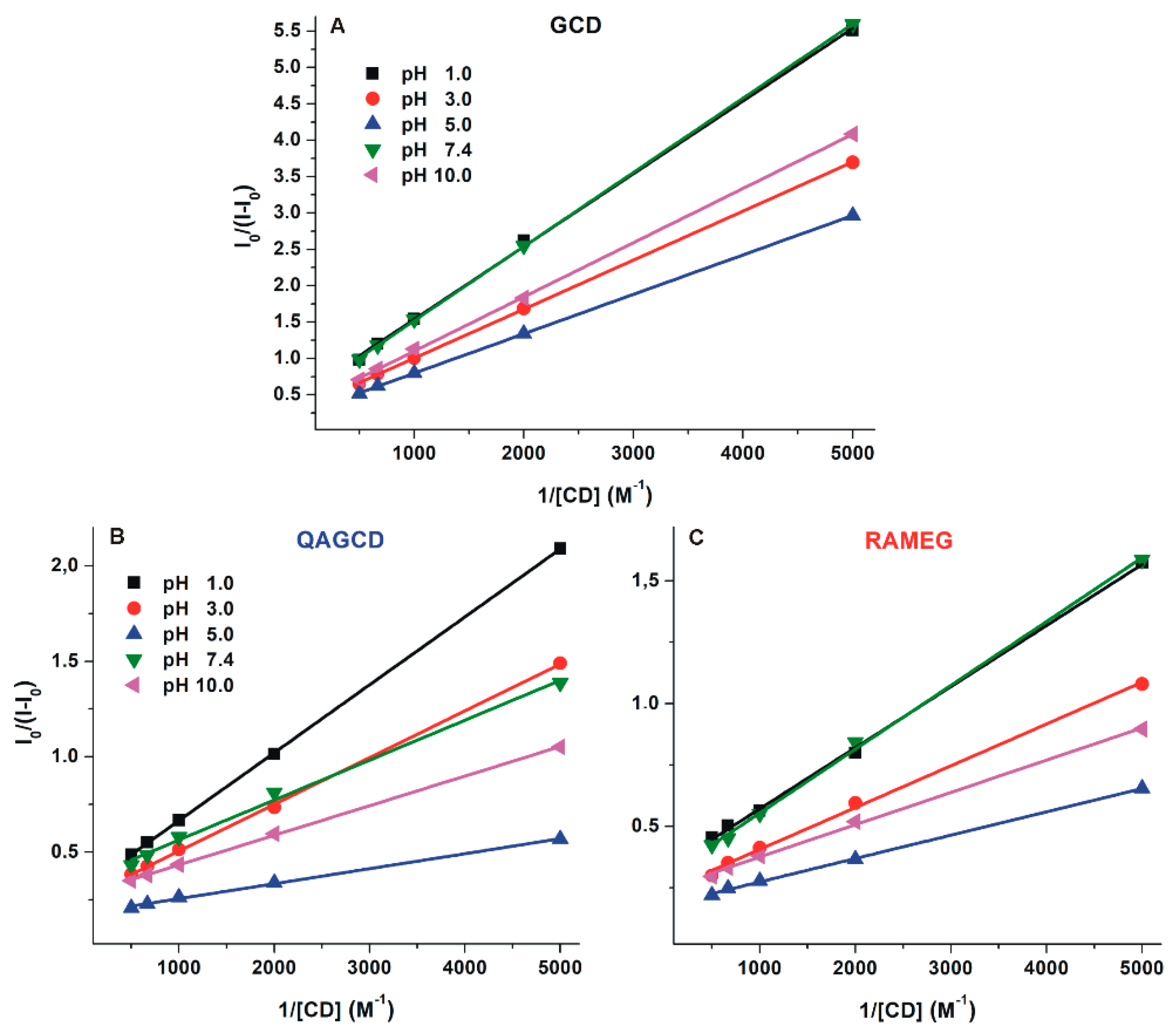
2.4. Interaction of DHC with γ-Cyclodextrins
2.5. Modeling Studies
3. Materials and Methods
3.1. Reagents
3.2. Fluorescence Spectroscopic Measurements
3.3. Modeling Studies
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Geisen, R.; Schmidt-Heydt, M.; Touhami, N.; Himmelsbach, A. New aspects of ochratoxin A and citrinin biosynthesis in Penicillium. Curr. Opin. Food Sci. 2018, 23, 23–31. [Google Scholar] [CrossRef]
- da Rocha, M.E.B.; Freire, F.C.O.; Maia, F.E.F.; Guedes, M.I.F.; Rondina, D. Mycotoxins and their effects on human and animal health. Food Control 2014, 36, 59–165. [Google Scholar] [CrossRef]
- de Oliveira Filho, J.W.G.; Islam, M.T.; Ali, E.S.; Uddin, S.J.; Santos, J.V.O.; de Alencar, M.V.O.B.; Júnior, A.L.G.; Paz, M.F.C.J.; de Brito, M.D.R.M.; E Sousa, J.M.C.; et al. A comprehensive review on biological properties of citrinin. Food Chem. Toxicol. 2017, 110, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Kitabatake, N.; Trivedi, A.B.; Doi, E. Thermal Decomposition and Detoxification of Citrinin under Various Moisture Conditions. J. Agric. Food Chem. 1991, 39, 2240–2244. [Google Scholar] [CrossRef]
- Dunn, B.B.; Stack, M.E.; Park, D.L.; Joshi, A.; Friedman, L.; King, R.L. Isolation and identification of dihydrocitrinone, a urinary metabolite of citrinin in rats. J. Toxicol. Environ. Health 1983, 12, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Degen, G.H.; Ali, N.; Gundert-Remy, U. Preliminary data on citrinin kinetics in humans and their use to estimate citrinin exposure based on biomarkers. Toxicol. Lett. 2018, 282, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Föllmann, W.; Behm, C.; Degen, G.H. Toxicity of the mycotoxin citrinin and its metabolite dihydrocitrinone and of mixtures of citrinin and ochratoxin A in vitro. Arch. Toxicol. 2014, 88, 1097–1107. [Google Scholar] [CrossRef]
- Faisal, Z.; Vörös, V.; Lemli, B.; Derdák, D.; Kunsági-Máté, S.; Bálint, M.; Hetényi, C.; Csepregi, R.; Kőszegi, T.; Bergmann, D.; Sueck, F.; Humpf, H.-U.; Hübner, F.; Poór, M. Interaction of the mycotoxin metabolite dihydrocitrinone with serum albumin. Mycotoxin Res. 2018. [Google Scholar] [CrossRef]
- Gerding, J.; Ali, N.; Schwartzbord, J.; Cramer, B.; Brown, D.L.; Degen, G.H.; Humpf, H.-U. A comparative study of the human urinary mycotoxin excretion patterns in Bangladesh, Germany and Haiti using a rapid and sensitive LC-MS/MS approach. Mycotoxin Res. 2015, 31, 127–136. [Google Scholar] [CrossRef]
- Ali, N.; Blaszkewicz, M.; Degen, G.H. Occurrence of the mycotoxin citrinin and its metabolite dihydrocitrinone in urines of German adults. Arch. Toxicol. 2015, 89, 573–578. [Google Scholar] [CrossRef]
- Ali, N.; Hossain, K.; Degen, G.H. Blood plasma biomarkers of citrinin and ochratoxin A exposure in young adults in Bangladesh. Mycotoxin Res. 2018, 34, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Szente, L.; Szejtli, J. Highly soluble cyclodextrin derivatives: Chemistry, properties, and trends in development. Adv. Drug Deliv. Rev. 1999, 36, 17–28. [Google Scholar] [CrossRef]
- Szente, L.; Szemán, J. Cyclodextrins in Analytical Chemistry: Host−Guest Type Molecular Recognition. Anal. Chem. 2013, 85, 8024–8030. [Google Scholar] [CrossRef] [PubMed]
- Crini, G. Review: A history of cyclodextrins. Chem. Rev. 2014, 114, 10940–10975. [Google Scholar] [CrossRef] [PubMed]
- Aghamohammadi, M.; Alizadeh, N. Fluorescence enhancement of the aflatoxin B1 by forming inclusion complexes with some cyclodextrins and molecular modeling study. J. Lumin. 2007, 127, 575–582. [Google Scholar] [CrossRef]
- Amadasi, A.; Dall’asta, C.; Ingletto, G.; Pela, R.; Marchelli, R.; Cozzini, P. Explaining cyclodextrin–mycotoxin interactions using a ’natural’ force field. Bioorg. Med. Chem. 2007, 15, 4585–4594. [Google Scholar] [CrossRef]
- Verrone, R.; Catucci, L.; Cosma, P.; Fini, P.; Agostiano, A.; Lippolis, V.; Pascale, M. Effect of b-cyclodextrin on spectroscopic properties of ochratoxin A in aqueous solution. J. Incl. Phenom. Macrocycl. Chem. 2007, 57, 475–479. [Google Scholar] [CrossRef]
- Dall’Asta, C.; Faccini, A.; Galaverna, G.; Corradini, R.; Dossena, A.; Marchelli, R. Complexation of zearalenone and zearalenols with native and modified b-cyclodextrins. J. Incl. Phenom. Macrocycl. Chem. 2009, 64, 331–340. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, J.; Dong, L.; Lu, L.; Chen, F.; Hu, D.; Wang, X. A study of fluorescence properties of citrinin in b-cyclodextrin aqueous solution and different solvents. J. Lumin. 2012, 132, 1437–1445. [Google Scholar] [CrossRef]
- Poór, M.; Kunsági-Máté, S.; Sali, N.; Kőszegi, T.; Szente, L.; Peles-Lemli, B. Interactions of zearalenone with native and chemically modified cyclodextrins and their potential utilization. J. Photochem. Photobiol. B Biol. 2015, 151, 63–68. [Google Scholar] [CrossRef]
- Poór, M.; Kunsági-Máté, S.; Szente, L.; Matisz, G.; Secenji, G.; Czibulya, Z.; Kőszegi, T. Interaction of ochratoxin A with quaternary ammonium beta-cyclodextrin. Food Chem. 2015, 172, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Poór, M.; Zand, A.; Szente, L.; Lemli, B.; Kunsági-Máté, S. Interaction of α- and β-zearalenols with β-cyclodextrins. Molecules 2017, 22, 1910. [Google Scholar] [CrossRef]
- Maragos, C.M.; Appell, M.; Lippolis, V.; Visconti, A.; Catucci, L.; Pascale, M. Use of cyclodextrins as modifiers of fluorescence in the detection of mycotoxins. Food Addit. Contam. 2008, 25, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Dall’Asta, C.; Faccini, A.; Galaverna, G.; Corradini, R.; Dossena, A.; Marchelli, R. Complexation of the mycotoxin zearalenone with β-cyclodextrin: Study of the interaction and first promising applications. Mycotoxin Res. 2008, 24, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Galaverna, G.; Dall’Asta, C.; Corradini, R.; Dossena, A.; Marchelli, R. Cyclodextrins as selectors for mycotoxin recognition. World Mycotoxin J. 2008, 1, 397–406. [Google Scholar] [CrossRef]
- Appell, M.; Jackson, M.A. Synthesis and evaluation of cyclodextrin-based polymers for patulin extraction from aqueous solutions. J. Incl. Phenom. Macrocycl. Chem. 2010, 68, 117–122. [Google Scholar] [CrossRef]
- Appell, M.; Jackson, M.A. Sorption of ochratoxin A from aqueous solutions using β-cyclodextrin–polyurethane polymer. Toxins 2012, 4, 98–109. [Google Scholar] [CrossRef]
- Poór, M.; Faisal, Z.; Zand, A.; Bencsik, T.; Lemli, B.; Kunsági-Máté, S.; Szente, L. Removal of Zearalenone and Zearalenols from Aqueous Solutions Using Insoluble Beta-Cyclodextrin Bead Polymer. Toxins 2018, 10, 216. [Google Scholar] [CrossRef]
- Poór, M.; Matisz, G.; Kunsági-Máté, S.; Derdák, D.; Szente, L.; Lemli, B. Fluorescence spectroscopic investigation of the interaction of citrinin with native and chemically modified cyclodextrins. J. Lumin. 2016, 172, 23–28. [Google Scholar] [CrossRef]
- Tölgyesi, Á.; Stroka, J.; Tamosiunas, V.; Zwickel, T. Simultaneous analysis of Alternaria toxins and citrinin in tomato: An optimised method using liquid chromatography-tandem mass spectrometry. Food Addit. Contam. Part A Chem. 2015, 32, 1512–1522. [Google Scholar] [CrossRef]
- Dobretsov, G.E.; Syrejschikova, T.I.; Smolina, N.V. On mechanisms of fluorescence quenching by water. Biophysics 2014, 59, 183–188. [Google Scholar] [CrossRef]
- Bergmann, D.; Hübner, F.; Wibbeling, B.; Daniliuc, C.; Cramer, B.; Humpf, H.-U. Large-scale total synthesis of 13C3-labeled citrinin and its metabolite dihydrocitrinone. Mycotoxin Res. 2018, 34, 141–150. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | DHC–BCD | DHC–QABCD | DHC–RAMEB | |||
|---|---|---|---|---|---|---|
| logK ± SEM | I/I0 ± SEM | logK ± SEM | I/I0 ± SEM | logK ± SEM | I/I0 ± SEM | |
| 1.0 | 2.29 ± 0.04 | 5.59 ± 0.12 | 2.76 ± 0.01 | 10.17 ± 0.11 | 2.76 ± 0.02 | 6.98 ± 0.11 |
| 3.0 | 2.34 ± 0.04 | 9.23 ± 0.35 | 2.81 ± 0.02 | 12.89 ± 0.18 | 2.79 ± 0.04 | 12.89 ± 0.19 |
| 5.0 | 2.47 ± 0.05 | 13.14 ± 0.63 | 3.23 ± 0.05 | 19.53 ± 0.06 | 2.78 ± 0.02 | 17.59 ± 0.12 |
| 7.4 | 2.42 ± 0.06 | 6.75 ± 0.06 | 3.21 ± 0.02 | 9.92 ± 0.07 | 2.90 ± 0.03 | 9.31 ± 0.14 |
| 10.0 | 2.22 ± 0.02 | 7.40 ± 0.36 | 3.03 ± 0.02 | 14.22 ± 0.34 | 2.85 ± 0.04 | 12.74 ± 0.34 |
| pH | DHC–GCD | DHC–QAGCD | DHC–RAMEG | |||
|---|---|---|---|---|---|---|
| logK ± SEM | I/I0 ± SEM | logK ± SEM | I/I0 ± SEM | logK ± SEM | I/I0 ± SEM | |
| 1.0 | 2.69 ± 0.03 | 2.09 ± 0.03 | 2.98 ± 0.03 | 2.96 ± 0.06 | 3.12 ± 0.01 | 3.12 ± 0.08 |
| 3.0 | 2.78 ± 0.05 | 2.41 ± 0.10 | 3.13 ± 0.05 | 3.70 ± 0.12 | 3.20 ± 0.06 | 4.21 ± 0.09 |
| 5.0 | 2.73 ± 0.04 | 3.05 ± 0.07 | 3.38 ± 0.04 | 6.01 ± 0.09 | 3.22 ± 0.03 | 5.64 ± 0.08 |
| 7.4 | 2.75 ± 0.03 | 2.00 ± 0.01 | 3.18 ± 0.02 | 3.48 ± 0.16 | 3.05 ± 0.02 | 3.46 ± 0.05 |
| 10.0 | 2.74 ± 0.06 | 2.32 ± 0.07 | 3.24 ± 0.02 | 3.83 ± 0.02 | 3.18 ± 0.05 | 4.42 ± 0.03 |
| Ionic State of DHC | DHC–BCD | DHC–GCD | ||||||
| logK | ΔG | ΔH | ΔS | logK | ΔG | ΔH | ΔS | |
| 0 | 2.3 | −18.65 | −14.60 | 13.6 | 2.7 | −21.90 | −22.85 | −3.19 |
| −1 | 2.5 | −20.28 | −16.02 | 14.3 | 2.7 | −21.94 | −26.45 | −15.13 |
| −2 | 2.2 | −17.84 | −13.19 | 15.6 | 2.7 | −21.97 | −28.19 | −20.86 |
| DHC–RAMEB | DHC–QABCD | |||||||
| logK | ΔG | ΔH | ΔS | logK | ΔG | ΔH | ΔS | |
| 0 | 2.7 | −21.90 | −17.85 | 13.6 | 2.7 | −21.90 | −17.85 | 13.6 |
| −1 | 2.8 | −22.71 | −18.45 | 14.3 | 3.3 | −26.76 | −25.07 | 5.7 |
| −2 | 2.8 | −22.71 | −18.06 | 15.6 | 3.0 | −24.33 | −22.49 | 6.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faisal, Z.; Kunsági-Máté, S.; Lemli, B.; Szente, L.; Bergmann, D.; Humpf, H.-U.; Poór, M. Interaction of Dihydrocitrinone with Native and Chemically Modified Cyclodextrins. Molecules 2019, 24, 1328. https://doi.org/10.3390/molecules24071328
Faisal Z, Kunsági-Máté S, Lemli B, Szente L, Bergmann D, Humpf H-U, Poór M. Interaction of Dihydrocitrinone with Native and Chemically Modified Cyclodextrins. Molecules. 2019; 24(7):1328. https://doi.org/10.3390/molecules24071328
Chicago/Turabian StyleFaisal, Zelma, Sándor Kunsági-Máté, Beáta Lemli, Lajos Szente, Dominik Bergmann, Hans-Ulrich Humpf, and Miklós Poór. 2019. "Interaction of Dihydrocitrinone with Native and Chemically Modified Cyclodextrins" Molecules 24, no. 7: 1328. https://doi.org/10.3390/molecules24071328
APA StyleFaisal, Z., Kunsági-Máté, S., Lemli, B., Szente, L., Bergmann, D., Humpf, H.-U., & Poór, M. (2019). Interaction of Dihydrocitrinone with Native and Chemically Modified Cyclodextrins. Molecules, 24(7), 1328. https://doi.org/10.3390/molecules24071328