
Ultrafast Fluorescence Spectroscopy via Upconversion and Its Applications in Biophysics


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
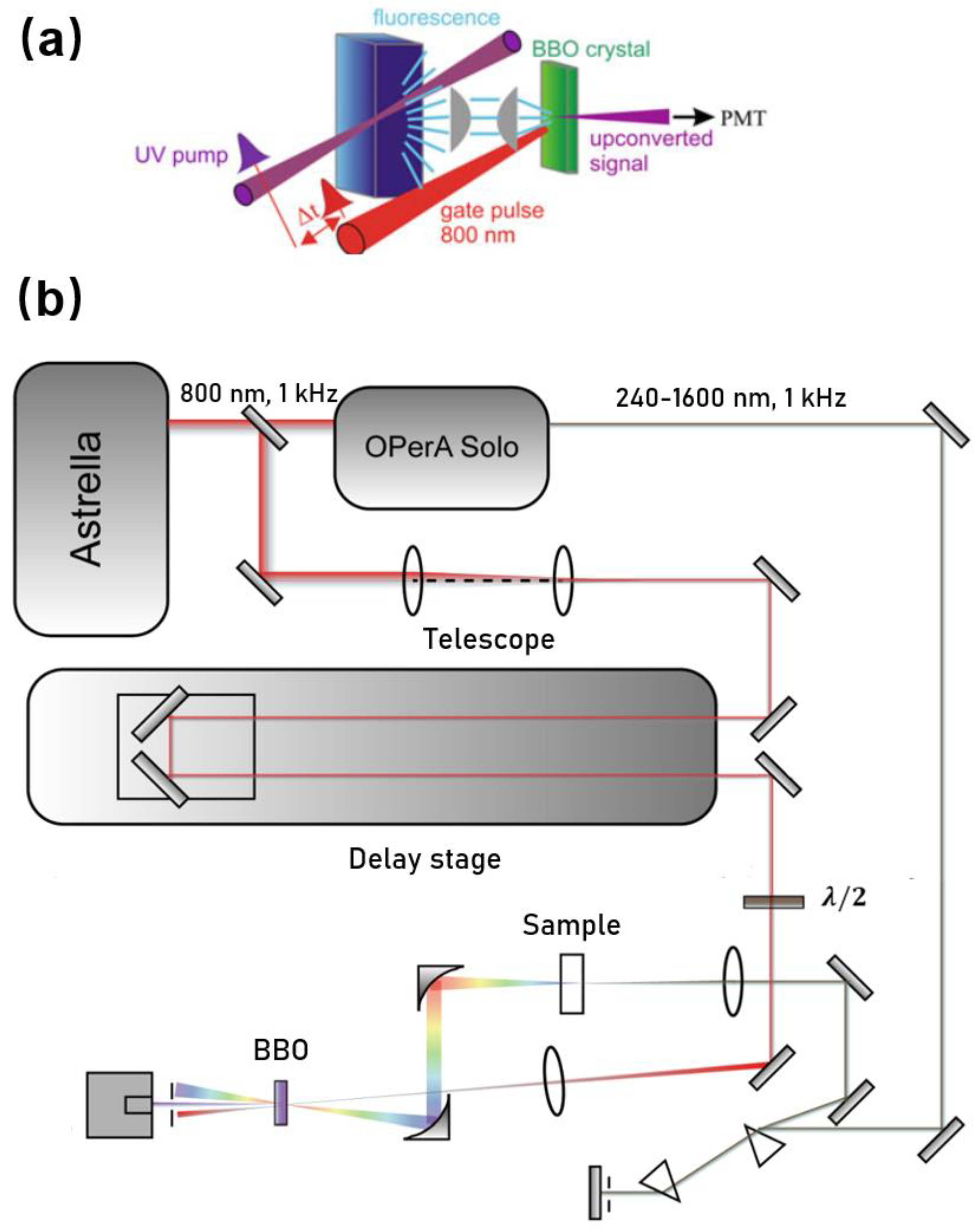
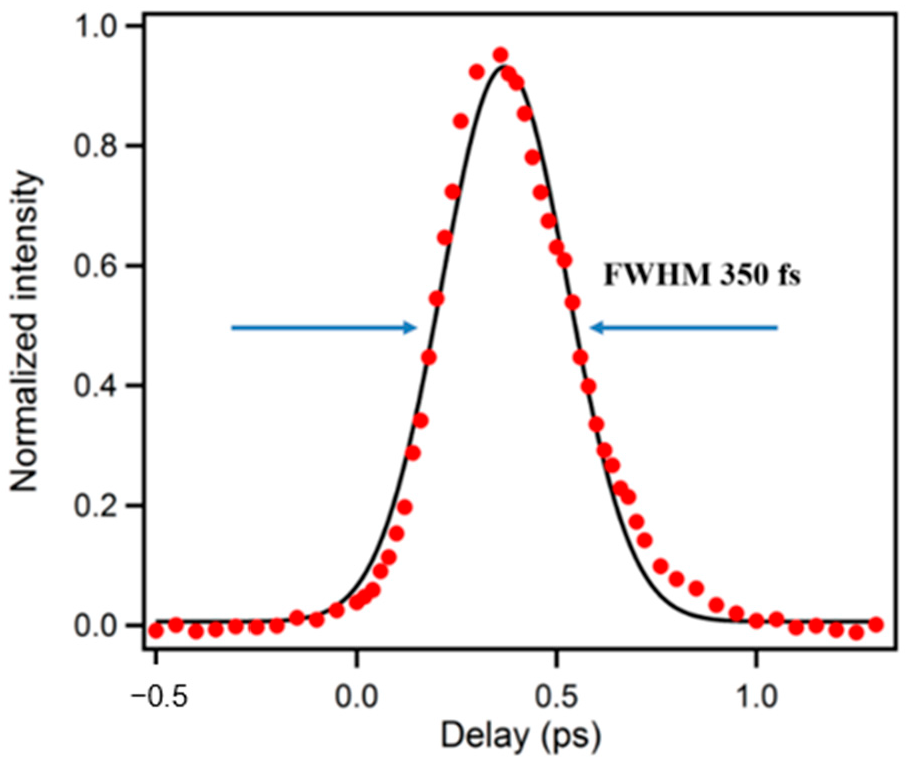
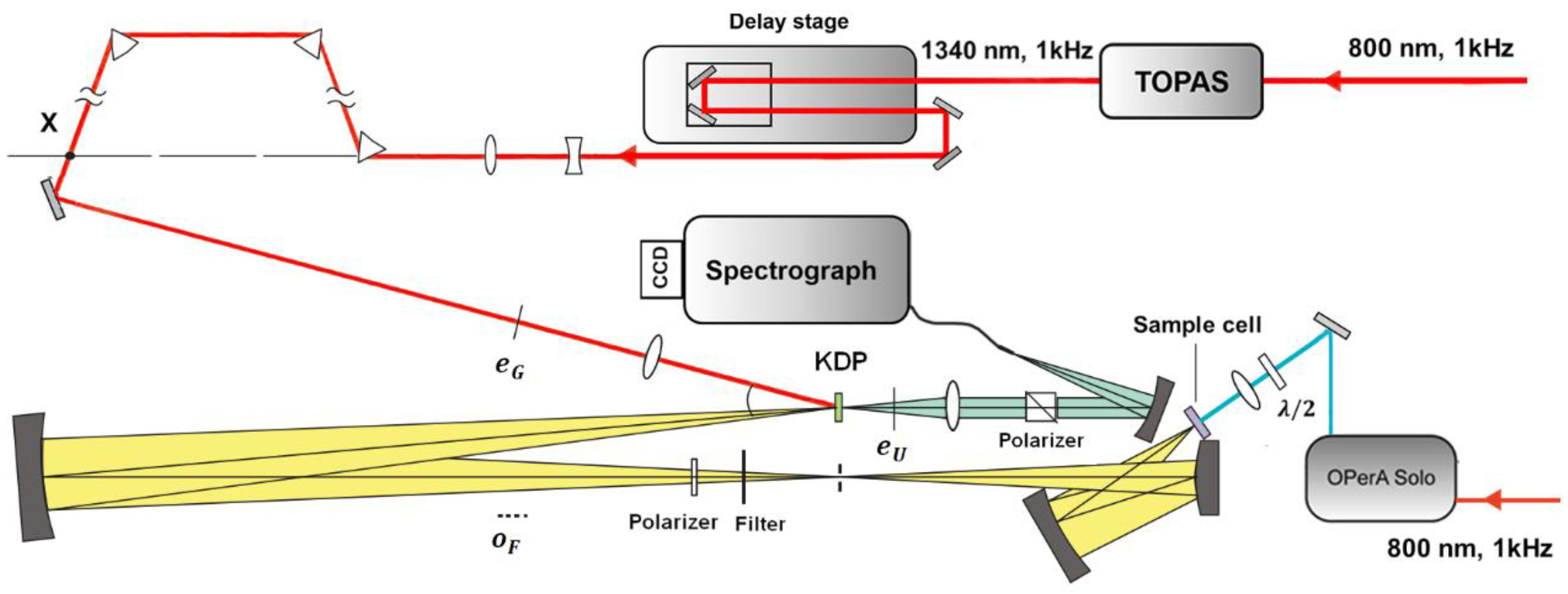
2. Experimental Set-Up and Considerations
3. Solvation Dynamics of Biomolecules: Bulk Water and “Biological” Water
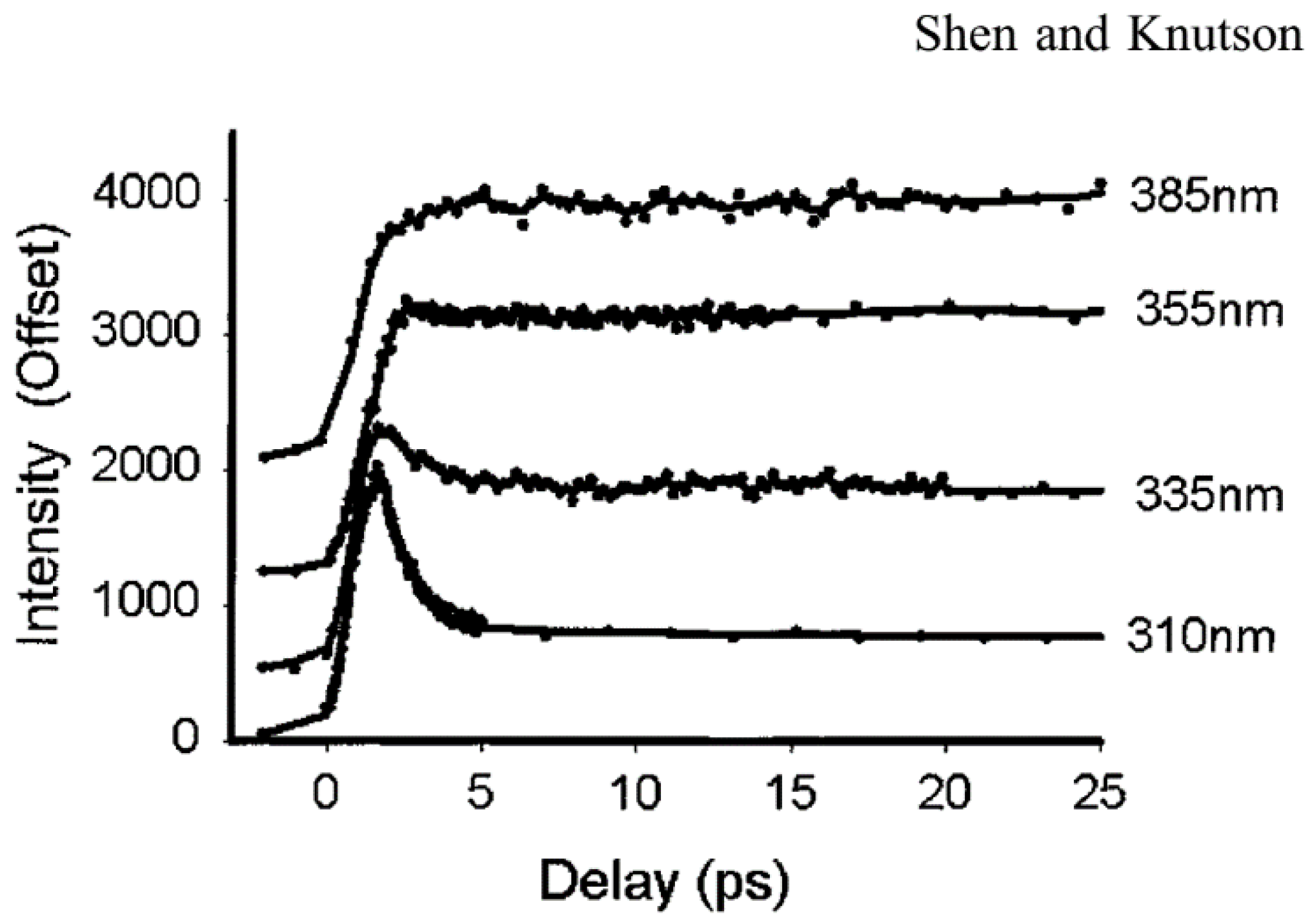
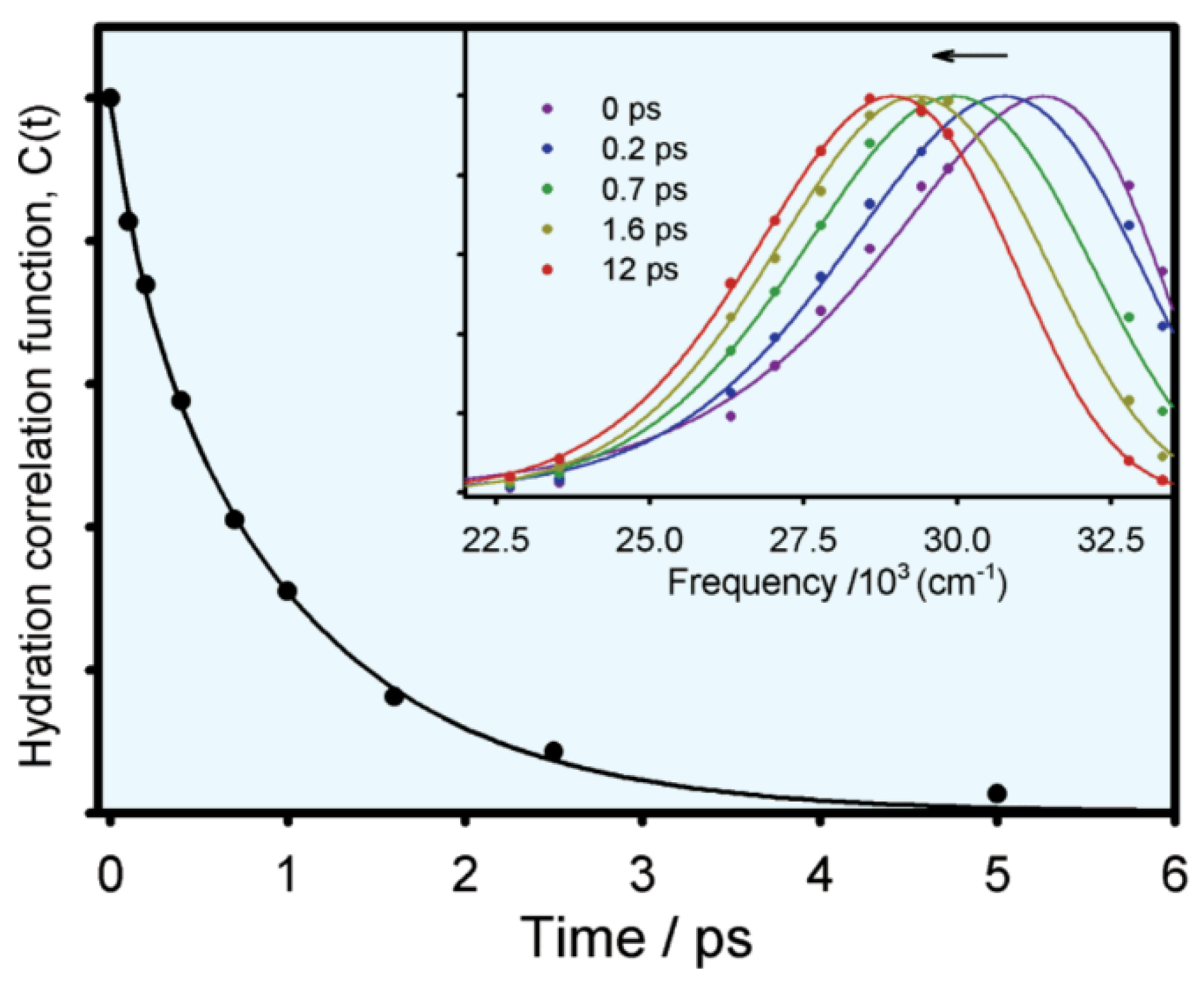
3.1. Solvent Relaxation of Tryptophan Alone in Water
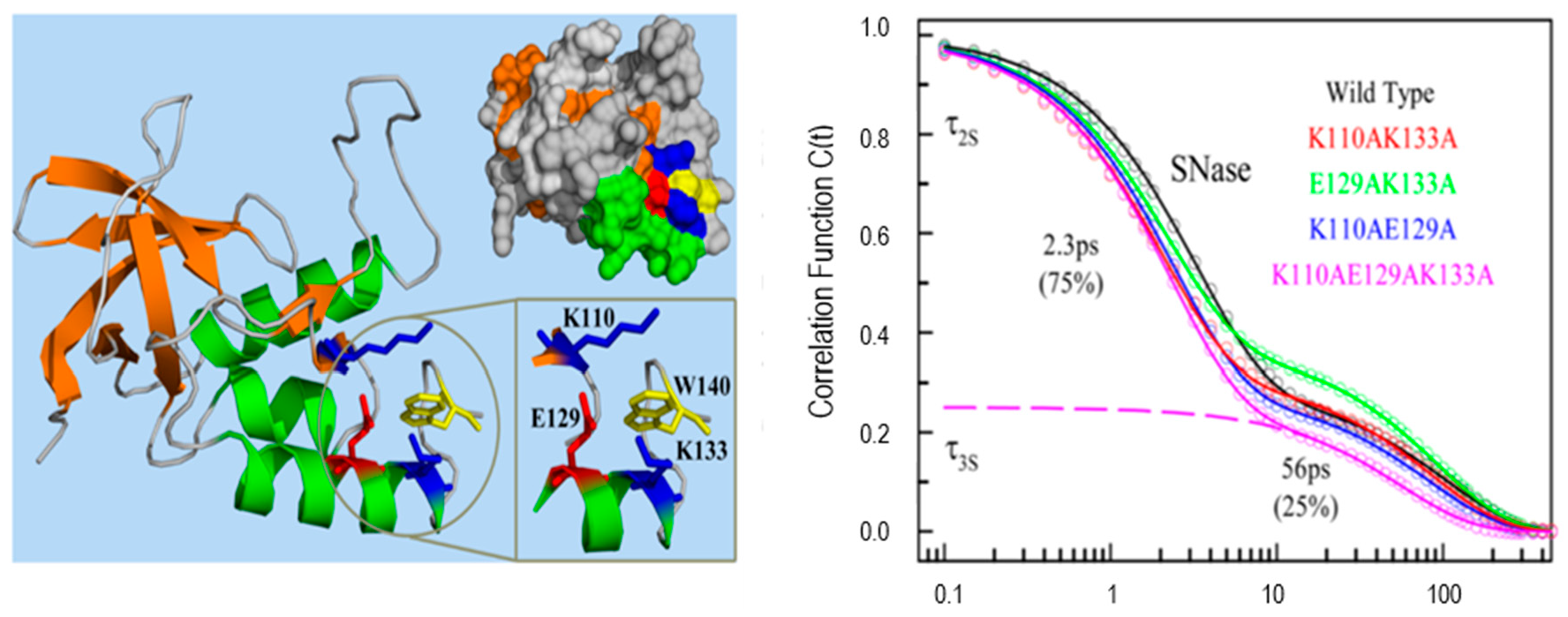
3.2. Solvation Dynamics at the Surface of Proteins
3.3. Solvation Dynamics in Free NADH
4. QSSQ in Biomolecules
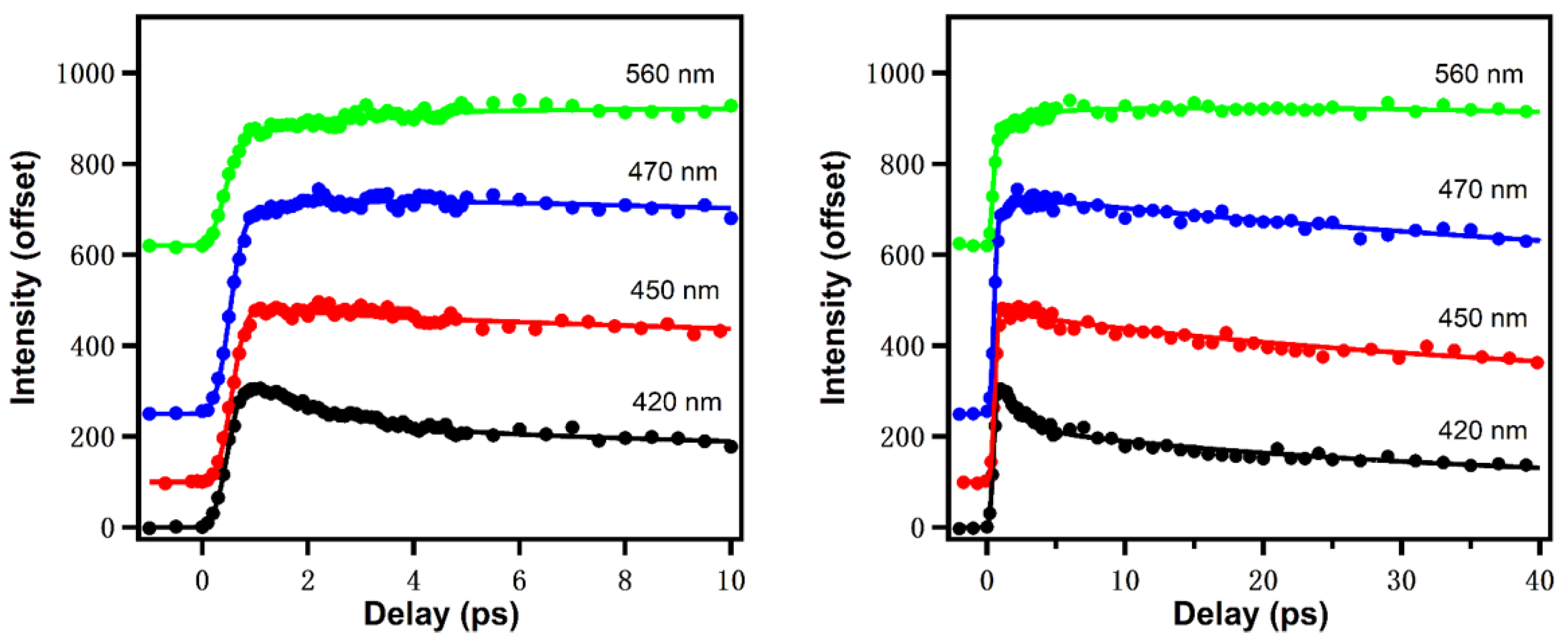
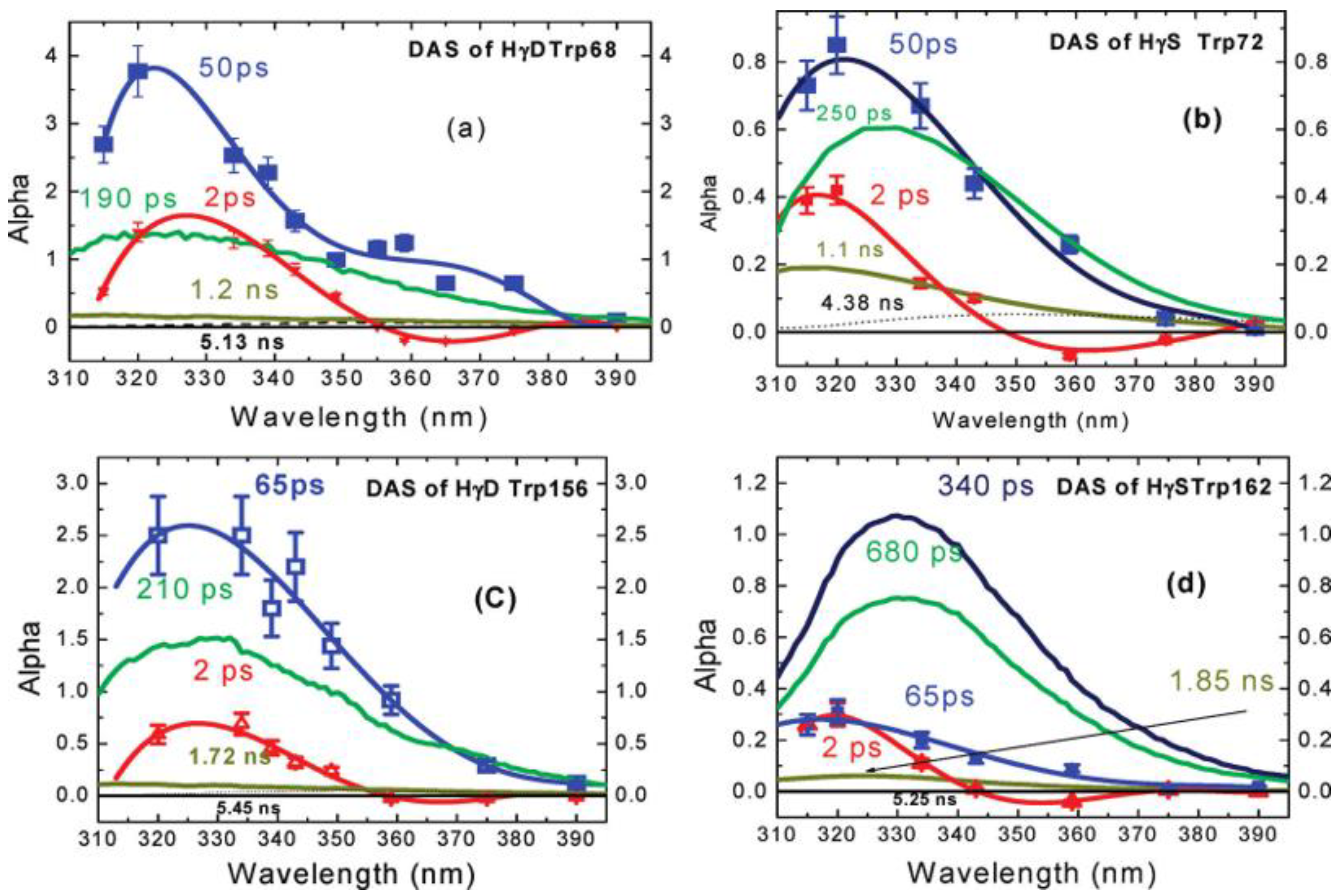
4.1. QSSQ of Tryptophan in Dipeptides and Proteins
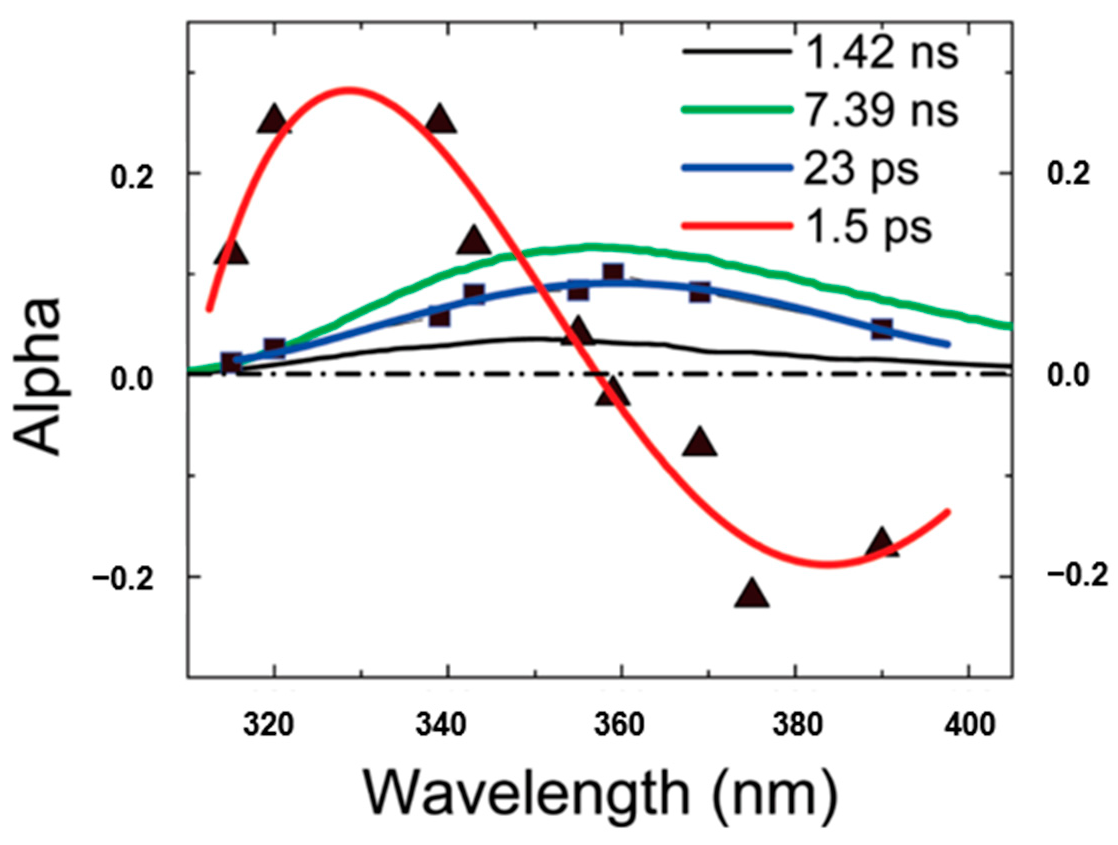
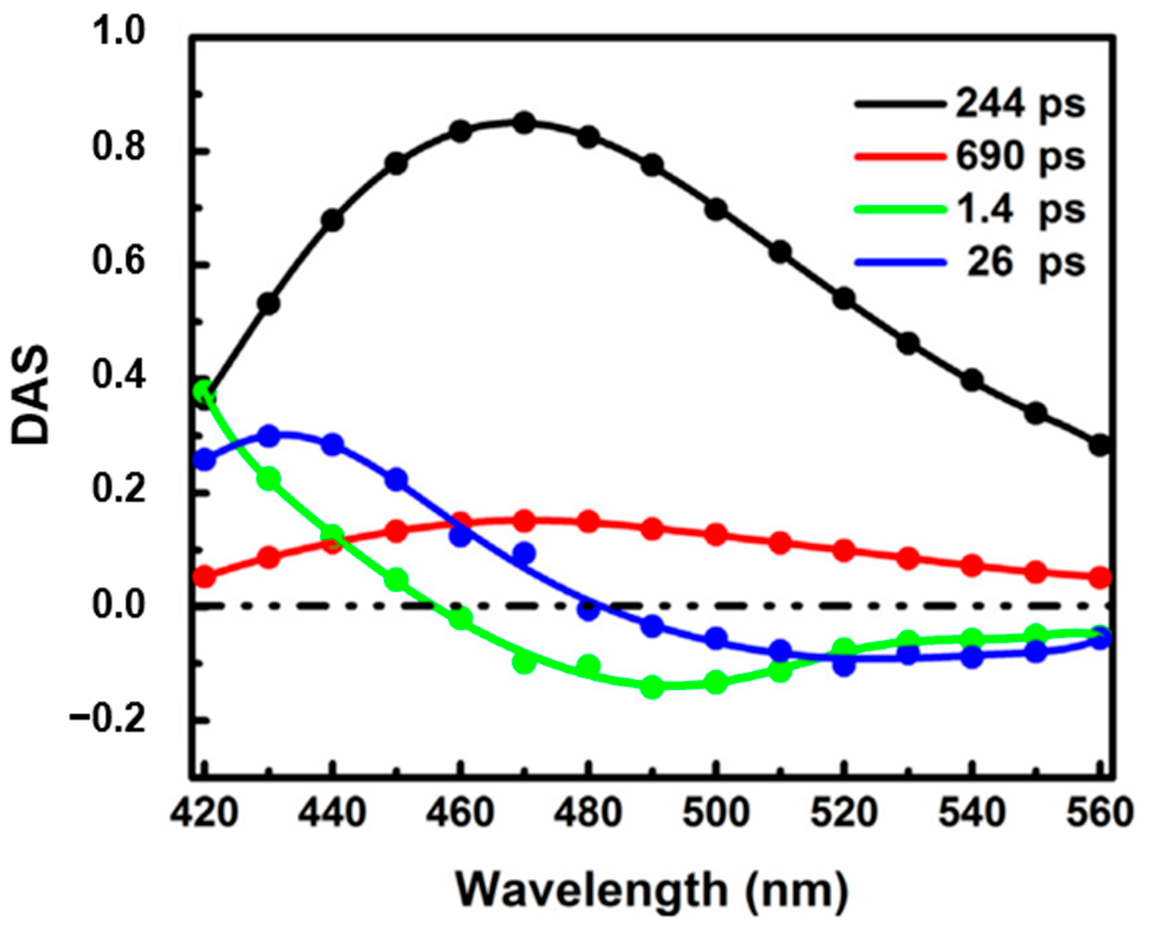
4.2. QSSQ in Free NADH
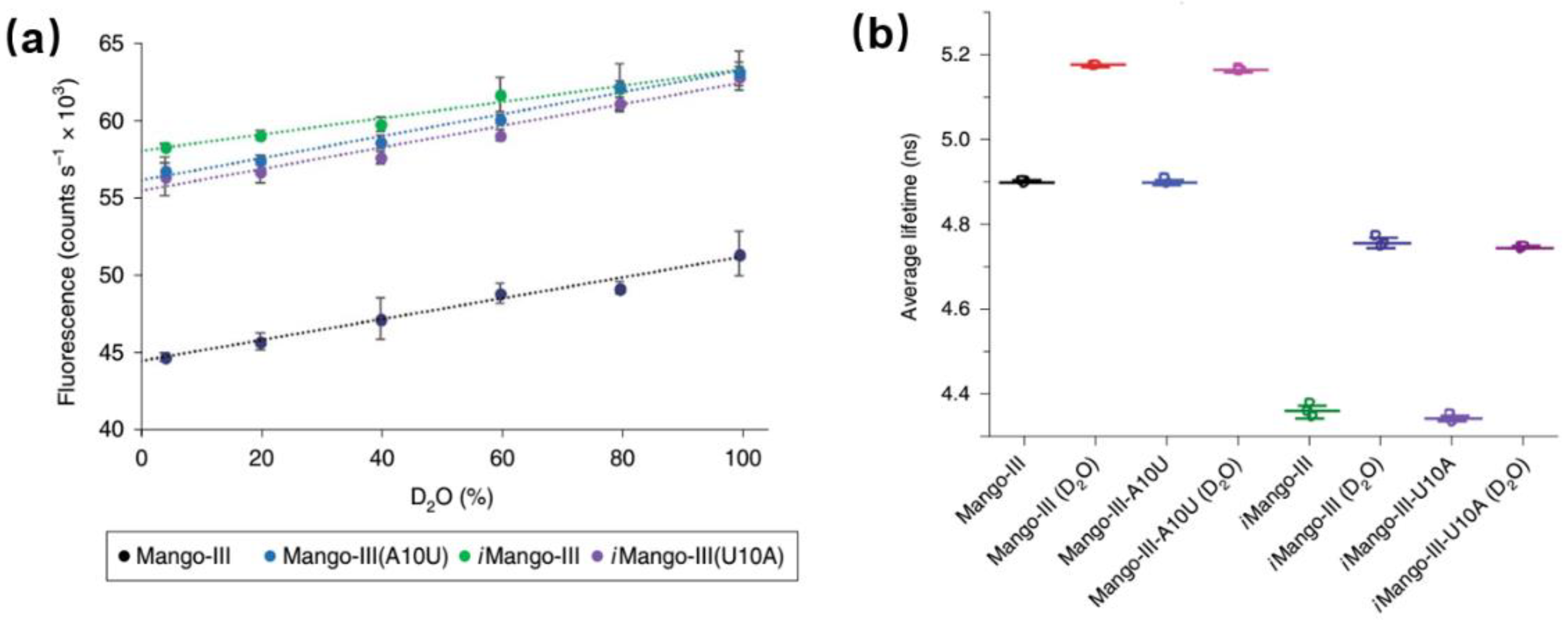
5. Application of RNA “Turn-On” Aptamers in Live Cell Imaging
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Das, A.; Sharma, G.; Kamatham, N.; Prabhakar, R.; Sen, P.; Ramamurthy, V. Ultrafast solvation dynamics reveal the octa acid capsule’s interior dryness depends on the guest. J. Phys. Chem. A 2019, 123, 5928–5936. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Santra, S.; Yesylevskyy, S.; Maiti, J.; Jana, M.; Das, R. Picosecond solvation dynamics in nanoconfinement: Role of water and host-guest complexation. J. Phys. Chem. B 2018, 122, 3996–4005. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Chaterjee, S.; Halder, R.; Jana, B.; Singh, P.C. Role of dispersive fluorous interaction in the solvation dynamics of the perfluoro group containing molecules. J. Phys. Chem. B 2017, 121, 7681–7688. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Knutson, J.R. Ultrafast fluorescence spectroscopy via upconversion applications to biophysics. Methods Enzymol. 2008, 450, 159–183. [Google Scholar]
- Qin, Y.; Jia, M.; Yang, J.; Wang, D.; Wang, L.; Xu, J.; Zhong, D. Molecular origin of ultrafast water–protein coupled interactions. J. Phys. Chem. Lett. 2016, 7, 4171–4177. [Google Scholar] [CrossRef]
- Zhang, L.; Kao, Y.-T.; Qiu, W.; Wang, L.; Zhong, D. Femtosecond studies of tryptophan fluorescence dynamics in proteins: Local solvation and electronic quenching. J. Phys. Chem. B 2006, 110, 18097–18103. [Google Scholar] [CrossRef]
- Qiu, W.; Zhang, L.; Okobiah, O.; Yang, Y.; Wang, L.; Zhong, D.; Zewail, A.H. Ultrafast solvation dynamics of human serum albumin: Correlations with conformational transitions and site-selected recognition. J. Phys. Chem. B 2006, 110, 10540–10549. [Google Scholar] [CrossRef]
- Qiu, W.; Kao, Y.-T.; Zhang, L.; Yang, Y.; Wang, L.; Stites, W.E.; Zhong, D.; Zewail, A.H. Protein surface hydration mapped by site-specific mutations. Proc. Natl. Acad. Sci. USA 2006, 103, 13979–13984. [Google Scholar] [CrossRef]
- Lu, W.; Kim, J.; Qiu, W.; Zhong, D. Femtosecond studies of tryptophan solvation: Correlation function and water dynamics at lipid surfaces. Chem. Phys. Lett. 2004, 388, 120–126. [Google Scholar] [CrossRef]
- Bhattacharyya, S.M.; Wang, Z.-G.; Zewail, A.H. Dynamics of water near a protein surface. J. Phys. Chem. B 2003, 107, 13218–13228. [Google Scholar] [CrossRef]
- Peon, J.; Pal, S.K.; Zewail, A.H. Hydration at the surface of the protein monellin: Dynamics with femtosecond resolution. Proc. Natl. Acad. Sci. USA 2002, 99, 10964–10969. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.K.; Peon, J.; Bagchi, B.; Zewail, A.H. Biological water: Femtosecond dynamics of macromolecular hydration. J. Phys. Chem. B 2002, 106, 12376–12395. [Google Scholar] [CrossRef]
- Xu, J.; Knutson, J.R. Quasi-static self-quenching of Trp-X and X-Trp dipeptides in water: Ultrafast fluorescence decay. J. Phys. Chem. B 2009, 113, 12084–12089. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Chen, J.; Toptygin, D.; Tcherkasskaya, O.; Callis, P.; King, J.; Brand, L.; Knutson, J.R. Femtosecond fluorescence spectra of tryptophan in human gamma-crystallin mutants: Site-dependent ultrafast quenching. J. Am. Chem. Soc. 2009, 131, 16751–16757. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Toptygin, D.; Graver, K.J.; Albertini, R.A.; Savtchenko, R.S.; Meadow, N.D.; Roseman, S.; Callis, P.R.; Brand, L.; Knutson, J.R. Ultrafast fluorescence dynamics of tryptophan in the proteins monellin and IIAGlc. J. Am. Chem. Soc. 2006, 128, 1214–1221. [Google Scholar] [CrossRef]
- Cao, S.; Zhou, Z.; Li, H.; Jia, M.; Liu, Y.; Wang, M.; Zhang, M.; Zhang, S.; Chen, J.; Xu, J.; et al. A fraction of NADH in solution is “dark”: Implications for metabolic sensing via fluorescence lifetime. Chem. Phys. Lett. 2019, 726, 18–21. [Google Scholar] [CrossRef]
- Cao, S.; Li, H.; Liu, Y.; Zhang, M.; Wang, M.; Zhou, Z.; Chen, J.; Zhang, S.; Xu, J.; Knutson, J.R. Femtosecond fluorescence spectra of NADH in solution: Ultrafast solvation dynamics. J. Phys. Chem. B 2020, 124, 771–776. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, D.; Su, N.; Bao, B.; Xie, X.; Zuo, F.; Yang, L.; Wang, H.; Jiang, L.; Lin, Q.; et al. Visualizing RNA dynamics in live cells with bright and stable fluorescent RNAs. Nat. Biotechnol. 2019, 37, 1287–1293. [Google Scholar] [CrossRef]
- Babendure, J.R.; Adams, S.R.; Tsien, R.Y. Aptamers switch on fluorescence of triphenylmethane dyes. J. Am. Chem. Soc. 2003, 125, 14716–14717. [Google Scholar] [CrossRef]
- Paige, J.S.; Wu, K.Y.; Jaffrey, S.R. RNA mimics of green fluorescent protein. Science 2011, 333, 642–646. [Google Scholar] [CrossRef]
- Dolgosheina, E.V.; Jeng, S.C.Y.; Panchapakesan, S.S.S.; Cojocaru, R.; Chen, P.S.K.; Wilson, P.D.; Hawkins, N.; Wiggins, P.A.; Unrau, P.J. RNA mango aptamer-fluorophore: A bright, high-affinity complex for RNA labeling and tracking. ACS Chem. Biol. 2014, 9, 2412–2420. [Google Scholar] [CrossRef] [PubMed]
- Gustavsson, T.; Baldacchino, G.; Mialocq, J.-C.; Pommeret, M.S. A femtosecond fluorescence up-conversion study of the dynamic Stokes shift of the DCM dye molecule in polar and non-polar solvents. Chem. Phys. Lett. 1995, 236, 587–594. [Google Scholar] [CrossRef]
- Cannizzo, A.; Bram, O.; Zgrablic, G.; Tortschanoff, A.; Oskouei, A.A.; Mourik, F.V.; Chergui, M. Femtosecond fluorescence upconversion setup with broadband detection in the ultraviolet. Opt. Lett. 2007, 32, 3555–3557. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-X.; Würth, C.; Zhao, L.; Resch-Genger, U.; Ernsting, N.P.; Sajadi, M. Femtosecond broadband fluorescence upconversion spectroscopy: Improved setup and photometric correction. Rev. Sci. Instrum. 2011, 82, 063108. [Google Scholar] [CrossRef]
- Schanz, R.; Kovalenko, S.A.; Kharlanov, V.; Ernsting, N.P. Broad-band fluorescence upconversion for femtosecond spectroscopy. Appl. Phys. Lett. 2001, 79, 566–568. [Google Scholar] [CrossRef]
- Gerecke, M.; Bierhance, G.; Gutmann, M.; Ernsting, N.P.; Rosspeintner, A. Femtosecond broadband fluorescence upconversion spectroscopy: Spectral coverage versus efficiency. Rev. Sci. Instrum. 2016, 87, 053115. [Google Scholar] [CrossRef]
- Becker, W.; Studier, H.; Wetzker, C. Ultra-fast HPM detectors improve NAD(P)H FLIM. In Proceedings of the Multiphoton Microscopy in the Biomedical Sciences XVIII, San Francisco, CA, USA, 28–30 January 2018. [Google Scholar] [CrossRef]
- Ruggiero, A.J.; Todd, D.C.; Fleming, G.R. Subpicosecond fluorescence anisotropy studies of tryptophan in water. J. Am. Chem. Soc. 1990, 112, 1003–1014. [Google Scholar] [CrossRef]
- Shen, X.; Knutson, J.R. Subpicosecond fluorescence spectra of tryptophan in water. J. Phys. Chem. B 2001, 105, 6260–6265. [Google Scholar] [CrossRef]
- Grant, E.H.; Sheppard, R.J.; South, G.P. Dielectric behavior of biological molecules in solution. In Monographs on Physical Biochemistry; Clarendon: Oxford, UK, 1978. [Google Scholar]
- Otting, G.; Liepinsh, E.; Wuthrich, K. Protein hydration in aqueous solution. Science 1991, 254, 974–980. [Google Scholar] [CrossRef]
- Denisov, V.P.; Halle, B. Protein hydration dynamics in aqueous solution. Faraday Discuss. 1996, 103, 227–244. [Google Scholar] [CrossRef] [PubMed]
- Denisov, V.P.; Jonsson, B.-H.; Halle, B. Hydration of denatured and molten globule proteins. Nat. Struct. Biol. 1999, 6, 253–260. [Google Scholar] [PubMed]
- Yang, J.; Zhang, L.; Wang, L.; Zhong, D. Femtosecond conical intersection dynamics of tryptophan in proteins and validation of slowdown of hydration layer dynamics. J. Am. Chem. Soc. 2012, 134, 16460. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Zhang, L.; Kao, Y.-T.; Lu, W.; Li, T.; Kim, J.; Sollenberger, G.M.; Wang, L.; Zhong, D. Ultrafast hydration dynamics in melittin folding and aggregation: Helix formation and tetramer self-assembly. J. Phys. Chem. B 2005, 109, 16901–16910. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Hassanali, A.A.; Kao, Y.-T.; Zhong, D.; Singer, S.J. Hydration dynamics and time scales of coupled water-protein fluctuations. J. Am. Chem. Soc. 2007, 129, 3376–3382. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Yang, J.; Qin, Y.; Wang, D.; Pan, H.; Wang, L.; Xu, J.; Zhong, D. Determination of protein surface hydration by systematic charge mutations. J. Phys. Chem. Lett. 2015, 6, 5100–5105. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.F.; Knutson, J.R.; Ziffer, H.; Porter, D. Fluorescence of tryptophan dipeptides: Correlations with the rotamer model. Biochemistry 1991, 30, 5184–5195. [Google Scholar] [CrossRef]
- Chen, J.; Flaugh, S.L.; Callis, P.R.; King, J. Mechanism of the highly efficient quenching of tryptophan fluorescence in human γD-crystallin. Biochemistry 2006, 45, 11552–11563. [Google Scholar] [CrossRef]
- Chen, J.; Toptygin, D.; Brand, L.; King, J. Mechanism of the efficient tryptophan fluorescence quenching in human γd-crystallin studied by time-resolved fluorescence. Biochemistry 2008, 47, 10705–10721. [Google Scholar] [CrossRef]
- Schaefer, P.M.; Kalinina, S.; Rueck, A.; Arnim, C.A.F.V.; Einem, B.V. NADH Autofluorescence—A marker on its way to boost bioenergetic research. Cytom. Part A 2019, 95, 34–46. [Google Scholar] [CrossRef]
- Rück, A.; Hauser, C.; Mosch, S.; Kalinina, S. Spectrally resolved fluorescence lifetime imaging to investigate cell metabolism in malignant and nonmalignant oral mucosa cells. J. Biomed. Opt. 2014, 19, 96005. [Google Scholar] [CrossRef] [PubMed]
- Drozdowicztomsia-Tomsia, K.; Anwer, A.G.; Cahill, M.A.; Madlum, K.N.; Maki, A.M.; Baker, M.S.; Goldys, E.M. Multiphoton fluorescence lifetime imaging microscopy reveals free-to-bound NADH ratio changes associated with metabolic inhibition. J. Biomed. Opt. 2014, 19, 086016. [Google Scholar] [CrossRef] [PubMed]
- Torno, K.; Wright, B.K.; Jones, M.R.; Digman, M.A.; Gratton, E.; Phillips, M. Real-time analysis of metabolic activity within Lactobacillus acidophilus by phasor fluorescence lifetime imaging microscopy of NADH. Curr. Microbiol. 2013, 66, 365–367. [Google Scholar] [CrossRef] [PubMed]
- Visser, A.J.W.G.; Hoek, A.V. The fluorescence decay of reduced nicotinamides in aqueous solution after excitation with a UV-mode locked Ar ion laser. Photochem. Photobiol. 1981, 33, 35–40. [Google Scholar] [CrossRef]
- Hull, R.V.; Conger, P.S., III; Hoobler, R.J. Conformation of NADH studied by fluorescence excitation transfer spectroscopy. Biophys. Chem. 2001, 90, 9–16. [Google Scholar] [CrossRef]
- Yang, X.; Bi, S.; Yang, L.; Zhu, Y.; Wang, X. Multi-NMR and fluorescence spectra study the effects of aluminum (III) on coenzyme NADH in aqueous solutions. Spectrochim. Acta Part A 2003, 59, 2561–2569. [Google Scholar] [CrossRef]
- Formoso, E.; Mujika, J.I.; Grabowski, S.J.; Lopez, X. Aluminum and its effect in the equilibrium between folded/unfolded conformation of NADH. J. Inorg. Biochem. 2015, 152, 139–146. [Google Scholar] [CrossRef]
- Heiner, Z.; Roland, T.; Leonard, J.; Haacke, S.; Groma, G.I. Kinetics of light-induced intramolecular energy transfer in different conformational states of NADH. J. Phys. Chem. B 2017, 92, 331. [Google Scholar] [CrossRef]
- Blacker, T.S.; Nicolaou, N.; Duchen, M.R.; Bain, A.J. Polarised two-photon absorption and heterogeneous fluorescence dynamics in NAD (P) H. J. Phys. Chem. B 2019, 123, 4705–4717. [Google Scholar] [CrossRef]
- Boldridge, D.W.; Morton, T.H.; Scott, G.W.; Clark, J.H.; Philips, L.A.; Webb, S.P.; Yeh, S.M.; Eikeren, P.V. Excited-state dynamics of NADH and 1-N-Propyl-1,4 Dihydronicotinamide. In Ultrafast Phenomena IV; Springer: Berlin/Heidelberg, Germany, 1984. [Google Scholar]
- Xu, J.; Chen, B.; Callis, P.; Muino, P.L.; Rozeboom, H.; Broos, J.; Toptygin, D.; Brand, L.; Knutson, J.R. Picosecond fluorescence dynamics of tryptophan and 5-fluorotryptophan in monellin: Slow water-protein relaxation unmasked. J. Phys. Chem. B 2019, 119, 4230–4239. [Google Scholar] [CrossRef]
- Neubacher, S.; Hennig, S. RNA structure and cellular applications of fluorescent light-up aptamers. Angew. Chem. Int. Ed. 2019, 58, 1266–1279. [Google Scholar] [CrossRef] [PubMed]
- Autour, A.; Jeng, S.C.Y.; Cawte, A.D.; Abdolahzadeh, A.; Galli, A.; Panchapakesan, S.S.S.; Rueda, D.; Ryckelynck, M.; Unrau, P.J. Fluorogenic RNA Mango aptamers for imaging small non-coding RNAs in mammalian cells. Nat. Commun. 2018, 9, 656. [Google Scholar] [CrossRef] [PubMed]
- Trachman, R.J., III; Autour, A.; Jeng, S.C.Y.; Abdolahzadeh, A.; Andreoni, A.; Cojocaru, R.; Garipov, R.; Dolgosheina, E.V.; Knutson, J.R.; Ryckelynck, M.; et al. Structure and functional reselection of the Mango-III fluorogenic RNA aptamer. Nat. Chem. Biol. 2019, 15, 472. [Google Scholar] [CrossRef] [PubMed]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy, 3rd ed.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1999. [Google Scholar]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, S.; Li, H.; Zhao, Z.; Zhang, S.; Chen, J.; Xu, J.; Knutson, J.R.; Brand, L. Ultrafast Fluorescence Spectroscopy via Upconversion and Its Applications in Biophysics. Molecules 2021, 26, 211. https://doi.org/10.3390/molecules26010211
Cao S, Li H, Zhao Z, Zhang S, Chen J, Xu J, Knutson JR, Brand L. Ultrafast Fluorescence Spectroscopy via Upconversion and Its Applications in Biophysics. Molecules. 2021; 26(1):211. https://doi.org/10.3390/molecules26010211
Chicago/Turabian StyleCao, Simin, Haoyang Li, Zenan Zhao, Sanjun Zhang, Jinquan Chen, Jianhua Xu, Jay R. Knutson, and Ludwig Brand. 2021. "Ultrafast Fluorescence Spectroscopy via Upconversion and Its Applications in Biophysics" Molecules 26, no. 1: 211. https://doi.org/10.3390/molecules26010211
APA StyleCao, S., Li, H., Zhao, Z., Zhang, S., Chen, J., Xu, J., Knutson, J. R., & Brand, L. (2021). Ultrafast Fluorescence Spectroscopy via Upconversion and Its Applications in Biophysics. Molecules, 26(1), 211. https://doi.org/10.3390/molecules26010211