
Ultra-High Hydrostatic Pressure Pretreatment on White Que Zui Tea: Chemical Constituents, Antioxidant, Cytoprotective, and Anti-Inflammatory Activities


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Materials and Treatment
2.3. Extraction of Different Phenolic Fractions
2.4. UHPLC-MS/MS Analysis of WQT
2.5. Determination of the Content of Total Phenolics (TPC) and Total Flavonoids (TFC)
2.6. Evaluation of Antioxidant Activity by DPPH, ABTS, and FRAP Assays
2.6.1. DPPH Assay
2.6.2. ABTS Antioxidant Assay
2.6.3. Determination of Ferric-Reducing Antioxidant Power (FRAP)
2.7. Intracellular Antioxidant Assay
2.7.1. Cytotoxic Test
2.7.2. Measurement of Reactive Oxygen Species
2.7.3. Evaluation of SOD, CAT Activities, and GSH Content
2.8. Determination of Cellular Apoptosis Assay
2.9. Determination of NO Level and Pro-Inflammatory Cytokines (TNF-α, IL-6, and IL-1β) Assay
2.10. Statistical Analysis
3. Results and Discussion
3.1. The Chemical Constituents in WQT
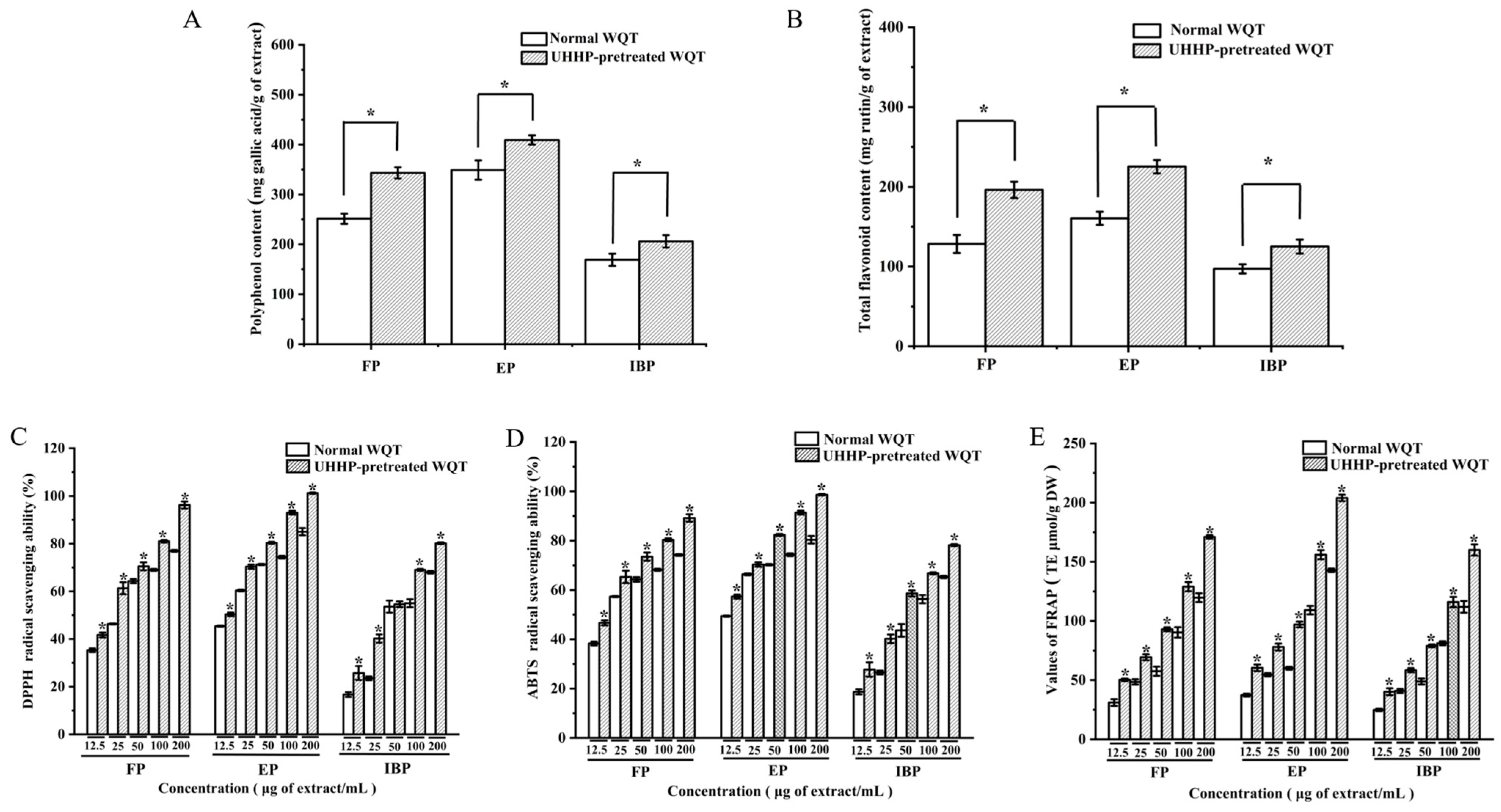
3.2. The Contents of Total Phenolics (TPC) and Total Flavonoids (TFC) in Different Phenolic Fractions from WQT
3.3. Antioxidant Activities of Different Phenolic Fractions from WQT
3.3.1. DPPH Radical Scavenging Abilities of Different Phenolic Fractions from WQT
3.3.2. ABTS Radical Scavenging Abilities of Different Phenolic Fractions from WQT
3.3.3. FRAP Evaluation of Different Phenolic Fractions from WQT
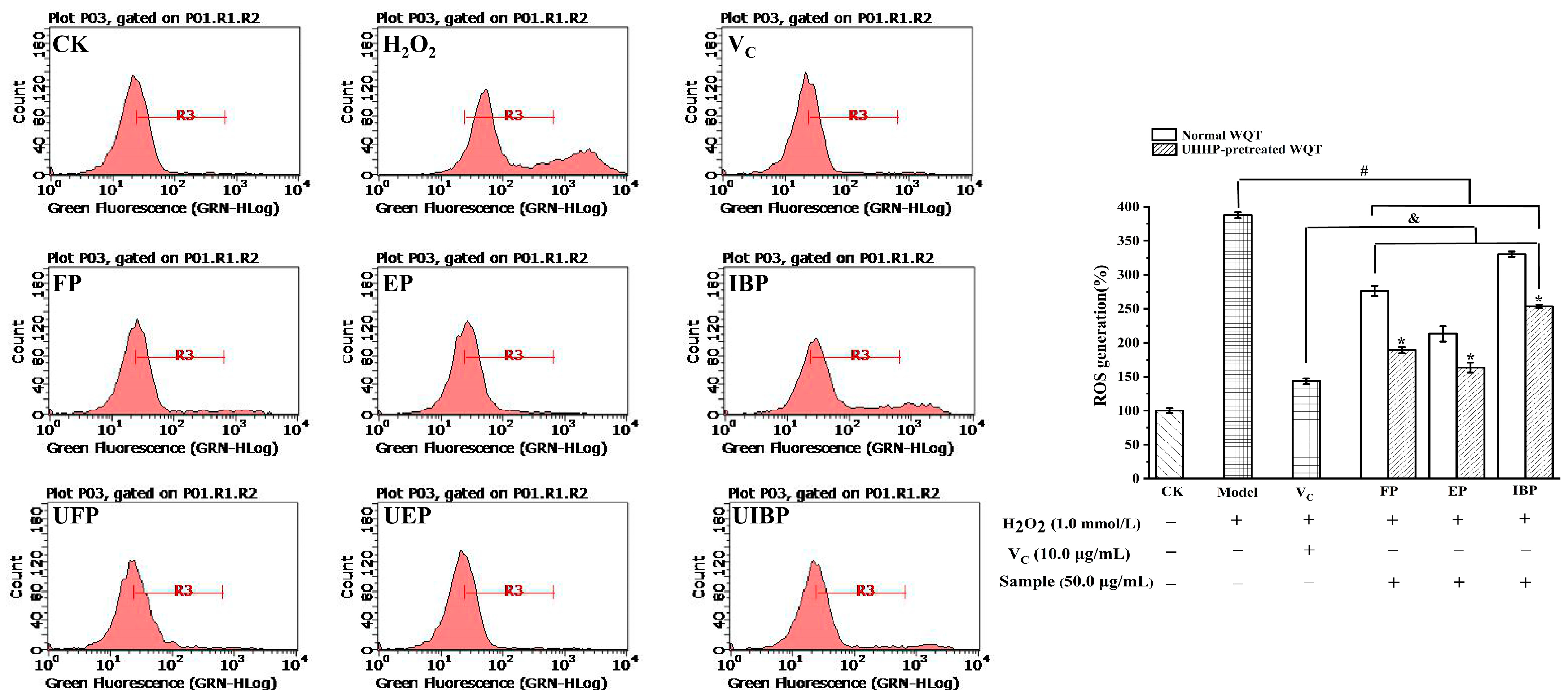
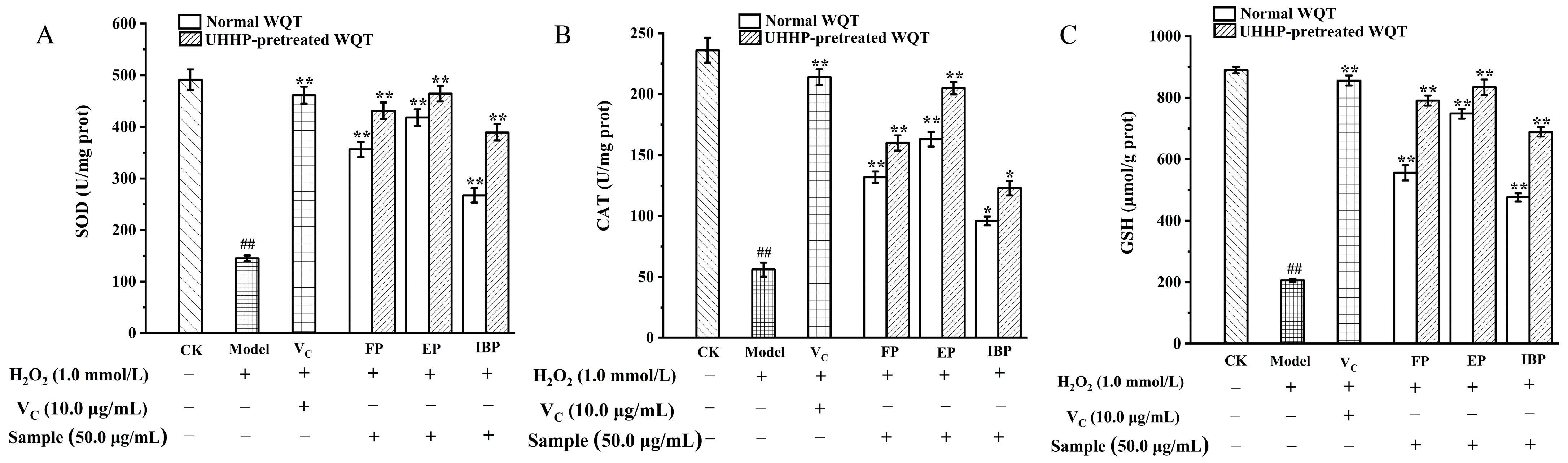
3.4. Intracellular Antioxidant Activities of Different Phenolic Fractions from WQT
3.4.1. Inhibitory Effect on ROS Production
3.4.2. Effects of Different Phenolics Fractions on Cellular SOD, CAT Activities, and GSH Level
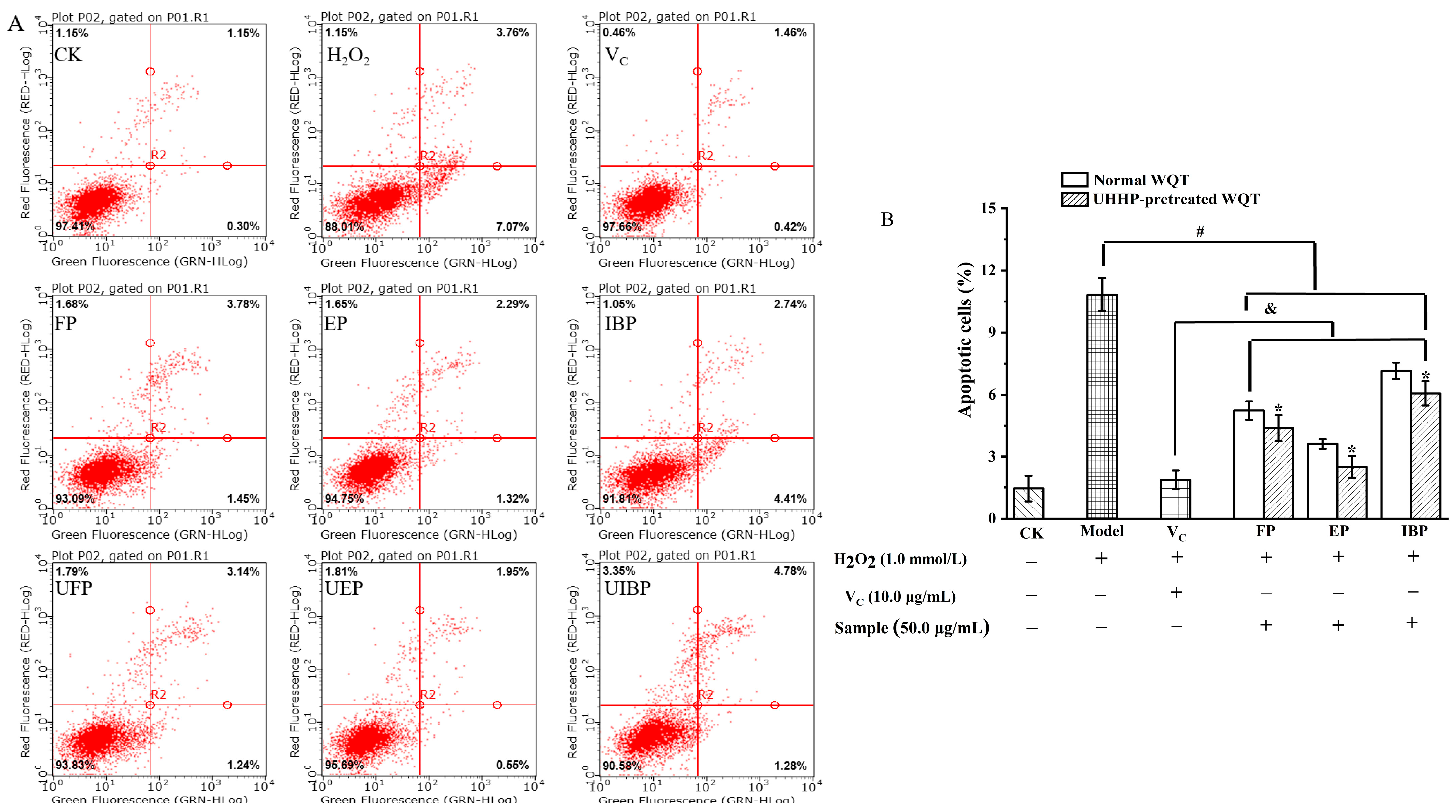
3.5. Inhibitory Effect on H2O2-Induced Cell Apoptosis of Different Phenolic Fractions from WQT
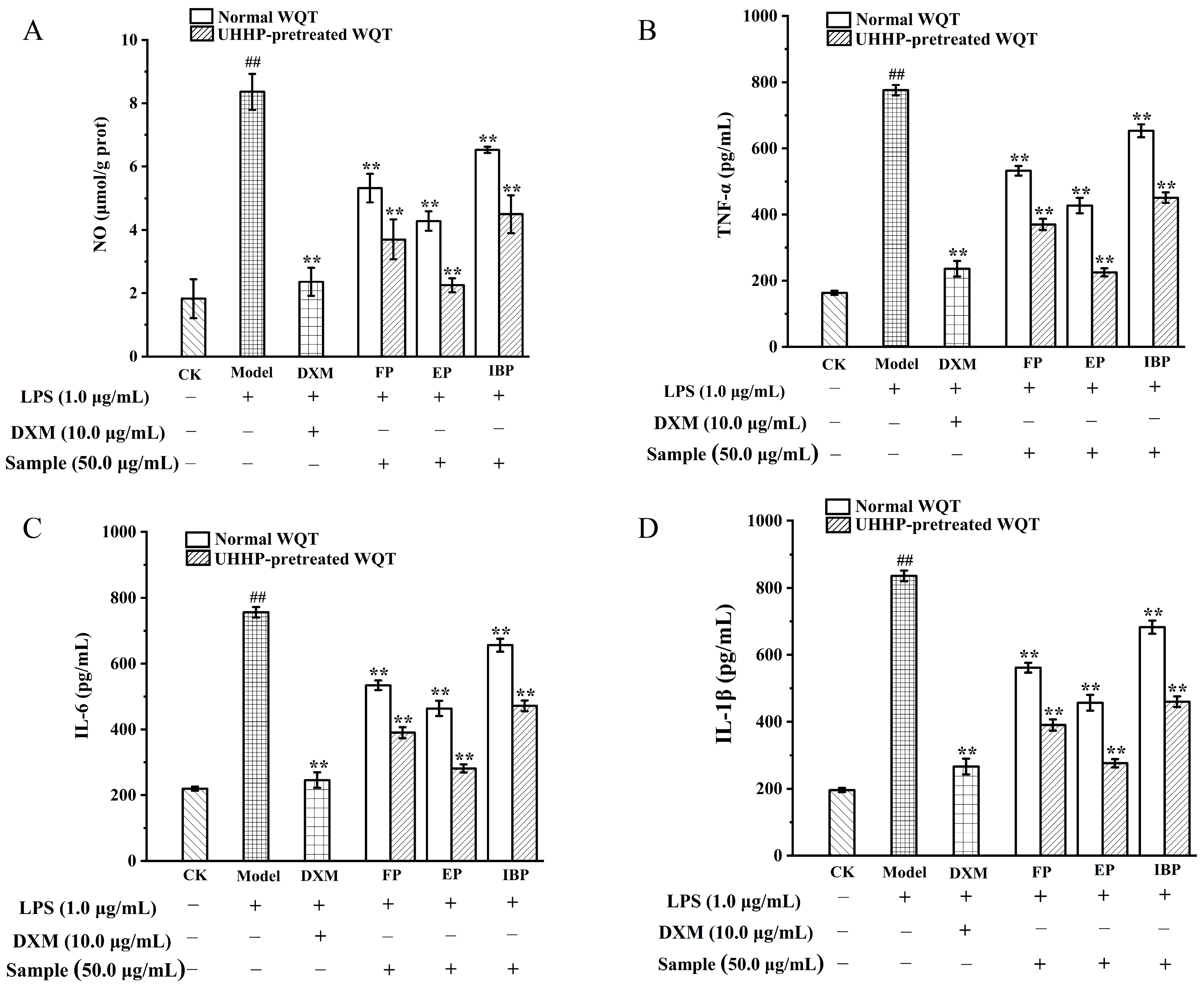
3.6. Anti-Inflammatory Activities of Different Phenolic Fractions from WQT
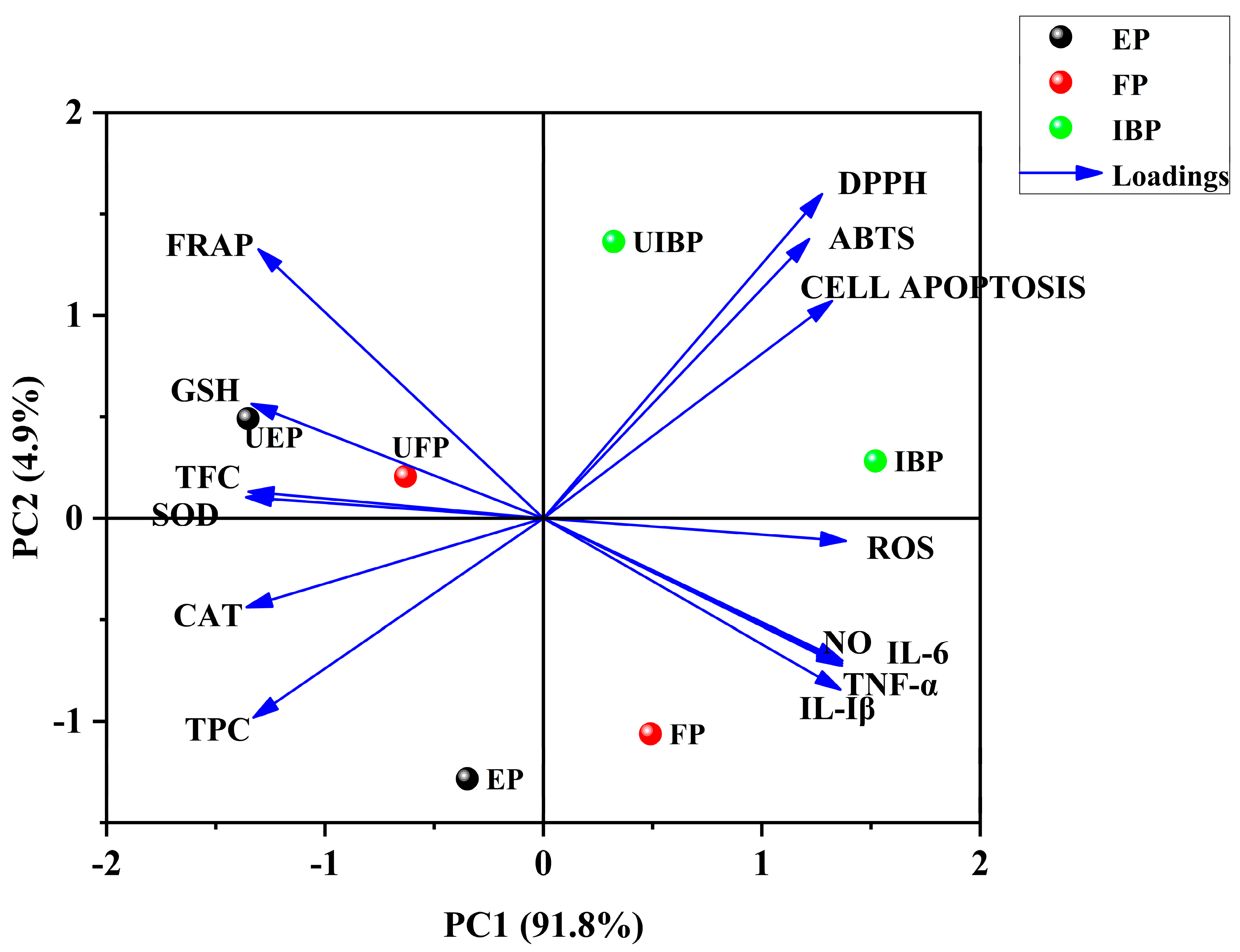
3.7. PCA Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABTS | 2,2′-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) |
| CAT | Catalase |
| DCFH-DA | 2′,7′-dichlorofluorescin diacetate |
| DMEM | dulbecco′s modified eagle′s medium |
| DPPH | 2-diphenyl-1-picrylhydrazyl radical |
| DXM | Dexamethasone |
| FBS | fetal bovine serum |
| GSH | Glutathione |
| IL-1β | interleukin-1β |
| IL-6 | interleukin-6 |
| LPS | Lipopolysaccharide |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenylte-trazolium bromide |
| SOD | superoxide dismutase |
| TNF-α | tumor necrosis factor-α |
| TPTZ | 1,3,5-tri(2-pyridyl)-2,4,6-triazine |
References
- Wu, S.M.; Yu, Q.Y.; Shen, S.; Shan, X.J.; Hua, J.J.; Zhu, J.Y.; Qiu, J.R.; Deng, Y.L.; Zhou, Q.H.; Jiang, Y.W.; et al. Non-targeted metabolomics and electronic tongue analysis reveal the effect of rolling time on the sensory quality and nonvolatile metabolites of congou black tea. Lwt-Food Sci. Technol. 2022, 169, 113971. [Google Scholar] [CrossRef]
- Chandrasekara, A.; Shahidi, F. Herbal beverages: Bioactive compounds and their role in disease risk reduction—A review. J. Tradit. Compl. Med. 2018, 8, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.M.; Huang, L.H.; Suh, J.H.; Wang, Y. Bioactivity-guided isolation and identification of antiadipogenic compounds in Shiya Tea (leaves of Adinandra nitida). J. Agric. Food Chem. 2019, 67, 6785–6791. [Google Scholar] [CrossRef] [PubMed]
- Muller, M.; De Beer, D.D.; Truzzi, C.; Annibaldi, A.; Carloni, P.; Girolametti, F.; Damiani, E.; Joubert, E. Cold brewing of rooibos tea affects its sensory profile and physicochemical properties compared to regular hot, and boiled brewing. Lwt-Food Sci. Technol. 2020, 132, 109919. [Google Scholar] [CrossRef]
- Zhu, C.L.; Yang, Z.B.; He, L.; Lu, X.; Tang, J.N.; Laghi, L. The longer the storage time, the higher the price, the better the quality? a 1H-NMR based metabolomic investigation of aged Ya′an Tibetan Tea (Camellia sinensis). Foods 2022, 11, 2986. [Google Scholar] [CrossRef] [PubMed]
- Barroso, M.V.; Graça-Reis, A.; Cattani-Cavalieri, I.; Gitirana, L.B.; Valenca, S.S.; Lanzetti, M. Mate tea reduces high fat diet-induced liver and metabolic disorders in mice. Biomed. Pharmacother. 2018, 109, 1547–1555. [Google Scholar] [CrossRef]
- Naithani, V.; Nair, S.; Kakkar, P. Decline in antioxidant capacity of Indian herbal teas during storage and its relation to phenolic content. Food Res. Int. 2006, 2, 176–181. [Google Scholar] [CrossRef]
- Zhao, C.N.; Tang, G.Y.; Cao, S.Y.; Xu, X.Y.; Gan, R.Y.; Liu, Q.; Mao, Q.Q.; Shang, A.; Li, H.B. Phenolic profiles and antioxidant activities of 30 tea infusions from green, black, oolong, white, yellow and dark teas. Antioxidants 2019, 7, 215. [Google Scholar] [CrossRef]
- Grgić, J.; Šelo, G.; Planinić, M.; Tišma, M.; Bucić-Kojić, A. Role of the encapsulation in bioavailability of phenolic compounds. Antioxidants 2020, 9, 923. [Google Scholar] [CrossRef]
- Zhang, J.K.; Zhou, X.L.; Wang, X.Q.; Zhang, J.X.; Yang, M.L.; Liu, Y.P.; Cao, J.X.; Cheng, G.G. Que Zui tea ameliorates hepatic lipid accumulation and oxidative stress in high fat diet induced nonalcoholic fatty liver disease. Food Res. Int. 2022, 156, 111196. [Google Scholar] [CrossRef]
- Ren, L.; Zhang, J.; Zhang, T.H. Immunomodulatory activities of polysaccharides from Ganoderma on immune effector cells. Food Chem. 2021, 340, 127933. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.H.; Zhong, S.N.; Li, T.Z.; Zhang, J. Saponins as modulators of nuclear receptors. Crit. Rev. Food Sci. Nutr. 2020, 1, 94–107. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.Y.; Ye, H.Q.; Kamaraj, R.; Zhang, T.H.; Zhang, J.; Pavek, P. A review on pharmacological activities and synergistic effect of quercetin with small molecule agents. Phytomedicine 2021, 92, 153736. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ren, L.; Zhang, T.H.; Pavek, P.; Kamaraj, R. Dietary phytochemicals as modulators of human pregnane X receptor. Crit. Rev. Food Sci. Nutr. 2021, 1–23. [Google Scholar] [CrossRef]
- Wang, Y.P.; Wang, Z.X.; Xue, Q.W.; Zhen, L.; Wang, Y.D.; Cao, J.X.; Liu, Y.P.; Khan, A.; Zhao, T.R.; Cheng, G.G. Effect of ultra-high pressure pretreatment on the phenolic profiles, antioxidative activity and cytoprotective capacity of different phenolic fractions from Que Zui tea. Food Chem. 2023, 409, 135271. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Y.D.; Xue, Q.W.; Zhao, T.R.; Khan, A.; Wang, Y.F.; Liu, Y.P.; Cao, J.X.; Cheng, G.G. The effect of ultra-high pretreatment on free, esterified and insoluble-bound phenolics from mango leaves and their antioxidant and cytoprotective activities. Food Chem. 2022, 368, 130864. [Google Scholar] [CrossRef]
- Deng, J.L.; Xiang, Z.Y.; Lin, C.B.; Zhu, Y.Q.; Yang, K.J.; Liu, T.H.; Xia, C.; Chen, J.; Zhang, W.H.; Zhang, Y.H.; et al. Identification and quantification of free, esterified, and insoluble-bound phenolics in grains of hulless barley varieties and their antioxidant activities. Lwt-Food Sci. Technol. 2021, 151, 112001. [Google Scholar] [CrossRef]
- Thu Dao, T.A.; Webb, H.K.; Malherbe, F. Optimisation of pectin extraction from fruit peels by response surface method: Conventional versus microwave-assisted heating. Food Hydrocolloid 2021, 113, 106475. [Google Scholar] [CrossRef]
- Gu, J.Y.; Zhang, H.H.; Zhang, J.X.; Wen, C.T.; Ma, H.L.; Duan, Y.Q.; He, Y.Q. Preparation, characterization and bioactivity of polysaccharide fractions from Sagittaria sagittifolia L. Carbohyd. Polym. 2019, 229, 115355. [Google Scholar] [CrossRef]
- Lv, X.J.; Li, Y.; Ma, S.G.; Qu, J.; Liu, Y.B.; Li, Y.H.; Zhang, D.; Li, L.; Yu, S.S. Antiviral triterpenes from the twigs and leaves of Lyonia ovalifolia. J. Nat. Prod. 2016, 384, 132517. [Google Scholar] [CrossRef]
- Zhao, D.R.; Su, L.H.; Li, R.T.; Chen, X.Q.; Li, H.M. Chemical constituents from the twigs and leaves of Lyonia ovalifolia. Biochem. Syst. Ecol. 2018, 78, 1–4. [Google Scholar] [CrossRef]
- Gao, S.H.; Zhao, T.R.; Liu, Y.P.; Wang, Y.F.; Cheng, G.G.; Cao, J.X. Phenolic constituents, antioxidant activity and neuroprotective effects of ethanol extracts of fruits, leaves and flower buds from Vaccinium dunalianum Wight. Food Chem. 2021, 374, 131752. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.R.; Sun, M.X.; Kong, L.P.; Xue, Q.W.; Wang, Y.D.; Wang, Y.F.; Khan, A.; Cao, J.X.; Cheng, G.G. Bioactivity-guided isolation of phytochemicals from Vaccinium dunalianum Wight and their antioxidant and enzyme inhibitory activities. Molecules 2021, 26, 2075. [Google Scholar] [CrossRef] [PubMed]
- Minh, T.N.; Van, T.M.; Khanh, T.D.; Xuan, T.D. Isolation and identification of constituents exhibiting antioxidant, antibacterial, and antihyperuricemia activities in piper methysticum root. Foods 2022, 11, 3889. [Google Scholar] [CrossRef] [PubMed]
- He, S.Y.; Cui, X.Y.; Khan, A.; Liu, Y.P.; Wang, Y.D.; Cui, Q.M.; Zhao, T.R.; Cao, J.X.; Cheng, G.G. Activity guided isolation of phenolic compositions from Anneslea fragrans Wall. and their cytoprotective effect against hydrogen peroxide induced oxidative stress in HepG2 cells. Molecules 2021, 26, 3690. [Google Scholar] [CrossRef]
- Oyeniran, O.H.; Ademiluyi, A.O.; Oboh, G. African mistletoe (Tapinanthus bangwensis Lor.) infestation improves the phenolic constituents, antioxidative and antidiabetic effects of almond (Terminalia catappa Linn.) host leaf in sucrose-rich diet-induced diabetic-like phenotypes in fruit fly (Drosophila melanogaster Meigen). Food Front. 2021, 2, 77–90. [Google Scholar] [CrossRef]
- Yang, M.L.; Wang, Y.D.; Patel, G.; Xue, Q.W.; Singor Njateng, G.S.; Cai, S.B.; Cheng, G.G.; Kai, G.Y. In vitro and in vivo anti-inflammatory effects of different extracts from Epigynum auritum through down-regulation of NF-κB and MAPK signaling pathways. J. Ethnopharmacol. 2020, 261, 113105. [Google Scholar] [CrossRef]
- Gu, C.Z.; Yang, M.L.; Zhou, Z.H.; Khan, A.; Cao, J.X.; Cheng, G.G. Purification and characterization of four benzophenone derivatives from Mangifera indica L. leaves and their antioxidant, immunosuppressive and α-glucosidase inhibitory activities. J. Funct. Foods 2018, 52, 709–714. [Google Scholar] [CrossRef]
- Ding, J.; Mei, S.H.; Gao, L.; Wang, Q.; Ma, H.L.; Chen, X.M. Tea processing steps affect chemical compositions, enzyme activities, and antioxidant and anti-inflammatory activities of coffee leaves. Food Front. 2022, 3, 505–516. [Google Scholar] [CrossRef]
- Zhou, J.X.; Ma, Y.L.; Jia, Y.J.; Pang, M.J.; Cheng, G.G.; Cai, S.B. Phenolic profiles, antioxidant activities and cytoprotective effects of different phenolic fractions from oil palm (Elaeis guineensis Jacq.) fruits treated by ultra-high pressure. Food Chem. 2019, 288, 68–77. [Google Scholar] [CrossRef]
- Arruda, H.S.; Pereira, G.A.; de Morais, D.R.; Eberlin, M.N.; Pastore, G.M. Determination of free, esterified, glycosylated and insoluble-bound phenolics composition in the edible part of araticum fruit (Annona crassiflora Mart.) and its by-products by HPLC-ESI-MS/MS. Food Chem. 2017, 245, 738–749. [Google Scholar] [CrossRef] [PubMed]
- Camboim Rockett, F.; de Oliveira Schmidt, H.; Schmidt, L.; Rodrigues, E.; Tischer, B.; Ruffo de Oliveira, V.; Lima da Silva, V.; Rossini Augusti, P.; Hickmann Flôres, S.; Rios, A. Phenolic compounds and antioxidant activity in vitro and in vivo of Butia and Opuntia fruits. Food Res. Int. 2020, 137, 109740. [Google Scholar] [CrossRef] [PubMed]
- Kolodziejczyk-Czepas, J.; Nowak, P.; Moniuszko-Szajwaj, B.; Kowalska, I.; Stochmal, A. Free radical scavenging actions of three Trifoliumspecies in the protection of blood plasma antioxidant capacityin vitro. Pharm. Biol. 2015, 53, 1277–1284. [Google Scholar] [CrossRef]
- Leopoldini, M.; Marino, T.; Russo, N.; Toscano, M. Antioxidant properties of phenolic compounds: H-atom versus electron transfer mechanism. J. Phys. Chem. A 2004, 108, 4916–4922. [Google Scholar] [CrossRef]
- Palafox-Carlos, H.; Gil-Chávez, J.; Sotelo-Mundo, R.; Namiesnik, J.; Gorinstein, S.; González-Aguilar, G. Antioxidant interactions between major phenolic compounds found in ‘Ataulfo’ mango pulp: Chlorogenic, gallic, protocatechuic and vanillic acids. Molecules 2012, 17, 12657–12664. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.M.; Wang, Y.D.; Zhou, W.B.; He, S.Y.; Yang, M.L.; Xue, Q.W.; Wang, Y.F.; Zhao, T.R.; Cao, J.X.; Khan, A.; et al. Phenolic composition, antioxidant and cytoprotective effects of aqueous-methanol extract from Anneslea fragrans leaves as affected by drying methods. Int. J. Food Sci. 2021, 56, 4807–4819. [Google Scholar] [CrossRef]
- Hu, Y.M.; Lu, S.Z.; Li, Y.S.; Wang, H.; Shi, Y.; Zhang, L.; Tu, Z.C. Protective effect of antioxidant peptides from grass carp scale gelatin on the H2O2-mediated oxidative injured HepG2 cells. Food Chem. 2021, 373, 131539. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, S.N.; Sheng, Z.L.; Li, X.; Chang, Y.N.; Dai, W.C.; Chang, S.K.; Liu, J.M.; Yang, Y.C. Effect of pinolenic acid on oxidative stress injury in HepG2 cells induced by H2O2. Food Sci. Nutr. 2021, 9, 5689–5697. [Google Scholar] [CrossRef]
- Xu, Y.; Ji, Y.; Li, X.; Ding, J.Z.; Chen, L.Q.; Huang, Y.F.; Wei, W.X. URI1 suppresses irradiation-induced reactive oxygen species (ROS) by activating autophagy in hepatocellular carcinoma cells. Int. J. Biol. Sci. 2021, 17, 3091–3103. [Google Scholar] [CrossRef]
- Liu, J.; Meng, C.G.; Yan, Y.H.; Shan, Y.N.; Kan, J.; Jin, C.H. Protocatechuic acid grafted onto chitosan: Characterization and antioxidant activity. Int. J. Biol. Macromol. 2016, 89, 518–526. [Google Scholar] [CrossRef]
- Yang, J.X.; Guo, J.; Yuan, J.F. In vitro antioxidant properties of rutin. LWT-Food Sci. Technol. 2008, 41, 1060–1066. [Google Scholar] [CrossRef]
- Yen, G.C.; Duh, P.D.; Tsai, H.L. Antioxidant and pro-oxidant properties of ascorbic acid and gallic acid. Food Chem. 2002, 3, 307–313. [Google Scholar] [CrossRef]
- Niki, E. Assessment of antioxidant capacity in vitro and in vivo. Free. Radic. Bio. Med. 2010, 49, 503–515. [Google Scholar] [CrossRef]
- Wang, L.Y.; Ding, L.; Yu, Z.P.; Zhang, T.; Ma, S.; Liu, J.B. Intracellular ROS scavenging and antioxidant enzyme regulating capacities of corn gluten meal-derived antioxidant peptides in HepG2 cells. Food Res. Int. 2017, 90, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Li, F.Y.; Lu, S.Y.; Ren, L.M.; Bian, S.; Liu, M.C.; Zhao, D.Q.; Wang, S.M.; Wang, J.W. Ginseng root extract attenuates inflammation by inhibiting the MAPK/NF-κB signaling pathway and activating autophagy and p62-Nrf2-Keap1 signaling in vitro and in vivo. J. Ethnopharmacol. 2021, 283, 114739. [Google Scholar] [CrossRef] [PubMed]
- Thi Ngoc Anh, P.; Hae Lim, K.; Dong-Ryung, L.; Bong-Keun, C.; Seung Hwan, Y. Anti-inflammatory effects of scrophularia buergeriana extract mixture fermented with lactic acid bacteria. Biotechnol. Bioproc. E 2022, 27, 370–378. [Google Scholar] [CrossRef]
- Saji, N.; Francis, N.; Schwarz, L.J.; Blanchard, C.L.; Santhakumar, A.B. The antioxidant and anti-inflammatory properties of rice bran phenolic extracts. Foods 2020, 6, 829. [Google Scholar] [CrossRef]
- Gu, I.; Brownmiller, C.; Stebbins, N.B.; Mauromoustakos, A.; Howard, L.; Lee, S.-O. Berry phenolic and volatile extracts inhibit pro-inflammatory cytokine secretion in LPS-stimulated RAW264.7 cells through suppression of NF-κB signaling pathway. Antioxidants 2020, 9, 871. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Li, M.; Zhen, L.; Wang, Y.; Wang, Y.; Qin, Y.; Zhang, Z.; Zhao, T.; Cao, J.; Liu, Y.; et al. Ultra-High Hydrostatic Pressure Pretreatment on White Que Zui Tea: Chemical Constituents, Antioxidant, Cytoprotective, and Anti-Inflammatory Activities. Foods 2023, 12, 628. https://doi.org/10.3390/foods12030628
Zhang X, Li M, Zhen L, Wang Y, Wang Y, Qin Y, Zhang Z, Zhao T, Cao J, Liu Y, et al. Ultra-High Hydrostatic Pressure Pretreatment on White Que Zui Tea: Chemical Constituents, Antioxidant, Cytoprotective, and Anti-Inflammatory Activities. Foods. 2023; 12(3):628. https://doi.org/10.3390/foods12030628
Chicago/Turabian StyleZhang, Xiaoyu, Mengcheng Li, Li Zhen, Yudan Wang, Yifen Wang, Yuyue Qin, Zhihong Zhang, Tianrui Zhao, Jianxin Cao, Yaping Liu, and et al. 2023. "Ultra-High Hydrostatic Pressure Pretreatment on White Que Zui Tea: Chemical Constituents, Antioxidant, Cytoprotective, and Anti-Inflammatory Activities" Foods 12, no. 3: 628. https://doi.org/10.3390/foods12030628
APA StyleZhang, X., Li, M., Zhen, L., Wang, Y., Wang, Y., Qin, Y., Zhang, Z., Zhao, T., Cao, J., Liu, Y., & Cheng, G. (2023). Ultra-High Hydrostatic Pressure Pretreatment on White Que Zui Tea: Chemical Constituents, Antioxidant, Cytoprotective, and Anti-Inflammatory Activities. Foods, 12(3), 628. https://doi.org/10.3390/foods12030628