A New Monoterpene from the Leaves of a Radiation Mutant Cultivar of Perilla frutescens var. crispa with Inhibitory Activity on LPS-Induced NO Production



Abstract
:1. Introduction
2. Results
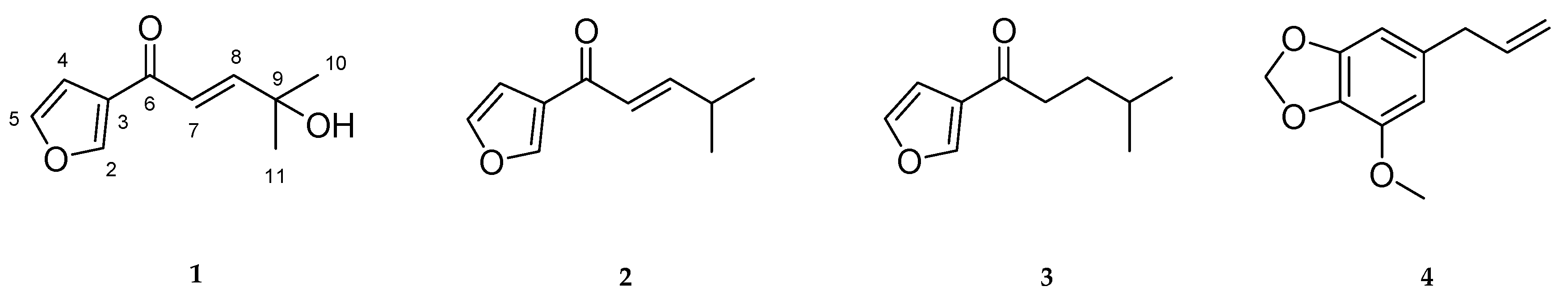
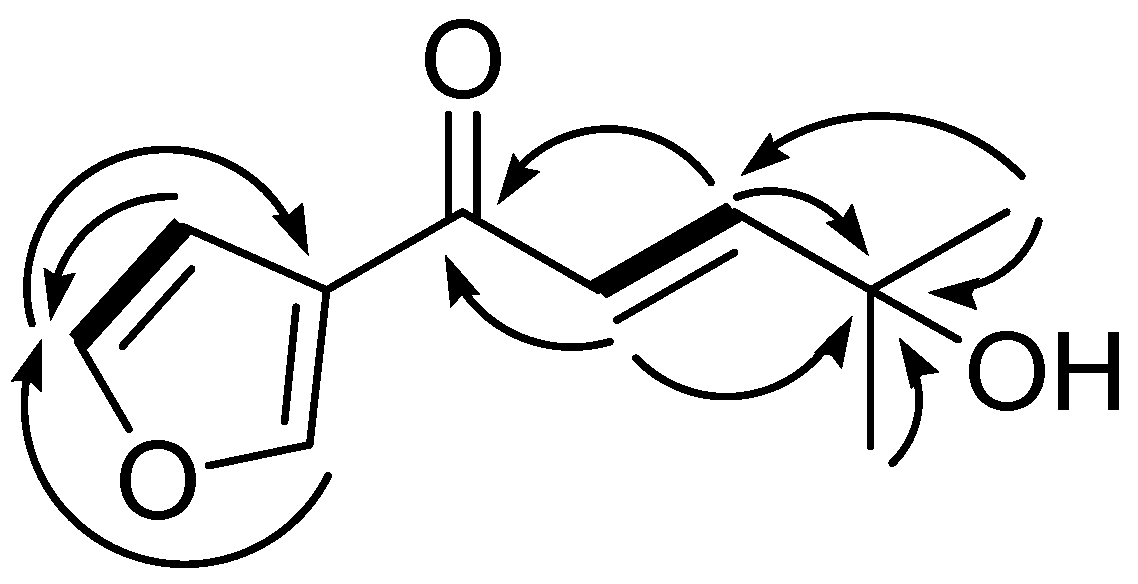
2.1. Structure Elucidation of Compound 1
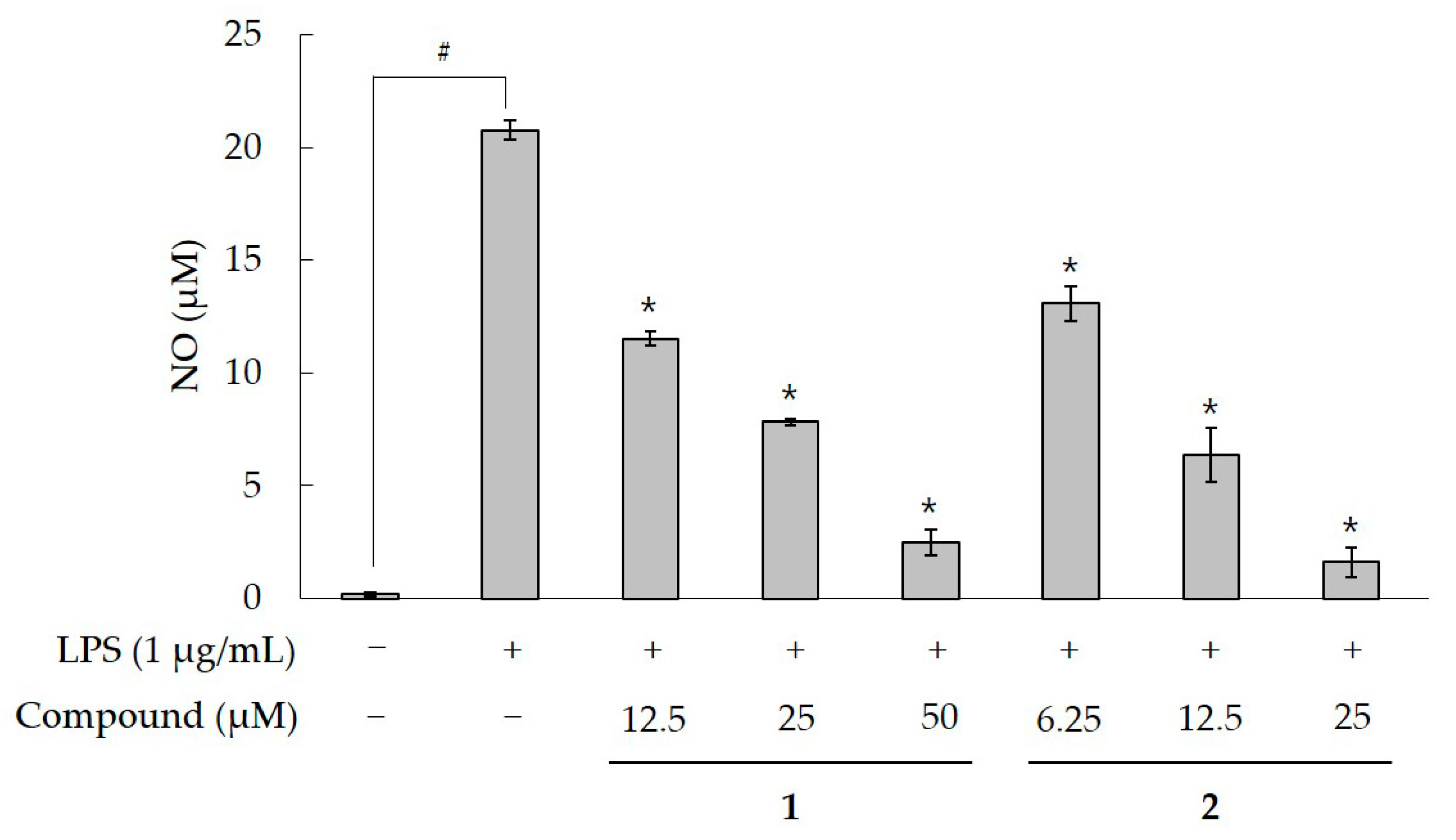
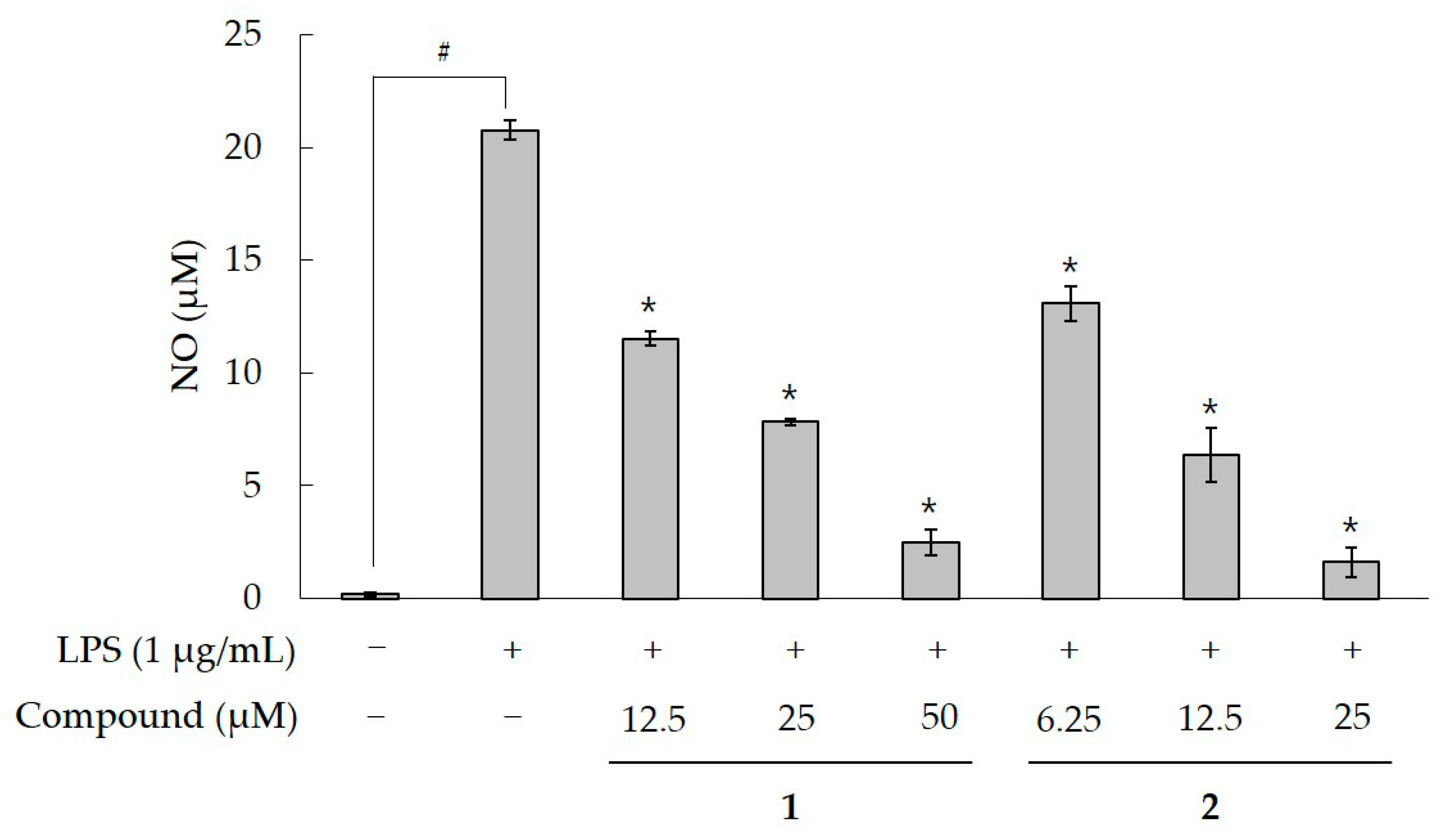
2.2. Effect of Compound 1 on NO production in LPS-Activatied RAW 264.7 Cells
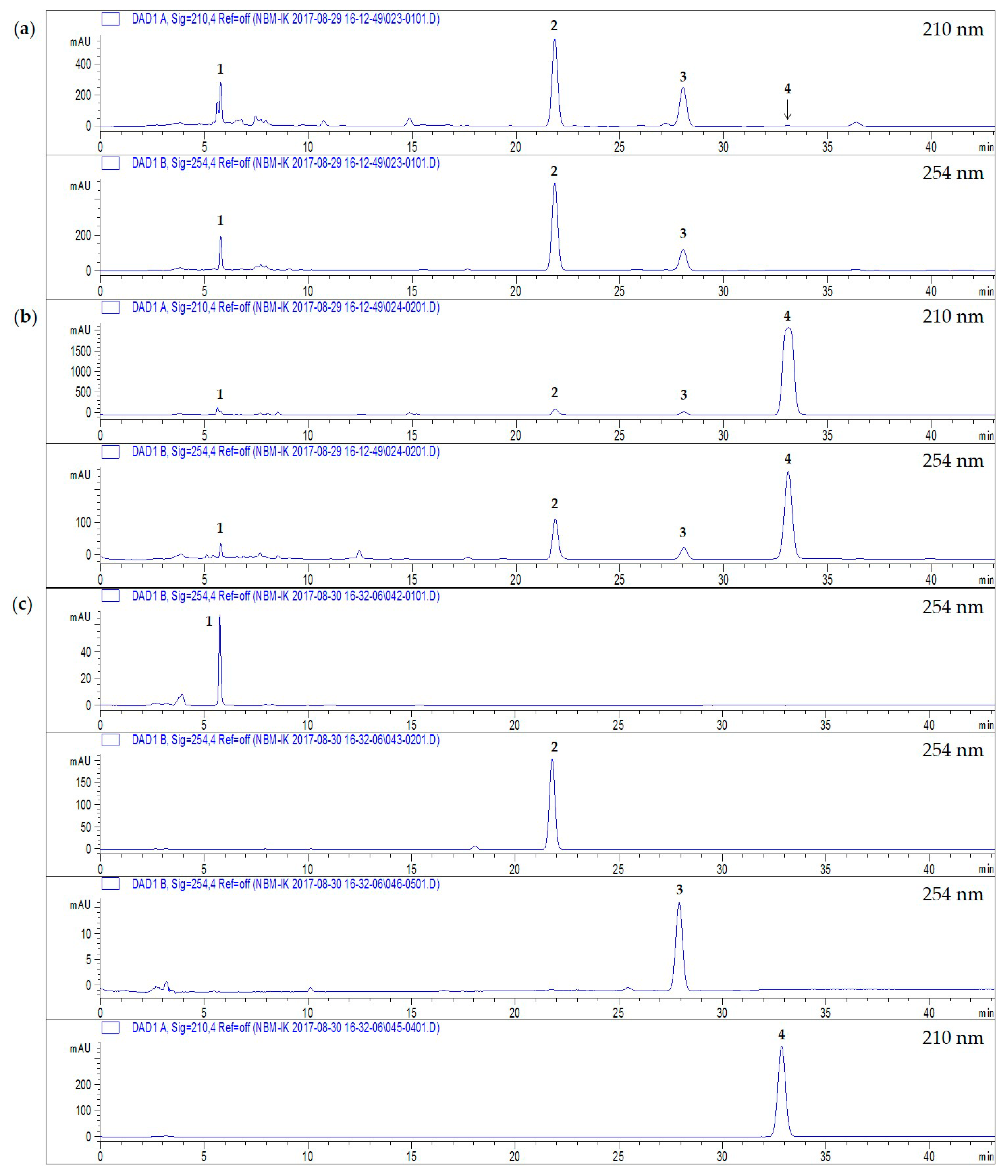
2.3. Quantitative Analysis of Four Compounds from Two Perilla Cultivars
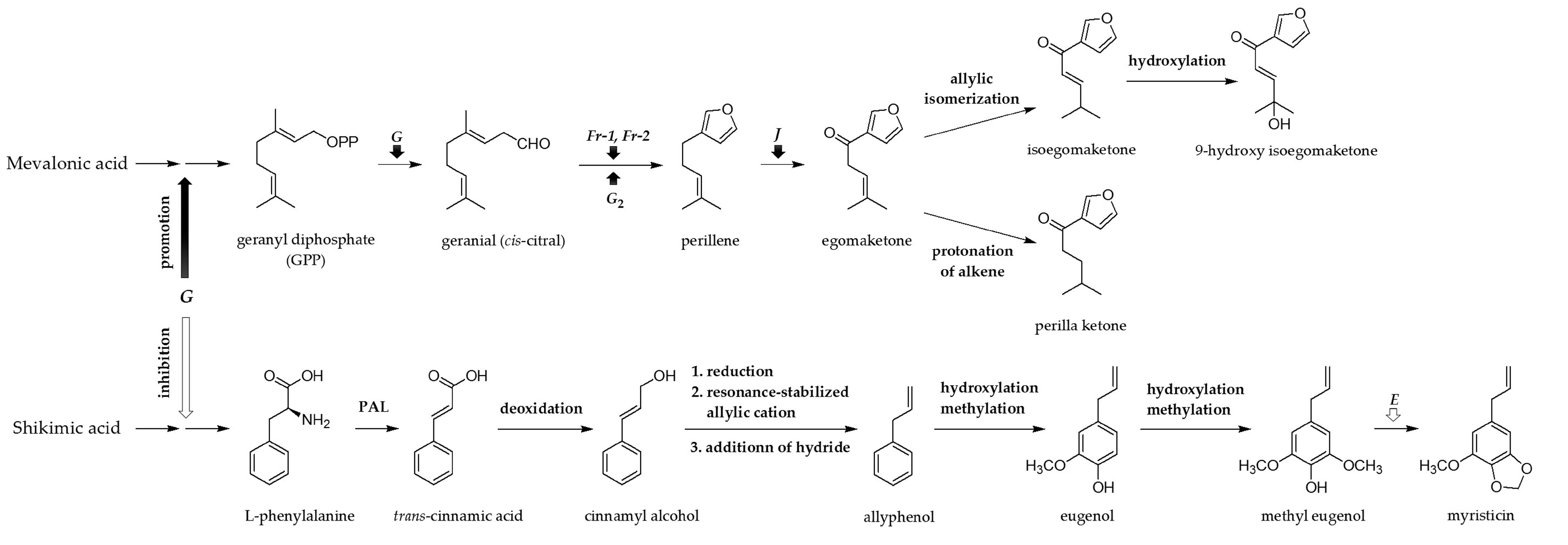
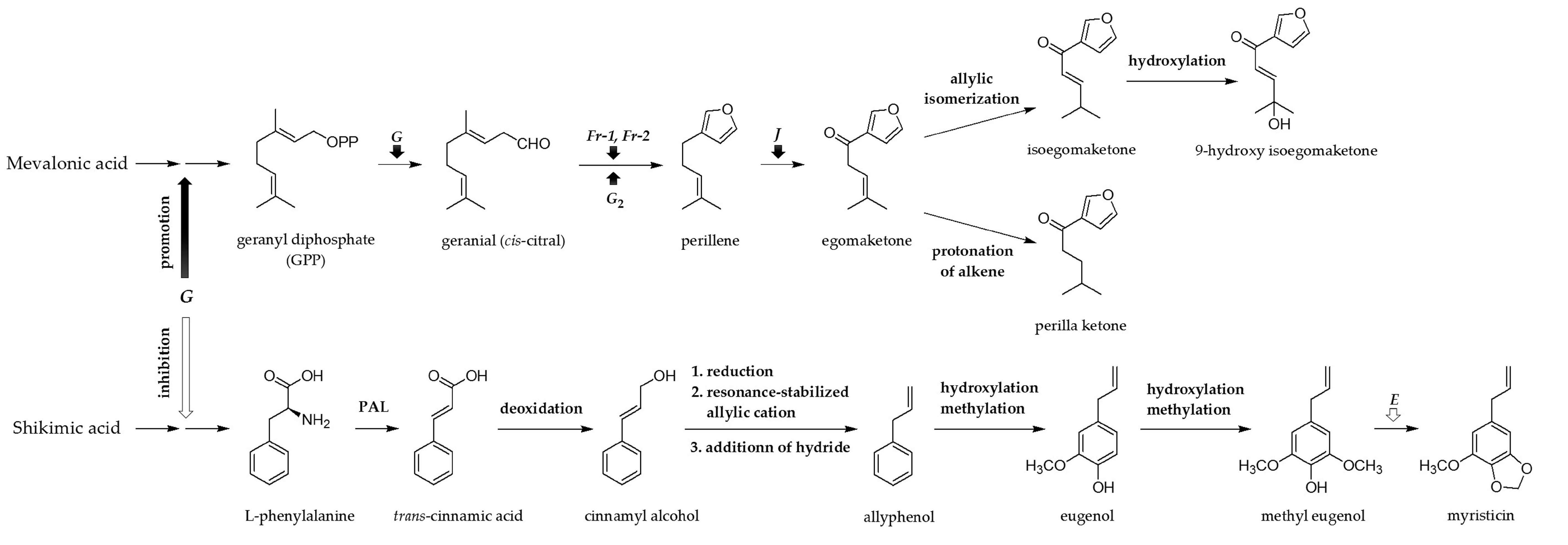
3. Discussion
4. Materials and Methods
4.1. General Procedures
4.2. Plant Materials
4.3. Extraction and Isolation
4.4. HPLC-DAD Anlysis
4.5. Measurement of NO Production on LPS-Stimulated RAW 264.7 Cells
4.6. Cytotoxicity Assay
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhu, Y.-P. Chinese Materia Medica: Chemistry, Pharmacology and Applications; CRC Press: Boca Raton, FL, USA, 1988; pp. 54–55. [Google Scholar]
- Tan, F.; Chen, Y.; Tan, X.; Ma, Y.; Peng, Y. Chinese materia medica used in medicinal diets. J. Ethnopharmcol. 2017, 206, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Bumblauskienl, L.; Jakstas, V.; Janulis, V.; Mazdzieriene, R.; Ragazinskiene, O. Preliminary analysis on essential oil composition of Perilla L. cultivated in Lithuania. Acta Pol. Pharm. 2009, 66, 409–413. [Google Scholar]
- Tian, J.; Wang, Y.; Lu, Z.; Sun, C.; Zhang, M.; Zhu, A.; Peng, X. Perillaldehyde, a promising antifungal agent used in food preservation, triggers apoptosis through a metacaspase-dependent pathway in Aspergillus flavus. J. Agric. Food Chem. 2016, 64, 7404–7413. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Li, Y.; Fu, Q.; Ma, S. Perillaldehyde attenuates cerebral ischemia-reperfusion injury-triggered overexpression of inflammatory cytokines via modulating Akt/JNK pathway in the rat brain cortex. Biochem. Biophys. Res. Commun. 2014, 454, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.H.; Lee, H.; Lee, J.S.; Young, C.Y.F. Perillyl alcohol inhibits the expression and function of the androgen receptor in human prostate cancer cells. Cancer Lett. 2006, 236, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Cho, B.O.; Jin, C.H.; Park, Y.D.; Ryu, H.W.; Byun, M.W.; Seo, K.I.; Jeong, I.Y. Isoegomaketone induces apoptosis through caspase-dependent and caspase-independent pathways in human DLD1 cells. Biosci. Biotechnol. Biochem. 2011, 75, 1306–1311. [Google Scholar] [CrossRef] [PubMed]
- Loutrari, H.; Hatziapostolou, M.; Skouridou, V.; Papadimitriou, E.; Roussos, C.; Kolisis, F.N.; Papapetropoulos, A. Perillyl alcohol is an angiogenesis inhibitor. J. Pharmacol. Exp. Ther. 2004, 311, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.H.; Lee, H.J.; Park, Y.D.; Choi, D.S.; Kim, D.S.; Kang, S.-Y.; Seo, K.-I.; Jeong, I.Y. Isoegomaketone inhibits lipopolysaccharide-induced nitric oxide production in RAW 264.7 macrophages through the heme oxygenase-1 induction and inhibition of the interferon-β-STAT-1 pathway. J. Agric. Food Chem. 2010, 58, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.H.; So, Y.; Kim, J.-B.; Han, S.N. Isoegomaketone upregulates heme oxygenase-1 in RAW264.7 cells via ROS/p38 MAPK/Nrf2 pathway. Biomol. Ther. 2016, 24, 510–516. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Kondo, T.; Kameda, K.; Goto, T. Structure of anthocyanins isolated from purple leaves of Perilla ocimoides L. var. crispa Benth and their isomerization by irradiation of light. Agric. Biol. Chem. 1990, 54, 1745–1751. [Google Scholar]
- Tada, M.; Matsumoto, R.; Yamaguchi, H.; Chiba, K. Novel antioxidants isolated from Perilla frutescens Britton var. crispa (Thunb.). Biosci. Biotechnol. Biochem. 1996, 60, 1093–1095. [Google Scholar]
- Park, Y.D.; Lee, Y.M.; Kang, M.A.; Lee, H.J.; Jin, C.H.; Choi, D.S.; Kim, D.S.; Kang, S.-Y.; Kim, W.-G.; Jeong, I.Y. Phytochemical profiles and in vitro anti-inflammatory properties of Perilla frutescens cv. Chookyoupjaso mutants induced by mutagenesis with γ-ray. Food Sci. Biotechnol. 2010, 19, 305–311. [Google Scholar] [CrossRef]
- Kwon, K.H.; Kim, K.I.; Jun, W.J.; Shin, D.H.; Cho, H.Y.; Hong, B.S. In vitro and in vivo effects of macrophage-stimulatory polysaccharide from leaves of Perilla frutescens var. crispa. Biol. Pharm. Bull. 2002, 25, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, A.; Yamamoto, Y.; Yoshinaka, N.; Namba, M.; Matsuo, H.; Okuyama, T.; Yoshigai, E.; Okumura, T.; Nishizawa, M.; Ikeya, Y. A new flavanone and other flavonoids from green perilla leaf extract inhibit nitric oxide production in interleukin 1β-treated hepatocytes. Biosci. Biotechnol. Biochem. 2015, 79, 138–146. [Google Scholar] [CrossRef] [PubMed]
- So, Y.; Lee, S.Y.; Han, A.-R.; Kim, J.-B.; Jeong, H.G.; Jin, C.H. Rosmarinic acid methyl ester inhibits LPS-induced NO production via suppression of MyD88-dependent and -independent pathways and induction of HO-1 in RAW 264.7 cells. Molecules 2016, 21, 1083. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.H.; Yang, H.S.; Choi, D.S.; Byun, M.W.; Kim, W.G.; Jeong, I.Y. Rosmarinic acid attenuated SIN-1-induced cytotoxicity in HepG2 cells through the HO-1 induction and radical scavenging activity. Food Sci. Biotechnol. 2013, 22, 549–556. [Google Scholar] [CrossRef]
- The Joint FAO/IAEA Mutant Variety Database. Available online: https://mvd.iaea.org (accessed on 4 September 2017).
- Park, H.C.; So, Y.; Kim, J.-B.; Yuk, H.S.; Jin, C.H. Comparison study of anti-inflammatory activity of extracts with supercritical carbon dioxide from radiation mutant Perilla frutescens (L.) Britton and wild-type. J. Radiat. Ind. 2016, 10, 97–104. [Google Scholar]
- Jin, C.H.; Park, H.C.; So, Y.; Nam, B.; Han, S.N.; Kim, J.-B. Comparison of the anti-inflammatory activities of supercritical carbon dioxide versus ethanol extracts from leaves of Perilla frutescens Britt. radiation mutant. Molecules 2017, 22, 311. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, W.; Yeo, S.; Choi, Y. Supercritical carbon dioxide extraction of Perilla seed oil. Food Sci. Biotechnol. 1996, 5, 300–304. [Google Scholar]
- Chiappini, L.; Perraudin, E.; Durand-Jolibois, R.; Doussin, J.F. Development of a supercritical fluid extraction-gas chromatography mass spectrometry method for the identification of highly polar compounds in secondary organic aerosols formed from biogenic hydrocarbons in smog chamber experiments. Anal. Bioanal. Chem. 2006, 386, 1749–1759. [Google Scholar] [CrossRef] [PubMed]
- You, C.X.; Jiang, H.Y.; Zhang, W.J.; Guo, S.S.; Yang, K.; Lei, N.; Ma, P.; Geng, Z.F.; Du, S.S. Contact toxicity and repellency of the main components from the essential oil of Clausena anisum-olens against two stored product insects. J. Insect Sci. 2015, 15, 87. [Google Scholar] [CrossRef] [PubMed]
- Pavia, D.L.; Lampman, G.M.; Kriz, G.S. Introduction to Spectroscopy, 3rd ed.; Thomson Learning, Ltd.: London, UK, 2001; pp. 353–386. [Google Scholar]
- Park, Y.D.; Jin, C.H.; Choi, D.S.; Byun, M.-W.; Jeong, I.Y. Biological evaluation of isoegomaketone isolated from Perilla frutescens and its synthetic derivatives as anti-inflammatory agents. Arch. Pharm. Res. 2011, 34, 1277–1282. [Google Scholar] [CrossRef] [PubMed]
- Zamora, R.; Vodovotz, Y.; Billiar, T.R. Inducible nitric oxide synthase and inflammatory diseases. Mol. Med. 2000, 6, 347–373. [Google Scholar] [PubMed]
- Nam, B.; Lee, S.Y.; Kim, J.-B.; Kang, S.-Y.; Jin, C.H. Simultaneous determination of isoegomaketone and perillaketone in Perilla frutescens (L.) Britton leaves by HPLC-DAD. korean J. Pharmacogn. 2016, 47, 79–83. [Google Scholar]
- Tabata, M. Genetics of monoterpene biosynthesis in Perilla plants. Plant Biotechnol. 2000, 17, 273–280. [Google Scholar] [CrossRef]
- Koezuka, Y.; Honda, G.; Tabata, M. Genetic control of the chemical composition of volatile oils in Perilla frutescens. Phytochemistry 1986, 25, 859–863. [Google Scholar] [CrossRef]
- Yuba, A.; Honda, G.; Koezuka, Y.; Tabata, M. Genetic analysis of essential oil variants in Perilla frutescens. Biochem. Genet. 1995, 33, 341–348. [Google Scholar]
- Nishizawa, A.; Honda, G.; Tabata, M. Genetic control of perillene accumulation in Perilla frutescens. Phytochemisty 1990, 29, 2873–2875. [Google Scholar] [CrossRef]
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach, 2nd ed.; John Wiley & Sons Ltd.: West Sussex, UK, 2002; pp. 7–34, 121–140. [Google Scholar]
- Waltenberger, B; Mocan, A.; Šmejkal, K.; Heiss, E.H.; Atanasov, A.G. Natural products to counteract the epidemic of cardiovascular and metabolic disorders. Molecules 2016, 21, 807. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [PubMed]
- Kinghorn, A.D.; Pan, L.; Fletcher, J.N.; Chai, H. The relevance of higher plants in lead compound discovery programs. J. Nat. Prod. 2011, 74, 1539–1555. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | tR (min) | Linear Range (μg/mL) | Regression Equation (y = ax + b) 1 | Correlation Coefficient (r2) | LOD 2 (μg/mL) | LOQ 3 (μg/mL) |
|---|---|---|---|---|---|---|
| 1 | 5.78 | 20−100 | y = 13.154x − 8.1524 | 0.9991 | 0.103 | 0.641 |
| 2 | 21.87 | 20−164 | y = 44.574x − 27.463 | 0.9992 | 0.067 | 0.952 |
| 3 | 28.04 | 20−100 | y = 7.8236x − 0.9952 | 0.9992 | 0.006 | 0.116 |
| 4 | 33.10 | 100−1000 | y = 10.824x − 67.536 | 0.9992 | 0.124 | 0.375 |
| Peak | Compound | Contents (mg/g) | |
|---|---|---|---|
| PFCA | PFC | ||
| 1 | 9-hydroxy-isoegomaketone (1) | 1.33 ± 0.07 | 0.15 ± 0.02 |
| 2 | isoegomaketone (2) | 2.76 ± 0.05 | 0.51 ± 0.02 |
| 3 | perilla ketone (3) | 6.96 ± 0.17 | 1.71 ± 0.12 |
| 4 | myristicin (4) | 0.042 ± 0.003 | 36.77 ± 5.60 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nam, B.; So, Y.; Kim, H.-Y.; Kim, J.-B.; Jin, C.H.; Han, A.-R. A New Monoterpene from the Leaves of a Radiation Mutant Cultivar of Perilla frutescens var. crispa with Inhibitory Activity on LPS-Induced NO Production. Molecules 2017, 22, 1471. https://doi.org/10.3390/molecules22091471
Nam B, So Y, Kim H-Y, Kim J-B, Jin CH, Han A-R. A New Monoterpene from the Leaves of a Radiation Mutant Cultivar of Perilla frutescens var. crispa with Inhibitory Activity on LPS-Induced NO Production. Molecules. 2017; 22(9):1471. https://doi.org/10.3390/molecules22091471
Chicago/Turabian StyleNam, Bomi, Yangkang So, Hyo-Young Kim, Jin-Baek Kim, Chang Hyun Jin, and Ah-Reum Han. 2017. "A New Monoterpene from the Leaves of a Radiation Mutant Cultivar of Perilla frutescens var. crispa with Inhibitory Activity on LPS-Induced NO Production" Molecules 22, no. 9: 1471. https://doi.org/10.3390/molecules22091471
APA StyleNam, B., So, Y., Kim, H.-Y., Kim, J.-B., Jin, C. H., & Han, A.-R. (2017). A New Monoterpene from the Leaves of a Radiation Mutant Cultivar of Perilla frutescens var. crispa with Inhibitory Activity on LPS-Induced NO Production. Molecules, 22(9), 1471. https://doi.org/10.3390/molecules22091471