Fatty Acid, Lipid Classes and Phospholipid Molecular Species Composition of the Marine Clam Meretrix lyrata (Sowerby 1851) from Cua Lo Beach, Nghe An Province, Vietnam

 , , , ,
, , , ,
Abstract
1. Introduction
2. Results and Discussion
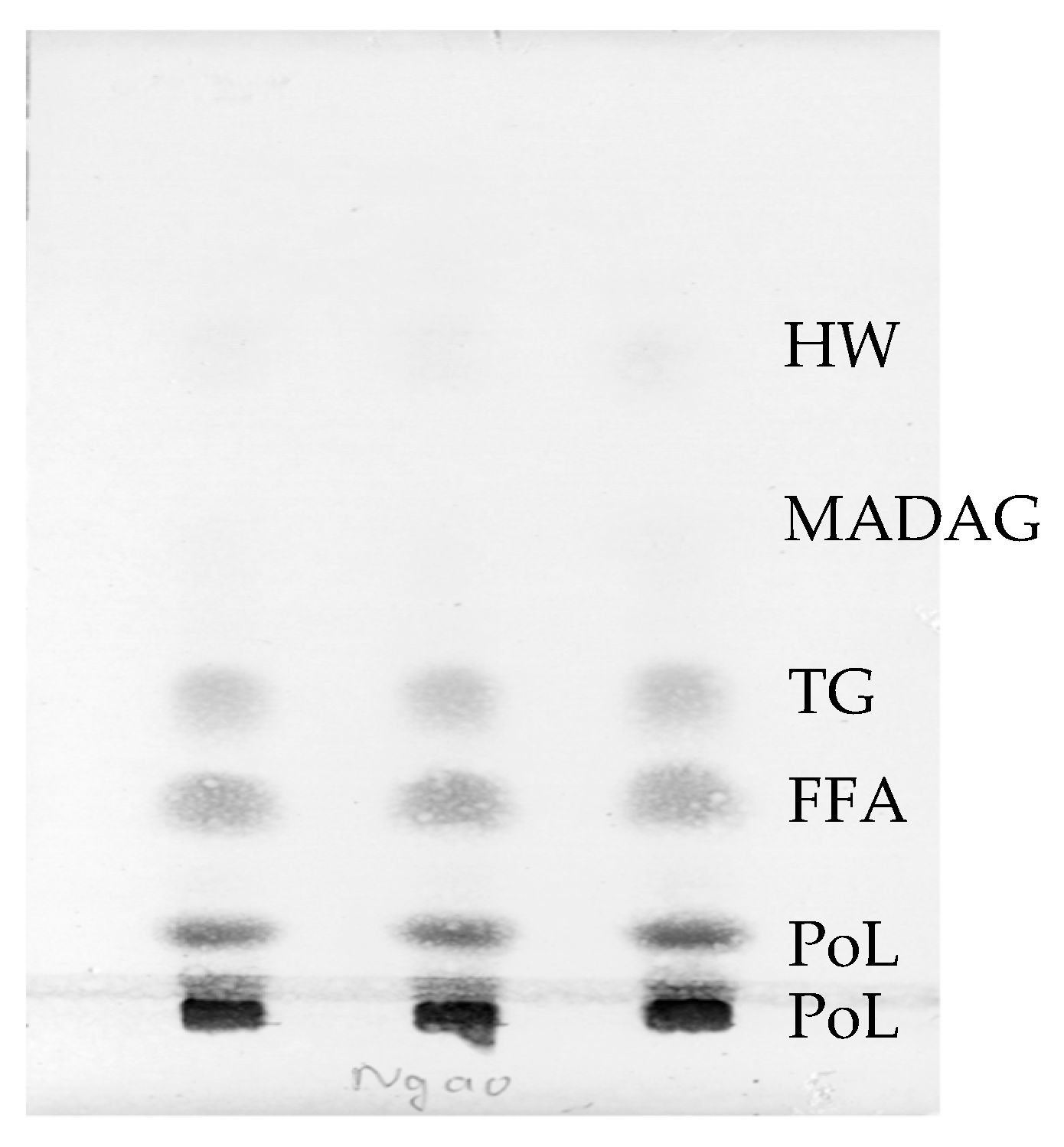
2.1. Total Lipid
2.2. Fatty Acids in Total Lipids
2.3. Fatty Acids in Fractions of Phospholipid
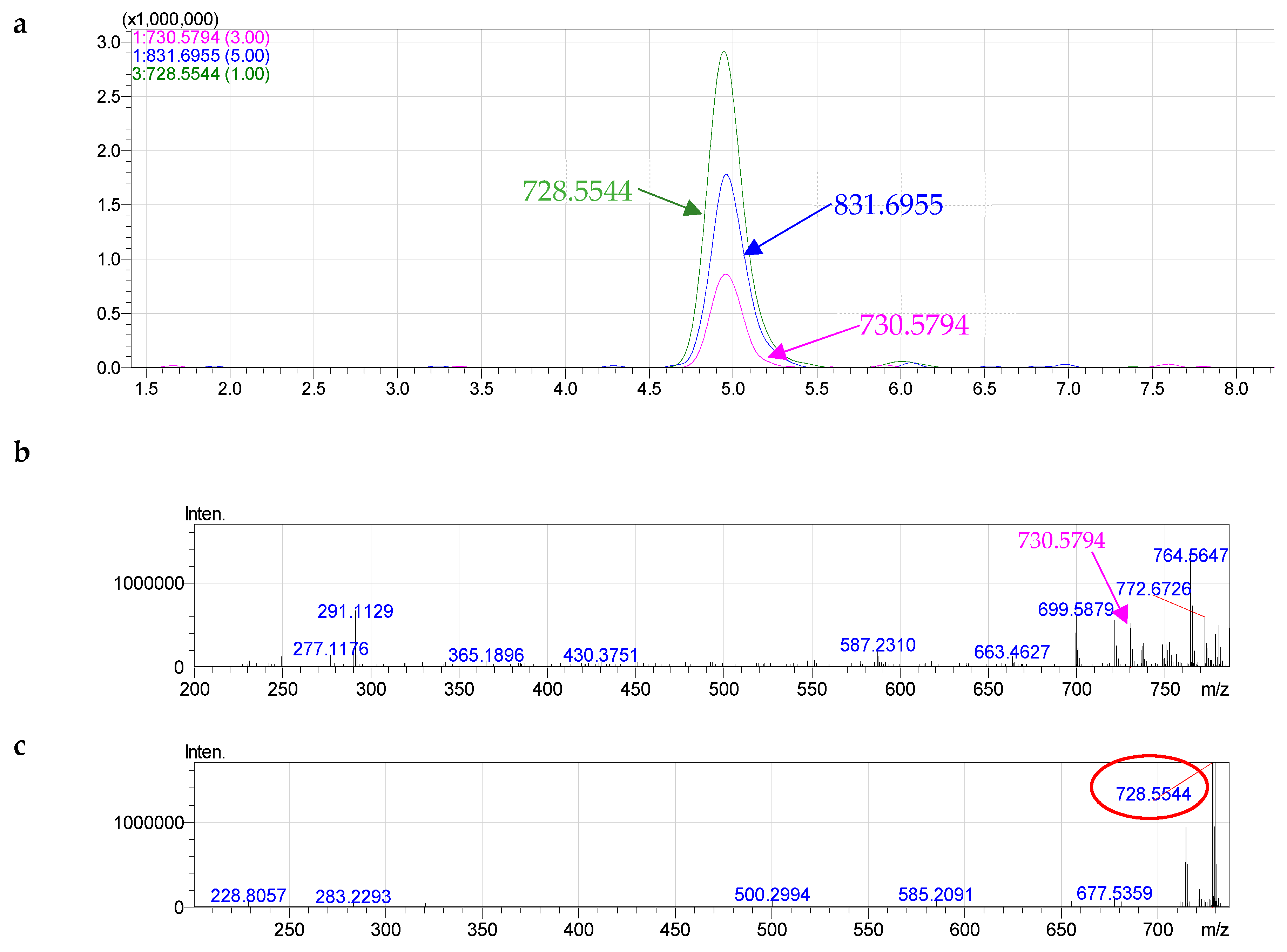
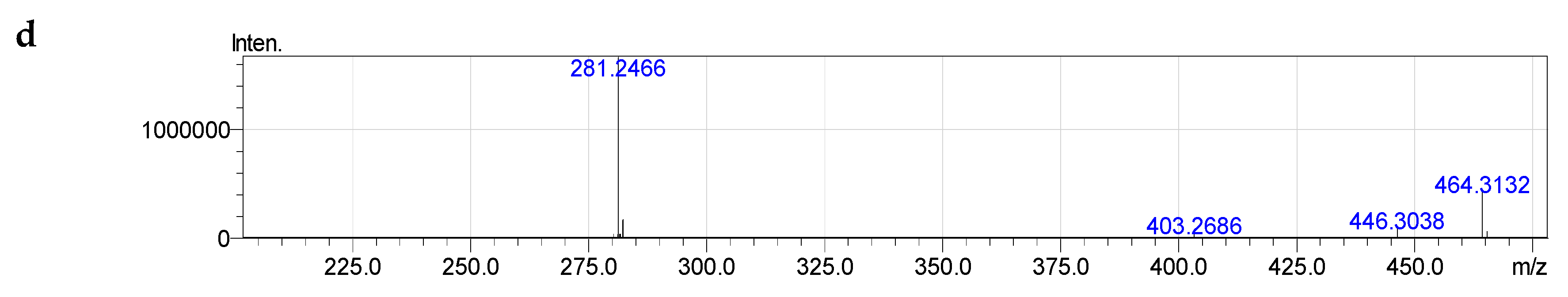
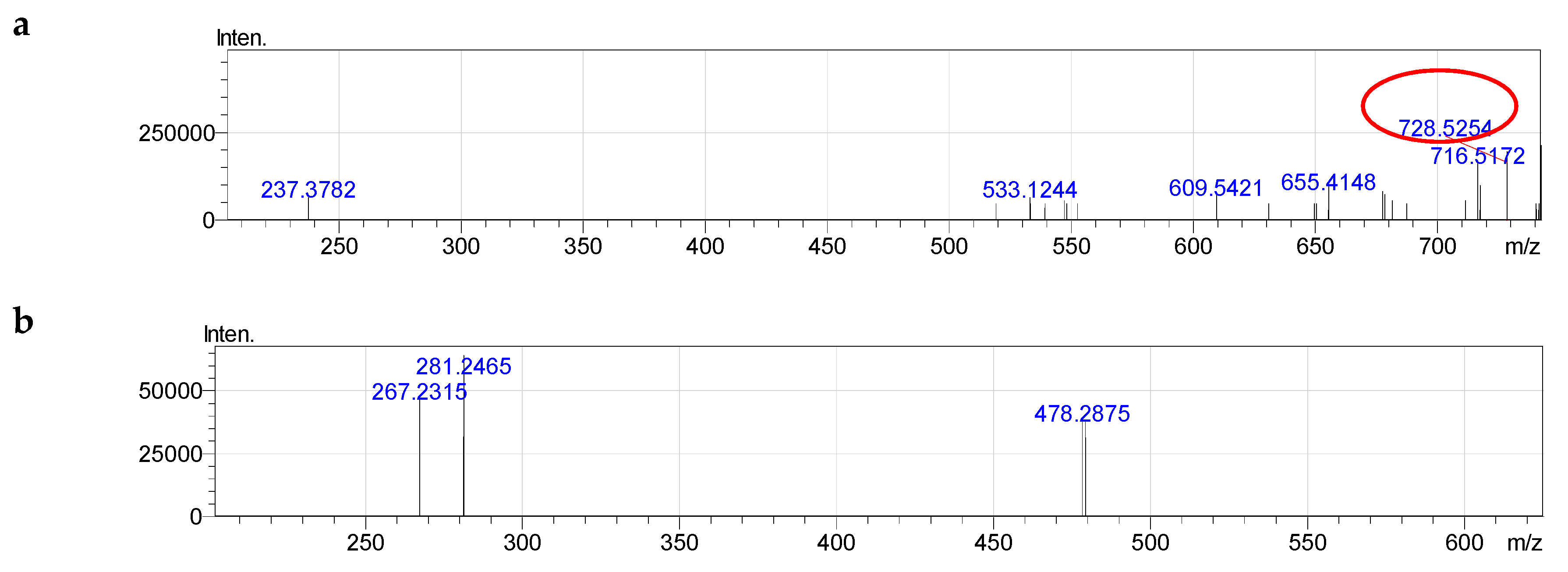
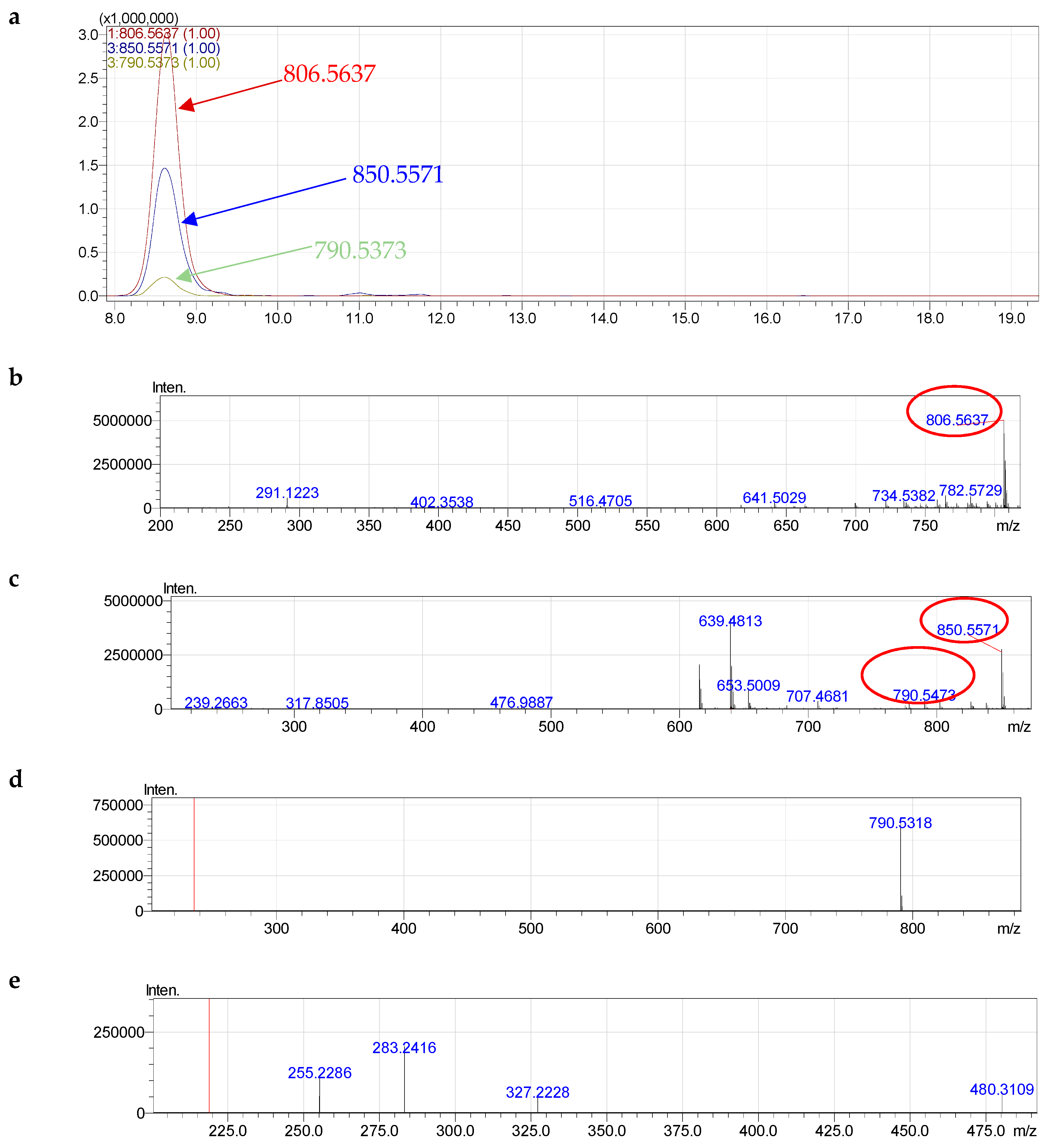
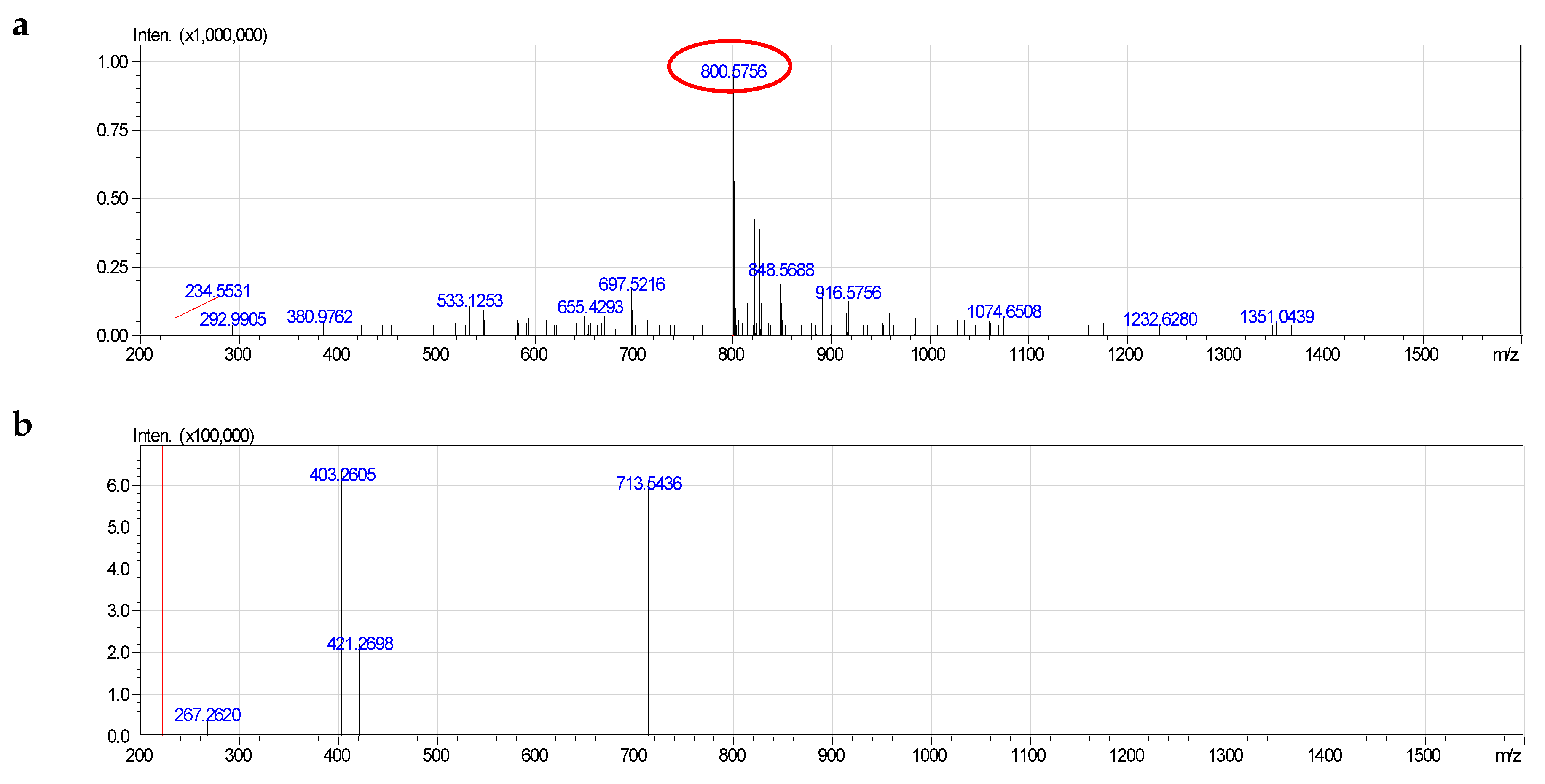
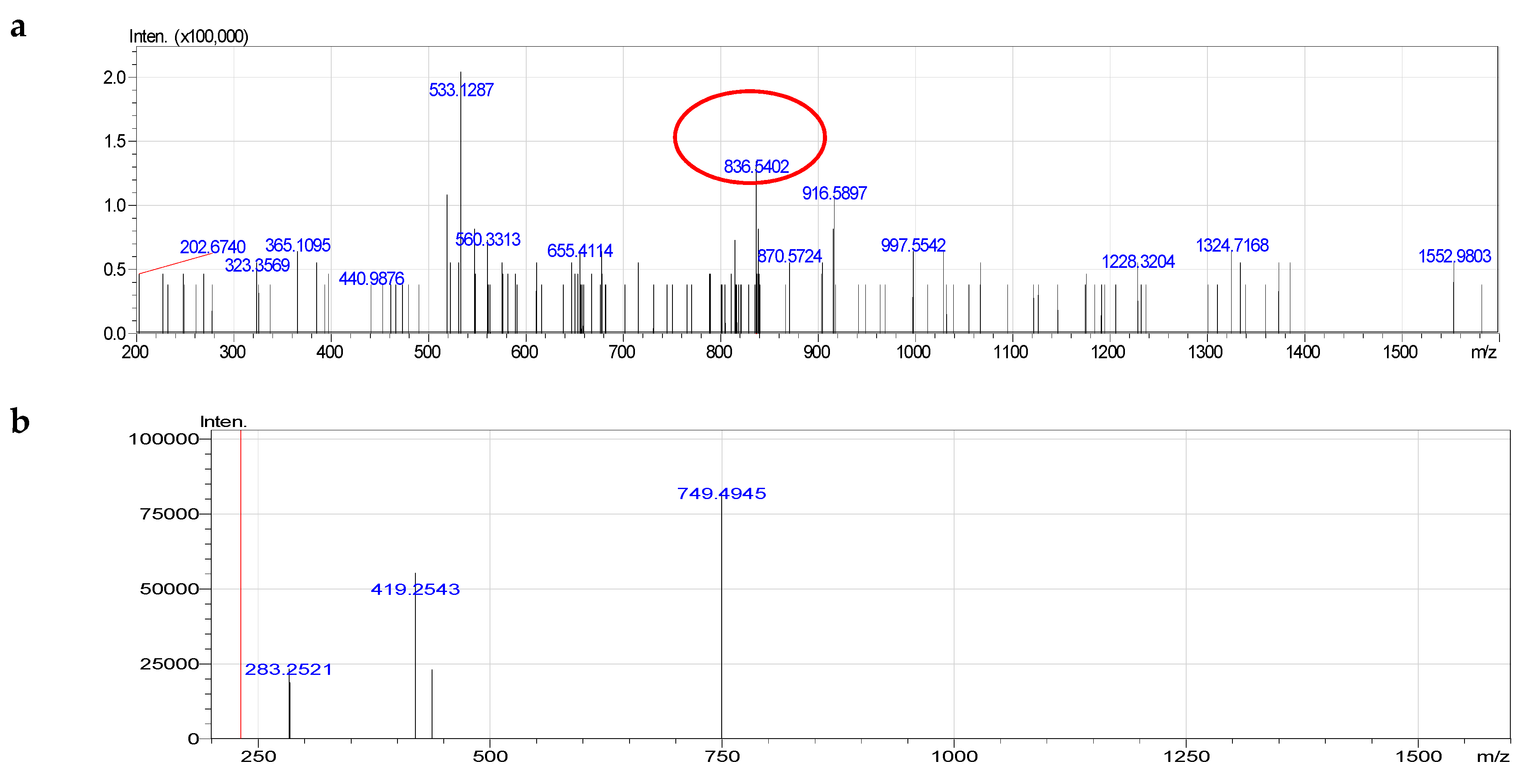
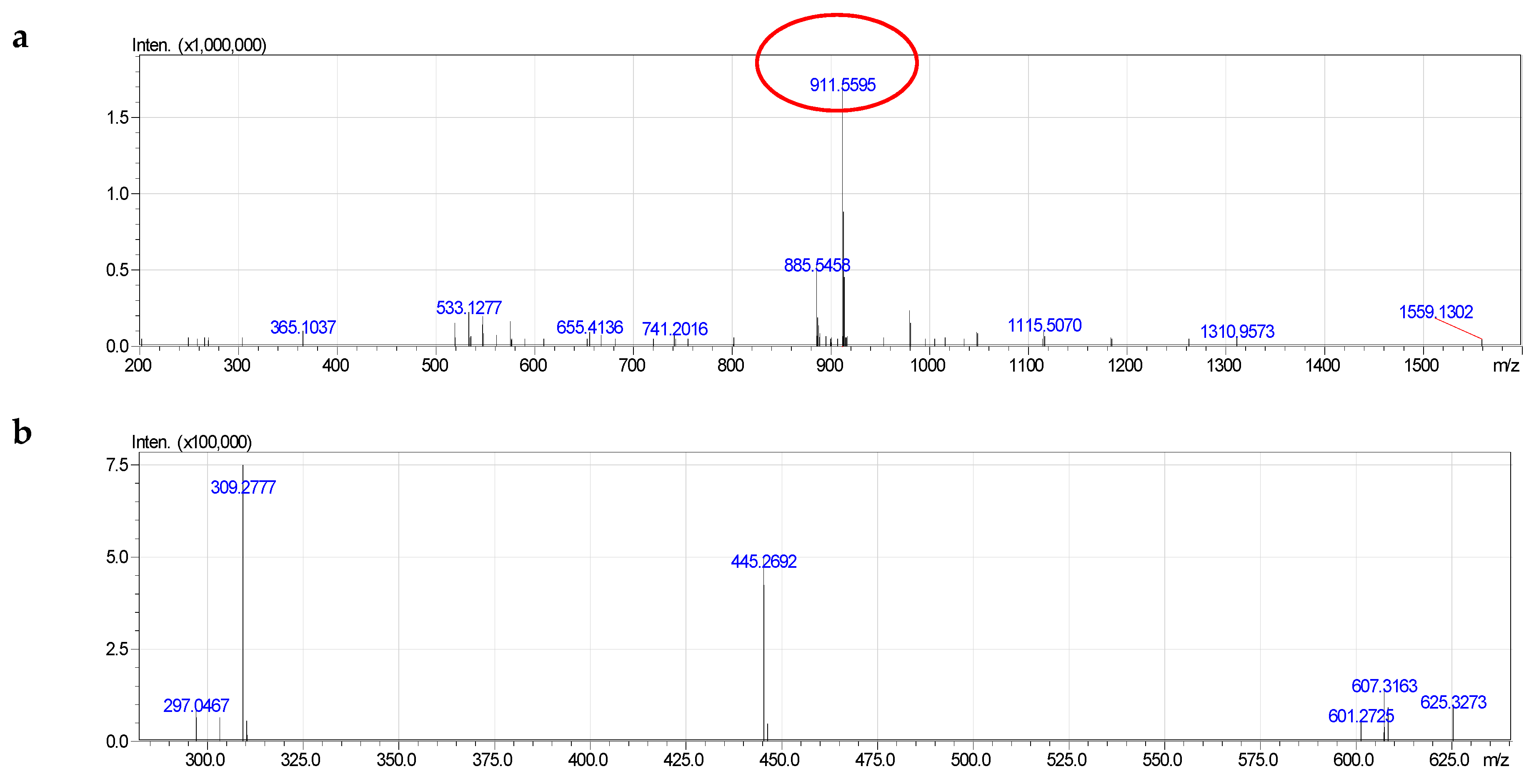
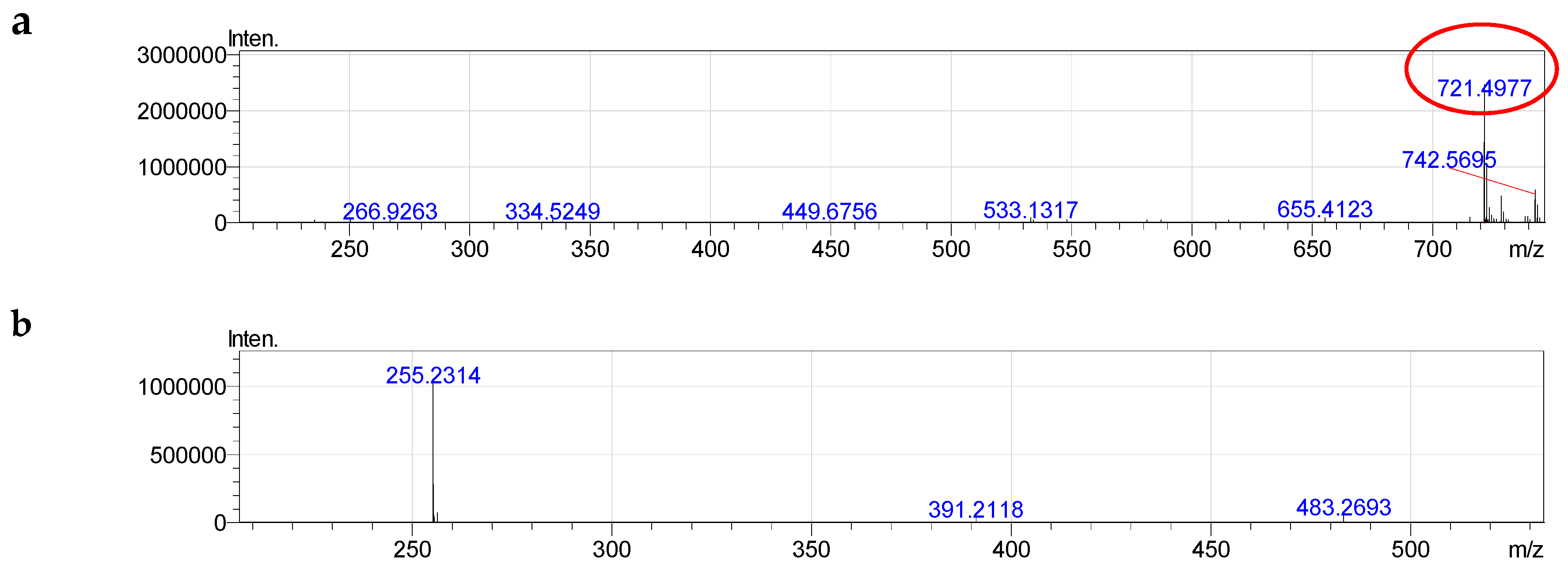
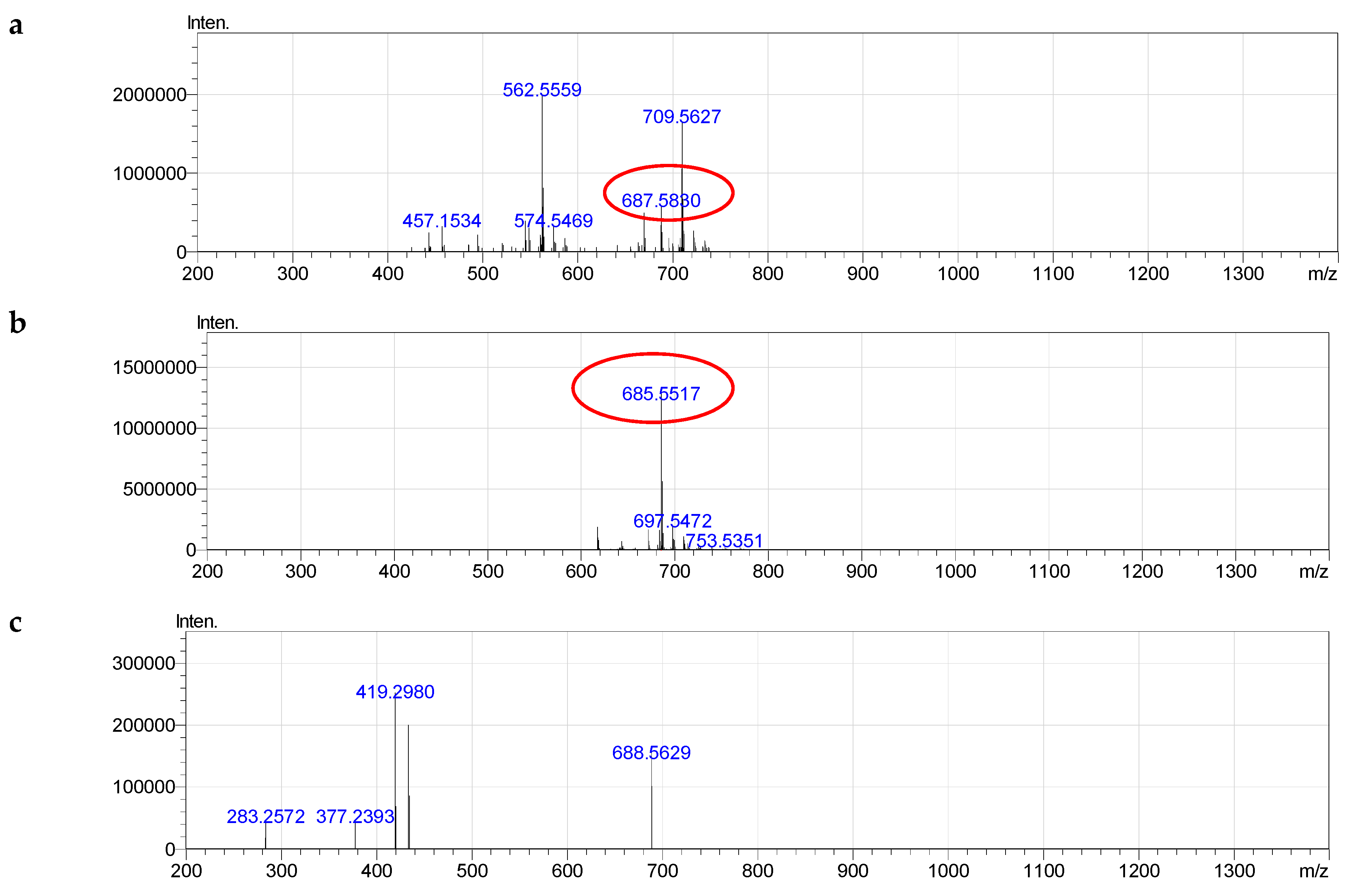
2.4. Molecular Species of Phospholipid
3. Materials and Methods
3.1. Material
3.2. Total Lipid Extraction
3.3. Analyses of Lipid Classes
3.4. Separation of Phospholipid Classes
3.5. Analyses of Fatty Acids
3.6. Analysis of Molecular Species of Phospholipids
3.7. Instrumental Equipment
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | Arachidonic acid |
| CAEP | Ceramide aminoethylphosphonate |
| ChoGpl | Choline glycerophospholipids |
| DHA | Docosahexaenoic acid |
| DMA | Dimethylacetal |
| DPA | Docosapentaenoic acid |
| EPA | Eicosapentaenoic acid |
| FFA | Free fatty acids |
| GC | Gas chromatography |
| GCMS | Gas chromatography—mass spectrometry |
| HPA | Heneicosapentaenoic acid |
| HPLC-HRMS | High performance liquid chromatography–high resolution mass spectrometry |
| HRMS | High resolution mass spectrometry |
| HUFA | Highly unsaturated fatty acid |
| HW | Hydrocarbon and wax |
| LC-PUFA | Long-chain polyunsaturated fatty acid |
| MADAG | Monoalkyl diacylglycerol |
| MS | Mass spectrometry |
| MS/MS | Mass spectrometry / Mass spectrometry (or tandem mass spectrometry) |
| MUFA | Monounsaturated fatty acid |
| NMI-PUFA | Non-methylene interrupted polyunsaturated fatty acid |
| PakCho | 1-O-alkyl-2-acyl-sn-glycero-3-phosphocholine |
| PC | Phosphatidylcholine |
| PE | Phosphatidylethalnolamine |
| PG | Phosphatidylglycerol |
| PI | Phosphatidylinositol |
| PL | Phospholipid |
| PlsEtn | Ethanolamine plasmalogen |
| PoL | Polar lipid |
| PS | Phosphatidylserine |
| PtdIns | Phosphatidylinositol |
| PUFA | Poly-unsaturated fatty acid |
| SAFA | Saturated fatty acid |
| ST | Sterol |
| TAG | Triacylglycerol |
| TL | Total lipid |
| TLC | Thin-layer chromatography |
| USAFA | Unsaturated fatty acid |
References
- Loureiro, D.R.P.; Soares, J.X.; Costa, J.C.; Magalhães, A.F.; Azevedo, C.M.G.; Pinto, M.M.M.; Afonso, C.M.M. Structures, Activities and Drug-Likeness of Anti-Infective Xanthone Derivatives Isolated from the Marine Environment: A Review. Molecules 2019, 24, 243. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.C.; Wu, W.T.; Yang, F.L.; Chiu, Y.H.; Peng, T.C.; Hsu, B.G.; Liao, K.W.; Lee, R.P. Effects of Freshwater Clam Extract Supplementation on Time to Exhaustion, Muscle Damage, Pro/Anti-Inflammatory Cytokines, and Liver Injury in Rats after Exhaustive Exercise. Molecules 2013, 18, 3825–3838. [Google Scholar] [CrossRef] [PubMed]
- Greetham, D.; Zaky, A.; Makanjuola, O.; Du, C. A brief review on bioethanol production using marine biomass, marine microorganism and seawater. Curr. Opin. Green Sustainable Chem. 2018, 14, 53–59. [Google Scholar] [CrossRef]
- Patil, M.P.; Kim, G.D. Marine microorganisms for synthesis of metallic nanoparticles and their biomedical applications. Colloids Surf. B 2018, 172, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Amorello, D.; Orecchio, S.; Pace, A.; Barreca, S. Discrimination of almonds (Prunus dulcis) geographical origin by minerals and fatty acids profiling. Nat. Prod. Res. 2016, 30, 2107–2110. [Google Scholar] [CrossRef] [PubMed]
- FAO Yearbook of Fishery and Aquaculture Statistics 2018. Available online: http://www.fao.org/fishery/statistics/en (accessed on 20 February 2019).
- Fernández, M.J.; Labarta, U.; Albentosa, M.; Pérez-Camacho, A. Lipid composition of Ruditapes philipponarum spat: Effect of ration and diet quality. Comp. Biochem. Physiol. 2006, 144, 229–237. [Google Scholar]
- Orban, E.; Di, L.G.; Nevigato, T.; Casini, I.; Caproni, R.; Santaroni, G.; Giulini, G. Nutritional and commercial quality of the striped venus clam, Chamelea gallina, from the Adriatic sea. Food Chem. 2007, 101, 1063–1070. [Google Scholar] [CrossRef]
- Karnjanapratum, S.; Benjakul, S.; Kishimura, H.; Tsai, Y. Chemical compositions and nutritional value of Asian hard clam (Meretrix lusoria) from the coast of Andaman Sea. Food Chem. 2013, 141, 4138–4145. [Google Scholar] [CrossRef] [PubMed]
- Boselli, E.; Pacetti, D.; Lucci, P.; Frega, N. Characterization of phospholipid molecular species in the edible parts of bony fish and shellfish. J. Agric. Food Chem. 2012, 60, 3234–3245. [Google Scholar] [CrossRef] [PubMed]
- Janssen, C.I.; Kiliaan, A.J. Long chain polyunsaturated fatty acids (LC-PUFA) from genesis to senecscence: The influence of LC-PUFA on neural development, aging and neurodegeneration. Prog. Lipid Res. 2014, 5, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lorente-Cebrian, S.; Costa, A.; Navas-Carretero, S.; Zabala, M.; Martinez, J.; Moreno-Aliaga, M. Role of omega-3 fatty acids in obesity, metabolic syndrome and cardiovascular diseases: A review of the evidence. J. Physiol. Biochem. 2013, 69, 633–651. [Google Scholar] [CrossRef] [PubMed]
- Cottin, S.; Sanders, T.A.; Hall, W.L. The differential effects of EPA and DHA on cardiovascular risk factors. Proc. Nutr. Soc. 2011, 70, 215–231. [Google Scholar] [CrossRef] [PubMed]
- Edelstein, S. Food Science: An Ecological Approach; Jones & Bartlett Publishers: Burlington, MA, USA, 2014; pp. 70–71. [Google Scholar]
- Miller, M.; Stone, N.J.; Ballantyne, C.; Bittner, V.; Criqui, M.H.; Ginsberg, H.N. Triglycerides and cardiovascular disease: A scientific statement from the American Heart Association. Circulation 2011, 123, 2292–2333. [Google Scholar] [CrossRef] [PubMed]
- Weintraub, H.S. Overview of prescription omega-3 fatty acid products for hypertriglyceridemia. Postgrad Med. 2014, 126, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Saito, H. Identification of novel n-4 series polyunsaturated fatty acids in a deep-sea clam, Calyptogena phaseoliformis. J. Chromatogr. A 2007, 1163, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Saito, H.; Murata, M.; Hashimoto, J. Lipid characteristics of a seep clam, Mesolinga soliditesta: Comparison with those of two coastal clams, Meretrix lamarckii and Ruditapes philippinarum. Deep Sea Res. Part I 2014, 94, 150–158. [Google Scholar] [CrossRef]
- FAO Yearbook. Fishery and Aquaculture Statistics 2016. Available online: http://www.fao.org/fishery/static/Yearbook/YB2016_USBcard/navigation/index_intro_e.htm (accessed on 20 February 2019).
- Hamli, H.; Idris, M.H.; Rajaee, A.H.; Kamal, A.H.M. Reproductive Cycle of Hard Clam, Meretrix lyrata Sowerby, 1851 (Bivalvia: Veneridae) from Sarawak, Malaysia. Trop Life Sci. Res. 2015, 26, 59–72. [Google Scholar] [PubMed]
- Thao, N.T.; Nha le, Q. Effects of glucose and probiotic supplementation in nursing juvenile clam, Meretrix lyrata. Commun. Agric. Appl. Biol. Sci. 2013, 78, 304–307. [Google Scholar] [PubMed]
- Klingensmith, J.S.; Stillway, L.W. Lipid composition of selected tissues of the hardsell clam, Mercenaria mercenaria. Comp. Biochem. Physiol. 1981, 71B, 111–112. [Google Scholar]
- Caers, M.; Coutteau, P.; Sorgeloos, P. Impact of starvation and of feeding algal and artificial diets on the lipid content and composition of juvenile oysters (Crassostrea gigas) and clams (Tapes philippinarum). J. Mar. Biol. 2000, 136, 891–899. [Google Scholar] [CrossRef]
- Aouini, F.; Ghribi, F.; Boussoufa, D.; Bejaoui, S.; Navarro, J.; Cafsi, M.E. Changes in lipid classes, fatty acid composition and lipid peroxidation in the gills of the clam Donax Trunculus after permethrin exposure. Research Gate 2016, 186. [Google Scholar]
- Beninger, P.G. Seasonal variation of the mayor lipid classes in relation to the reproductive activity of two species of clams raised in a common habitat: Tapes decssatus L. (Jeffreys, 1863) and Philippinraum (Adams and Reeve), 1850. J. Exp. Mar. Bio. Ecol. 1984, 79, 79–90. [Google Scholar] [CrossRef]
- Prato, E.; Danieli, A.; Michele, M.; Francesca, B. Lipid and Fatty Acid Compositions of Mytilus galloprovincialis Cultured in the Mar Grande of Taranto (Southern Italy): Feeding Strategies and Trophic Relationships. Zool. Stud. 2010, 49, 211–219. [Google Scholar]
- Bernsdorff, C.; Winter, R. Differential properties of the sterols cholesterol, ergosterol, β-sitosterol, trans-7-dehydrocholesterol, stigmasterol and lanosterol on DPPC bilayer order. J. Phys. Chem. A 2003, 107, 10658–10664. [Google Scholar] [CrossRef]
- Chen, T.C.C. A comparison of the behavior of cholesterol, 7-dehydrocholesterol and ergosterol in phospholipid membranes. Biochim. Biophys. Acta 2012, 1818, 1673–1681. [Google Scholar] [CrossRef] [PubMed]
- Dennis, F.; Riccardo, G.; Agnese, P.; Deborah, P.; Barrara, Z.; Luciiano, B.; Enrico, M. Comparison of eleven extraction methods for quantitative evalution of total lipids and fatty acids in the clam Anadara Inaequivalvis (Bruguiere). J. Shellfish Res. 2013, 32, 285–290. [Google Scholar]
- Andray, B.I.; Ly, P.D.; Viacheslav, G.R.; Vasily, I.S. Fatty acid, Lipid class, and Phospholipid Molecular Species Composition of the Soft Coral Xenia sp. (Nha Trang Bay, the South China Sea, Vietnam). Springer 2015, 50, 575–589. [Google Scholar]
- Liu, Z.Y.; Zhou, D.-Y.; Wu, Z.-X.; Yin, F.-W.; Zhao, Q.; Xie, H.-K.; Zhang, J.-R.; Qin, L.; Shahidi, F. Extraction and detailed characterization of phospholipid-enriched oils from six species of edible clams. Food Chem. 2017, 239, 1175–1181. [Google Scholar] [CrossRef] [PubMed]
- Peterson, B.L.; Cummings, B.S. A review of chromatographic methods for the assessment of phospholipids in biological samples. Biomed. Chromatogr. 2006, 20, 227–243. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Phys. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Vaskovsky, V.E.; Terekhova, T.A. HPTLC of phospholipid mixtures containing phosphatidylglycerol. J. High. Resolut. Chromatogr. Chromatogr. Commun. 1979, 671–672. [Google Scholar] [CrossRef]
- Vaskovsky, V.E.; Kostetsky, E.Y.; Vasendin, I.M. A universal reagent for phospholipid analysis. J. Chromatogr. 1975, 114, 129–141. [Google Scholar] [CrossRef]
- Christie, W. Equivalent chain lengths of methyl ester derivatives of fatty acids on gas chromatography a reappraisal. J. Chromatogr. 1988, 447, 305–314. [Google Scholar] [CrossRef]
- Mass Spectrometry of Fatty Acid Derivatives. Available online: http://lipidlibrary.aocs.org/ms/masspec.html (accessed on 10 August 2014).
- Harrabi, S.; Herchi, W.; Kallel, H.; Mayer, P.; Boukhchina, S. Liquid chromatographic-mass spectrometric analysis of glycerophospholipids in corn oil. Food Chem. 2009, 114, 712–716. [Google Scholar] [CrossRef]
Sample Availability: Not available. |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Lipid Class | Content (%) |
|---|---|---|
| 1 | Hydrocarbon and wax (HW) | 2.6 ± 0.1 |
| 2 | Monoalkyl diacylglycerol (MADAG) | 1.3 ± 0.2 |
| 3 | Triacylglycerol (TG) | 11.7 ± 0.3 |
| 4 | Free fatty acids (FFA) | 18.6 ± 0.8 |
| 5 | Sterols (ST) | 20.2 ± 0.5 |
| 6 | Polar lipid (PoL) | 45.7 ± 0.6 |
| R.t (min) | Area | Content (%) | |
|---|---|---|---|
| Σ SAFAs | 1,971,006 | 26.3 | |
| 14:0 * | 3.748 | 291,352.4 | 3.9 |
| 16:0 * | 5.491 | 1,197,827 | 16.0 |
| 17:0 * | 6.812 | 105,581.2 | 1.4 |
| 18:0 * | 8.686 | 376,245.5 | 5.0 |
| Σ MUFAs | 1,191,078 | 15.9 | |
| 16:1n-7 * | 5.863 | 469,554.2 | 6.3 |
| 18:1n-9 * | 9.228 | 355,129.6 | 4.7 |
| 18:1n-7 * | 9.378 | 147,985.6 | 2.0 |
| 20:1n-11 | 15.324 | 99,722.7 | 1.3 |
| 20:1n-9 * | 15.511 | 42,995.4 | 0.6 |
| 20:1n-7 * | 15.875 | 75,690.3 | 1.0 |
| Σ PUFAs | 2,831,606 | 37.7 | |
| 16:3n-3 | 7.292 | 56,506.2 | 0.8 |
| 18:2n-6 * | 10.411 | 63,571.7 | 0.8 |
| 18:3n-3 | 12.383 | 77,148.8 | 1.0 |
| 18:4n-3 * | 13.518 | 80,259.3 | 1.1 |
| 20:2n-6 * | 17.775 | 108,771.3 | 1.4 |
| 20:3n-6 | 19.233 | 31,176.5 | 0.4 |
| 20:4n-6 * | 20.617 | 283,411 | 3.8 |
| 20:4n-3 * | 23.19 | 36,235.8 | 0.5 |
| 20:5n-3 * | 24.929 | 588,733 | 7.8 |
| 21:3n-3 | 28.661 | 85,752.1 | 1.1 |
| 21:5n-6 | 29.156 | 144,854.4 | 1.9 |
| 21:5n-3 | 33.552 | 53,585.8 | 0.7 |
| 22:4n-6 * | 36.426 | 115,438.2 | 1.5 |
| 22:5n-6 | 39.584 | 96,337.1 | 1.3 |
| 22:5n-3 * | 44.249 | 161,865.8 | 2.2 |
| 22:6n-3 * | 48.476 | 901,545.1 | 12.0 |
| Octadecanal a | 7.882 | 808,630.9 | 10.8 |
| Other b | 702,316.4 | 9.4 |
| Fraction | PE + PG | PC | CAEP | PS + PI |
|---|---|---|---|---|
| Σ SAFAs | 40.5 | 52.2 | 75.8 | 32.5 |
| 14:0 | 4.3 | nd | nd | 0.7 |
| 16:0 | 22.0 | 36.9 | 57.0 | 12.9 |
| 17:0 | 3.2 | 4.1 | 4.3 | 2.1 |
| 18:0 | 11.0 | 9.9 | 13.0 | 16.2 |
| 20:00 | nd | 1.3 | 1.5 | 0.4 |
| Σ MUFAs | 25.3 | 14.4 | 11.3 | 22.5 |
| 16:1n-7 | 10.9 | 3.6 | nd | 1.2 |
| 18:1n-9 | 8.4 | 7.9 | 8.5 | 9.5 |
| 18:1n-7 | 3.5 | 1.5 | 0.4 | 0.7 |
| 20:1n-11 | 1.1 | 0.5 | 0.3 | 7.4 |
| 20:1n-9 | 0.6 | 0.5 | 0.7 | 1.2 |
| 20:1n-7 | 0.8 | 0.4 | 1.4 | 2.5 |
| Σ PUFAs | 14.9 | 29.9 | 9.1 | 22.7 |
| 16:3n-3 | 0.9 | 0.8 | nd | nd |
| 18:2n-6 | 0.9 | 1.0 | 2.1 | 1.5 |
| 18:3n-3 | 1.2 | 0.5 | 0.1 | nd |
| 18:4n-3 | 0.8 | 0.6 | nd | nd |
| 20:2n-6 | nd | 1.2 | 0.2 | nd |
| 20:3n-6 | nd | 0.7 | 0.3 | nd |
| 20:4n-6 | 1.7 | 2.4 | 1.7 | 5.8 |
| 20:4n-3 | nd | 0.3 | nd | nd |
| 20:5n-3 | 2.9 | 3.8 | 1.2 | 1.4 |
| 21:3n-6 | nd | 2.6 | 0.8 | nd |
| 21:3n-3 | 2.2 | 0.4 | 0.2 | 4.9 |
| 21:5n-6 | nd | 0.8 | 0.3 | nd |
| 21:5n-3 | nd | 0.1 | 1.1 | nd |
| 22:4n-6 | nd | 1.0 | nd | 0.7 |
| 22:5n-6 | 1.1 | 0.9 | nd | 1.5 |
| 22:5n-3 | nd | 1.8 | nd | 1.3 |
| 22:6n-3 | 3.2 | 11.0 | 1.0 | 5.5 |
| Σ DMAs | 12.9 | 16.0 | ||
| 16:0 | 0.4 | nd | nd | nd |
| 17:0 | 1.2 | nd | nd | 1.5 |
| 18:0 | 11.3 | nd | nd | 14.6 |
| Other | 6.3 | 3.5 | 3.8 | 6.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tran, Q.T.; Le, T.T.T.; Pham, M.Q.; Do, T.L.; Vu, M.H.; Nguyen, D.C.; Bach, L.G.; Bui, L.M.; Pham, Q.L. Fatty Acid, Lipid Classes and Phospholipid Molecular Species Composition of the Marine Clam Meretrix lyrata (Sowerby 1851) from Cua Lo Beach, Nghe An Province, Vietnam. Molecules 2019, 24, 895. https://doi.org/10.3390/molecules24050895
Tran QT, Le TTT, Pham MQ, Do TL, Vu MH, Nguyen DC, Bach LG, Bui LM, Pham QL. Fatty Acid, Lipid Classes and Phospholipid Molecular Species Composition of the Marine Clam Meretrix lyrata (Sowerby 1851) from Cua Lo Beach, Nghe An Province, Vietnam. Molecules. 2019; 24(5):895. https://doi.org/10.3390/molecules24050895
Chicago/Turabian StyleTran, Quoc Toan, Thi Thanh Tra Le, Minh Quan Pham, Tien Lam Do, Manh Hung Vu, Duy Chinh Nguyen, Long Giang Bach, Le Minh Bui, and Quoc Long Pham. 2019. "Fatty Acid, Lipid Classes and Phospholipid Molecular Species Composition of the Marine Clam Meretrix lyrata (Sowerby 1851) from Cua Lo Beach, Nghe An Province, Vietnam" Molecules 24, no. 5: 895. https://doi.org/10.3390/molecules24050895
APA StyleTran, Q. T., Le, T. T. T., Pham, M. Q., Do, T. L., Vu, M. H., Nguyen, D. C., Bach, L. G., Bui, L. M., & Pham, Q. L. (2019). Fatty Acid, Lipid Classes and Phospholipid Molecular Species Composition of the Marine Clam Meretrix lyrata (Sowerby 1851) from Cua Lo Beach, Nghe An Province, Vietnam. Molecules, 24(5), 895. https://doi.org/10.3390/molecules24050895