
Evaluation of the Inhibitory Effects of (E)-1-(2-hydroxy-4,6-dimethoxyphenyl)-3-(naphthalen-1-yl)prop-2-en-1-one (DiNap), a Natural Product Analog, on the Replication of Type 2 PRRSV In Vitro and In Vivo

 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
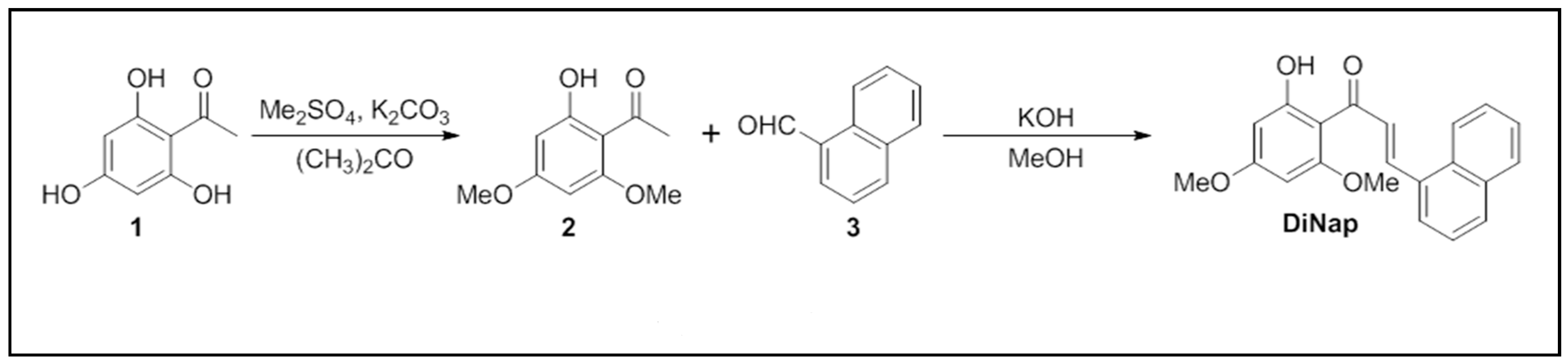
2.1. Chemistry
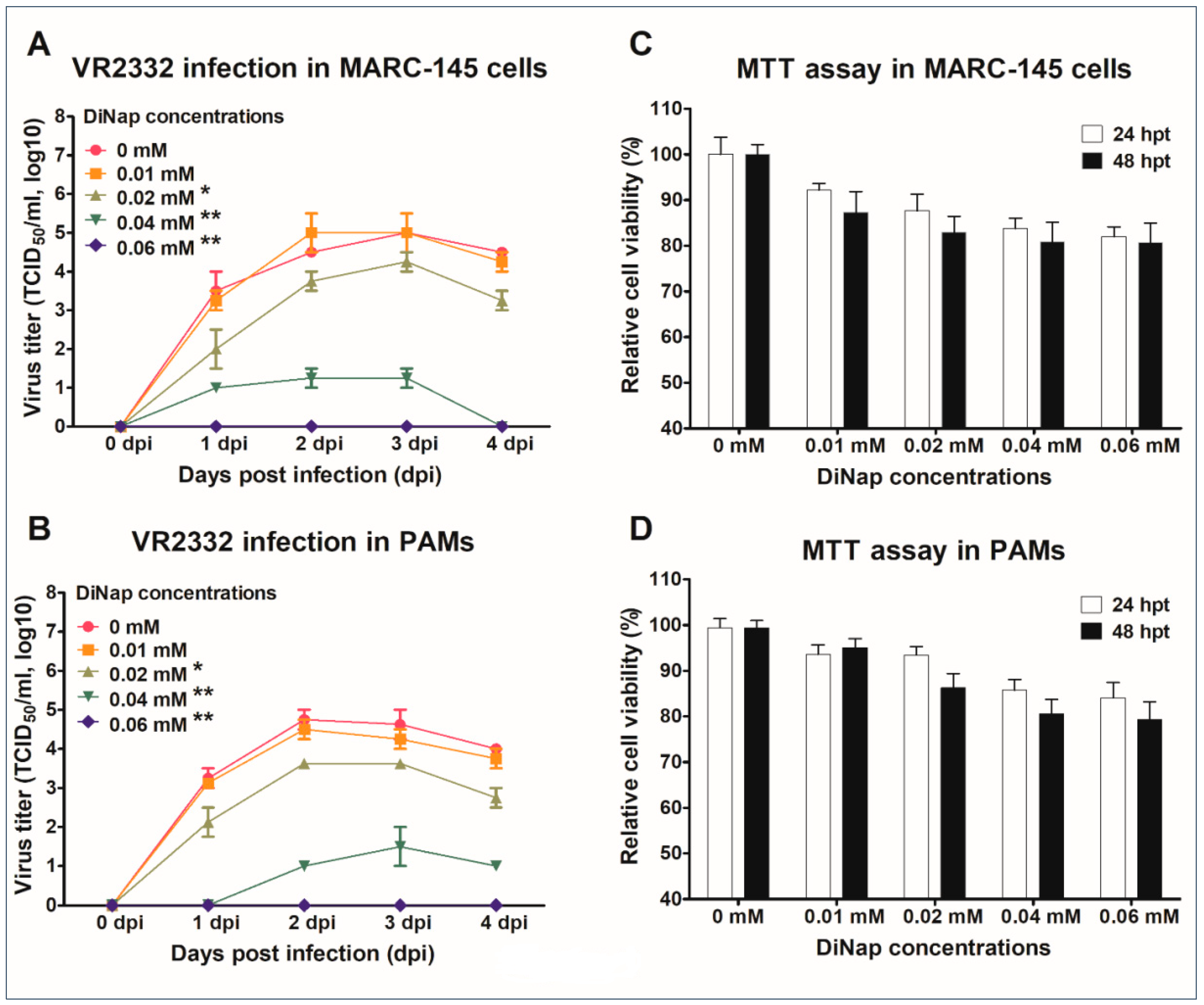
2.2. In Vitro Evaluation of the Effects of DiNap on PRRSV Replication
2.3. Inhibitory Effect of DiNap on PRRSV in Experimentally Infected Pigs
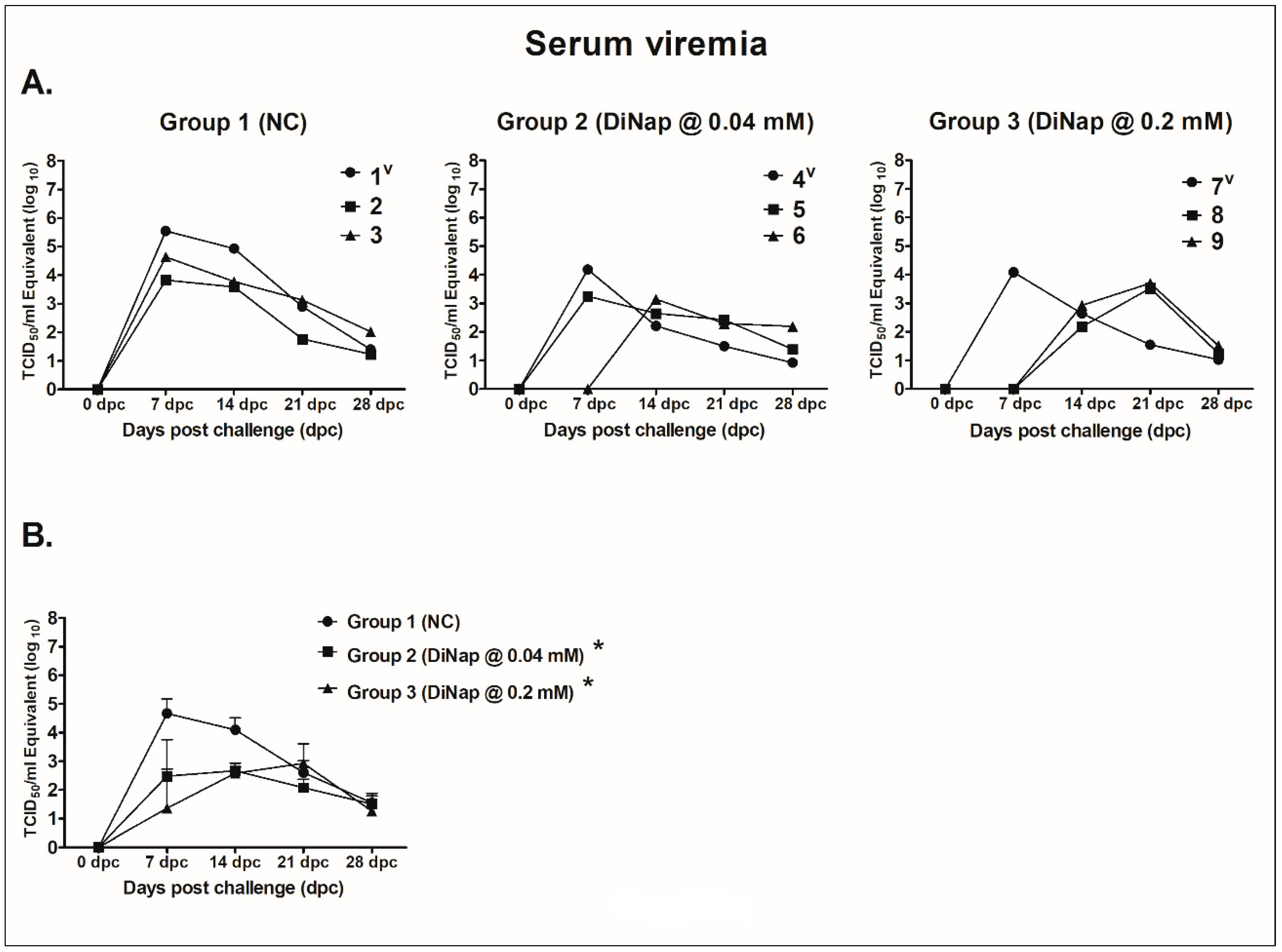
2.3.1. Serum Viremia
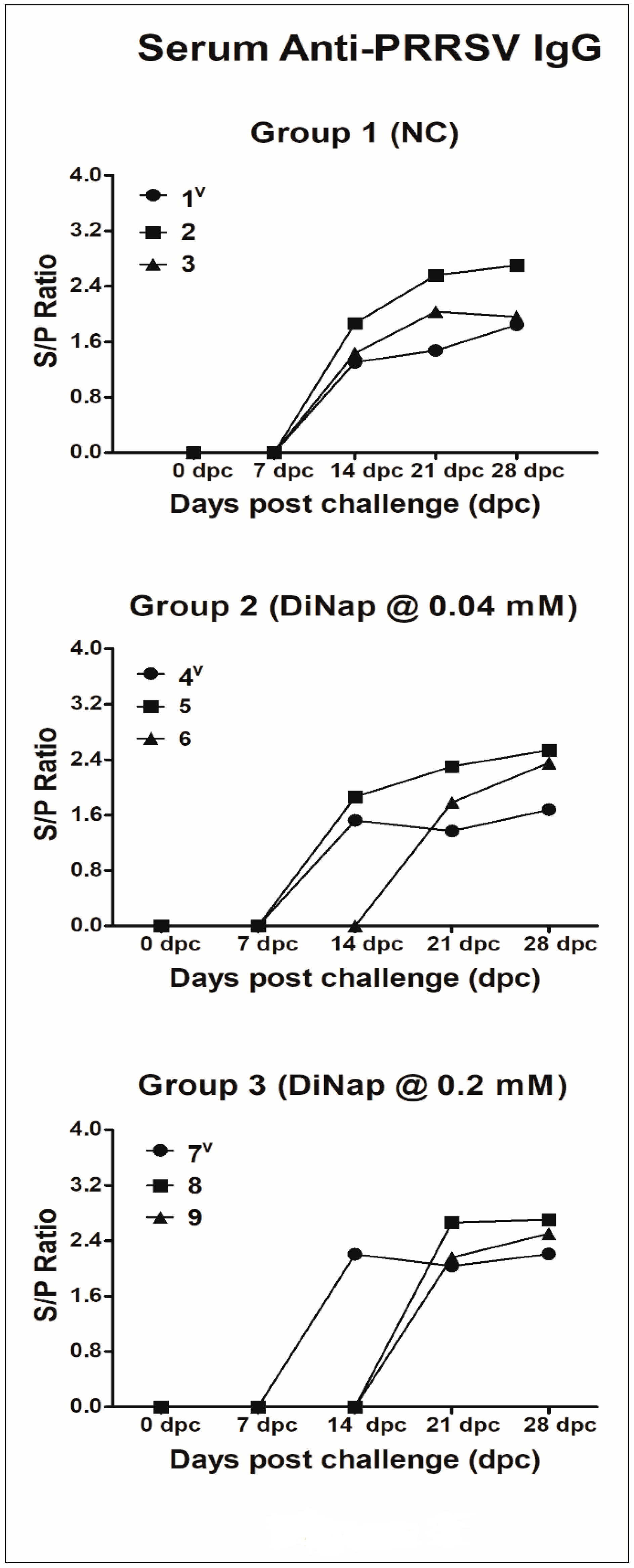
2.3.2. Anti-PRRSV IgG Response
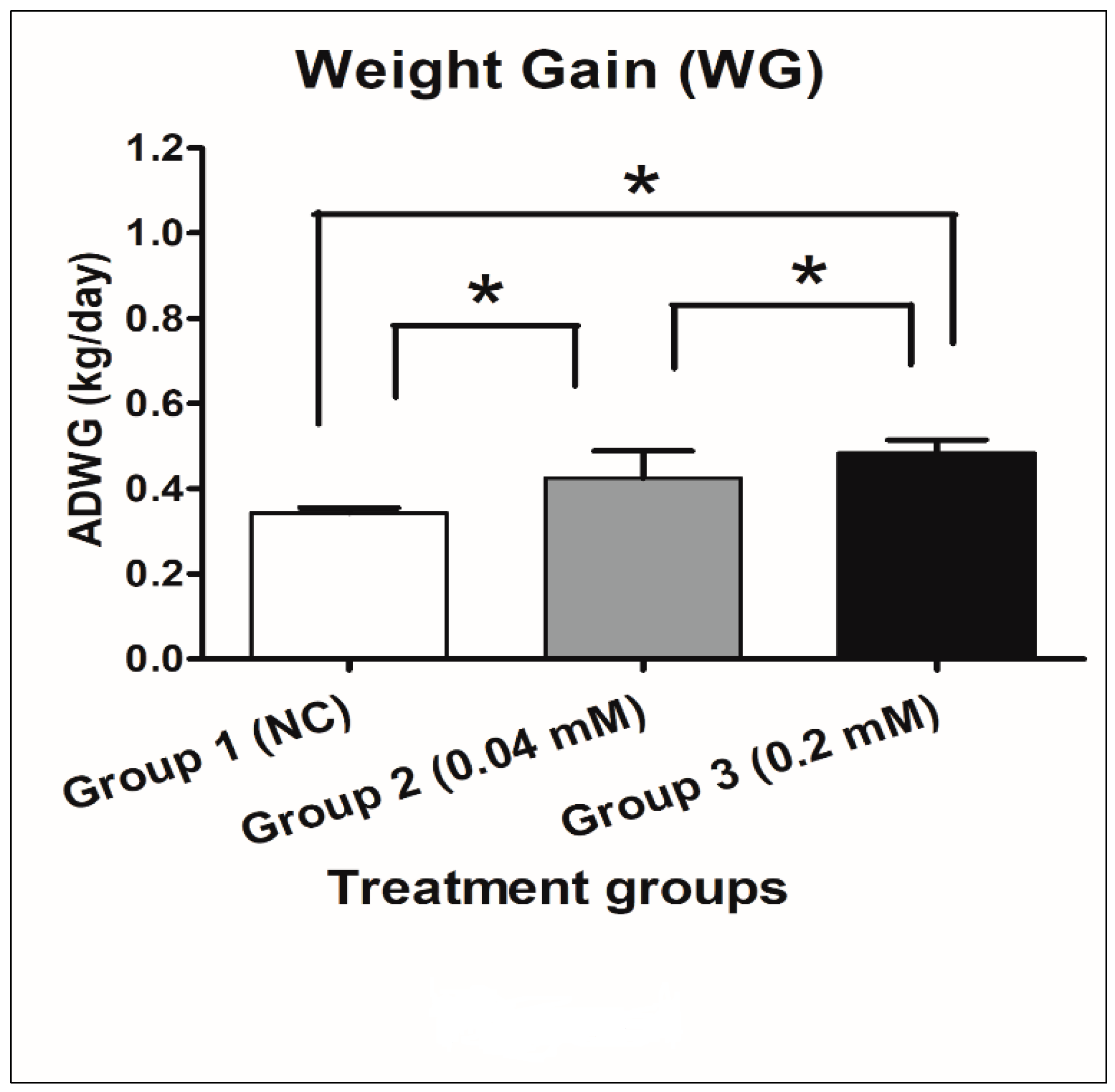
2.3.3. Average Daily Weight Gain
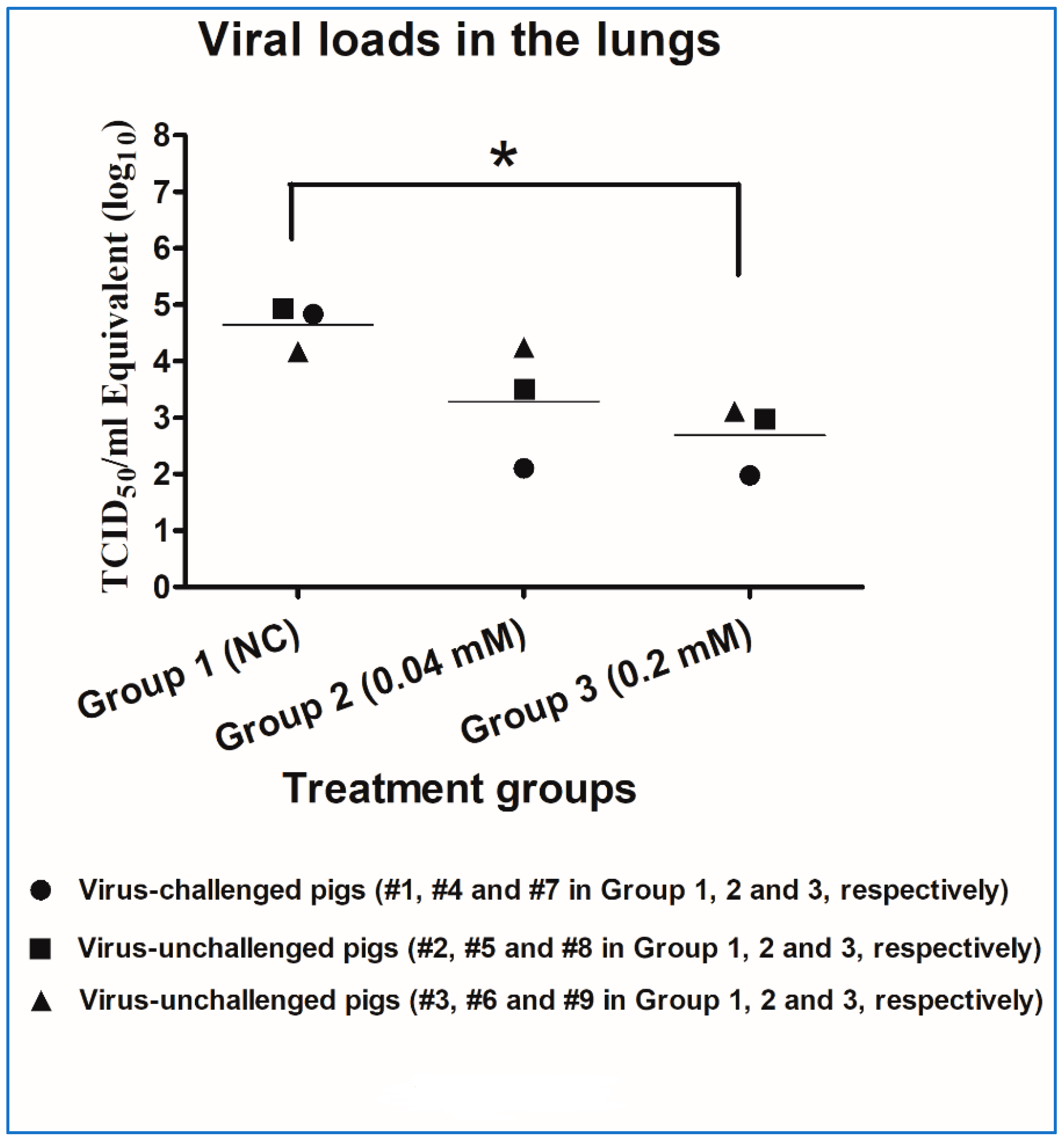
2.3.4. Lung Pathology and Residual Viral Loads in Lung Tissues
3. Discussion
4. Materials and Methods
4.1. Synthesis of DiNap, an Analog of a Natural Product
4.1.1. General Methods
4.1.2. 1-(2-Hydroxy-4,6-dimethoxyphenyl)ethanone (2)
4.1.3. (E)-1-(2-Hydroxy-4,6-dimethoxyphenyl)-3-(naphthalen-1-yl)prop-2-en-1-one (DiNap)
4.2. Viruses and Cells
4.3. Antiviral Effect of DiNap Against PRRSV Replication in Cells
4.4. MTT Assay
4.5. Animal Experiment and Samples
4.5.1. Serum ELISA
4.5.2. Lung Tissue Scoring and Pathology
4.5.3. Quantification of Viral Loads in Serum and Lung Tissue
4.6. Data Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Botner, A.; Strandbygaard, B.; Sorensen, K.J.; Have, P.; Madsen, K.G.; Madsen, E.S.; Alexandersen, S. Appearance of acute PRRS-like symptoms in sow herds after vaccination with a modified live PRRS vaccine. Vet. Rec. 1997, 141, 497–499. [Google Scholar] [CrossRef] [PubMed]
- Done, S.H.; Paton, D.J. Porcine Reproductive and Respiratory Syndrome—Clinical-Disease, Pathology and Immunosuppression. Vet. Rec. 1995, 136, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Holtkamp, D.J.; Kliebenstein, J.B.; Neumann, E.J.; Zimmerman, J.J.; Rotto, H.F.; Yoder, T.K.; Wang, C.; Yeske, P.E.; Mowrer, C.L.; Haley, C.A. Assessment of the economic impact of porcine reproductive and respiratory syndrome virus on United States pork producers. J. Swine Health Prod. 2013, 21, 72–84. [Google Scholar]
- Cavanagh, D. Nidovirales: A new order comprising Coronaviridae and Arteriviridae. Arch. Virol. 1997, 142, 629–633. [Google Scholar] [PubMed]
- Meulenberg, J.J.M.; Hulst, M.M.; Demeijer, E.J.; Moonen, P.L.J.M.; Denbesten, A.; Dekluyver, E.P.; Wensvoort, G.; Moormann, R.J.M. Lelystad Virus, the Causative Agent of Porcine Epidemic Abortion and Respiratory Syndrome (Pears), Is Related to Ldv and Eav. Virology 1993, 192, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Murtaugh, M.P.; Elam, M.R.; Kakach, L.T. Comparison of the structural protein coding sequences of the VR-2332 and Lelystad virus strains of the PRRS virus. Arch. Virol. 1995, 140, 1451–1460. [Google Scholar] [CrossRef] [PubMed]
- Firth, A.E.; Zevenhoven-Dobbe, J.C.; Wills, N.M.; Go, Y.Y.; Balasuriya, U.B.R.; Atkins, J.F.; Snijder, E.J.; Posthuma, C.C. Discovery of a small arterivirus gene that overlaps the GP5 coding sequence and is important for virus production. J. Gen. Virol. 2011, 92, 1097–1106. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Yoo, D. Engineering the PRRS virus genome: Updates and perspectives. Vet. Microbiol. 2014, 174, 279–295. [Google Scholar] [CrossRef] [PubMed]
- International Committee on Taxonomy of Viruses. Available online: https://talk.ictvonline.org/taxonomy/p/taxonomy-history?taxnode_id=20171832,id=20181833 (accessed on 20 February 2018).
- Shi, M.; Lam, T.T.; Hon, C.C.; Hui, R.K.; Faaberg, K.S.; Wennblom, T.; Murtaugh, M.P.; Stadejek, T.; Leung, F.C. Molecular epidemiology of PRRSV: A phylogenetic perspective. Virus Res. 2010, 154, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhang, C. Porcine reproductive and respiratory syndrome virus vaccines: Current status and strategies to a universal vaccine. Transbound. Emerg. Dis. 2014, 61, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Gravina, H.D.; Tafuri, N.F.; Silva, A., Jr.; Fietto, J.L.; Oliveira, T.T.; Diaz, M.A.; Almeida, M.R. In vitro assessment of the antiviral potential of trans-cinnamic acid, quercetin and morin against equid herpesvirus 1. Res. Vet. Sci. 2011, 91, e158–e162. [Google Scholar] [CrossRef] [PubMed]
- Pang, R.; Tao, J.Y.; Zhang, S.L.; Zhao, L.; Yue, X.; Wang, Y.F.; Ye, P.; Dong, J.H.; Zhu, Y.; Wu, J.G. In vitro antiviral activity of lutein against hepatitis B virus. Phytother. Res. 2010, 24, 1627–1630. [Google Scholar] [CrossRef] [PubMed]
- Schnepf, N.; Corvo, J.; Pors, M.J.; Mazeron, M.C. Antiviral activity of ganciclovir and artesunate towards human cytomegalovirus in astrocytoma cells. Antivir. Res. 2011, 89, 186–188. [Google Scholar] [CrossRef] [PubMed]
- Glatthaar-Saalmüller, B.; Rauchhaus, U.; Rode, S.; Haunschild, J.; Saalmüller, A. Antiviral activity in vitro of two preparations of the herbal medicinal product Sinupret® against viruses causing respiratory infections. Phytomedicine 2011, 15, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Karuppannan, A.K.; Wu, K.X.; Qiang, J.; Chu, J.J.; Kwang, J. Natural compounds inhibiting the replication of Porcine reproductive and respiratory syndrome virus. Antivir. Res. 2012, 94, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Sun, N.; Zhao, X.; Niu, L.; Song, M.; Sun, Y.; Jiang, J.; Guo, J.; Bai, Y.; He, J.; et al. In vitro screening for compounds derived from traditional chinese medicines with antiviral activities against porcine reproductive and respiratory syndrome virus. J. Microbiol. Biotechnol. 2013, 23, 1076–1083. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Zhang, W.; Wang, L.; Zhu, M.; Wang, H.; Feng, W.H.; Ng, T.B. A novel compound from the mushroom Cryptoporus volvatus inhibits porcine reproductive and respiratory syndrome virus (PRRSV) in vitro. PLoS ONE 2013, 8, e79333. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Zhang, W.; Sun, Y.; Yang, Q.; Ren, J.; Liu, J.; Wang, H.; Feng, W.H. Cryptoporus volvatus extract inhibits porcine reproductive and respiratory syndrome virus (PRRSV) in vitro and in vivo. PLoS ONE 2013, 8, e63767. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Liu, S.; Li, C.; Yang, L.; Zu, Y. In vitro evaluation of the antiviral activity of the synthetic epigallocatechin gallate analog-epigallocatechin gallate (EGCG) palmitate against porcine reproductive and respiratory syndrome virus. Viruses 2014, 6, 938–950. [Google Scholar] [CrossRef] [PubMed]
- Sun, N.; Zhao, X.; Bai, X.Y.; Niu, L.; Song, M.Q.; Sun, Y.G.; Jiang, J.B.; Li, H.Q. Anti-PRRSV effect and mechanism of sodium tanshinone IIA sulfonate in vitro. J. Asian Nat. Prod. Res. 2012, 14, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Jung, B.G.; Lee, J.A.; Lee, B.J. Antiviral effect of dietary germanium biotite supplementation in pigs experimentally infected with porcine reproductive and respiratory syndrome virus. J. Vet. Sci. 2013, 14, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Pringproa, K.; Khonghiran, O.; Kunanoppadol, S.; Potha, T.; Chuammitri, P. In Vitro Virucidal and Virustatic Properties of the Crude Extract of Cynodon dactylon against Porcine Reproductive and Respiratory Syndrome Virus. Vet. Med. Int. 2014, 2014, 947589. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Sun, N.; Zhao, J.X.; Sun, Y.G.; Huang, J.G.; Lei, H.M.; Guo, J.H.; Hu, Y.L.; Wang, W.K.; Li, H.Q. In vitro evaluation of antiviral activity of tea seed saponins against porcine reproductive and respiratory syndrome virus. Antivir. Ther. 2015, 20, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Chen, H.; Chen, K.; Gao, Y.; Gao, S.; Liu, X.; Li, J. Sulfated modification can enhance antiviral activities of Achyranthes bidentata polysaccharide against porcine reproductive and respiratory syndrome virus (PRRSV) in vitro. Int. J. Biol. Macromol. 2013, 52, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Sun, N.; Wang, Z.W.; Wu, C.H.; Li, E.; He, J.P.; Wang, S.Y.; Hu, Y.L.; Lei, H.M.; Li, H.Q. Antiviral activity and underlying molecular mechanisms of Matrine against porcine reproductive and respiratory syndrome virus in vitro. Res. Vet. Sci. 2014, 96, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Duan, E.; Wang, D.; Fang, L.; Ma, J.; Luo, J.; Chen, H.; Li, K.; Xiao, S. Suppression of porcine reproductive and respiratory syndrome virus proliferation by glycyrrhizin. Antivir. Res. 2015, 120, 122–125. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Bai, X.; Cui, T.; Zhou, H.; Chen, Y.; Xie, J.; Shi, Q.; Wang, H.; Zhang, G. In Vitro Antiviral Activity of Germacrone Against Porcine Reproductive and Respiratory Syndrome Virus. Curr. Microbiol. 2016, 73, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Duan, H.; Li, N.; Zhao, L.; Pu, F.; Huang, B.; Wu, C.; Nan, Y.; Du, T.; Mu, Y.; et al. Heme oxygenase-1 metabolite biliverdin, not iron, inhibits porcine reproductive and respiratory syndrome virus replication. Free Radic. Biol. Med. 2017, 102, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Sakagami, H.; Masuda, Y.; Tomomura, M.; Yokose, S.; Uesawa, Y.; Ikezoe, N.; Asahara, D.; Takao, K.; Kanamoto, T.; Terakubo, S.; et al. Quantitative Structure-Cytotoxicity Relationship of Chalcones. Anticancer Res. 2017, 37, 1091–1098. [Google Scholar] [PubMed]
- Naithani, R.; Huma, L.C.; Holland, L.E.; Shukla, D.; McCormick, D.L.; Mehta, R.G.; Moriarty, R.M. Antiviral activity of phytochemicals: A comprehensive review. Mini Rev. Med. Chem. 2008, 8, 1106–1133. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.R.; Prasad, S.; Sung, B.; Aggarwal, B.B. The role of chalcones in suppression of NF-kappaB-mediated inflammation and cancer. Int. Immunopharmacol. 2011, 11, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Nowakowska, Z. A review of anti-infective and anti-inflammatory chalcones. Eur. J. Med. Chem. 2007, 42, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Dawane, B.S.; Konda, S.G.; Shaikh, B.M.; Bhosale, R.B. An improved procedure for synthesis of some new 1,3-diaryl-2-propen-1-ones using PEG-400 as a recyclable solvent and their antimicrobial evaluation. Acta Pharm. 2009, 59, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Dimmock, J.R.; Elias, D.W.; Beazely, M.A.; Kandepu, N.M. Bioactivities of chalcones. Curr. Med. Chem. 1999, 6, 1125–1149. [Google Scholar] [PubMed]
- Go, M.L.; Wu, X.; Liu, X.L. Chalcones: An update on cytotoxic and chemoprotective properties. Curr. Med. Chem. 2005, 12, 483–499. [Google Scholar] [CrossRef]
- Ahmad, A.L.; Dowsett, A.B.; Tyrrell, D.A. Studies of rhinovirus resistant to an antiviral chalcone. Antivir. Res. 1987, 8, 27–39. [Google Scholar] [CrossRef]
- Gutierrez, R.M.P.; Muniz-Ramirez, A.; Sauceda, J.V. Review: The potential of chalcones as a source of drugs. Afr. J. Pharm. Pharmacol. 2015, 9, 237–257. [Google Scholar]
- Chang, C.T.; Hseu, Y.C.; Thiyagarajan, V.; Lin, K.Y.; Way, T.D.; Korivi, M.; Liao, J.W.; Yang, H.L. Chalcone flavokawain B induces autophagic-cell death via reactive oxygen species-mediated signaling pathways in human gastric carcinoma and suppresses tumor growth in nude mice. Arch. Toxicol. 2017, 91, 3341–3364. [Google Scholar] [CrossRef] [PubMed]
- Zenger, K.; Agnolet, S.; Schneider, B.; Kraus, B. Biotransformation of Flavokawains A, B, and C, Chalcones from Kava (Piper methysticum), by Human Liver Microsomes. J. Agric. Food Chem. 2015, 63, 6376–6385. [Google Scholar] [CrossRef] [PubMed]
- Baez-Santos, Y.M.; Barraza, S.J.; Wilson, M.W.; Agius, M.P.; Mielech, A.M.; Davis, N.M.; Baker, S.C.; Larsen, S.D.; Mesecar, A.D. X-ray structural and biological evaluation of a series of potent and highly selective inhibitors of human coronavirus papain-like proteases. J. Med. Chem. 2014, 57, 2393–2412. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Cao, S.; Hevener, K.E.; Truong, L.; Gatuz, J.L.; Patel, K.; Ghosh, A.K.; Johnson, M.E. Synergistic inhibitor binding to the papain-like protease of human SARS coronavirus: Mechanistic and inhibitor design implications. Chem. Med. Chem. 2013, 8, 1361–1372. [Google Scholar] [CrossRef] [PubMed]
- Mishra, L.; Sinha, R.; Itokawa, H.; Bastow, K.F.; Tachibana, Y.; Nakanishi, Y.; Kilgore, N.; Lee, K.H. Anti-HIV and cytotoxic activities of Ru(II)/Ru(III) polypyridyl complexes containing 2,6-(2′-benzimidazolyl)-pyridine/chalcone as co-ligand. Bioorg. Med. Chem. 2001, 9, 1667–1671. [Google Scholar] [CrossRef]
- Vogel, S.; Barbic, M.; Jurgenliemk, G.; Heilmann, J. Synthesis, cytotoxicity, anti-oxidative and anti-inflammatory activity of chalcones and influence of A-ring modifications on the pharmacological effect. Eur. J. Med. Chem. 2010, 45, 2206–2213. [Google Scholar] [CrossRef] [PubMed]
- Mahapatra, D.K.; Bharti, S.K.; Asati, V. Chalcone Derivatives: Anti-inflammatory Potential and Molecular Targets Perspectives. Curr. Top Med. Chem. 2017, 17, 3146–3169. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.H.; Park, S.Y. Synthesis of Flavokawain Analogues and their Anti-neoplastic Effects on Drug-resistant Cancer Cells Through Hsp90 Inhibition. Bull. Korean Chem. Soc. 2014, 35, 1154–1158. [Google Scholar] [CrossRef]
- Kim, H.S.; Kwang, J.; Yoon, I.J.; Joo, H.S.; Frey, M.L. Enhanced replication of porcine reproductive and respiratory syndrome (PRRS) virus in a homogeneous subpopulation of MA-104 cell line. Arch. Virol. 1993, 133, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Laguna, J.; Salguero, F.J.; Pallares, F.J.; Carrasco, L. Immunopathogenesis of porcine reproductive and respiratory syndrome in the respiratory tract of pigs. Vet. J. 2013, 195, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Van Breedam, W.; Delputte, P.L.; Van Gorp, H.; Misinzo, G.; Vanderheijden, N.; Duan, X.; Nauwynck, H.J. Porcine reproductive and respiratory syndrome virus entry into the porcine macrophage. J. Gen. Virol. 2010, 91, 1659–1667. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.J.; Wu, L.L.; Zimmerman, J.J.; Hill, H.T.; Platt, K.B. Antibody-dependent enhancement (ADE) of porcine reproductive and respiratory syndrome virus (PRRSV) infection in pigs. Viral Immunol. 1996, 9, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Khatun, A.; Shabir, N.; Seo, B.J.; Kim, B.S.; Yoon, K.J.; Kim, W.I. The Attenuation Phenotype of a Ribavirin-Resistant Porcine Reproductive and Respiratory Syndrome Virus Is Maintained during Sequential Passages in Pigs. J. Virol. 2016, 90, 4454–4468. [Google Scholar] [CrossRef] [PubMed]
- Greig, A. The use of a microtitration technique for the routine assay of African swine fever virus. Brief Report. Arch. Virol. 1975, 47, 287–289. [Google Scholar] [CrossRef] [PubMed]
- Khatun, A.; Shabir, N.; Yoon, K.-J.; Kim, W.-I. Effect of Ribavirin on the replication and genetic stability of porcine reproductive and respiratory syndrome virus. BMC Vet. Res. 2015, 11, 21. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Xu, J.M.; Song, S.T.; Tang, Z.M.; Jiang, Z.F.; Liu, X.Q.; Zhou, L.; Zhang, J.; Liu, X.W. Predictive chemotherapy of advanced breast cancer directed by MTT assay in vitro. Breast Cancer Res. Treat. 1999, 53, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Halbur, P.G.; Paul, P.S.; Frey, M.L.; Landgraf, J.; Eernisse, K.; Meng, X.J.; Lum, M.A.; Andrews, J.J.; Rathje, J.A. Comparison of the pathogenicity of two US porcine reproductive and respiratory syndrome virus isolates with that of the Lelystad virus. Vet. Pathol. 1995, 32, 648–660. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; McKeown, N.E.; Harmon, K.L.; Meng, X.J.; Halbur, P.G. Porcine circovirus type 2 infection decreases the efficacy of a modified live porcine reproductive and respiratory syndrome virus vaccine. Clin. Vaccine Immunol. 2006, 13, 923–929. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khatun, A.; Park, S.Y.; Shabir, N.; Nazki, S.; Kang, A.-R.; Jeong, C.-G.; Seo, B.-J.; Yang, M.-S.; Kim, B.; Seo, Y.H.; et al. Evaluation of the Inhibitory Effects of (E)-1-(2-hydroxy-4,6-dimethoxyphenyl)-3-(naphthalen-1-yl)prop-2-en-1-one (DiNap), a Natural Product Analog, on the Replication of Type 2 PRRSV In Vitro and In Vivo. Molecules 2019, 24, 887. https://doi.org/10.3390/molecules24050887
Khatun A, Park SY, Shabir N, Nazki S, Kang A-R, Jeong C-G, Seo B-J, Yang M-S, Kim B, Seo YH, et al. Evaluation of the Inhibitory Effects of (E)-1-(2-hydroxy-4,6-dimethoxyphenyl)-3-(naphthalen-1-yl)prop-2-en-1-one (DiNap), a Natural Product Analog, on the Replication of Type 2 PRRSV In Vitro and In Vivo. Molecules. 2019; 24(5):887. https://doi.org/10.3390/molecules24050887
Chicago/Turabian StyleKhatun, Amina, Sun You Park, Nadeem Shabir, Salik Nazki, A-Rum Kang, Chang-Gi Jeong, Byoung-Joo Seo, Myeon-Sik Yang, Bumseok Kim, Young Ho Seo, and et al. 2019. "Evaluation of the Inhibitory Effects of (E)-1-(2-hydroxy-4,6-dimethoxyphenyl)-3-(naphthalen-1-yl)prop-2-en-1-one (DiNap), a Natural Product Analog, on the Replication of Type 2 PRRSV In Vitro and In Vivo" Molecules 24, no. 5: 887. https://doi.org/10.3390/molecules24050887
APA StyleKhatun, A., Park, S. Y., Shabir, N., Nazki, S., Kang, A.-R., Jeong, C.-G., Seo, B.-J., Yang, M.-S., Kim, B., Seo, Y. H., & Kim, W.-I. (2019). Evaluation of the Inhibitory Effects of (E)-1-(2-hydroxy-4,6-dimethoxyphenyl)-3-(naphthalen-1-yl)prop-2-en-1-one (DiNap), a Natural Product Analog, on the Replication of Type 2 PRRSV In Vitro and In Vivo. Molecules, 24(5), 887. https://doi.org/10.3390/molecules24050887