
Chemical Characterization of an Encapsulated Red Wine Powder and Its Effects on Neuronal Cells



Abstract
1. Introduction
2. Results
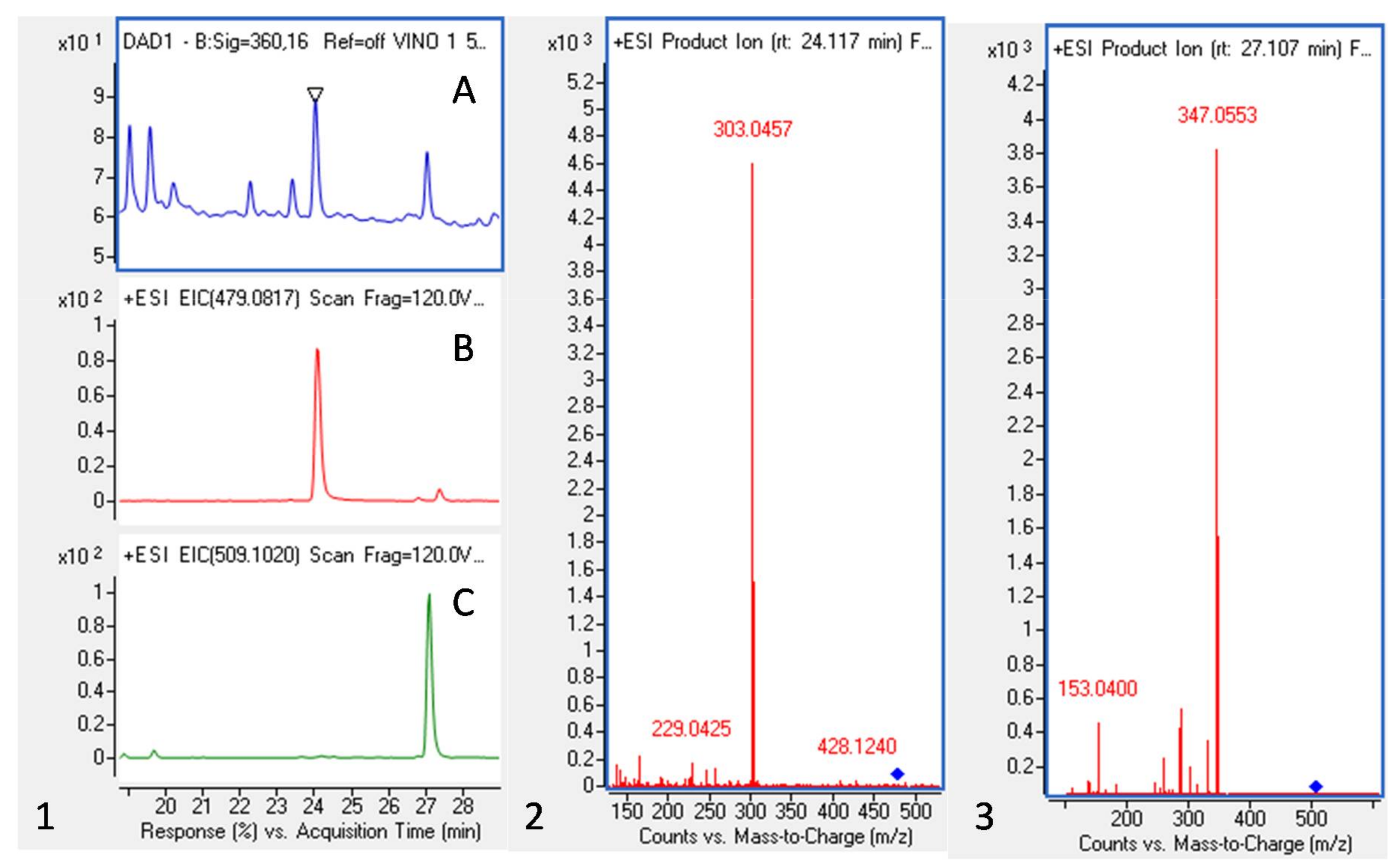
2.1. Red Wine Powder (WP) Polyphenolic Composition
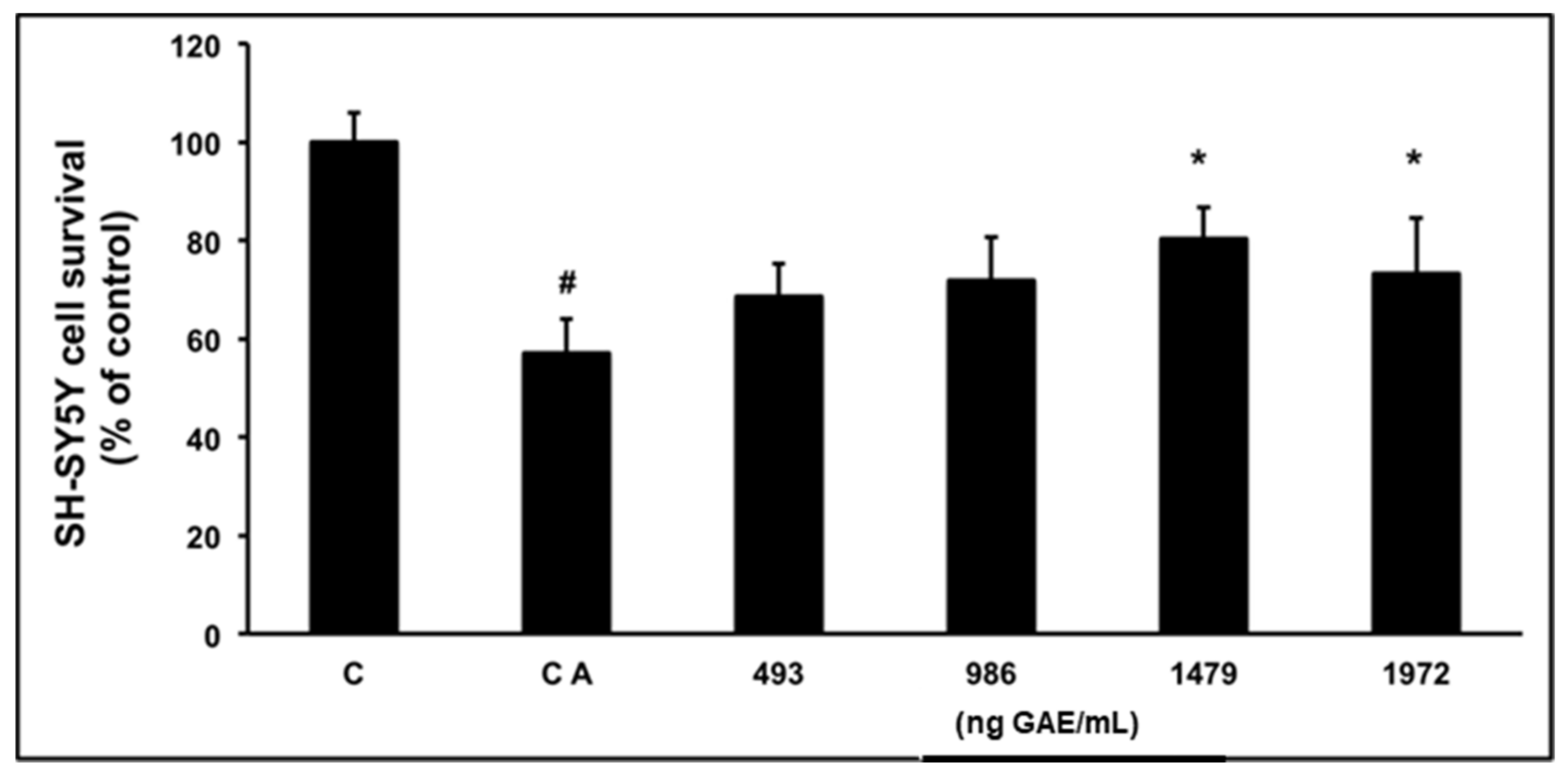
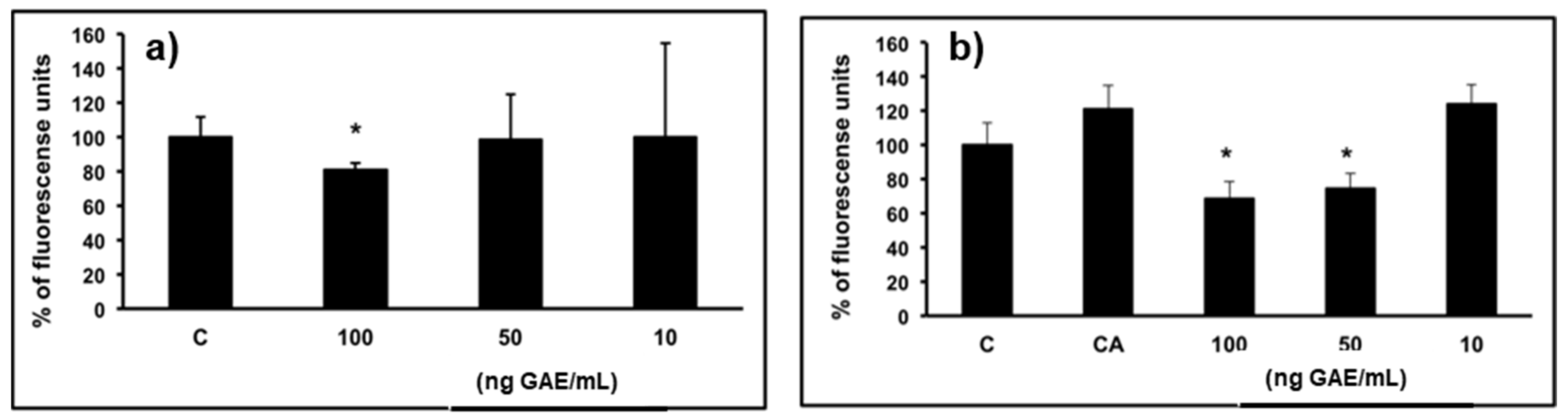
2.2. Cell Viability
2.3. Effect of WP on ROS Production
3. Discussion
3.1. Red Wine Powder (WP) Polyphenolic Composition
3.2. Cell Viability
3.3. Effect of WP on Reactive Oxygen Species (ROS) Production
4. Materials and Methods
4.1. Encapsulation Procedure
4.2. HPLC-QTOF Analysis
4.3. Cell Culture and Viability
4.4. Evaluation of Reactive Oxygen Species (ROS) Generation
4.5. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Stevenson, D.E.; Hurst, R.D. Polyphenolic phytochemicals—Just antioxidants or much more? Cell. Mol. Life Sci. 2007, 64, 2900–2916. [Google Scholar] [CrossRef] [PubMed]
- Cendrowski, A.; Ścibisz, I.; Kieliszek, M.; Kolniak-Ostek, J.; Mitek, M. UPLC-PDA-Q/TOF-MS profile of polyphenolic compounds of liqueurs from Rose petals (Rosa rugosa). Molecules 2017, 22, 1832. [Google Scholar] [CrossRef] [PubMed]
- Cendrowski, A.; Ścibisz, I.; Mitek, M.; Kieliszek, M.; Kolniak-Ostek, J. Profile of the Phenolic Compounds of Rosa rugosa Petals. J. Food Qual. 2017. [Google Scholar] [CrossRef]
- Garcia-Alonso, M.; Rimbach, G.; Rivas-Gonzalo, J.C.; de Pascual-Teresa, S. Antioxidant and cellular activities of anthocyanins and their corresponding vitisins A—Studies in platelets, monocytes, and human endothelial cells. J. Agric. Food. Chem. 2004, 52, 3378–3384. [Google Scholar] [CrossRef] [PubMed]
- Sun, A.Y.; Simonyi, A.Ñ.; Sun, G.Y. The ‘French Paradox’ and beyond: Neuroprotective effects of polyphenols. Free Radic. Biol. Med. 2002, 32, 314–318. [Google Scholar] [CrossRef]
- Corder, R.; Mullen, W.; Khan, N.Q.; Marks, S.C.; Wood, E.G.; Carrier, M.J.; Crozier, A. Oenology: Red wine procyanidins and vascular health. Nature 2006, 444, 566. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, A.; Lisa-Santamaría, P.; García-Marino, M.; Escribano-Bailón, M.T.; Rivas-Gonzalo, J.C.; Revuelta, J.L. The biological activity of the wine anthocyanins delphinidin and petunidin is mediated through Msn2 and Msn4 in Saccharomyces cerevisiae. FEMS Yeast Res. 2010, 10, 858–869. [Google Scholar] [CrossRef] [PubMed]
- German, J.B.; Walzem, R.L. The health benefits of wine. Annu. Rev. Nutr. 2000, 20, 561–593. [Google Scholar] [CrossRef] [PubMed]
- Renaud, S.; de Lorgeril, M. Wine, alcohol, platelets, and the French paradox for coronary heart disease. Lancet 1992, 339, 1523–1526. [Google Scholar] [CrossRef]
- Nikfardjam, M.S.P.; Márk, L.; Avar, P.; Figler, M.; Ohmacht, R. Polyphenols, anthocyanins, and trans-resveratrol in red wines from the Hungarian Villány region. Food Chem. 2006, 98, 453–462. [Google Scholar] [CrossRef]
- Soulat, T.; Philippe, C.; Sollier, C.B.D.; Brezillon, C.; Berge, N.; Teissedre, P.L.; Drouet, L. Wine constituents inhibit thrombosis but not atherogenesis in C57BL/6 apolipoprotein E-deficient mice. Br. J. Nutr. 2006, 96, 290. [Google Scholar] [CrossRef] [PubMed]
- Gorelik, S.; Ligumsky, M.; Kohen, R.; Kanner, J. A novel function of red wine polyphenols in humans: prevention of absorption of cytotoxic lipid peroxidation products. FASEB J. 2008, 22, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Carollo, C.; Presti, R.; Caimi, G. Wine, diet, and arterial hypertension. Angiology 2007, 58, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Karatzi, K.N.; Papamichael, C.M.; Karatzis, E.N.; Papaioannou, T.G.; Aznaouridis, K.A.; Katsichti, P.P.; Mavrikakis, M.E. Red wine acutely induces favorable effects on wave reflections and central pressures in coronary artery disease patients. Am. J. Hypertens. 2005, 18, 1161–1167. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Jiménez, J.; Serrano, J.; Tabernero, M.; Arranz, S.; Díaz-Rubio, M.E.; García-Diaz, L.; Saura-Calixto, F. Effects of grape antioxidant dietary fiber in cardiovascular disease risk factors. Nutrition 2008, 24, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Greenrod, W.; Stockley, C.S.; Burcham, P.; Abbey, M.; Fenech, M. Moderate acute intake of de-alcoholised red wine, but not alcohol, is protective against radiation-induced DNA damage ex vivo-Results of a comparative in vivo intervention study in younger men. Mutat. Res. 2005, 591, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Lecour, S.; Blackhurst, D.; Marais, D.; Opie, L. Lowering the degree of alcohol in red wine does not alter its cardioprotective effect. J. Mol. Cell. Cardiol. 2006, 40, 997–998. [Google Scholar] [CrossRef]
- Sanchez, V.; Baeza, R.; Galmarini, M.V.; Zamora, M.C.; Chirife, J. Freeze-Drying Encapsulation of Red Wine Polyphenols in an Amorphous Matrix of Maltodextrin. Food Bioprocess Technol. 2013, 6, 1350–1354. [Google Scholar] [CrossRef]
- Rocha-Parra, D.F.; Lanari, M.C.; Zamora, M.C.; Chirife, J. Influence of storage conditions on phenolic compounds stability, antioxidant capacity and colour of freeze-dried encapsulated red wine. LWT Food Sci. Technol. 2006, 70, 162–170. [Google Scholar] [CrossRef]
- Galmarini, M.V.; Maury, C.; Mehinagic, E.; Sanchez, V.; Baeza, R.I.; Mignot, S.; Zamora, C.; Chirife, J. Stability of Individual Phenolic Compounds and Antioxidant Activity During Storage of a Red Wine Powder. Food Bioprocess Technol. 2013, 6, 3585–3595. [Google Scholar] [CrossRef]
- Nicolini, G.; Rigolio, R.; Miloso, M.; Bertelli, A.A.E.; Tredici, G. Anti-apoptotic effect of trans-resveratrol on paclitaxel-induced apoptosis in the human neuroblastoma SH-SY5Y cell line. Neurosci. Lett. 2001, 302, 41–44. [Google Scholar] [CrossRef]
- Cavallini, G.; Straniero, S.; Donati, A.; Bergamini, E. Resveratrol requires red wine polyphenols for optimum antioxidant activity. J. Nutr. Health Aging 2016, 20, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A., Jr. Colorimetry of total phenolics with phosphomolybdic–phosphotungtic acid reagent. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Cantos, E.; García-Viguera, C.; de Pascual-Teresa, S.; Tomás-Barberán, F.A. Effect of postharvest ultraviolet irradiation on resveratrol and other phenolics of cv. Napoleon table grapes. J. Agric. Food. Chem. 2000, 10, 4606–4612. [Google Scholar] [CrossRef]
- Bimpilas, A.; Tsimogiannis, D.; Balta-Brouma, K.; Lymperopoulou, T.; Oreopoulou, V. Evolution of phenolic compounds and metal content of wine during alcoholic fermentation and storage. Food Chem. 2015, 178, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Dopico-García, M.S.; Fique, A.; Guerra, L.; Afonso, J.M.; Pereira, O.; Valentão, P.; Seabra, R.M. Principal components of phenolics to characterize red Vinho Verde grapes: Anthocyanins or non-coloured compounds? Talanta 2008, 75, 1190–1202. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Race, E.J.; Shrikhande, A.J. Anthocyanin transformation in Cabernet Sauvignon wine during aging. J. Agric. Food Chem. 2003, 51, 7989–7994. [Google Scholar] [CrossRef] [PubMed]
- De Pascual-Teresa, S.; Rivas-Gonzalo, J.C.; Santos-Buelga, C. Prodelphinidins and related flavanols in wine. International. J. Food Sci. Technol. 2000, 35, 33–40. [Google Scholar] [CrossRef]
- He, F.; Liang, N.N.; Mu, L.; Pan, Q.H.; Wang, J.; Reeves, M.J.; Duan, C.Q. Anthocyanins and their variation in red wines I. Monomeric anthocyanins and their color expression. Molecules 2012, 17, 1571–1601. [Google Scholar] [CrossRef] [PubMed]
- Li, S.Y.; He, F.; Zhu, B.Q.; Wang, J.; Duan, C.Q. Comparison of Phenolic and Chromatic Characteristics of Dry Red Wines Made from Native Chinese Grape Species and Vitis Vinifera. Int. J. Food Prop. 2017, 20, 2134–2146. [Google Scholar] [CrossRef]
- Shi, P.B.; Yue, T.X.; Ai, L.L.; Cheng, Y.F.; Meng, J.F.; Li, M.H.; Zhang, Z.W. Phenolic compound profiles in grape skins of Cabernet Sauvignon, Merlot, Syrah and Marselan cultivated in the Shacheng Area (China). S. Afr. J. Enol. Vitic. 2016, 37, 132–138. [Google Scholar]
- Shay, J.; Elbaz, H.A.; Lee, I.; Zielske, S.P.; Malek, M.H.; Hüttemann, M. Molecular Mechanisms and Therapeutic Effects of (−)-Epicatechin and Other Polyphenols in Cancer, Inflammation, Diabetes, and Neurodegeneration. Oxid. Med. Cell. Longev. 2015. [Google Scholar] [CrossRef] [PubMed]
- Miloso, M.; Bertelli, A.A.E.; Nicolini, G.; Tredici, G. Resveratrol-induced activation of the mitogen-activated protein kinases, ERK1 and ERK2, in human neuroblastoma SH-SY5Y cells. Neurosci. Lett. 1999, 264, 141–144. [Google Scholar] [CrossRef]
- Tredici, G.; Miloso, M.; Nicolini, G.; Galbiati, S.; Cavaletti, G.; Bertelli, A. Resveratrol, map kinases and neuronal cells: Might wine be a neuroprotectant? Drugs Exp. Clin. Res. 1999, 25, 99–103. [Google Scholar] [PubMed]
- Savaskan, E.; Olivieri, G.; Meier, F.; Seifritz, E.; Wirz-Justice, A.; Müller-Spahn, F. Red wine ingredient resveratrol protects from β-amyloid neurotoxicity. Gerontology 2003, 49, 380–383. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.K.; Kang, S.J.; Poncz, M.; Song, K.-J.; Park, K.S. Resveratrol protects SH-SY5Y neuroblastoma cells from apoptosis induced by dopamine. Exp. Mol. Med. 2007, 39, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Soundararajan, R.; Wishart, A.D.; Rupasinghe, H.P.V.; Arcellana-Panlilio, M.; Nelson, C.M.; Mayne, M.; Robertson, G.S. Quercetin 3-glucoside protects neuroblastoma (SH-SY5Y) cells in vitro against oxidative damage by inducing sterol regulatory element-binding protein-2-mediated cholesterol biosynthesis. J. Biol. Chem. 2008, 283, 2231–2245. [Google Scholar] [CrossRef] [PubMed]
- Bandaruk, Y.; Mukai, R.; Terao, J. Cellular uptake of quercetin and luteolin and their effects on monoamine oxidase-A in human neuroblastoma SH-SY5Y cells. Toxicol. Rep. 2014, 1, 639–649. [Google Scholar] [CrossRef] [PubMed]
- González-Sarrías, A.; Núñez-Sánchez, M.Á.; Tomás-Barberán, F.A.; Espín, J.C. Neuroprotective effects of bioavailable polyphenol-derived metabolites against oxidative stress-induced cytotoxicity in human neuroblastoma SH-SY5Y cells. J. Agric. Food Chem. 2016, 65, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Esteban-Fernández, A.; Rendeiro, C.; Spencer, J.P.E.; Gigorro del Coso, D.; González del Llano, M.D.; Bartolomé, B.; Moreno-Arribas, M.V. Neuroprotective Effects of Selected Microbial-Derived Phenolic Metabolites and Aroma Compounds from Wine in Human SH-SY5Y Neuroblastoma Cells and Their Putative Mechanisms of Action. Front. Nutr. 2017, 4, 3. [Google Scholar] [CrossRef] [PubMed]
- Pignatelli, P.; Ghiselli, A.; Buchetti, B.; Carnevale, R.; Natella, F.; Germanò, G.; Violi, F. Polyphenols synergistically inhibit oxidative stress in subjects given red and white wine. Atherosclerosis 2006, 188, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Guilford, J.M.; Pezzuto, J.M. Wine and health: A review. Am. J. Enol. Vitic. 2011, 62, 471–486. [Google Scholar] [CrossRef]
- Giovinazzo, G.; Grieco, F. Functional Properties of Grape and Wine Polyphenols. Plant Foods Hum. Nutr. 2015, 70, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Tseng, W.T.; Hsu, Y.W.; Pan, T.M. The ameliorative effect of Monascus purpureus NTU 568-fermented rice extracts on 6-hydroxydopamine-induced neurotoxicity in SH-SY5Y cells and the rat model of Parkinson’s disease. Food Funct. 2016, 7, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Rabelo, T.K.; Zeidán-Chuliá, F.; Vasques, L.M.; dos Santos, J.P.; da Rocha, R.F.; Pasquali, M.A.; Rybarczyk-Filho, J.L.; Araújo, A.A.; Moreira, J.C.; Gelain, D.P. Redox characterization of usnic acid and its cytotoxic effect on human neuron-like cells (SH-SY5Y). Toxicol. In Vitro 2012, 26, 304–314. [Google Scholar] [CrossRef] [PubMed]
- Basli, A.; Soulet, S.; Chaher, N.; Mérillon, J.-M.; Chibane, M.; Monti, J.-P.; Richard, T. Wine Polyphenols: Potential Agents in Neuroprotection. Oxid. Med. Cell. Longev. 2012, 805762. [Google Scholar] [CrossRef] [PubMed]
- Caruana, M.; Cauchi, R.; Vassallo, N. Putative Role of Red Wine Polyphenols against Brain Pathology in Alzheimer’s and Parkinson’s Disease. Front. Nutr. 2016, 3, 31. [Google Scholar] [CrossRef] [PubMed]
- Silván, J.M.; Reguero, M.; de Pascual-Teresa, S. A protective effect of anthocyanins and xanthophylls on UVB-induced damage in retinal pigment epithelial cells. Food Funct. 2016, 7, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polyphenol | Rt (min) | M+ | M− | Fragment | Abs Max (nm) | (mg/100 g WP) |
|---|---|---|---|---|---|---|
| Anthocyanins | 249.92 | |||||
| Delphinidin-3-O-glucoside | 7.8 | 465.10 | 303.04 | 520 | 5.90 | |
| Cyanidin-3-O-glucoside | 9.9 | 449.10 | 287.05 | 520 | 0.73 | |
| Petunidin-3-O-glucoside | 10.5 | 479.11 | 317.06 | 520 | 15.60 | |
| Peonidin-3-O-glucoside | 12.7 | 463.12 | 301.06 | 520 | 6.06 | |
| Malvidin-3-O-glucoside | 13.0 | 493.10 | 331.08 | 520 | 112.45 | |
| Delphindin-3-O-(6″-ac-glu) | 15.0 | 507.08 | 303.02 | 520 | 4.33 | |
| Cyanidin- 3-O-(6″-ac-glu) | 17.6 | 491.08 | 287.03 | 520 | 0.64 | |
| Petunidin- 3-O-(6″-ac-glu) | 18.1 | 521.09 | 317.04 | 520 | 7.74 | |
| Peonidin- 3-O-(6″-ac-glu) | 20.4 | 505.10 | 301.04 | 520 | 3.65 | |
| Malvidin 3-O-(6″-ac-glu) | 20.5 | 535.11 | 331.06 | 520 | 72.87 | |
| Delphinidin 3-O-(6″-cou-glu) | 22.0 | 611.13 | 303.04 | 520 | 0.24 | |
| Cyanidin 3-O-(6″-cou-glu) | 24.0 | 595.14 | 287.05 | 520 | 0.15 | |
| Petunidin 3-O-(6″-cou-glu) | 24.2 | 625.15 | 317.06 | 520 | 1.24 | |
| Peonidin 3-O-(6″-cou-glu) | 26.6 | 609.13 | 301.05 | 520 | 1.48 | |
| Malvidin 3-O-(6″-cou-glu) | 26.6 | 639.14 | 331.06 | 520 | 16.84 | |
| Flavonols | 8.34 | |||||
| Quercetin-3-O-glucoside | 23.1 | 465.10 | 303.04 | 360 | 0.27 | |
| Quercetin-3-O-glucuronide | 24.1 | 479.08 | 303.04 | 360 | 4.19 | |
| Syringenin-3-O-glucoside | 27.0 | 509.10 | 347.05 | 360 | 3.70 | |
| Syringenin- 3-O-(6″-ac-glu) | 32.3 | 551.10 | 347.05 | 360 | 0.18 | |
| Flavanols | 24.73 | |||||
| Gallocatechin | 7.4 | 307.07 | 139.03 | 280 | 0.95 | |
| Epigallocatechin | 10.3 | 307.07 | 139.03 | 280 | 0.18 | |
| Catechin | 12.6 | 291.07 | 139.03; 123.03 | 280 | 5.06 | |
| Epicatechin | 15.9 | 291.06 | 139.03; 123.03 | 280 | 5.75 | |
| Procyanidin B1 | 9.8 | 579.14 | 127.03; 409.08 | 280 | 5.18 | |
| Procyanidin B3 | 11.5 | 579.14 | 127.03; 287.04 | 280 | 1.56 | |
| Procyanidin B4 | 13.8 | 579.14 | 127.03 | 280 | 0.70 | |
| Procyanidin B2 | 14.1 | 579.14 | 127.03; 409.08 | 280 | 3.76 | |
| Procyanidin B7 | 18.4 | 579.11 | 127.03; 287.04 | 280 | 0.43 | |
| Procyanidin trimer EEC | 12.3 | 867.21 | 289.07; 579.13 | 280 | 0.66 | |
| Procyanidin trimer EEE | 17.1 | 867.16 | 291.05; 579.10 | 280 | 0.50 | |
| Phenolic acids | 29.10 | |||||
| Gallic acid | 5.3 | 169.01 | 125.01 | 280 | 16.75 | |
| Caffeic acid | 15.6 | 179.03 | 135.04 | 280 | 1.97 | |
| Syringic acid | 21.3 | 197.04 | 124.02; 169.02 | 280 | 10.37 | |
| Stilbenoids | 0.15 | |||||
| Resveratrol | 39.3 | 227.11 | 138.95; 185.05 | 280 | 0.07 | |
| Piceid (Res 3-O-glu) | 28.5 | 389.22 | 227.06 | 280 | 0.08 | |
| Total Polyphenols by HPLC | 312.23 |
| (−)-epicatechin (μM) | SH-SY5Y Cell Survival * (% of Control) |
|---|---|
| Activated Control | 58.0 ± 5.5 |
| 10 | 81.1 ± 6.5 * |
| 25 | 96.5 ± 4.8 * |
| 50 | 99.4 ± 9.5 * |
| 100 | 89.2 ± 10.9 * |
| 200 | 102.8 ± 6.3 * |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rocha-Parra, D.; Chirife, J.; Zamora, C.; De Pascual-Teresa, S. Chemical Characterization of an Encapsulated Red Wine Powder and Its Effects on Neuronal Cells. Molecules 2018, 23, 842. https://doi.org/10.3390/molecules23040842
Rocha-Parra D, Chirife J, Zamora C, De Pascual-Teresa S. Chemical Characterization of an Encapsulated Red Wine Powder and Its Effects on Neuronal Cells. Molecules. 2018; 23(4):842. https://doi.org/10.3390/molecules23040842
Chicago/Turabian StyleRocha-Parra, Diego, Jorge Chirife, Clara Zamora, and Sonia De Pascual-Teresa. 2018. "Chemical Characterization of an Encapsulated Red Wine Powder and Its Effects on Neuronal Cells" Molecules 23, no. 4: 842. https://doi.org/10.3390/molecules23040842
APA StyleRocha-Parra, D., Chirife, J., Zamora, C., & De Pascual-Teresa, S. (2018). Chemical Characterization of an Encapsulated Red Wine Powder and Its Effects on Neuronal Cells. Molecules, 23(4), 842. https://doi.org/10.3390/molecules23040842