
A Log Ratio-Based Analysis of Individual Changes in the Composition of the Oral Microbiota in Different Dietary Phases

 ,
,  , ,
, ,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. The Data
2.2. Analytic Strategy
- For each pair of bacteria, the ratio between the two fractions is built for each participant and phase. Such a pairwise ratio directly depicts the relation between two bacteria. They do not depend on the concentration of the other bacteria or the overall concentration. If a third bacterium has an unusually high or low concentration, this affects the overall concentration and hence the fractions, but it does not affect the ratio. Therefore, in this way we become independent of the behaviour of other bacteria. Ratios also have a simple direct interpretation describing how more frequent one bacterium is compared with the other. Note that a ratio between two fractions can also be interpreted directly as the ratio between two corresponding concentrations.
- The second step is to consider the logarithm with the base 10 of the ratio. This is useful because ratios tend to have a skewed and unstable distribution, whereas taking logarithms leads to a more symmetric distribution of the values. The basis 10 was chosen due to its strong interpretability: a value of 1 in the scale means the tenfold in the real concentration setting. A ratio of 1—i.e., equal concentrations—has a value of 0 in the setting. For ratios smaller than 1, the transformed value becomes negative. Therefore, a positive (negative) value means that the first bacteria is less (more) frequent than the second one.
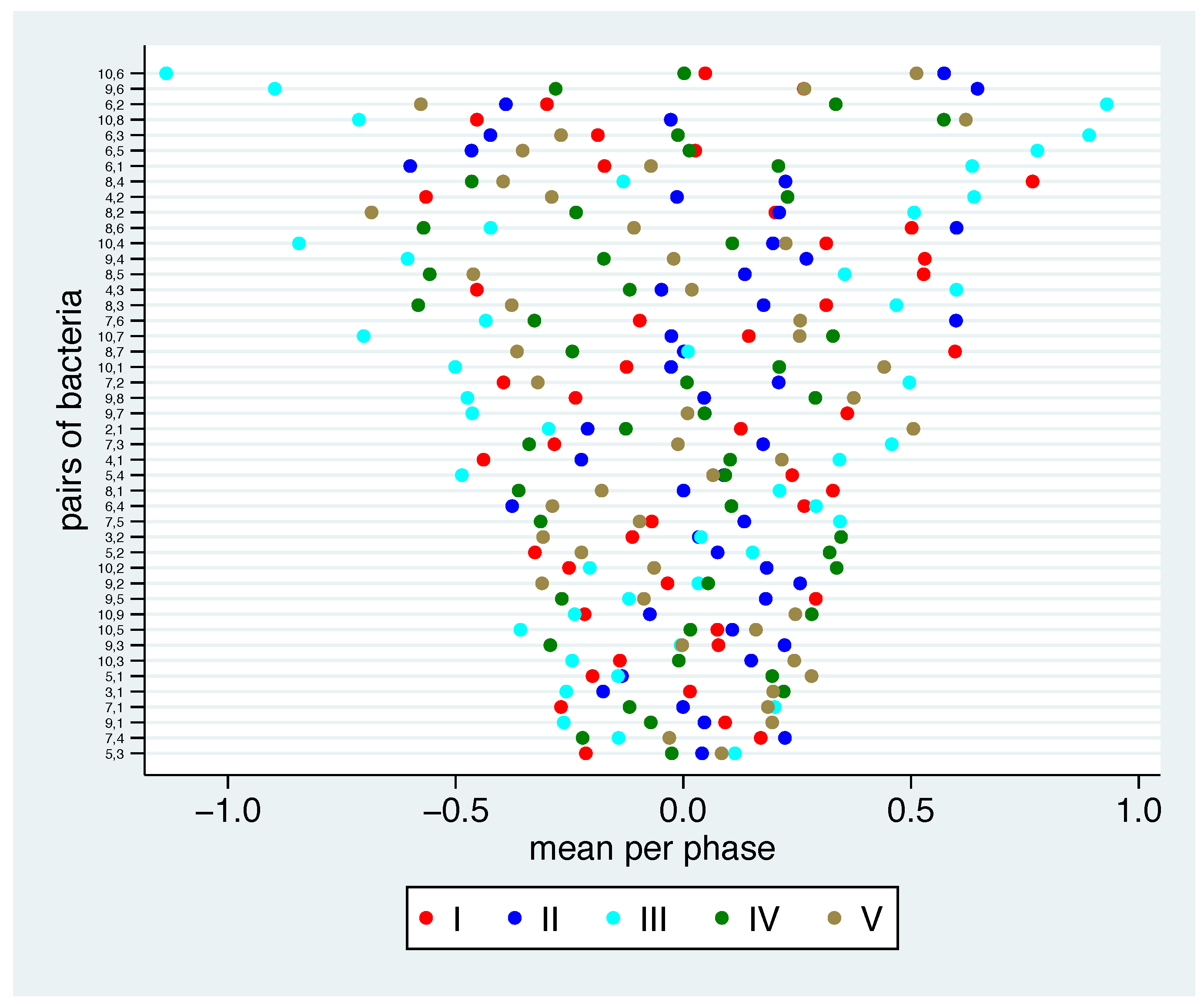
- Logarithmising the ratios leads to one number per participant and phase, which can be analysed just like any other number [15]. In particular, mean values and standard deviations can be calculated. In this step, for each pair of bacteria we consider the mean value of the log ratios across all participants per phase. It is interpretable as the typical relationship between these two bacteria in this phase. Therefore, this mean value offers the opportunity to check, whether the relationship between two bacteria is similar in different phases or whether it changes due to changes in the nutrition. Accordingly, we can analyse the stability between bacteria between the phases. Similarly as above, a mean value of 1 suggests that typically the first bacterium has ten times the concentration of the second bacterium.
- We also analyse the corresponding standard deviations, which are a measure of the stability of the relationship between two bacteria within one phase. The closer that the value is to 0, the more homogeneous are the values across participants, i.e., the relation between the two considered bacteria is stable and can thus be assumed as “typical”. For example, if we observe a standard deviation of 0.5 and a mean log ratio of 0 for two bacteria, then the application of the 2 sigma rule leads to a 95% range of (−1, 1), i.e., 95% of the observed ratios are between 1/10 and 10.
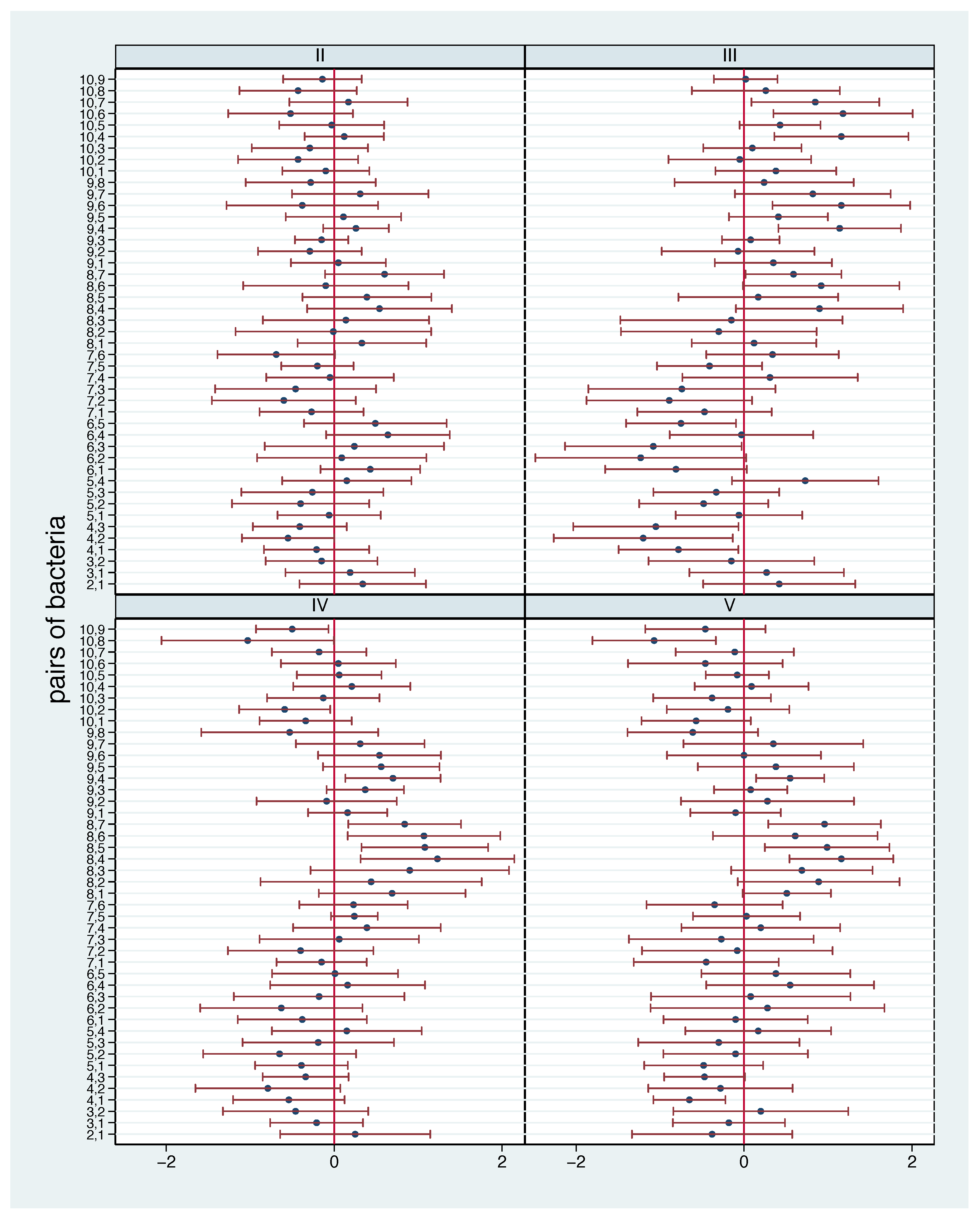
- As the aim is to study the impact of an additional standardised diet on the microbiome, we also depict the changes of the concentrations compared with the first phase. For a given pair of bacteria, for each participant and phase we consider the difference in the log ratio compared with the first phase. A difference above 0 means that the ratio increased, i.e., the first bacterium became more frequent relative to the second one. As a difference in the log ratio is equal to the logarithm of the ratio of ratios, we can also interpret the values themselves. A difference of 1 means, that the ratio has increased by a factor of ten. For example, this may reflect a change from 1.2 to 12, or from 0.3 to 3. Again, we can consider mean values and standard deviations across participants per phase for phases II to V. The mean values inform us about the typical change in the relation between the two bacteria. The standard deviations offer a hint about the stability of these changes across the participants.
- Finally, we also consider a bacterium-specific summary measure, which is based on averaging the log ratio changes for one bacterium over all other bacteria within a single participant If this way a bacterium has a value greater than 0, this can be interpreted as having increased its fraction on average relative to all other bacteria. The bacterium with the largest positive value is the one with the most distinct increase compared with all other bacteria. We refer to this value for each participant as the average log ratio change.
2.3. Analysing Change or Comparing Phases Directly
3. Results
3.1. A First Look at the Raw Data
3.2. Stability of the Relation between Bacterial Groups between Phases
3.3. Stability of the Relation between Bacterial Groups within Phases
3.4. Identification of Bacterial Pairs with a Change in Relation in Comparison with the Baseline
3.5. Stability of Change in Relation between Phases
3.6. Stability of Change in Relation within Phases
3.7. Interim Summary
3.8. Average Log Ratio Changes
Global p-Value Per Phase
3.9. Similarity of Change Pattern Across Bacterial Groups
4. Discussion
- The bacterial pair Gemella Granulicatella A. spp.–Streptococcus spp. group 2 is the one with high stability across all analyses.
- The bacterial pairs Actiomyces. spp.–HACEK and Actiomyces. spp.–Campylobacter spp. are the bacterial pairs with low stability across all analyses.
- The bacterial pair Neisseria spp.–Streptococcus spp. group 3 could be identified as the bacterial pair that most distinctly reacts to the dietary supplements.
- Bacterial pairs with high stability in at least both a within-phase and between-phase analysis were Gemella Granulicatella A. spp.–Capnocytophaga spp., Rothia spp.–Streptococcus spp. group 2, Capnocytophaga spp.–Streptococcus spp. group 3 and Streptococcus spp. group 2–Streptococcus spp. group 3.
- The most distinct changes in the spectrum were observed in phase III, i.e., the phase with additional milk products. In particular, the relation of Neisseria spp. to Streptococcus spp. group 2 and group 3 changed. When looking at the single bacterial species, the most distinct change was a decrease of Neisseria spp. and HACEK.
- In phases IV and V, when looking at the single bacterial species, the increase of Campylobacter spp. should be mentioned. On the other hand, the figures in phases IV and V were very similar, although in phase V the participants returned to their individual nutrition without specific additions.
- For Gemella Granulicatella A. spp., Rothia spp. and Streptococcus spp. group 3, we observed rather stable relations to other bacterial groups, while for HACEK and Campylobacter spp. the relations are rather variable.
Further Insights
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Appendix A. Description of the Bacterial Groups
- Gemella Granulicatella Abiotrophia spp.: Gemella morbillorum, Gemella haemolysans, Gemella sanguinis, Granulicatella adiacens, Granulicatella elegans, Abiotrophia defectiva
- Actinomyces spp.: Actinomyces oris, Actinomyces odontolyticus, Actinomyces dentalis, Actinomyces georgiae, Actinomyces naeslundii
- Rothia spp.: Rothia mucilaginosa, Rothia dentocariosa, Rothia aeria, Corynebacterium spp.
- Neisseria spp.: Neisseria macacae/mucosa, Neisseria oralis, Neisseria subflava, Neisseria bacilliformis, Neisseria elongata, Neisseria flavescens, Neisseria perflava, Neisseria cinerea, Lautrop mirabilis
- Capnocytophaga spp.: Capnocytophaga granulosa, Capnocytophaga gingivalis, Capnocytophaga ochracea, Capnocytophaga sputigena
- HACEK: Haemophilus haemolyticus, Haemophilus parahaemolyticus, Haemophilus parainfluenzae, Haemophilus influenzae, Cardiobacterium hominis, Eikenella corrodens, Kingella spp.
- Fusobacterium spp.: Fusobacterium nucleatum, Fusobacterium periodontium
- Campylobacter spp.: Campylobacter rectus, Campylobacter concisus, Campylobacter showae
- Streptococcus spp. group 2: Streptococcus oralis, Streptococcus mitis, Streptococcus infantis, Streptococcus australis, Streptococcus peroris, Streptococcus salivarius, Streptococcus vestibularis, Streptococcus anginosus group
- Streptococcus spp. group 3: Streptococcus sanguinis, Streptococcus parasanguinis, Streptococcus gordonii
Appendix B. Materials and Methods
Appendix B.1. Notation
Appendix B.1.1. Log Ratio-Based Analysis of Fractions
Appendix B.1.2. Pairwise Differences in Log Ratio Changes and Their Characteristics
Appendix B.1.3. Average Log Ratio Changes
Appendix B.2. Evidence of Composition Change
References
- Anderson, A.C.; Rothballer, M.; Altenburger, M.J.; Woelber, J.P.; Karygianni, L.; Vach, K.; Hellwig, E.; Al-Ahmad, A. Long-term fluctuation of oral biofilm microbiota following different dietary phases. Appl. Environ. Microbiol. 2020. [Google Scholar] [CrossRef]
- Bradshaw, D.J.; Lynch, R.J.M. Diet and the microbial aetiology of dental caries: New paradigms. Int. Dent. J. 2013, 63, 64–72. [Google Scholar] [CrossRef]
- Moynihan, P.; Petersen, P.E. Diet, nutrition and the prevention of dental diseases. Public Health Nutr. 2004, 7, 201–226. [Google Scholar] [CrossRef] [PubMed]
- Petti, S.; Simonetti, R.; Simonetti D’Arca, A. The effect of milk and sucrose consumption on caries in 6-to-11-year-old Italian schoolchildren. Eur. J. Epidemiol. 1997, 13, 659–664. [Google Scholar] [CrossRef]
- Aimutis, W.R. Bioactive Properties of Milk Proteins with Particular Focus on Anticariogenesis. J. Nutr. 2004, 134, 989S–995S. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, E.C.; Cai, F.; Shen, P.; Walker, G.D. Retention in Plaque and Remineralization of Enamel Lesions by Various Forms of Calcium in a Mouthrinse or Sugar-free Chewing Gum. J. Dent. Res. 2016. [Google Scholar] [CrossRef]
- Schlafer, S.; Ibsen, C.J.S.; Birkedal, H.; Nyvad, B. Calcium-Phosphate-Osteopontin Particles Reduce Biofilm Formation and pH Drops in in situ Grown Dental Biofilms. Caries Res. 2017, 51, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Nyvad, B. Caries ecology revisited: Microbial dynamics and the caries process. Caries Res. 2008, 42, 409–418. [Google Scholar] [CrossRef]
- Marsh, P.D. In Sickness and in Health—What Does the Oral Microbiome Mean to Us? An Ecological Perspective. Adv. Dent. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Mira, A. Oral Microbiome Studies: Potential Diagnostic and Therapeutic Implications. Adv. Dent. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Aas, J.A.; Griffen, A.L.; Dardis, S.R.; Lee, A.M.; Olsen, I.; Dewhirst, F.E.; Leys, E.J.; Paster, B.J. Bacteria of Dental Caries in Primary and Permanent Teeth in Children and Young Adults. J. Clin. Microbiol. 2008, 46, 1407–1417. [Google Scholar] [CrossRef] [PubMed]
- Levy, R.M.; Giannobile, W.V.; Feres, M.; Haffajee, A.D.; Smith, C.; Socransky, S.S. The Effect of Apically Repositioned Flap Surgery on Clinical Parameters and the Composition of the Subgingival Microbiota: 12-Month Data. Int. J. Periodontics Restor. Dent. 2002, 22, 209–219. [Google Scholar]
- Koopman, J.E.; van der Kaaij, N.C.W.; Buijs, M.J.; Elyassi, Y.; van der Veen, M.H.; Crielaard, W.; Ten Cate, J.M.; Zaura, E. The Effect of Fixed Orthodontic Appliances and Fluoride Mouthwash on the Oral Microbiome of Adolescents—A Randomized Controlled Clinical Trial. PLoS ONE 2015, 10, e0137318. [Google Scholar] [CrossRef]
- Velmurugan, S.; Gan, J.M.; Rathod, K.S.; Khambata, R.S.; Ghosh, S.M.; Hartley, A.; Van Eijl, S.; Sagi-Kiss, V.; Chowdhury, T.A.; Curtis, M.; et al. Dietary nitrate improves vascular function in patients with hypercholesterolemia: A randomized, double-blind, placebo-controlled study. Am. J. Clin. Nutr. 2016, 103, 25–38. [Google Scholar] [CrossRef]
- Greenacre, M.J. Compositional Data Analysis in Practice; CRC Press: Boca Raton, FL, USA; Taylor & Francis Group: Abingdon, UK, 2019. [Google Scholar]
- Lamont, R.J.; Koo, H.; Hajishengallis, G. The oral microbiota: Dynamic communities and host interactions. Nat. Reviews. Microbiol. 2018, 16, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, S.; Karygianni, L.; Filippi, A.; Anderson, A.C.; Zürcher, A.; Hellwig, E.; Vach, K.; Macchiarelli, G.; Al-Ahmad, A. Combining culture and culture-independent methods reveals new microbial composition of halitosis patients’ tongue biofilm. MicrobiologyOpen 2020, 9, e958. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.C.; Rothballer, M.; Altenburger, M.J.; Woelber, J.P.; Karygianni, L.; Lagkouvardos, I.; Hellwig, E.; Al-Ahmad, A. In-vivo shift of the microbiota in oral biofilm in response to frequent sucrose consumption. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Vach, K.; Al-Ahmad, A.; Anderson, A.; Woelber, J.P.; Karygianni, L.; Wittmer, A.; Hellwig, E. Analysing the Relationship between Nutrition and the Microbial Composition of the Oral Biofilm—Insights from the Analysis of Individual Variability. Antibiotics 2020, 9, 479. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.C.; Hellwig, E.; Vespermann, R.; Wittmer, A.; Schmid, M.; Karygianni, L.; Al-Ahmad, A. Comprehensive Analysis of Secondary Dental Root Canal Infections: A Combination of Culture and Culture-Independent Approaches Reveals New Insights. PLoS ONE 2012, 7, e49576. [Google Scholar] [CrossRef]
- Paster, B.J.; Boches, S.K.; Galvin, J.L.; Ericson, R.E.; Lau, C.N.; Levanos, V.A.; Sahasrabudhe, A.; Dewhirst, F.E. Bacterial Diversity in Human Subgingival Plaque. J. Bacteriol. 2001, 183, 3770–3783. [Google Scholar] [CrossRef]
- Zaura, E.; Brandt, B.W.; Teixeira de Mattos, M.J.; Buijs, M.J.; Caspers, M.P.M.; Rashid, M.U.; Weintraub, A.; Nord, C.E.; Savell, A.; Hu, Y.; et al. Same Exposure but Two Radically Different Responses to Antibiotics: Resilience of the Salivary Microbiome versus Long-Term Microbial Shifts in Feces. mBio 2015, 6, e01693-15. [Google Scholar] [CrossRef] [PubMed]
- Chastin, S.; Palarea-Albaladejo, J.; Dontje, M.; Skelton, D. Combined Effects of Time Spent in Physical Activity, Sedentary Behaviors and Sleep on Obesity and Cardio-Metabolic Health Markers: A Novel Compositional Data Analysis Approach. PLoS ONE 2015, 10, e0139984. [Google Scholar] [CrossRef] [PubMed]
- Martín Fernández, J.A.; Daunis i Estadella, J.; Mateu i Figueras, G. On the Interpretation of Differences between Groups for Compositional Data. Sort Stat. Oper. Res. Trans. 2015, 39, 231–252. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | bB | w | ||||||||
| 2 | b | bwBW | W | bwBW | ||||||
| 3 | bB | w | bBW | wW | wW | |||||
| 4 | wW | W | W | bw | B | |||||
| 5 | bBW | w | bB | W | bwB | |||||
| 6 | wBW | bB | bB | |||||||
| 7 | bB | bB | W | W | ||||||
| 8 | B | wW | bB | |||||||
| 9 | bwBW | bwW | w | B | B | |||||
| 10 | W | w | bB | w | bwW | w | bwW |
| Phase II | Phase III | Phase IV | Phase V | |
|---|---|---|---|---|
| p-value | 0.459 | 0.055 | 0.096 | 0.289 |
| Phase III | Phase IV | Phase V | |
|---|---|---|---|
| Phase II | 0.199 | 0.488 | 0.628 |
| Phase III | 0.528 | 0.205 | |
| Phase IV | 0.780 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vach, K.; Al-Ahmad, A.; Anderson, A.; Woelber, J.P.; Karygianni, L.; Wittmer, A.; Hellwig, E. A Log Ratio-Based Analysis of Individual Changes in the Composition of the Oral Microbiota in Different Dietary Phases. Nutrients 2021, 13, 793. https://doi.org/10.3390/nu13030793
Vach K, Al-Ahmad A, Anderson A, Woelber JP, Karygianni L, Wittmer A, Hellwig E. A Log Ratio-Based Analysis of Individual Changes in the Composition of the Oral Microbiota in Different Dietary Phases. Nutrients. 2021; 13(3):793. https://doi.org/10.3390/nu13030793
Chicago/Turabian StyleVach, Kirstin, Ali Al-Ahmad, Annette Anderson, Johan Peter Woelber, Lamprini Karygianni, Annette Wittmer, and Elmar Hellwig. 2021. "A Log Ratio-Based Analysis of Individual Changes in the Composition of the Oral Microbiota in Different Dietary Phases" Nutrients 13, no. 3: 793. https://doi.org/10.3390/nu13030793
APA StyleVach, K., Al-Ahmad, A., Anderson, A., Woelber, J. P., Karygianni, L., Wittmer, A., & Hellwig, E. (2021). A Log Ratio-Based Analysis of Individual Changes in the Composition of the Oral Microbiota in Different Dietary Phases. Nutrients, 13(3), 793. https://doi.org/10.3390/nu13030793