Butyrate Stimulates Histone H3 Acetylation, 8-Isoprostane Production, RANKL Expression, and Regulated Osteoprotegerin Expression/Secretion in MG-63 Osteoblastic Cells


Abstract
1. Introduction
2. Results
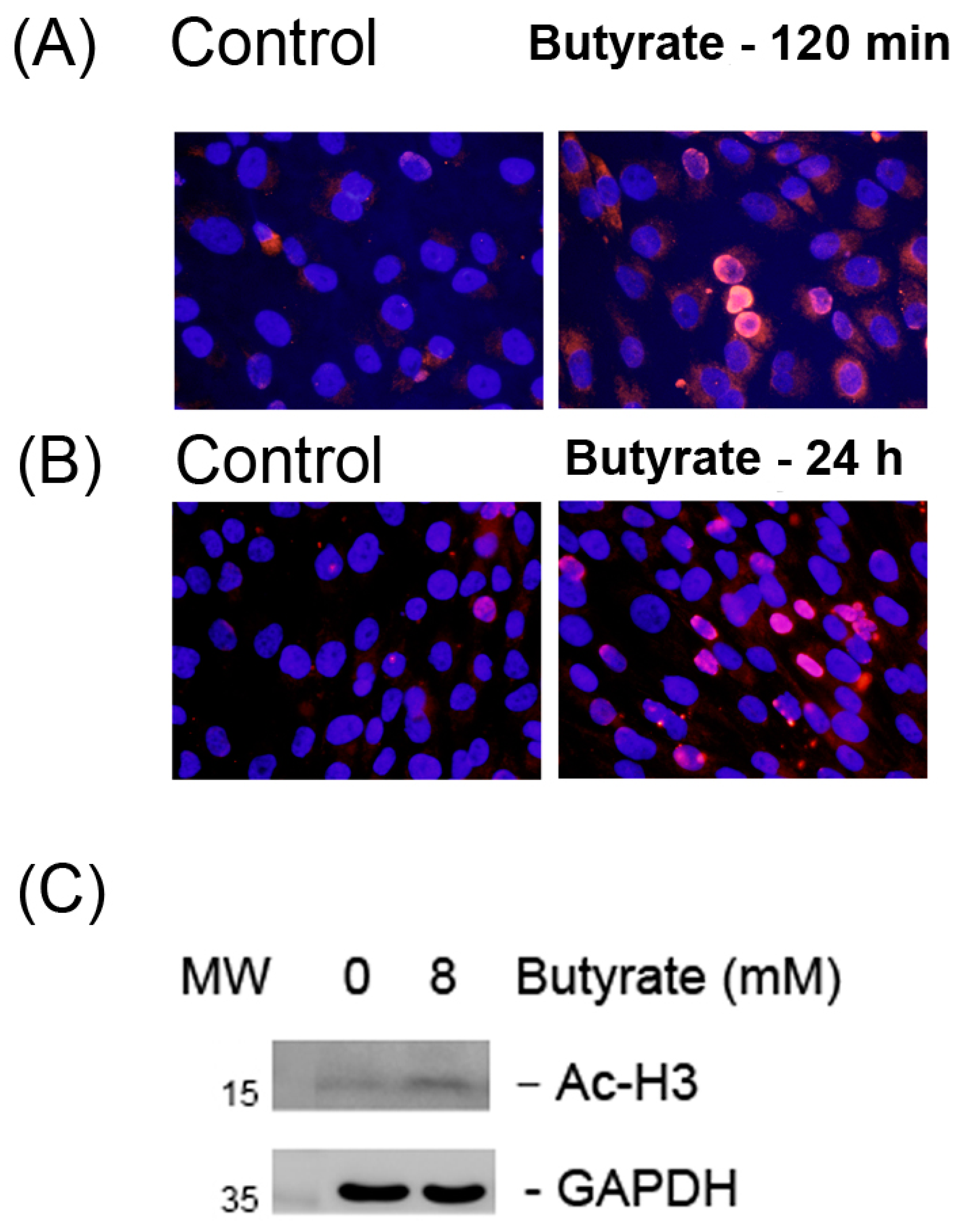
2.1. Stimulation of Histone H3 Acetylation by Butyrate
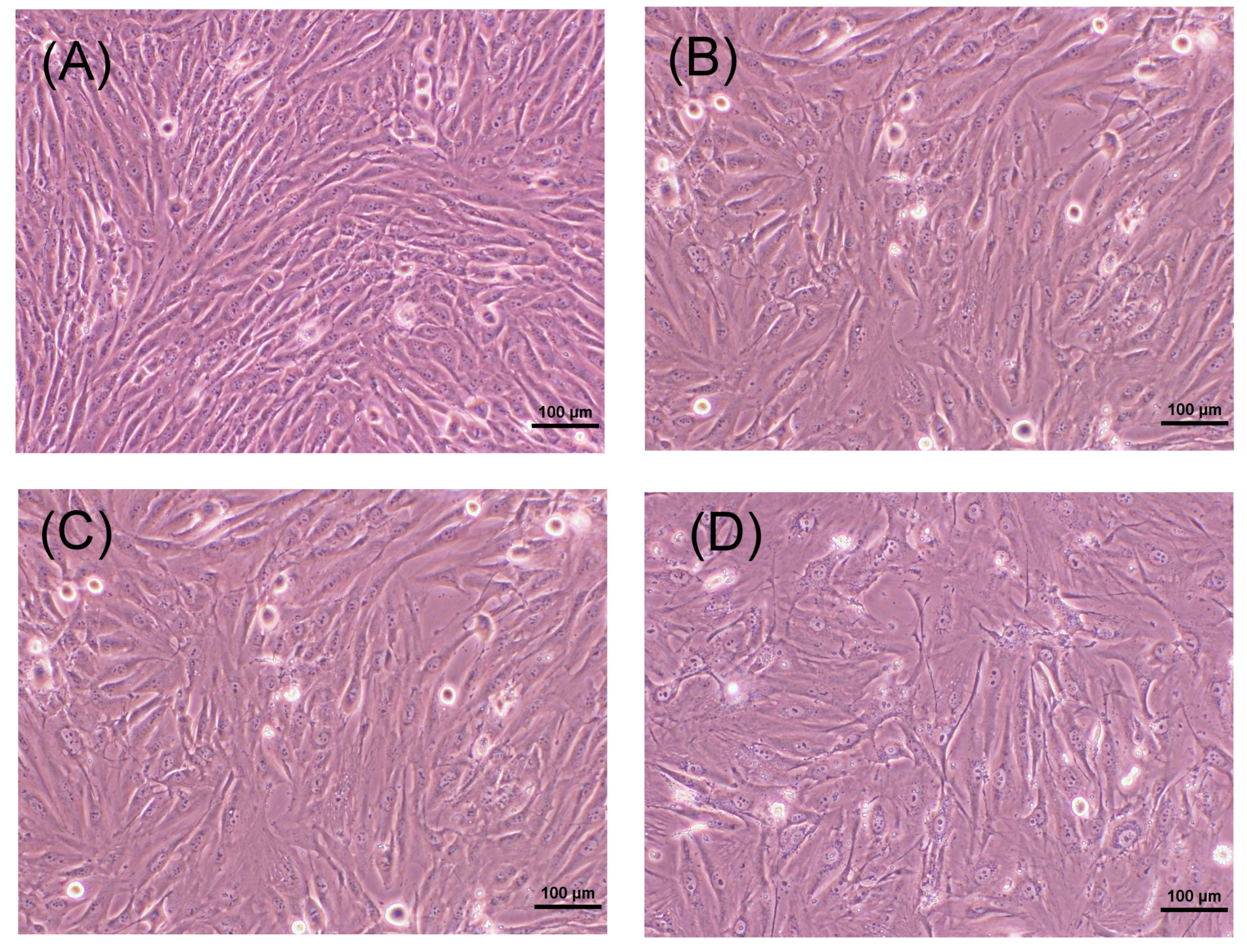
2.2. Morphology of MG-63 Cells after Exposure to Butyrate for Three Days
2.3. Effect of Butyrate on the Growth and Cell Viability of MG-63 Cells
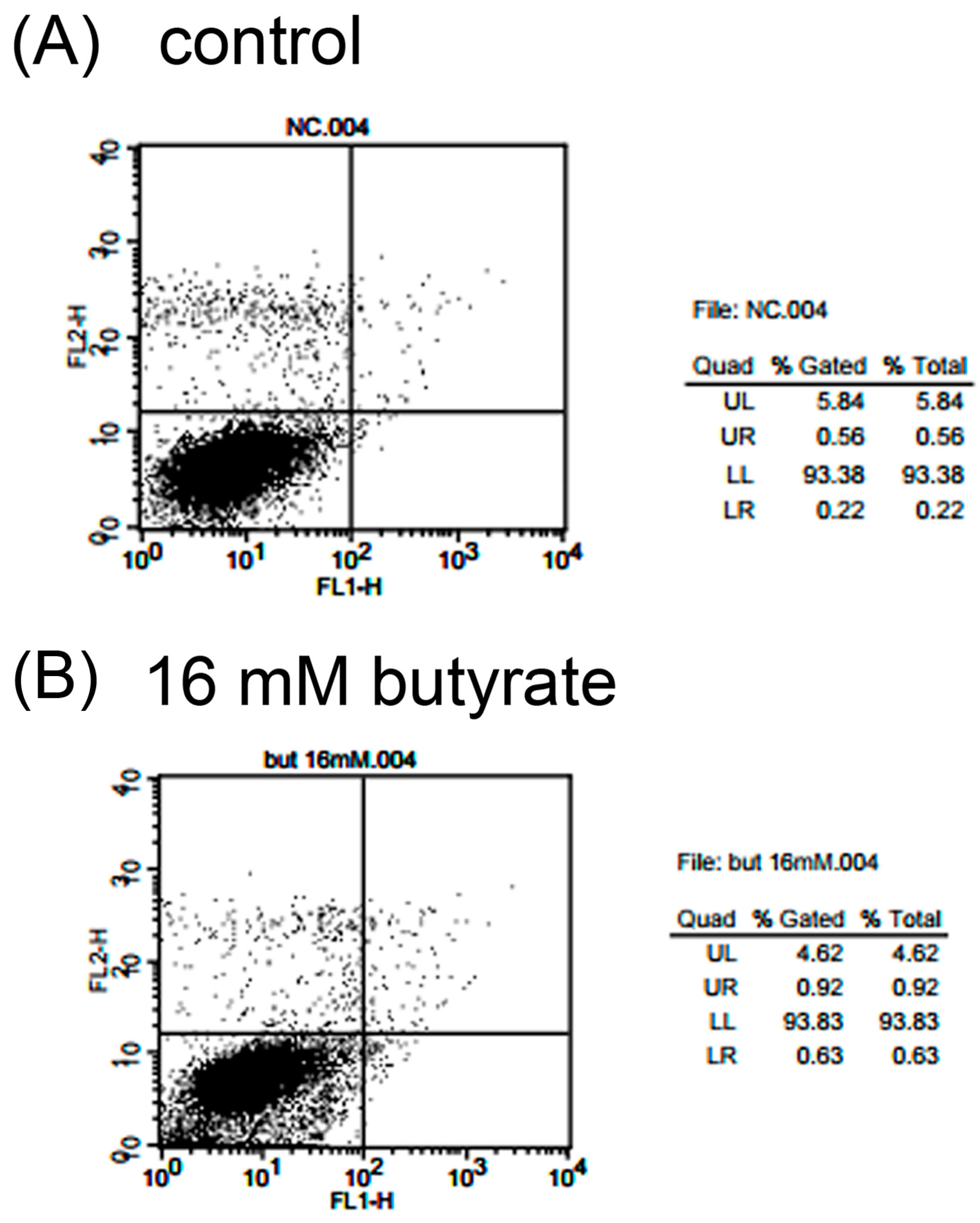
2.4. Effect of Butyrate on the Apoptosis and Necrosis of MG-63 Cells
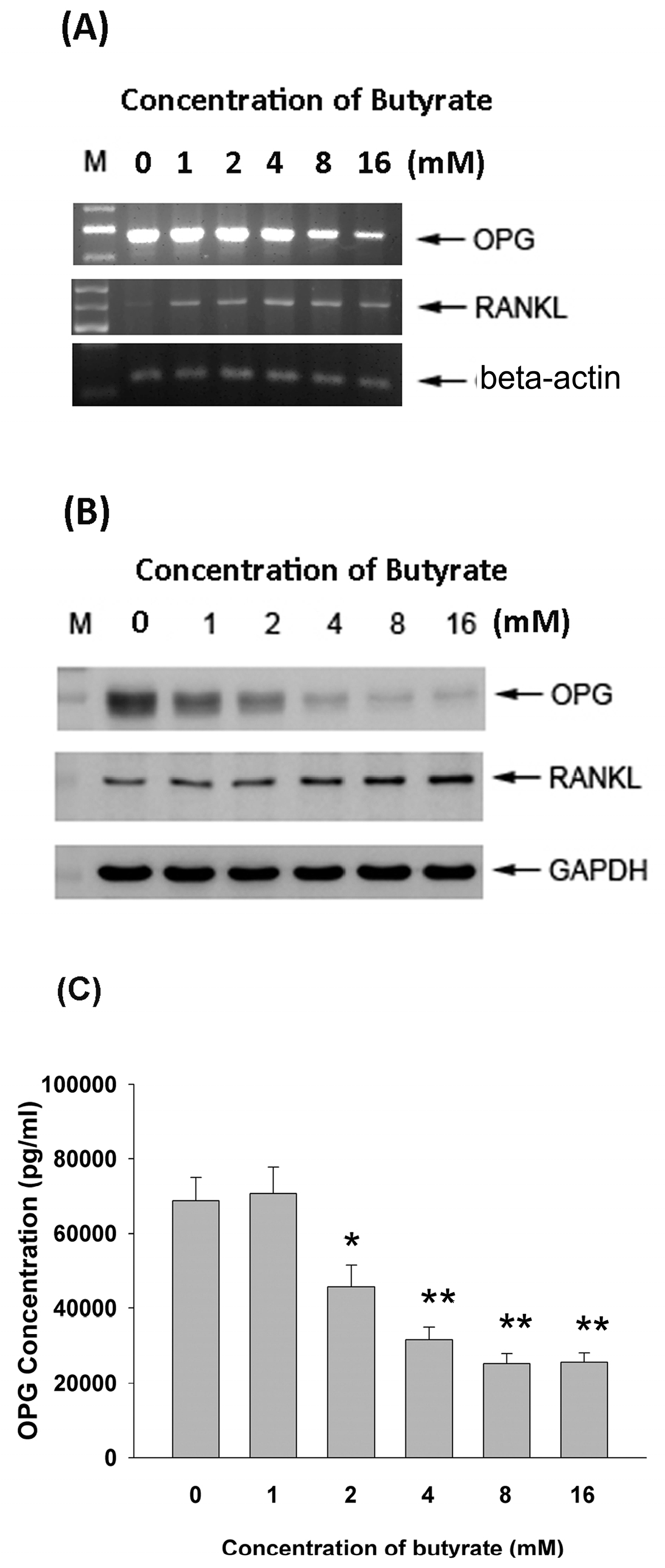
2.5. Effect of Butyrate on OPG and RANKL mRNA and Protein Expression
2.6. Effect of Butyrate on OPG and RANKL Secretion of MG-63 Cells
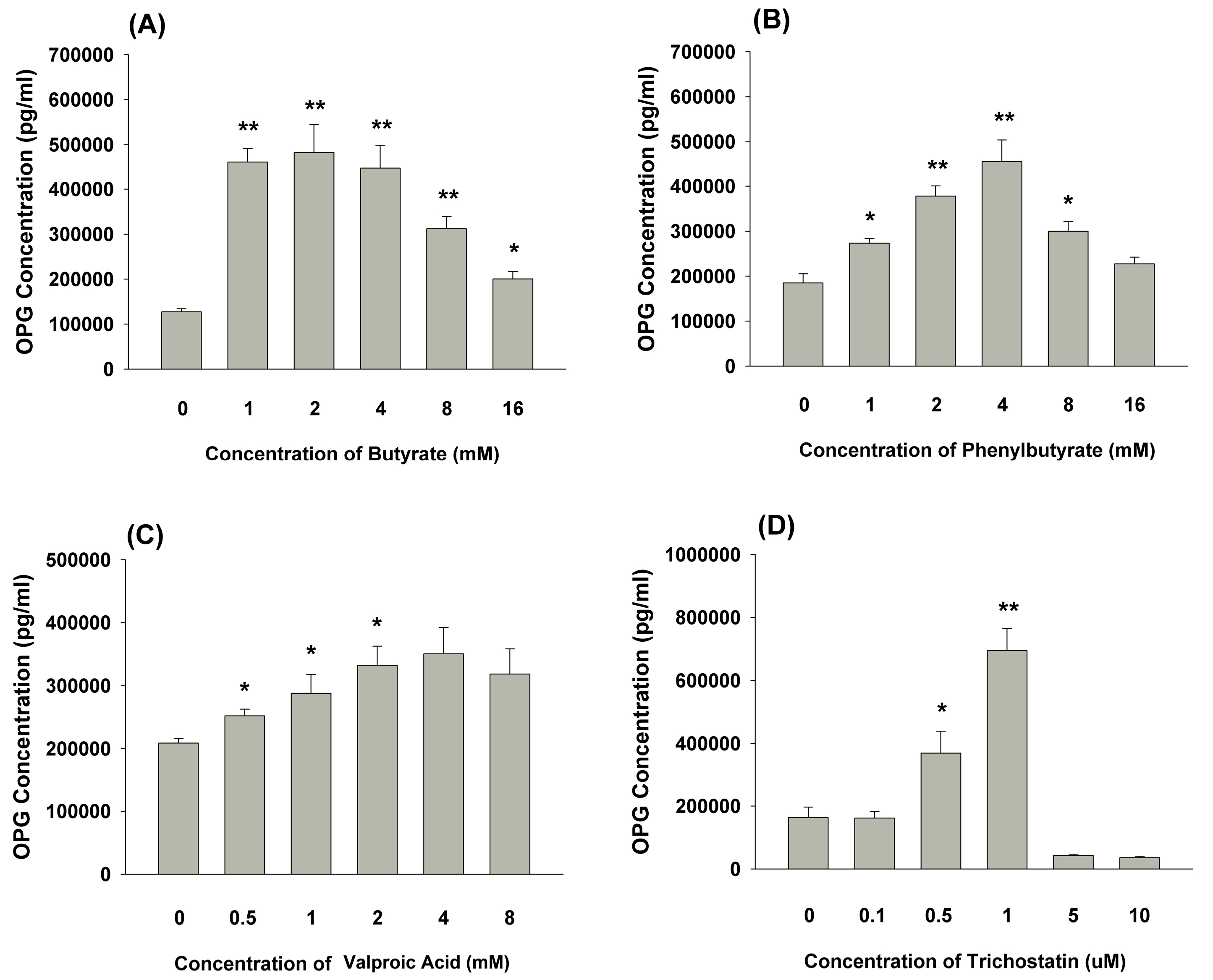
2.7. Effect of 3 days of Exposure of MG-63 Cells to Butyrate or other HDAC Inhibitors on OPG Secretion
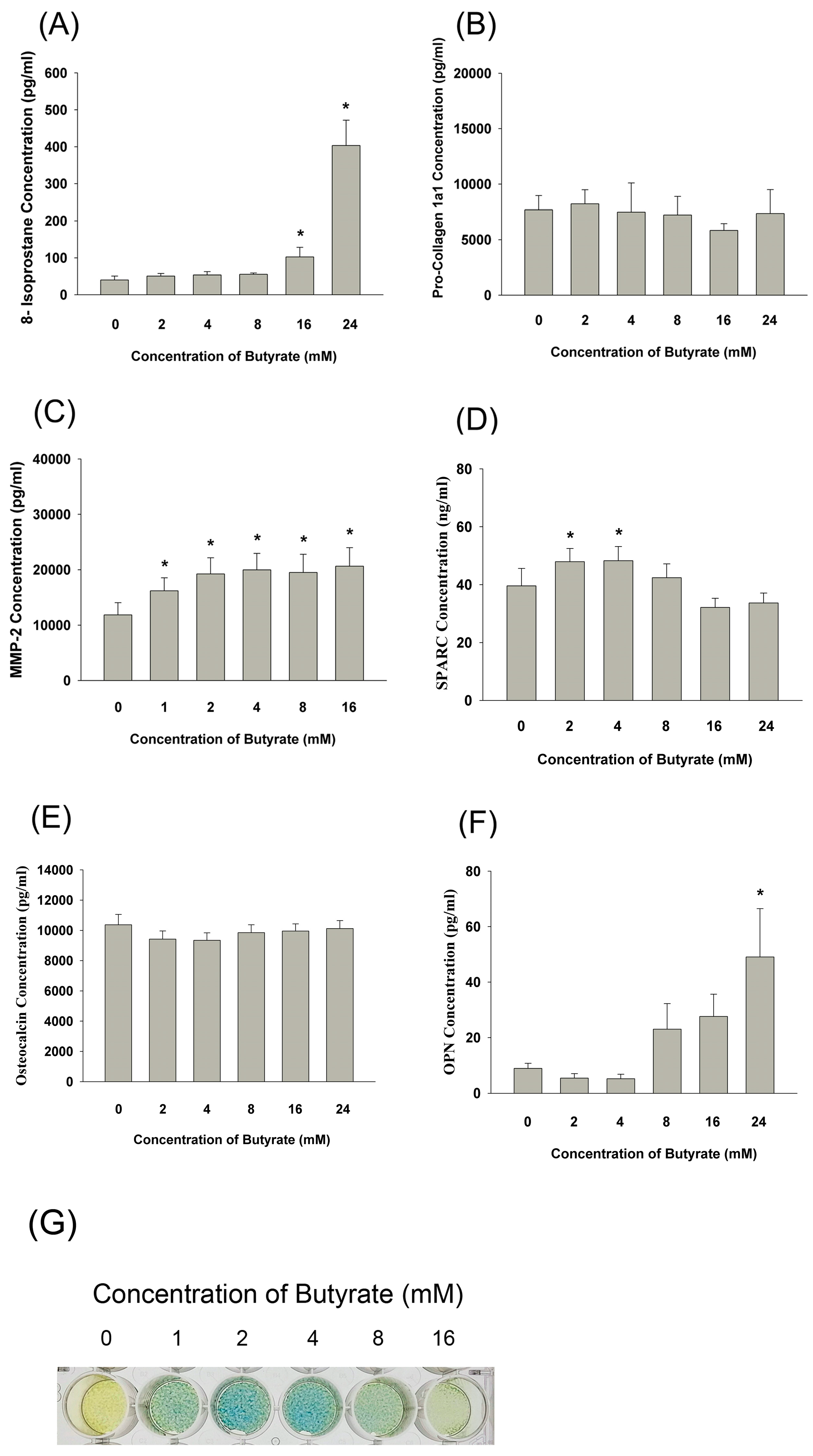
2.8. Effect of Butyrate on 8-Isoprostane, Pro-Collagen I, MMP-2, Osteopontin, Osteonectin, and Osteocalcin Secretion as well as the ALP Activity of MG-63 Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Immunofluorescent Staining of Ac-H3 Expression
4.3. Effect of Butyrate on the Proliferation and Viability of MG-63 Cells
4.4. Inducing the Apoptosis and Necrosis of MG-63 Cells by Butyrate: PI and Annexin V Flow Cytometric Analysis
4.5. Effect of Butyrate on the mRNA Expression of OPG, RANKL of Cultured MG-63 Cells
4.6. Effects of Butyrate on OPG, RANKL and Ac-H3 Protein Expression of MG-63 Cells
4.7. Effect of Butyrate on OPG, RANKL, 8-Isoprostane, Pro-Collagen I, MMP-2, Osteopontin, Osteonectin and Osteocalcin Secretion and the ALP activity of MG-63 Cells
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Yang, L.; Li, Z.; Li, X.; Wang, Z.; Wang, S.; Sasaki, Y.; Takai, H.; Ogata, Y. Butyric acid stimulates bone sialoprotein gene transcription. J. Oral Sci. 2010, 52, 231–237. [Google Scholar] [CrossRef]
- Tanner, A.; Maiden, M.F.; Macuch, P.J.; Murray, L.L.; Kent, R.L., Jr. Microbiota of health, gingivitis and initial periodontitis. J. Clin. Periodontol. 1998, 25, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Nishihara, T.; Koseki, T. Microbial etiology of periodontitis. Periodontol 2000 2004, 36, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Lamont, R.J.; Jenkinson, H.F. Life below the gum line: Pathogenic mechanisms of Porphyromonas gingivalis. Microbiol. Mol. Biol. Rev. 1998, 62, 1244–1263. [Google Scholar] [PubMed]
- Holt, S.C.; Kesavalu, L.; Walker, S.; Genco, C.A. Virulence factors of Porphyromonas gingivalis. Periodontol 2000 1999, 20, 168–238. [Google Scholar] [CrossRef] [PubMed]
- Isogai, H.; Isogai, E.; Yoshimura, F.; Suzuki, T.; Kagota, W.; Takano, K. Specific inhibition of adherence of an oral strain of Bacteroides gingivalis 381 to epithelial cells by monoclonal antibodies against the bacterial fimbriae. Arch. Oral Biol. 1988, 33, 479–485. [Google Scholar] [CrossRef]
- Njoroge, T.; Genco, R.J.; Sojar, H.T.; Hamada, N.; Genco, C.A. A role for fimbriae in Porphyromonas gingivalis invasion of oral epithelial cells. Infect. Immun. 1997, 65, 1980–1984. [Google Scholar]
- Wilson, M.; Reddi, K.; Henderson, B. Cytokine-inducing components of periodontopathogenic bacteria. J. Periodont. Res. 1996, 31, 393–407. [Google Scholar] [CrossRef] [PubMed]
- Pollanen, M.T.; Overman, D.O.; Salonen, J.I. Bacterial metabolites sodium butyrate and proprionate inhibit epithelial cell growth in vitro. J. Periodontal Res. 1997, 32, 326–334. [Google Scholar] [CrossRef]
- Niederman, R.; Buyle-Bodin, Y.; Lu, B.Y.; Robinson, P.; Naleway, C. Short-chain carboxylic acid concentration in human gingival crevicular fluid. J. Dent. Res. 1997, 76, 575–579. [Google Scholar] [CrossRef]
- Provenzano, J.C.; Rocas, I.N.; Tavares, L.F.; Neves, B.C.; Sigueura, J.F. Short-chain fatty acids in infected root canals of teeth with apical periodontitis before and after treatment. J. Endod. 2015, 41, 831–835. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.C.; Tsai, Y.L.; Chen, Y.W.; Chan, C.P.; Huang, C.F.; Lan, W.C.; Lin, C.C.; Lan, W.H.; Jeng, J.H. Butyrate induces reactive oxygen species production and affects cell cycle progression in human gingival fibroblasts. J. Periodontal Res. 2013, 48, 66–73. [Google Scholar] [CrossRef]
- Morozumi, A. High concentration of sodium butyrate suppresses osteoblastic differentiation and mineralized nodule formation in ROS17/2.8 cells. J. Oral Sci. 2011, 53, 509–516. [Google Scholar] [CrossRef]
- Niederman, R.; Zhang, J.; Kashket, S. Short-chain carboxylic-acid-stimulated, PMN-mediated gingival inflammation. Crit. Rev. Oral Biol. Med. 1997, 8, 269–290. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Z.; Duan, N.; Zhu, G.; Schwarz, E.M.; Xie, C. Osteoblast-osteoclast interactions. Connect. Tissue Res. 2018, 59, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Barreiros, D.; Pucinelli, C.M.; Oliveira, K.M.H.; Paula-Silva, F.W.G.; Nelson Filho, P.; Silva, L.A.B.D.; Kuchler, E.C.; Silva, B.A.B.D. Immunohistochemical and mRNA expression of RANK, RANKL, OPG, TLR2 and MyD88 during apical periodontitis progression in mice. J. Appl. Oral Sci. 2018, 26, e20170512. [Google Scholar] [CrossRef] [PubMed]
- Belibasakis, G.N.; Rechenberg, D.K.; Zehnder, M. The receptor activator of NF-κB ligand-osteoprotegerin system in pulpal and periapical disease. Int. Endod. J. 2013, 46, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Belibasakis, G.N.; Bostanci, N. The RANKL-OPG system in clinical periodontology. J. Clin. Periodontol. 2012, 39, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Reddi, D.; Brown, S.J.; Belibasakis, G.N. Porphyromonas gingivalis induces RANKL in bone marrow stromal cells: Involvement of the p38 MAPK. Microb. Pathog. 2011, 51, 415–420. [Google Scholar] [CrossRef]
- Han, X.; Lin, X.; Yu, X.; Lin, J.; Kawai, T.; LaRosa, K.B.; Taubman, M.A. Porphyromonas gingivalis infection-associated periodontal bone resorption is dependent on receptor activator of NF-κB ligand. Infect. Immun. 2013, 81, 1502–1509. [Google Scholar] [CrossRef]
- Jung, I.H.; Lee, D.E.; Yun, J.H.; Cho, A.R.; Kim, C.S.; You, Y.J.; Kim, S.J.; Choi, S.H. Anti-inflammatory effect of (−)-epigallocatechin-3-gallate on Porphyromonas gingivalis lipopolysaccharide-stimulated fibroblasts and stem cells derived from human periodontal ligament. J. Periodontal Implant Sci. 2012, 42, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Jeng, J.H.; Chan, C.P.; Ho, Y.S.; Lan, W.H.; Hsieh, C.C.; Chang, M.C. Effects of butyrate and proprionate on the adhesion, growth, cell cycle kinetics and protein synthesis of cultured human gingival fibroblast. J. Periodontol. 1999, 70, 1435–1442. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.C.; Tsai, Y.L.; Liou, E.J.; Tang, C.M.; Wang, T.M.; Liu, H.C.; Liao, M.W.; Yeung, S.Y.; Chan, C.P.; Jeng, J.H. Effect of butyrate on collagen expression, cell viability, cell cycle progression and related proteins expression of MG-63 osteoblastic cells. PLoS ONE 2016, 11, e0165438. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Bai, J.; Ren, S.; Wang, R.; Zhang, L.; Zuo, Y. Sodium butyrate restores ASC expression and induces apoptosis in LS174T cells. Int. J. Mol. Med. 2012, 30, 1431–1437. [Google Scholar] [CrossRef] [PubMed]
- Tailor, D.; Hahm, E.R.; Kale, R.K.; Singh, S.V.; Singh, R.P. Sodium butyrate induces DRP1-mediated mitochondrial fusion and apoptosis in human colorectal cancer cells. Mitochondrion 2014, 16, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.C.; Chang, Y.C. Effects of a bacterial lipid byproduct on human pulp fibroblasts in vitro. J. Endod. 2007, 33, 437–441. [Google Scholar] [CrossRef] [PubMed]
- Bolognini, D.; Tobin, A.B.; Milligan, G.; Moss, C.E. The pharmacology and function of receptors for short-chain fatty acids. Mol. Pharmacol. 2016, 89, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Cushing, K.; Alvarado, D.M.; Ciorba, M.A. Butyrate and mucosa inflammation: New scientific evidence supports clinical observation. Clin. Transl. Gastroenterol. 2015, 6, e108. [Google Scholar] [CrossRef]
- Li, M.; van Esch, B.C.A.M.; Henricks, P.A.J.; Folkerts, G.; Garssen, J. The anti-inflammatory effects of short chain fatty acids on lipopolysaccharide- or tumor necrosis factor-α-stimulated endothelial cells via activation of GPR41/43 and inhibition of HDACs. Front. Pharmacol. 2018, 9, 533. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Y.; Wu, Y.; Ni, B.; Liang, Z. Sodium butyrate promotes the differentiation of rat bone marrow mesenchymal stem cells to smooth muscle cells through histone acetylation. PLoS ONE 2014, 9, e116183. [Google Scholar] [CrossRef]
- Chen, T.H.; Chen, W.H.; Hsu, K.H.; Kuo, C.D.; Hung, S.C. Sodium butyrate activates ERK to regulate differentiation of mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2007, 355, 913–918. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, T.M.; Westendorf, J.J. Histone deacetylase inhibitors promotes osteoblast maturation. J. Bone Miner. Res. 2005, 20, 2254–2263. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Suh, J.H.; Kim, A.Y.; Lee, Y.S.; Park, S.Y.; Kim, J.B. Histone deacetylase 1-mediated histone modification regulates osteoblast differentiation. Mol. Endocrinol. 2006, 20, 2432–2443. [Google Scholar] [CrossRef] [PubMed]
- Delanty, N.; Reilly, M.; Pratico, D.; FitzGerald, D.J.; Lawson, J.A.; FitzGerald, G.A. 8-Epi PGF2α: Specific analysis of an isoeicosanoid as an index of oxidant stress in vivo. Br. J. Clin. Pharmacol. 1996, 42, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Pratico, D.; Rokach, J.; Lawson, J.; FitzGerald, G.A. F2-isoprostanes as indices of lipid peroxidation in inflammatory diseases. Chem. Phys. Lipids 2004, 128, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Sottero, B.; Rossin, D.; Poli, G.; Biasi, F. Lipid oxidation products in the pathogenesis of inflammation-related gut disease. Curr. Med. Chem. 2018, 25, 1311–1326. [Google Scholar] [CrossRef] [PubMed]
- Musiek, E.S.; Morrow, J.D. F2-isoprostanes as markers of oxidant stress: An overview. Curr. Protoc. Toxicol. 2005, 24. [Google Scholar] [CrossRef]
- Huang, Y.; Zhu, M.; Li, Z.; Sa, R.; Chu, Q.; Zhang, Q.; Zhang, H.; Tang, W.; Zhang, M.; Yin, H. Mass spectrometry-based metabolomics profiling identifies alterations in salivary redox status and fatty acid metabolism in response to inflammation and oxidative stress in periodontal disease. Free Radic. Biol. Med. 2014, 70, 223–232. [Google Scholar] [CrossRef]
- Vengerfeldt, V.; Mandar, R.; Saag, M.; Piir, A.; Kullisaar, T. Oxidative stress in patients with endodontic pathologies. J. Pain Res. 2017, 10, 2031–3040. [Google Scholar] [CrossRef]
- Khocht, A.; Rogers, T.; Janal, M.N.; Brown, M. Gingival fluid inflammatory biomarkers and hypertension in African Americans. JDR Clin. Trans. Res. 2017, 2, 269–277. [Google Scholar] [CrossRef]
- Pradeep, A.R.; Rao, N.S.; Bajaj, P.; Agarwal, E. 8-Isoprostane: A lipid peroxidation product in gingival crevicular fluid in healthy, gingivitis and chronic periodontitis subjects. Arch. Oral Biol. 2013, 58, 500–504. [Google Scholar] [CrossRef] [PubMed]
- Parhami, F.; Morrow, A.D.; Balucan, J.; Leitinger, N.; Watson, A.D.; Tintut, Y.; Berliner, J.A.; Demer, L.L. Lipid oxidation products have opposite effects on calcifying vascular cell and bone cell differentiation: A possible explanation for the paradox of arterial calcification in osteoporotic patients. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Ma, E.; Ingram, K.H.; Milne, G.L.; Garvey, W.T. F2-Isoprostane reflect oxidative stress correlated with lean mass and bone density but not insulin resistance. J. Endocr. Soc. 2017, 1, 436–448. [Google Scholar] [CrossRef] [PubMed]
- Czerska, M.; Zielinski, M.; Gromadzinska, J. Isoprostanes—A novel major group of oxidative stress markers. Int. J. Occup. Med. Environ. Health 2016, 29, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Xu, J.K.; Figliomeni, L.; Huang, L.; Pavlos, N.J.; Rogers, M.; Tan, A.; Price, P.; Zheng, M.H. Expression of RANKL and OPG mRNA in periodontal disease: Possible involvement in bone destruction. Int. J. Mol. Med. 2003, 11, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Rechenberg, D.K.; Bostanci, N.; Zehnder, M.; Belibasakis, G.N. Periapical fluid RANKL and IL-8 are differentially regulated in pulpitis and apical periodontitis. Cytokine 2014, 69, 116–119. [Google Scholar] [CrossRef]
- Menezes, R.; Bramante, C.M.; Da Silva Paiva, K.B.; Letra, A.; Carneiro, E.; Fernando Zambuzzi, W.; Granjeiro, J.M. Receptor activator NF-κB-ligand and osteoprotegerin protein expression in human periapical cysts and granuloma. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2006, 102, 404–409. [Google Scholar] [CrossRef]
- Fan, R.; Sun, B.; Zhang, C.F.; Lu, Y.L.; Xuan, W.; Wang, Q.Q.; Yin, X.Z. Receptor activator of nuclear factor κB ligand and osteoprotegerin expression in chronic apical periodontitis: Possible association with inflammatory cells. Chin. Med. J. 2011, 124, 2162–2166. [Google Scholar]
- Katono, T.; Kawato, T.; Tanabe, N.; Suzuki, N.; Iida, T.; Morozumi, A.; Ochiai, K.; Maeno, M. Sodium butyrate stimulates mineralized nodule formation and osteoprotegerin expression by human osteoblasts. Arch. Oral Biol. 2008, 53, 903–909. [Google Scholar] [CrossRef]
- McBeath, R.; Pirone, D.M.; Nelson, C.M.; Bhadriraju, K.; Chen, C.S. Cell shape, cytoskeletal tension and RhoA regulate stem cell lineage commitment. Dev. Cell 2004, 6, 483–495. [Google Scholar] [CrossRef]
- Iwami, K.; Moriyama, T. Effects of short chain fatty acid, sodium butyrate, on osteoblastic and osteoclastic cells. Int. J. Biochem. 1993, 25, 1631–1635. [Google Scholar] [CrossRef]
- Kim, T.I.; Han, J.E.; Jung, H.M.; Oh, J.H.; Woo, K.M. Analysis of histone deacetylase inhibitor-induced response in human periodontal ligament fibroblasts. Biotechnol. Lett. 2013, 35, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Iida, T.; Kawato, T.; Tanaka, H.; Tanabe, N.; Nakai, K.; Zhao, N.; Suzuki, N.; Ochiai, K.; Naeno, M. Sodium butyrate induces the production of cyclooxygenases and prostaglandin E2 in ROS17/2.8 osteoblastic cells. Arch. Oral Biol. 2011, 56, 678–686. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.C.; Lee, J.J.; Chen, Y.J.; Lin, S.I.; Lin, L.D.; Liou, J.W.; Huang, W.L.; Chan, C.P.; Huang, C.C.; Jeng, J.H. Lysophosphatidylcholine induces cytotoxicity/apoptosis and IL-8 production of human endothelial cells: Related mechanisms. Oncotarget 2017, 8, 106177–106189. [Google Scholar] [CrossRef]
- Chang, M.C.; Chan, C.P.; Wang, Y.J.; Lee, P.H.; Chen, L.I.; Tsai, Y.L.; Lin, B.R.; Wang, Y.L.; Jeng, J.H. Induction of necrosis and apoptosis of KB cancer cells by sanguinarine is associated with reactive oxygen species production and mitochondrial membrane depolarization. Toxicol. Appl. Pharmacol. 2007, 218, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.C.; Chen, L.I.; Chan, C.P.; Lee, J.J.; Wang, T.M.; Yang, T.T.; Lin, P.S.; Lin, H.J.; Chang, H.H.; Jeng, J.H. The role of reactive oxygen species and hemeoxygenase-1 expression in the cytotoxicity, cell cycle alteration and apoptosis of dental pulp cells induced by BisGMA. Biomaterials 2010, 31, 8164–8171. [Google Scholar] [CrossRef]
- Sooampon, S.; Manokawinchoke, J.; Pavasant, P. Transient receptor potential vanilloid-1 regulates osteoprotegerin/RANKL homeostasis in human periodontal ligament cells. J. Periodontal Res. 2013, 48, 22–29. [Google Scholar] [CrossRef]
- Chang, M.C.; Wu, H.L.; Lee, J.J.; Lee, P.H.; Chang, H.H.; Hahn, L.J.; Lin, B.R.; Chen, Y.J.; Jeng, J.H. The induction of prostaglandin E2 production, interleukin-6 production, cell cycle arrest, and cytotoxicity in primary oral keratinocytes and κB cancer cells by areca nut ingredients is differentially regulated by MEK/ERK activation. J. Biol. Chem. 2004, 279, 50676–50683. [Google Scholar] [CrossRef]
- Jeng, J.H.; Ho, Y.S.; Chan, C.P.; Wang, Y.J.; Hahn, L.J.; Lei, D.; Hsu, C.C.; Chang, M.C. Areca nut extract up-regulates prostaglandin production, cyclooxygenase-2 mRNA and protein expression of human oral keratinocytes. Carcinogenesis 2000, 21, 1365–1370. [Google Scholar] [CrossRef]
- Chang, H.H.; Guo, M.K.; Kasten, F.H.; Chang, M.C.; Huang, G.F.; Wang, Y.L.; Wang, R.S.; Jeng, J.H. Stimulation of glutathione depletion, ROS production and cell cycle arrest of dental pulp cells and gingival epithelial cells by HEAM. Biomaterials 2005, 26, 745–753. [Google Scholar] [CrossRef]
- Chang, H.H.; Chang, M.H.; Wu, I.H.; Huang, G.F.; Huang, W.L.; Wang, Y.L.; Lee, S.Y.; Yeh, C.Y.; Guo, M.K.; Chan, C.P.; et al. Role of ALK5/Smad2/3 and MEK/ERK signaling in transformning growth factor-beta 1-modulated growth, collagen turnover, and differentiation of stem cells from apical papilla of human tooth. J. Endod. 2015, 41, 1272–1280. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Chang, M.C.; Chen, Y.J.; Liou, J.U.; Chang, H.H.; Huang, W.L.; Liao, W.C.; Chan, C.P.; Jeng, P.Y.; Jeng, J.H. Basic fibroblast growth factor regulates gene and protein expression related to proliferation, differentiation, and matrix production of human dental pulp cells. J. Endod. 2017, 43, 936–942. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Quadrant | Control | 8 mM Butyrate | 16 mM Butyrate | 24 mM Butyrate |
|---|---|---|---|---|
| UR | 0.85 ± 0.10 | 1.25 ± 0.19 | 1.28 ± 0.2 | 1.43 ± 0.40 |
| UL | 4.19 ± 1.12 | 4.19 ± 0.57 | 4.24 ± 0.51 | 4.79 ± 0.33 |
| LR | 0.41 ± 0.06 | 0.86 ± 0.21 | 1.05 ± 0.30 | 0.75 ± 0.24 |
| LL | 94.54 ± 1.05 | 93.69 ± 0.31 | 93.41 ± 0.18 | 93.02 ± 0.36 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, M.-C.; Chen, Y.-J.; Lian, Y.-C.; Chang, B.-E.; Huang, C.-C.; Huang, W.-L.; Pan, Y.-H.; Jeng, J.-H. Butyrate Stimulates Histone H3 Acetylation, 8-Isoprostane Production, RANKL Expression, and Regulated Osteoprotegerin Expression/Secretion in MG-63 Osteoblastic Cells. Int. J. Mol. Sci. 2018, 19, 4071. https://doi.org/10.3390/ijms19124071
Chang M-C, Chen Y-J, Lian Y-C, Chang B-E, Huang C-C, Huang W-L, Pan Y-H, Jeng J-H. Butyrate Stimulates Histone H3 Acetylation, 8-Isoprostane Production, RANKL Expression, and Regulated Osteoprotegerin Expression/Secretion in MG-63 Osteoblastic Cells. International Journal of Molecular Sciences. 2018; 19(12):4071. https://doi.org/10.3390/ijms19124071
Chicago/Turabian StyleChang, Mei-Chi, Yunn-Jy Chen, Yun-Chia Lian, Bei-En Chang, Chih-Chia Huang, Wei-Ling Huang, Yu-Hwa Pan, and Jiiang-Huei Jeng. 2018. "Butyrate Stimulates Histone H3 Acetylation, 8-Isoprostane Production, RANKL Expression, and Regulated Osteoprotegerin Expression/Secretion in MG-63 Osteoblastic Cells" International Journal of Molecular Sciences 19, no. 12: 4071. https://doi.org/10.3390/ijms19124071
APA StyleChang, M.-C., Chen, Y.-J., Lian, Y.-C., Chang, B.-E., Huang, C.-C., Huang, W.-L., Pan, Y.-H., & Jeng, J.-H. (2018). Butyrate Stimulates Histone H3 Acetylation, 8-Isoprostane Production, RANKL Expression, and Regulated Osteoprotegerin Expression/Secretion in MG-63 Osteoblastic Cells. International Journal of Molecular Sciences, 19(12), 4071. https://doi.org/10.3390/ijms19124071