Antioxidant and Hepatoprotective Effects of Croton hypoleucus Extract in an Induced-Necrosis Model in Rats

 , ,
, ,  and
and
Abstract
1. Introduction
2. Results
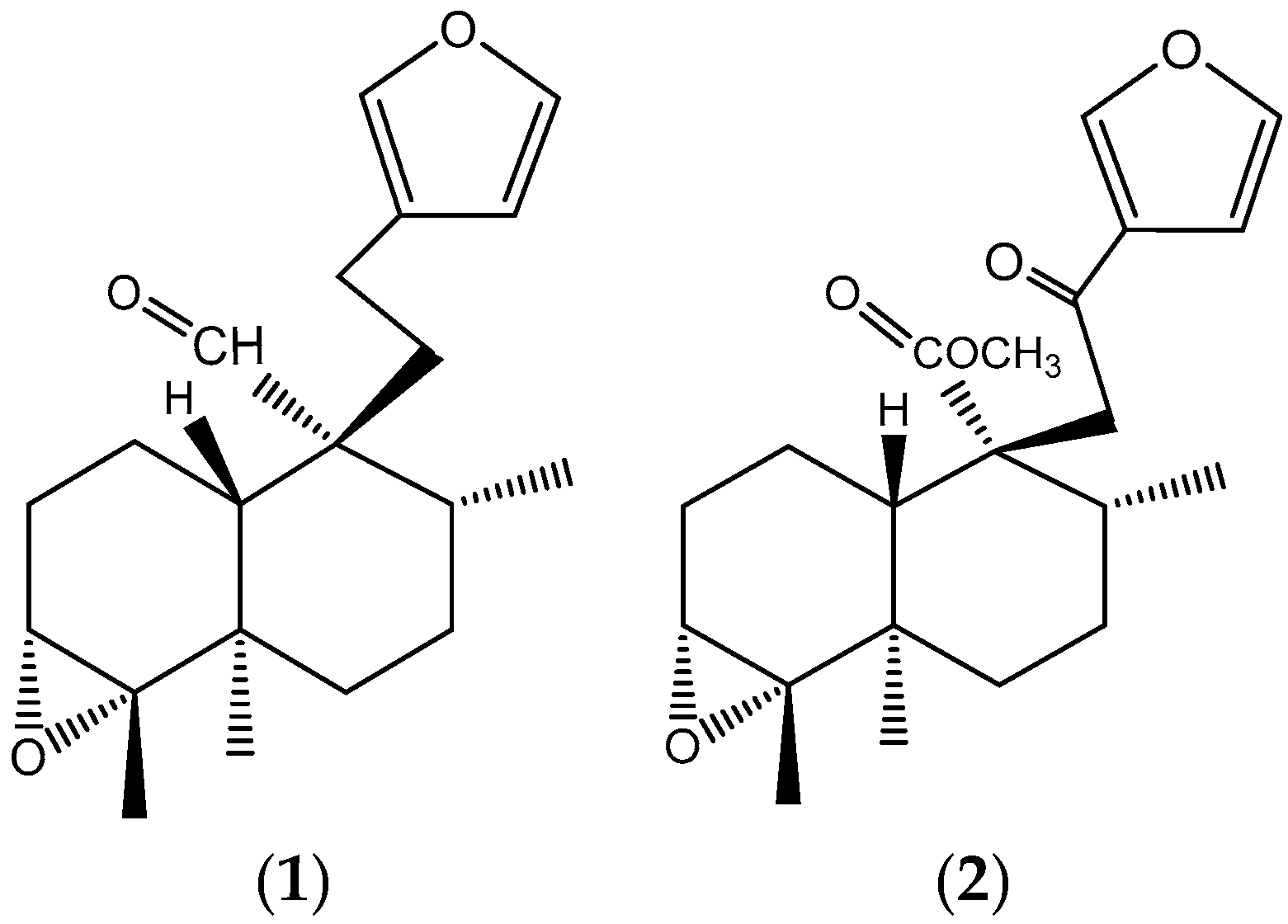
2.1. Purification of Main Compounds in Dichloromethane Fraction
2.2. Antioxidant Activity of EC
2.3. Acute Toxicity of EC
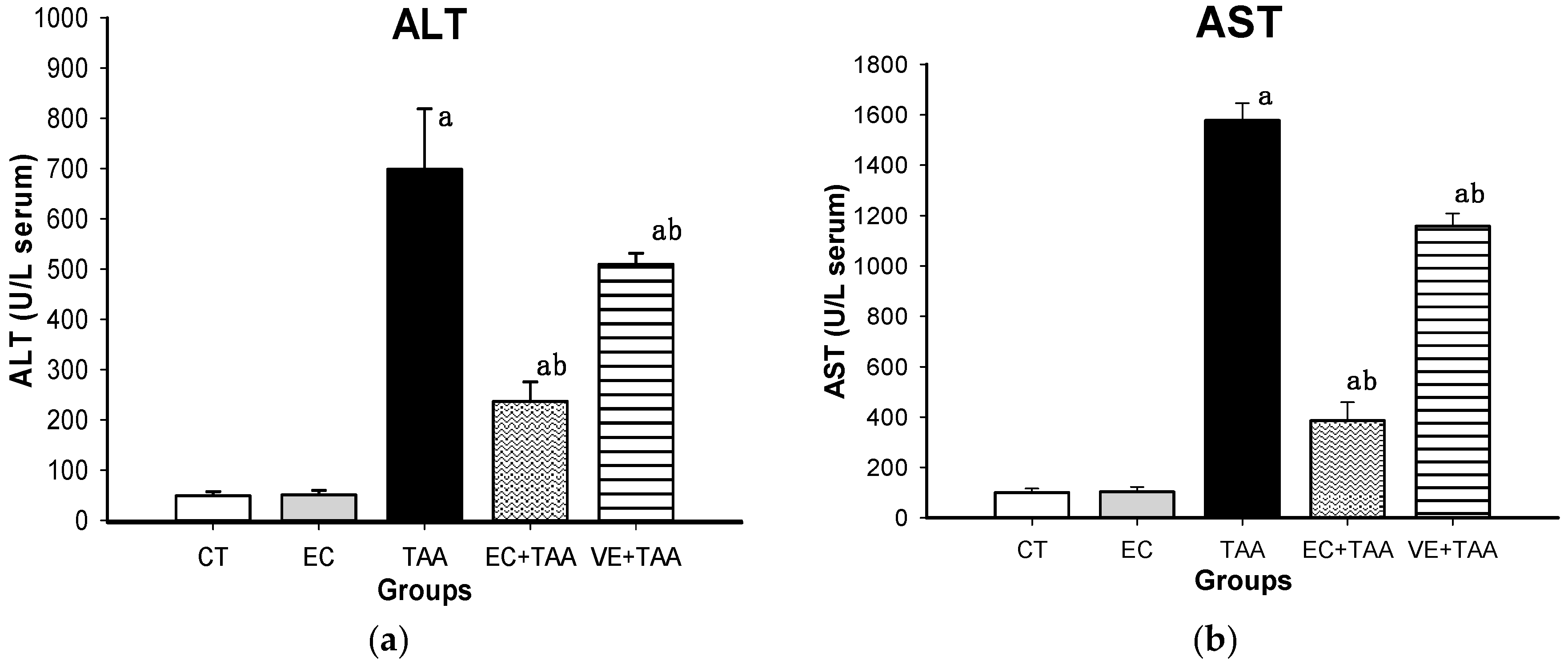
2.4. Liver Damage Biomarkers
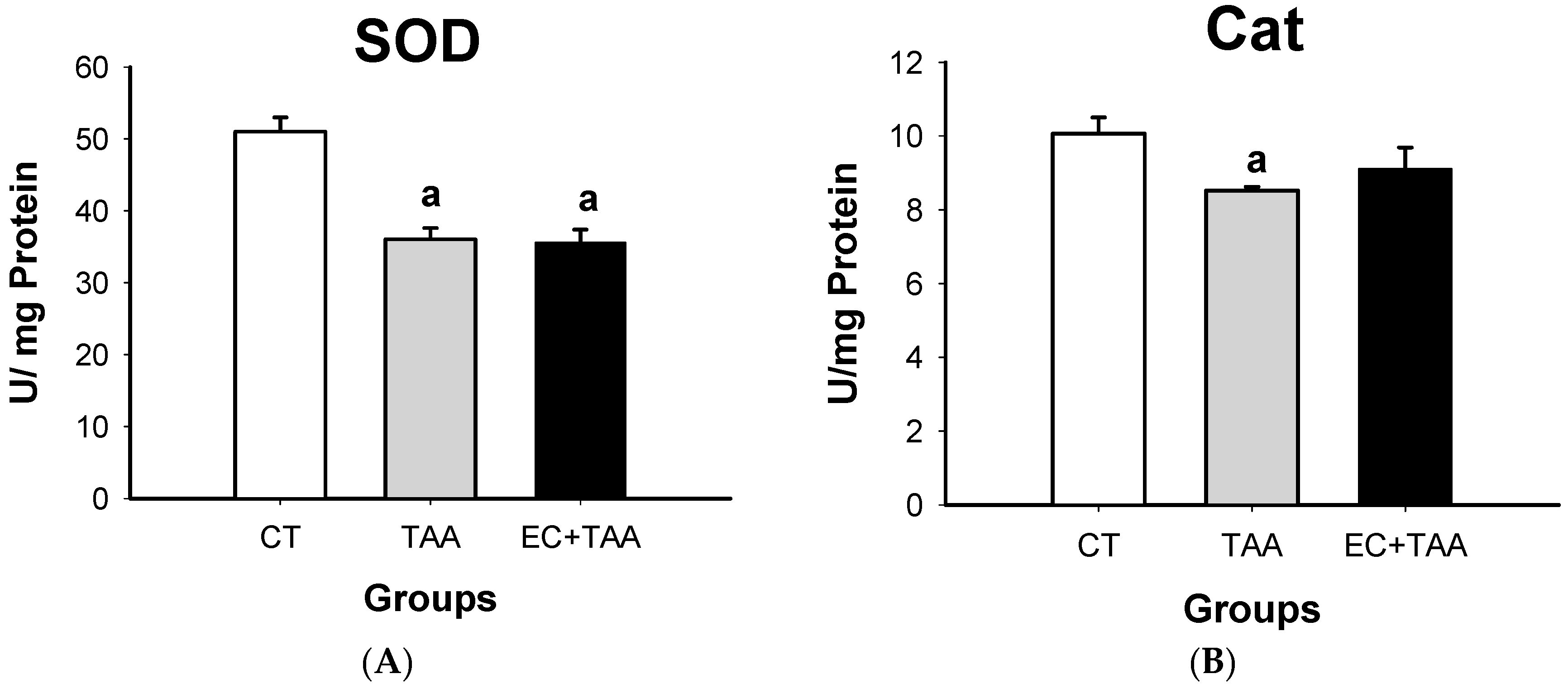
2.5. SOD and Cat Evaluation from Antioxidant Enzyme Activity
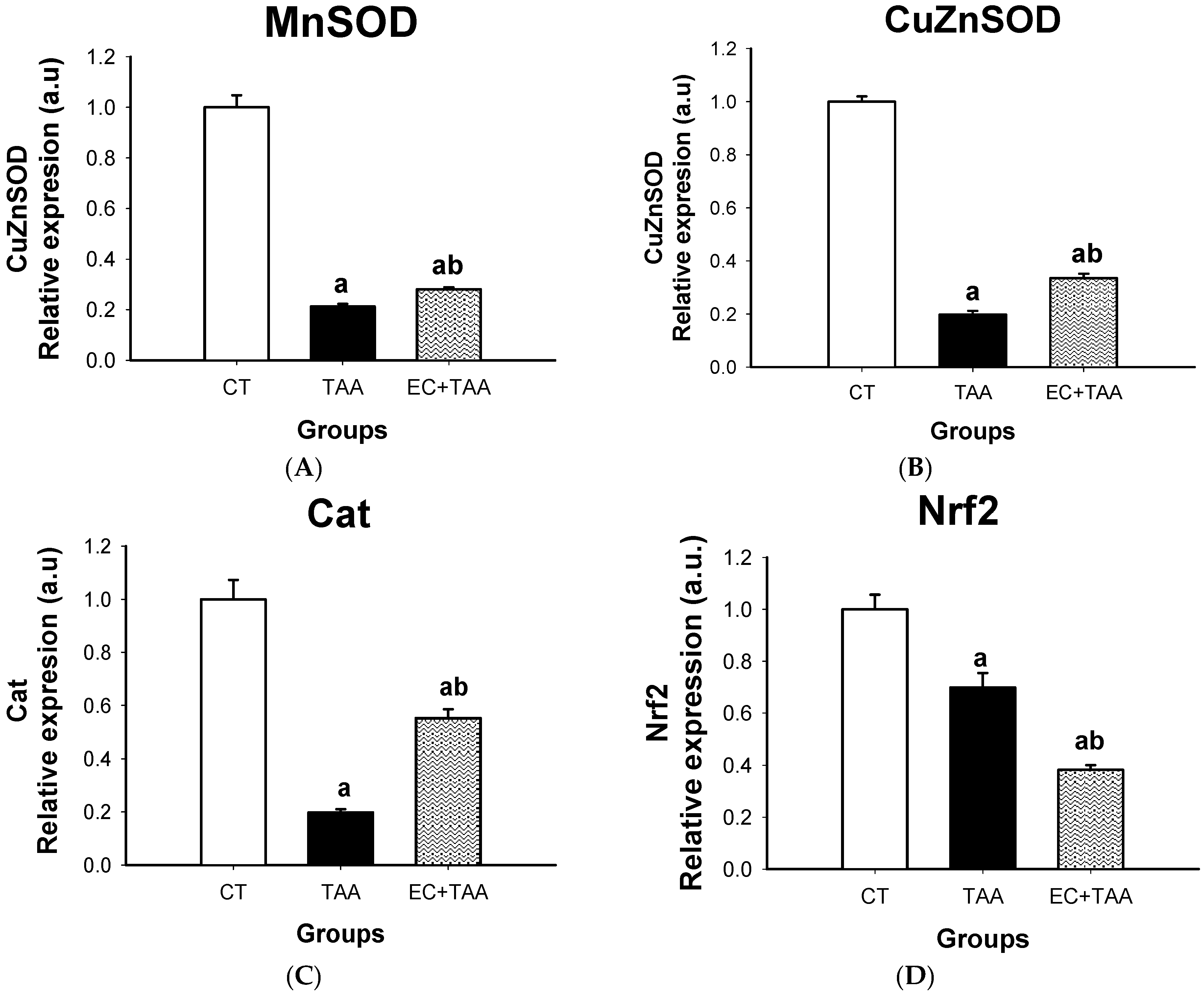
2.6. MnSOD, CuZnSOD, Cat, and Nrf2 mRNA Expression
3. Discussion
3.1. Main Compounds in Dichloromethane Fraction
3.2. Antioxidant Activity of EC
3.3. Acute Toxicity of EC
3.4. Liver Damage Biomarkers
3.5. SOD and Cat System
3.6. MnSOD, CuZnSOD, Cat, and Nrf2 mRNA Gene Expression
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Preparation of EC
4.3. Purification of Main Compounds in Dichloromethane Fraction
4.4. Antioxidant Capacity of EC
4.5. Experimental Animals
4.6. Acute Toxicity
4.7. Thioacetamide-Induced Hepatotoxicity
4.7.1. Biochemical Parameters
4.7.2. SOD and Cat in Vitro Activity
4.7.3. Extraction and Quantification of RNA by RT-PCR
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bonini, S.A.; Premoli, M.; Tambaro, S.; Kumar, A.; Maccarinelli, G.; Memo, M.; Mastinu, A. Cannabis sativa: A comprehensive ethnopharmacological review of a medicinal plant with a long history. J. Ethnopharmacol. 2018, 227, 300–315. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). The World Medicines Situation, Traditional Medicines: Global Situation, Issues and Challenges; WHO: Geneva, Switzerland, 2011; 12p. [Google Scholar]
- Salatino, A.; Salatino, M.L.F.; Negri, G. Traditional uses, chemistry and pharmacology of Croton species (Euphorbiaceae). J. Braz. Chem. Soc. 2007, 18, 11–33. [Google Scholar] [CrossRef]
- Kumar, A.; Premoli, M.; Bonini, S.A.; Maccarinelli, G.; Gianoncelli, A.; Memo, M.; Mastinu, A. Cannabimimetic plants: Are they new cannabbinoidergic modulators? Planta 2019, 269, 1681–1694. [Google Scholar] [CrossRef] [PubMed]
- Adewusi, E.A.; Afolayan, A.J. A review of natural products with hepatoprotective activity. JMPR 2010, 4, 1318–1334. [Google Scholar] [CrossRef]
- Govind, P. Medicinal plants against liver diseases. Int. Res. J. Pharm. 2011, 2, 115–151. [Google Scholar]
- Jannu, V.; Baddam, P.G.; Boorgula, A.K.; Jambula, S.R. A Review on Hepatoprotective Plants. Int. J. Drug Dev. Res. 2012, 4, 1–8. [Google Scholar]
- Lima, F.C.; Sousa, D.F.; Ferreira, J.M. Croton zehntneri essential oil prevents acetaminophen- induced acute hepatotoxicity in mice. Rec. Nat. Prod. 2008, 2, 135–140. [Google Scholar]
- Ahmed, B.; Alam, T.; Varshney, M.; Khan, S.A. Hepatoprotective activity of two plants belonging to the Apiaceae and the Euphorbiaceae family. J. Ethnopharmacol. 2002, 79, 313–316. [Google Scholar] [CrossRef]
- Jaya, S.E.; Beaulah, A.; Sadiq, A.M.; Chakkaravaarthy, M.V. Hepatoprotective activity of methanolic extract of Croton sparciflorus on DEN induced hepatotoxicity in wistar albino rats. J. Pharm. Chem. Biol. Sci. 2014, 4, 1002–1011. [Google Scholar]
- Velázquez-Jiménez, R.; Vargas-Mendoza, D.; Gayosso-de-Lucio, J.A.; González-Montiel, S.; Villagómez-Ibarra, J.R. Three novel epoxy-clerodanes bearing a furan ring from Croton hypoleucus. Phytochem. Lett. 2018, 24, 21–26. [Google Scholar] [CrossRef]
- Pandit, A.; Sachdeva, T.; Bafna, P. Drug-Induced Hepatotoxicity: A Review. J. Appl. Pharm. Sci. 2012, 2, 233–243. [Google Scholar] [CrossRef]
- Robin, S.; Kumar, S.; Rana, A.C.; Sharma, N. Different models of hepatotoxicity and related liver diseases: A review. Int. Res. J. Pharm. 2012, 3, 86–95. [Google Scholar]
- Luo, M.; Dong, L.; Li, J.; Wang, Y.; Shang, B. Protective effects of pentoxifylline on acute liver injury induced by thioacetamide in rats. Int. J. Clin. Exp. Pathol. 2015, 8, 8990–8996. [Google Scholar] [PubMed]
- Zhang, W.; Wang, M.; Xie, H.Y.; Zhou, L.; Meng, X.Q.; Shi, J.; Zheng, S. Role of reactive oxygen species in mediating hepatic ischemia-reperfusion injury and its therapeutic applications in liver transplantation. Transpl. Proc. 2007, 39, 1332–1337. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q. Xenobiotic-activated receptors: From transcription to drug metabolism to disease. Chem. Res. Toxicol. 2008, 21, 1651–1671. [Google Scholar] [CrossRef] [PubMed]
- Holmström, K.M.; Kostov, V.; Dinkova-Kostova, A.T. The multifaceted role of Nrf2 in mitochondrial function. Curr. Opin. Toxicol. 2016, 1, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Prasad, K.N. Simultaneous activation of Nrf2, elevation of antioxidants and reduction in glutamate level: An essential strategy for prevention and improved management of neurodegenerative Diseases. J. Alzheimers Dis. Park. 2016, 6, 6. [Google Scholar] [CrossRef]
- Akhtar, T.; Sheikh, N. An overview of thioacetamide-induced hepatotoxicity. Toxin Rev. 2013, 32, 43–46. [Google Scholar] [CrossRef]
- Ramahia, S.K.; Apte, U.; Mehendale, H.M. Cytochrome P4502E1 induction increases thioacetamide liver injury in diet-restricted rats. Drug Metab. Diapos. 2001, 269, 1088–1095. [Google Scholar]
- Hajovsky, H.; Hu, G.; Koen, Y.; Sarma, D.; Cui, W.; Moore, D.S.; Staudinger, J.L.; Hanzlik, R.P. Metabolism and Toxicity of Thioacetamide and Thioacetamide S -Oxide in Rat Hepatocytes. Chem. Res. Toxicol. 2012, 25, 1955–1963. [Google Scholar] [CrossRef]
- Chilakapati, J.; Shankar, K.; Korrapati, M.C.; Hill, R.A.; Mehendale, H.M. Saturation toxicokineticsof thioacetamide: Role in initiation of liver injury. Drug Metab. Dispos. 2005, 33, 2877–2885. [Google Scholar] [CrossRef]
- Xu, W.H.; Liu, W.Y.; Lang, Q. Chemical constituents from Croton Species and their biological activities. Molecules 2018, 23, 2333. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, M.; Ren, Q.; Li, S.; Xu, J.; Ohizumi, Y.; Xie, C.; jing, D.-Q.; Guo, Y. Two novel clerodane diterpenes with NGF-potentiating activities from the twings of Croton yanhuii. Fitorerapia 2014, 95, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Zhang, H.; Li, S.; Liu, Q.; Song, S. Review: Diterpenoids from croton genus (Euphorbiaceae) and their biological activity. Asian J. Tradit. Med. 2018, 13, 242–262. [Google Scholar]
- Chao, W.-W.; Lin, B.-F. Hepatoprotective diterpenoids isolated from Andrographis paniculate. Chin. Med. J. 2011, 3, 136–143. [Google Scholar] [CrossRef][Green Version]
- Alqasoumi, S.I.; Farraj, A.I.; Abdel-Kader, M.S. Study of the hepatoprotective effect of Janiperus phoenicea constituents. Pak. J. Pharm. Sci. 2013, 26, 999–1008. [Google Scholar] [PubMed]
- Park, E.J.; Zhao, Y.Z.; Young, H.K.; Jung, J.L.; Dong, H.S. Acanthoic acid from Acanthopanax koreanum protects against liver injury induced by tert-butyl hydroperoxide or carbon tetrachloride in vitro and in vivo. Planta Med. 2004, 70, 321–327. [Google Scholar]
- Krishnamurthy, T.P.; Bajaj, J.; Sharma, A.; Maimaran, S.; Bommenahalli, R.P.K.; Pottkad, V. Hepatoprotective activity of terpenoids and terpenoid fractions of Scoparia dulcis L. Orien. Pharm. Exp. Med. 2010, 10, 263–270. [Google Scholar] [CrossRef]
- Tian, J.-L.; Yao, G.-D.; Wang, Y.-X.; Gao, P.-Y.; Wang, D.; Li, L.-Z.; Lin, B.; Huang, X.-X.; Song, S.-J. Cytotoxic clerodane diterpenoids from Croton crassifolius. Bioorg. Med. Chem. Lett. 2017, 27, 1237–1242. [Google Scholar] [CrossRef]
- Kubo, I.; Asaka, Y.; Shibata, K. Insect growth inhibitory nor-diterpenes, cisdehydrocrotonin and trans-dehydrocrotonin, from Croton cajucara. Phytochemistry 1991, 30, 2545–2546. [Google Scholar] [CrossRef]
- Rodrígues, G.; Marcolin, E.; Bona, S.; Porawski, M.; Lehmann, M.; Possa, M.N. Hepatics alterations and genotoxic effects of Croton cajucara Beth (SACACA) in diabetic rats. Arq. Gastroenterol. 2010, 47, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Z.; Zhang, R.; Hou, P.; Bi, K.; Chen, X. Nephrotoxicity evaluation of a new cembrane diterpene from Euphorbiae pekinensis Radix with HEK 293T cells and the toxicokinetics study in rats using a sensitive and reliable UFLC–MS/MS. J. Pharm. Biomed. Anal. 2016, 119, 159–165. [Google Scholar] [CrossRef] [PubMed]
- da Silva Brito, S.S.; Silva, F.; Malheiro, R.; Baptista, P.; Pereira, J.A. Croton argyrophyllus Kunth and Croton heliotropiifolius Kunth: Phytochemical characterization and bioactive properties. Ind. Crops Prod. 2018, 113, 308–315. [Google Scholar] [CrossRef]
- Abdalaziz, M.N.; Ali, A.; Kabbashi, A. In vitro antioxidant activity and phytochemical screening of Croton zambesicus. J. Pharmacogn. Phytochem. 2016, 5, 12–16. [Google Scholar]
- Atoui, A.; Mansouri, A.; Boskou, G.; Kefalas, P. Tea and herbal infusions: Their antioxidant activity and phenolic profile. Food Chem. 2005, 89, 27–36. [Google Scholar] [CrossRef]
- Teixeira, S.; Mendes, A.; Alves, A.; Santos, L. Simultaneous distillation–extraction of high-value volatile compounds from Cistus ladanifer L. Anal. Chim. Acta 2007, 584, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Min, B.; McClung, A.M.; Chen, M.-H. Phytochemicals and Antioxidant Capacities in Rice Brans of Different Color. J. Food Sci. 2011, 76, C117–C126. [Google Scholar] [CrossRef]
- Dudonné, A.; Vitrac, X.; Woillez, M.; Mérillon, J.M. Comparative Study of Antioxidant Properties and Total Phenolic Content of 30 Plant Extracts of Industrial Interest Using DPPH, ABTS, FRAP, SOD, and ORAC Assays. J. Agric. Food Chem. 2009, 57, 1768–1774. [Google Scholar] [CrossRef]
- Furlan, C.M.; Pereira, S.K.; Sedano-Partida, M.D.; Barbosa, D.L.; Santos, D.Y.A.C.; Salatino, M.L.F.; Negri, G.; Berry, P.E.; Van Ee, B.; Salatino, A. Flavonoids and antioxidant potential of nine Argentinian species of roton (Euphorbiaceae). Braz. J. Bot. 2015, 38, 693–702. [Google Scholar] [CrossRef]
- Dos Santos, K.P.; Motta, L.B.; Santos, D.Y.; Salatino, M.L.; Salatino, A.; Ferreira, M.J.; Lago, J.H.; Ruíz, A.L.; Carvaho, J.E.; Furlan, C.M. Antiproliferative activity of flavonoids from Croton sphaerogynus Baill. (Euphorbiaceae). BioMed Res. Int. 2015, 2015, 212809. [Google Scholar] [CrossRef]
- Letha, N.; Ganesan, K.; Nair, P.S.K.; Azalewor, H.G.; Gani, S.B. Evaluation of In Vitro Antioxidant Activity and Phytochemical Screening of Croton macrostachyus Hochst. by using Different Solvent Extracts. Am. J. PharmTech Res. 2016, 6, 73–85. [Google Scholar]
- Lila, M.A.; Raskin, I. Health-related Interactions of Phytochemicals. J. Food Sci. 2005, 70, R20–R27. [Google Scholar] [CrossRef]
- Marchyshak, T.; Yakovenko, T.; Shmarakov, I.; Tkachuk, Z. The potential protective effect of oligoribonucleotides-d-mannitol complexes against thioacetamide-induced hepatotoxicity in mice. Pharmaceuticals 2018, 11, 77. [Google Scholar] [CrossRef] [PubMed]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef] [PubMed]
- Shon, M.-Y.; Lee, J.; Choi, J.-H.; Choi, S.-Y.; Nam, S.-H.; Seo, K.I.; Sang-Won, L.; Sung, N.J.; Park, S.K. Antioxidant and free radical scavenging activity of methanol extract of chungkukjang. J. Food Compos. Anal. 2007, 20, 113–118. [Google Scholar] [CrossRef]
- Gad, S.C. Single-Dose (Acute) and Pilot (DRF) Toxicity Testing in Drug Safety Evaluation. In Drug Safety Evaluation; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011; pp. 185–233. [Google Scholar] [CrossRef]
- Bulus, T.; Atawodi, S.E.; Mamman, M. Acute toxicity effect on the aqueous extract of Termelia avicennioides on white albino rats. Sci. World J. 2011, 6, 1–4. [Google Scholar]
- Asare, G.A.; Sittie, A.; Bugyei, K.; Gyan, B.A.; Adjei, S.; Addo, P.; Wiredu, E.K.; Nyarko, A.K.; Out-Nyarko, L.S.; Adjei, D.N. Acute toxicity studies of Croton membranaceus root extract. J. Ethnopharmacol. 2011, 135, 398–934. [Google Scholar] [CrossRef]
- Sridhar, N. Comparative anti-inflammatory and anti-oxidant evaluation of Jatropha gossypifolia and Croton bonplandianm. Int. J. Res. Pharm. Sci. 2013, 4, 16–27. [Google Scholar]
- Meireles, D.R.P.; Fernandes, H.M.B.; Rolim, T.L.; Batista, T.M.; Mangueira, V.M.; de Sousa, T.K.G.; Pita, J.C.L.R.; Xavier, A.L.; Beltrão, D.M.; Tavares, J.F.; et al. Toxicity and antitumor efficacy of Croton polyandrus oil against Ehrlich ascites carcinoma cells. Rev. Bras. Farmacogn. 2016, 26, 751–758. [Google Scholar] [CrossRef]
- Lorke, D. A new approach to practical acute toxicity testing. Arch. Toxicol. 1983, 54, 275–287. [Google Scholar] [CrossRef]
- Amin, K.A.; Mohamed, B.M.; El-wakil, M.A.M.; Ibrahem, S.O. Impact of Breast Cancer and Combination Chemotherapy on Oxidative Stress, Hepatic and Cardiac Markers. J. Breast Cancer. 2012, 15, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Bhat, T.K.; Sharma, O.P. Clinical Biochemistry of Hepatotoxicity. J. Clin. Toxicol. 2011, S4, 1–19. [Google Scholar] [CrossRef]
- Ozer, J.; Ratner, M.; Shaw, M.; Bailer, W.; Schomaker, S. The current state of serum biomarkers of hepatotoxicity. Toxicology 2008, 245, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Bautista, M.; Velazquez-González, C.; De la O Arciniega, M.; Morales-González, J.; Benedí, J.; Gayosso-De-Lucio, J. Chemical composition and hepatotoxic effect of Geranium schiedeanum in a thioacetamide-induced liver injury model. Pharmacogn. Mag. 2014, 10, 574. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Schnackenberg, L.K.; Shi, Q.; Salminen, W.F. Hepatic toxicity biomarkers. In Biomarkers; Gupta, R.C., Ed.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 241–260. [Google Scholar]
- Aydin, A.F.; Kusku-Kiraz, S.Z.; Dogru-Abbasoglu, M.; Gulluoglu, M.U.; Kocak-Toker, N. Effect of carnosine against thioacetamide-induced liver cirrhosis in rat. Peptides 2010, 31, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Atef, M.A. Hepatoprotective influence of vitamin C on thioacetamide-induced liver cirrhosis in wistar male rats. J. Toxicol. Pharmacol. 2011, 6, 218–233. [Google Scholar] [CrossRef][Green Version]
- Miguel, F.M.; Schemitt, E.G.; Colares, J.R.; Hartmann, R.M.; Morgan-Martins, M.I.; Marroni, N.P. Actio of vitamin E on experimental severe acute liver failure. Arq. Gastroenterol. 2017, 54, 123–129. [Google Scholar] [CrossRef]
- Braunwld, E.; Ghany, M.; Hoofnagle, J.; Berk, P.; Wolkoff, A.; Dienstag, J. Harrison. Principios de Medicina Interna, 17th ed.; Editorial Mc Graw-Hill Interamericana: México City, Mexico, 2009. [Google Scholar]
- Everhart, J.E.; Wright, E.C. Association of γ-glutamyl transferase (GGT) activity with treatment and clinical outcomes in chronic hepatitis C (HCV). Hepatology 2013, 57, 1725–1733. [Google Scholar] [CrossRef]
- Lee, D.-H.; Jacobs, D.R. Association between serum gamma-glutamyltransferase and C-reactive protein. Atherosclerosis 2005, 178, 327–330. [Google Scholar] [CrossRef]
- Bigoniya, P.; Singh, C.S.; Shukla, A. A comprehensive review of different liver toxicants used in experimental pharmacology. Int. J. Pharm. Sci. Drug Res. 2009, 1, 124–135. [Google Scholar]
- Zimmerman, H.J. Drug-induced liver disease. Clin. Liver Dis. 2000, 4, 79–96. [Google Scholar] [CrossRef]
- Khalaf, N.A.; Shakya, A.K.; Al-Othman, A.; El-Agbar, Z.; Farah, H. Antioxidant Activity of Some Common Plants. Turk. J. Biol. 2008, 32, 51–55. [Google Scholar]
- Pandey, K.B.; Rizvi, S.I. Markers of Oxidative Stress in Erythrocytes and Plasma During Aging in Humans. Oxidative Med. Cell. Longev. 2010, 3, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Gao, Y.; Chu, S.; Zhang, Z.; Xia, C.; Mou, Z.; Song, X.-Y.; He, W.-B.; Guo, X.-F.; Chen, N.-H. Nrf2 pathway activation contributes to anti-fibrosis effects of ginsenoside Rg1 in a rat model of alcohol- and CCl4-induced hepatic fibrosis. Acta Pharmacol. Sin. 2014, 35, 1031–1044. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.-N.; Maitra, A.; Lee, K.-F.; Jan, Y.-Y.; Chen, M.-F. Thioacetamide-induced intestinal-type cholangiocarcinoma in rat: An animal model recapitulating the multi-stage progression of human cholangiocarcinoma. Carcinogenesis 2003, 25, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, K.; Motohashi, H.; Yamamoto, M. Molecular mechanisms of the Keap1-Nrf2 pathway in stress response and cancer evolution: Molecular mechanisms of the Keap1-Nrf2 pathway. Genes Cells 2011, 16, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Rada, P.; Rojo, A.I.; Chowdhry, S.; McMahon, M.; Hayer, J.D.; Cuadrado, A. SCF/b-TrCP Promotes Glycogen Synthase kinase 3-dependent degradation of the Nrf2 transcription factor in a keap1-independent manner. Mol. Cell. Biol. 2011, 31, 1121–1133. [Google Scholar] [CrossRef]
- Bataille, A.M.; Manautou, J.E. Nrf2 a potential target to new therapeutics in liver disease. Clin. Pharmacol. Ther. 2012, 92, 340–348. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT—Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- SAGARPA. Norma Oficial Mexicana NOM-062-ZOO-1999. Especificaciones técnicas para la producción, cuidado y manejo de animales de laboratorio. Diario Oficial de la Federación 2001, 107–165. [Google Scholar]
- Murray, R. Alanine aminotransferase. In Clinical Chemistry: Theory, Analysis, and Correlation, 2nd ed.; CV Mosby: St. Louis, MO, USA, 1989; pp. 898–989. [Google Scholar]
- Rej, R.; Horder, M. Aspartate aminotransferase. L-aspartate: 2-oxoglutarate aminotranferase, EC 2.6.2.1. Routine, U.V. method. In Methods of Enzymatic Analysis; Verlag-CHemie: Weinheim, Germany, 1987; pp. 416–424. [Google Scholar]
- Bessey, O.A.; Lowry, O.H.; Brock, M.J. A method for the rapid determination of alkaline phosphatase with five cubic millimeters of serum. J. Biol. Chem. 1946, 164, 321–329. [Google Scholar] [PubMed]
- Theodorsen, L.; Strømme, J. Gamma-glutamyl-3-c arboxy4-nitroanilide: The substrate of choice for routine determinatinations of y-glutamyl-transferase activity in serum? Clin. Chim. Acta 1976, 72, 205–210. [Google Scholar] [CrossRef]
- Vanderlinde, R.E. Measurement of total lactate dehydrogenase activity. Ann. Clin. Lab. Sci. 1985, 15, 13–31. [Google Scholar] [PubMed]
- Martinek, R.G. Improved micro-method for determination of serum bilirubin. Clin. Chim. Acta 1966, 13, 161–170. [Google Scholar] [CrossRef]
- Neha, J.; Mishra, R.N. Antioxidant activity of Trikatu megaExt. Int. J. Res. Pharm. Biosci. 2011, 2, 624–628. [Google Scholar]
- Aebi, H. Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Livak, K.J.; Schimittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EC | Intragastric Doses (mg/kg) | ||
|---|---|---|---|
| Phase I | 10 | 100 | 1000 |
| Mortality | 0/3 | 0/3 | 0/3 |
| Phase II | 1600 | 2900 | 5000 |
| Mortality | 0/3 | 0/3 | 0/3 |
| LD50 | >5000 | ||
| Mn-SOD | sense: 50-ACTGAAGTTCAATGGCGGG-30 and antisense: 50-TCCAGCAACTCTCCTTTGGG-30 |
| CuZn-SOD | sense: 50-CTTCGAGCAGAAGGCAAGCG-30and antisense: 50-GACATGGAACCCATGCTCGC-30 |
| Cat | sense: 50-ATCAGGGATGCCATGTTGTT-30 and antisense: 50-GGGTCCTTCAGGTGAGTTTG-30 |
| Nrf2 | sense: 50-TTGTAGATGACCATGAGTCGC-30 and antisense: 50-GAGCTATCGAGTGACTGAGCC-30 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urrutia-Hernández, T.A.; Santos-López, J.A.; Benedí, J.; Sánchez-Muniz, F.J.; Velázquez-González, C.; De la O-Arciniega, M.; Jaramillo-Morales, O.A.; Bautista, M. Antioxidant and Hepatoprotective Effects of Croton hypoleucus Extract in an Induced-Necrosis Model in Rats. Molecules 2019, 24, 2533. https://doi.org/10.3390/molecules24142533
Urrutia-Hernández TA, Santos-López JA, Benedí J, Sánchez-Muniz FJ, Velázquez-González C, De la O-Arciniega M, Jaramillo-Morales OA, Bautista M. Antioxidant and Hepatoprotective Effects of Croton hypoleucus Extract in an Induced-Necrosis Model in Rats. Molecules. 2019; 24(14):2533. https://doi.org/10.3390/molecules24142533
Chicago/Turabian StyleUrrutia-Hernández, Thania Alejandra, Jorge Arturo Santos-López, Juana Benedí, Francisco Jose Sánchez-Muniz, Claudia Velázquez-González, Minarda De la O-Arciniega, Osmar Antonio Jaramillo-Morales, and Mirandeli Bautista. 2019. "Antioxidant and Hepatoprotective Effects of Croton hypoleucus Extract in an Induced-Necrosis Model in Rats" Molecules 24, no. 14: 2533. https://doi.org/10.3390/molecules24142533
APA StyleUrrutia-Hernández, T. A., Santos-López, J. A., Benedí, J., Sánchez-Muniz, F. J., Velázquez-González, C., De la O-Arciniega, M., Jaramillo-Morales, O. A., & Bautista, M. (2019). Antioxidant and Hepatoprotective Effects of Croton hypoleucus Extract in an Induced-Necrosis Model in Rats. Molecules, 24(14), 2533. https://doi.org/10.3390/molecules24142533