
Enhanced Zinc Intake Protects against Oxidative Stress and Its Consequences in the Brain: A Study in an In Vivo Rat Model of Cadmium Exposure

 , ,
, ,
Abstract
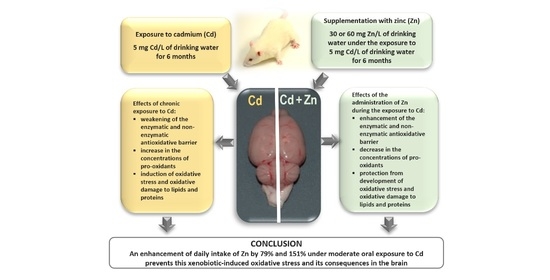
:
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Sources of Cd Exposure and Zn Supplementation
2.3. Study Protocol
- Control: Received drinking water without the addition of Cd and Zn and the standard rodent chow;
- Zn 30: Received drinking water containing 30 mg Zn/L;
- Zn 60: Received drinking water containing 60 mg Zn/L;
- Cd: Treated with Cd in drinking water at the concentration of 5 mg/L;
- Cd + Zn 30: Received drinking water containing Cd and Zn at the concentrations of 5 mg/L and 30 mg/L, respectively; and
- Cd + Zn 60: Received drinking water containing Cd and Zn at the concentrations of 5 mg/L and 60 mg/L, respectively.
2.4. Estimation of the Oxidative/Antioxidative Status of the Brain and Serum
2.4.1. Preparation of the Brain Tissue Homogenates
2.4.2. Assay of the Markers of the Oxidative/Antioxidative Status and Quantification of the Severity of Oxidative Stress in the Brain
Antioxidants in the Brain
Pro-Oxidants in the Brain
Indices of Oxidative Injury of Lipids and Proteins
2.4.3. Determination of the Severity of Oxidative Stress in the Serum
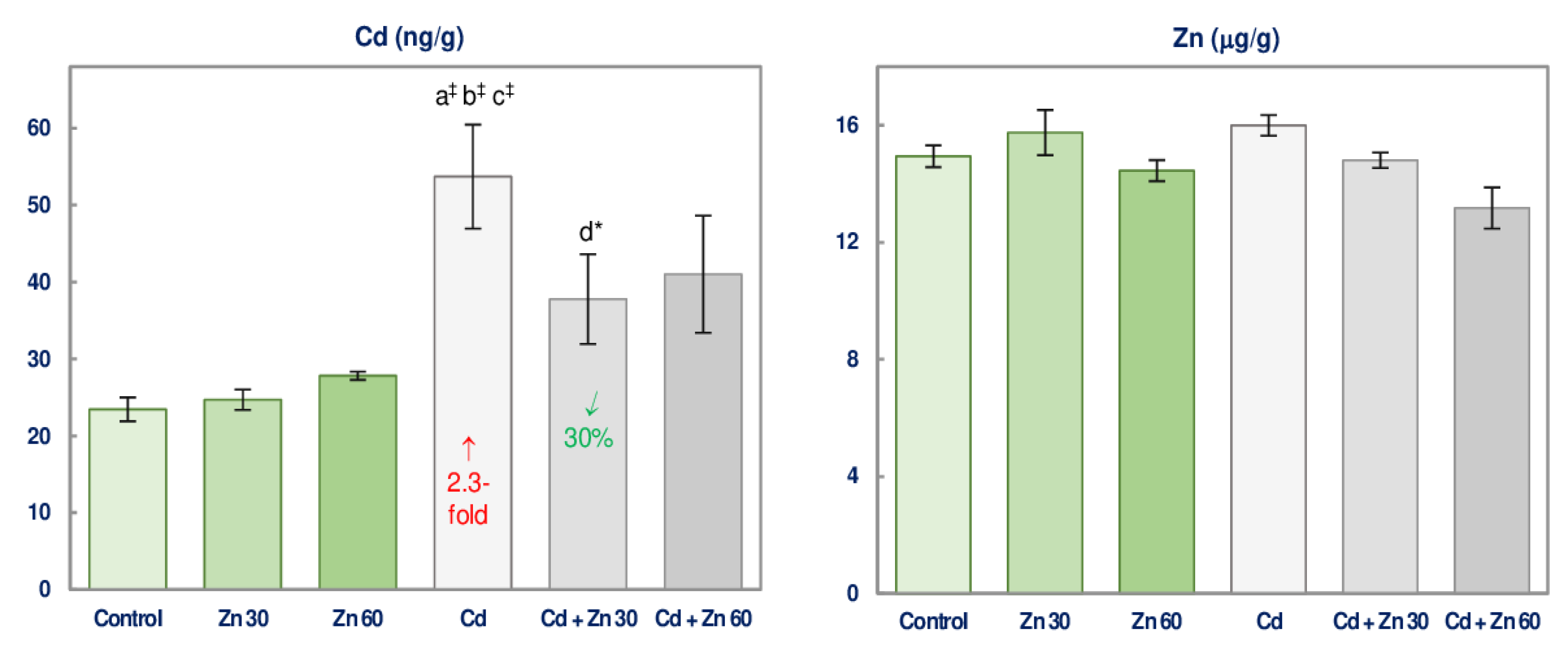
2.5. Determination of the Concentration of Zn and Cd in the Brain
2.6. Statistical Analysis
3. Results
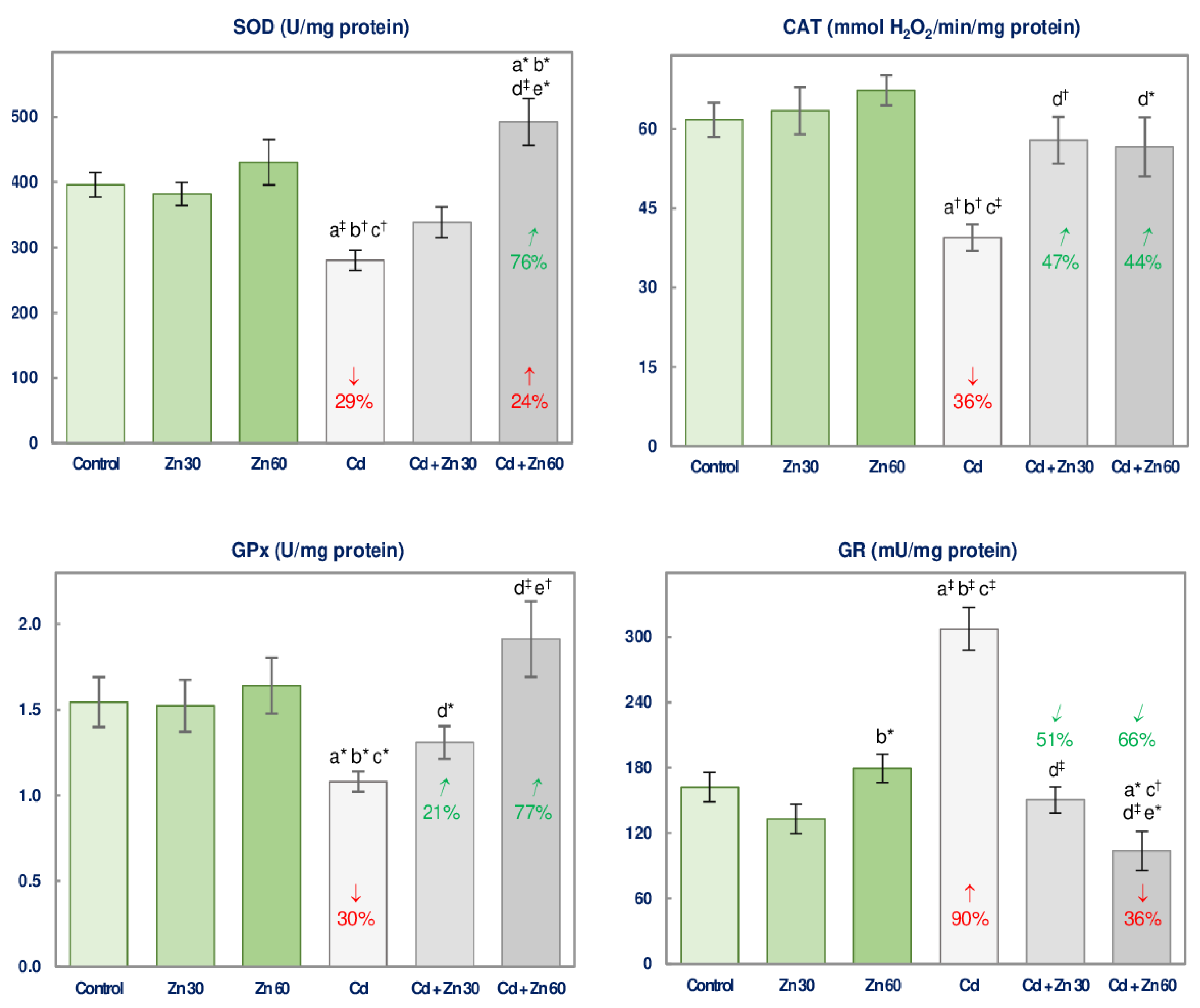
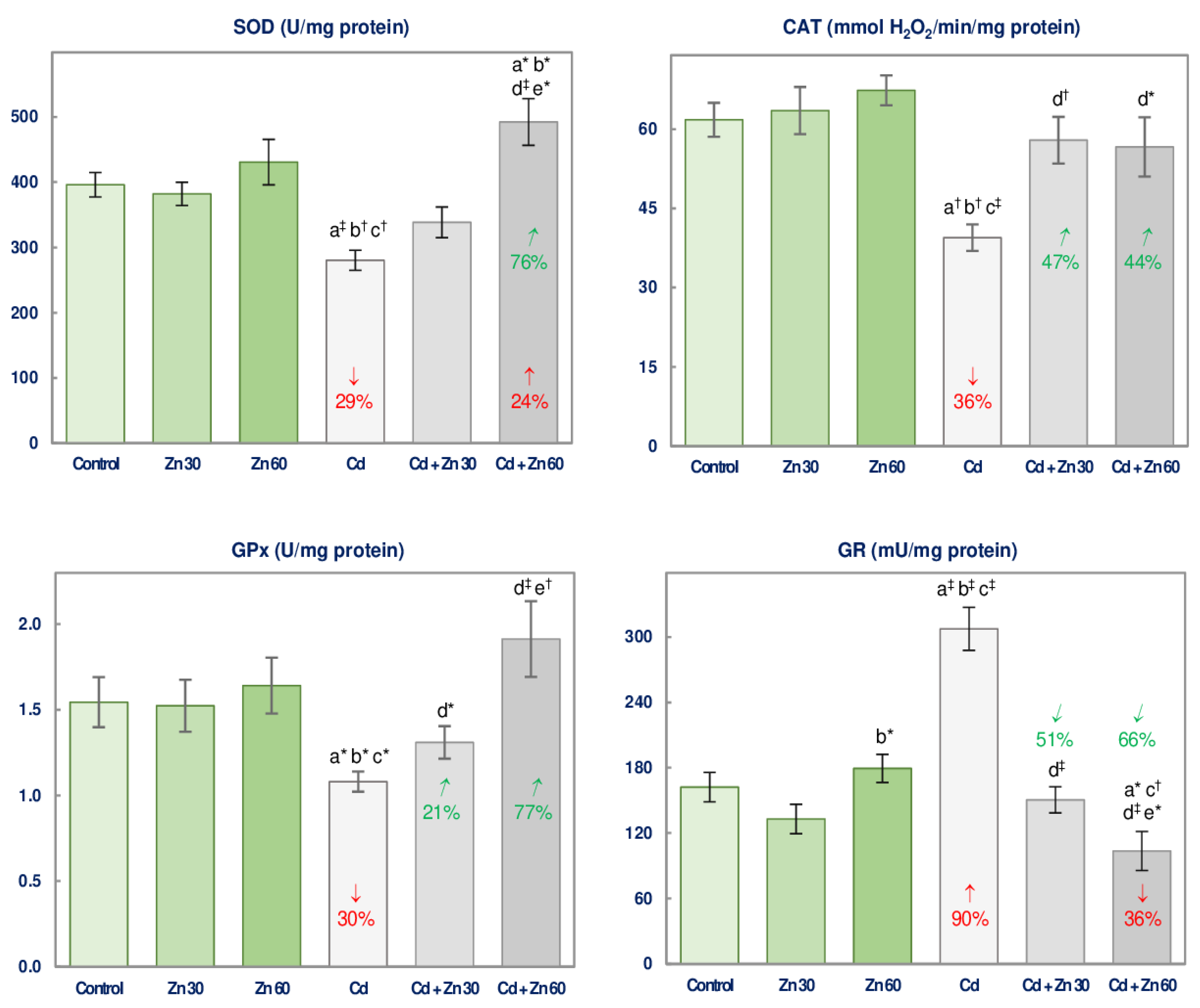
3.1. The Impact of Cd and/or Zn on the Enzymatic Antioxidative Status of the Brain
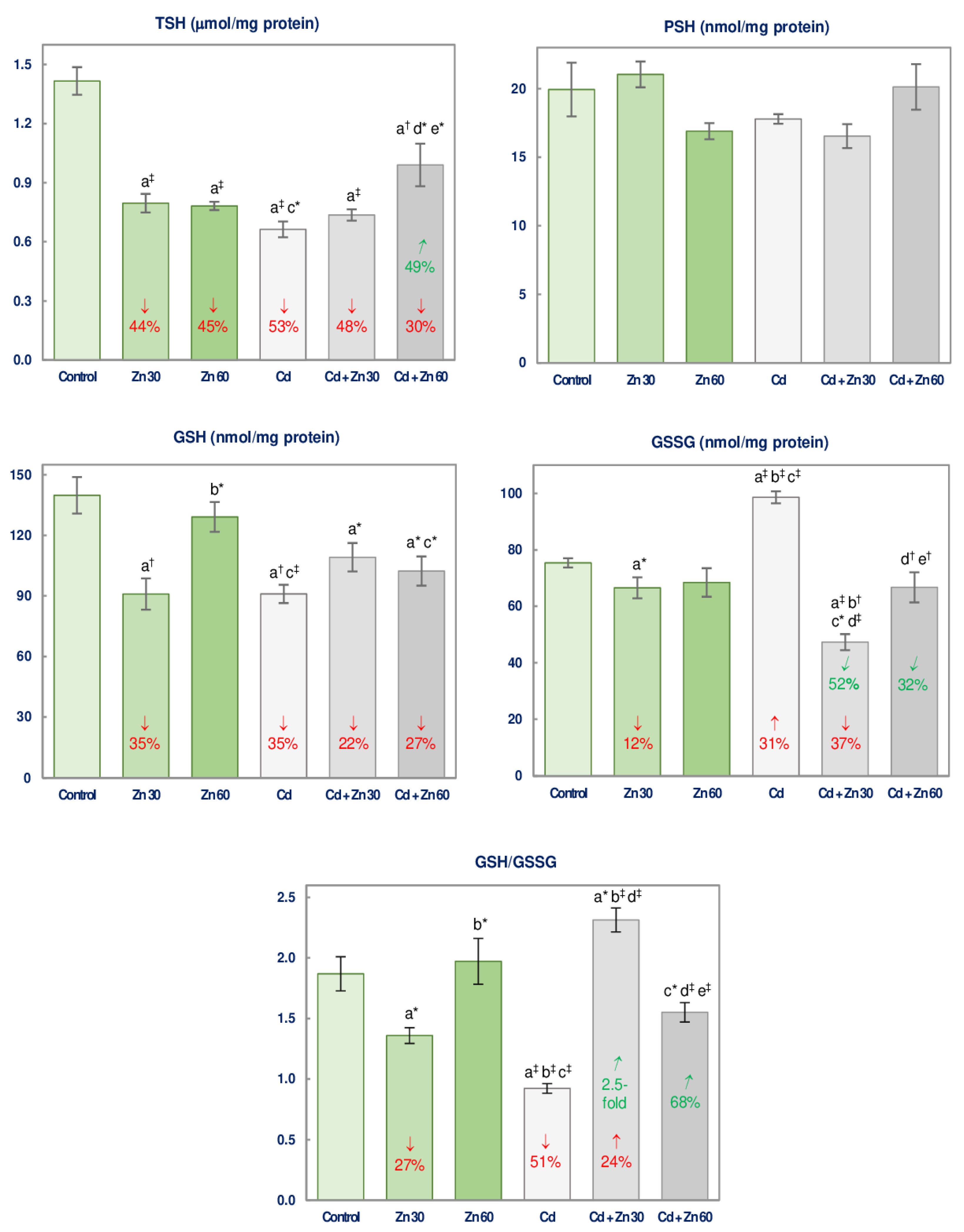
3.2. The Impact of Cd and/or Zn on the Non-Enzymatic Antioxidative Status of the Brain
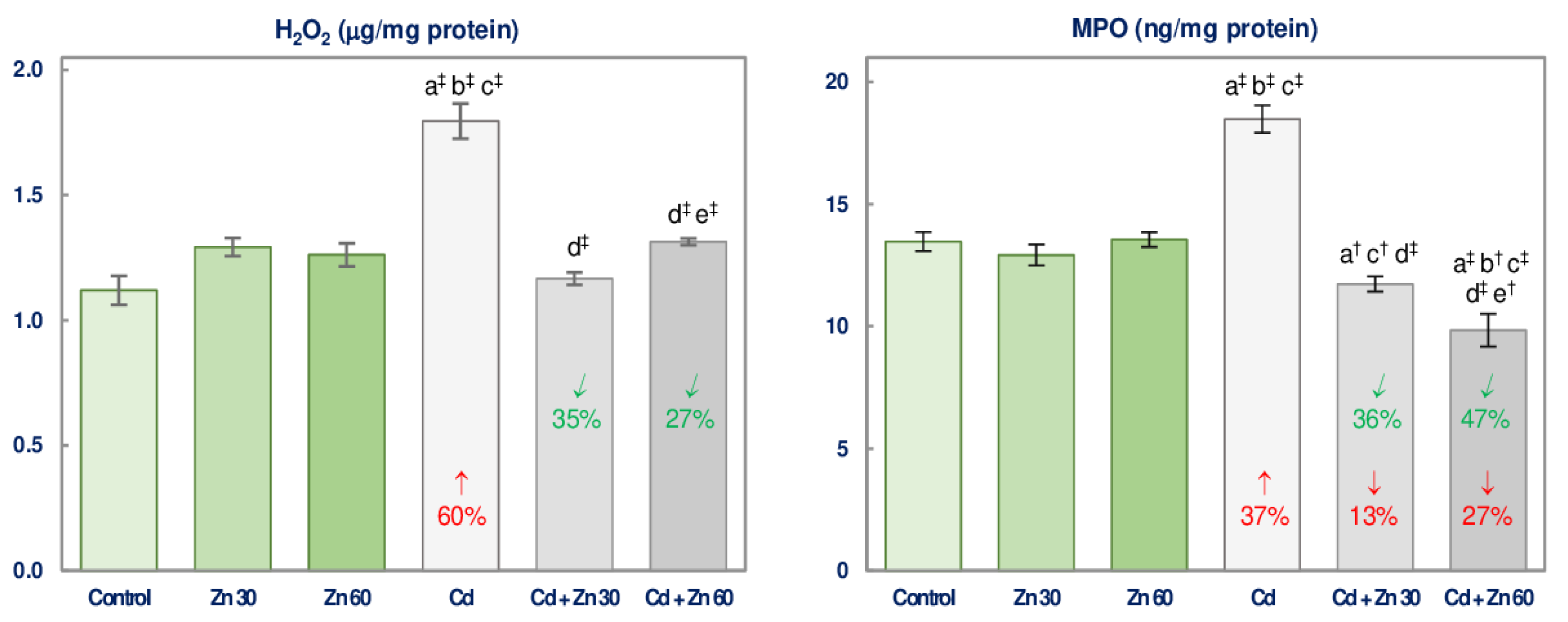
3.3. The Impact of Cd and/or Zn on Pro-oxidants Concentration in the Brain
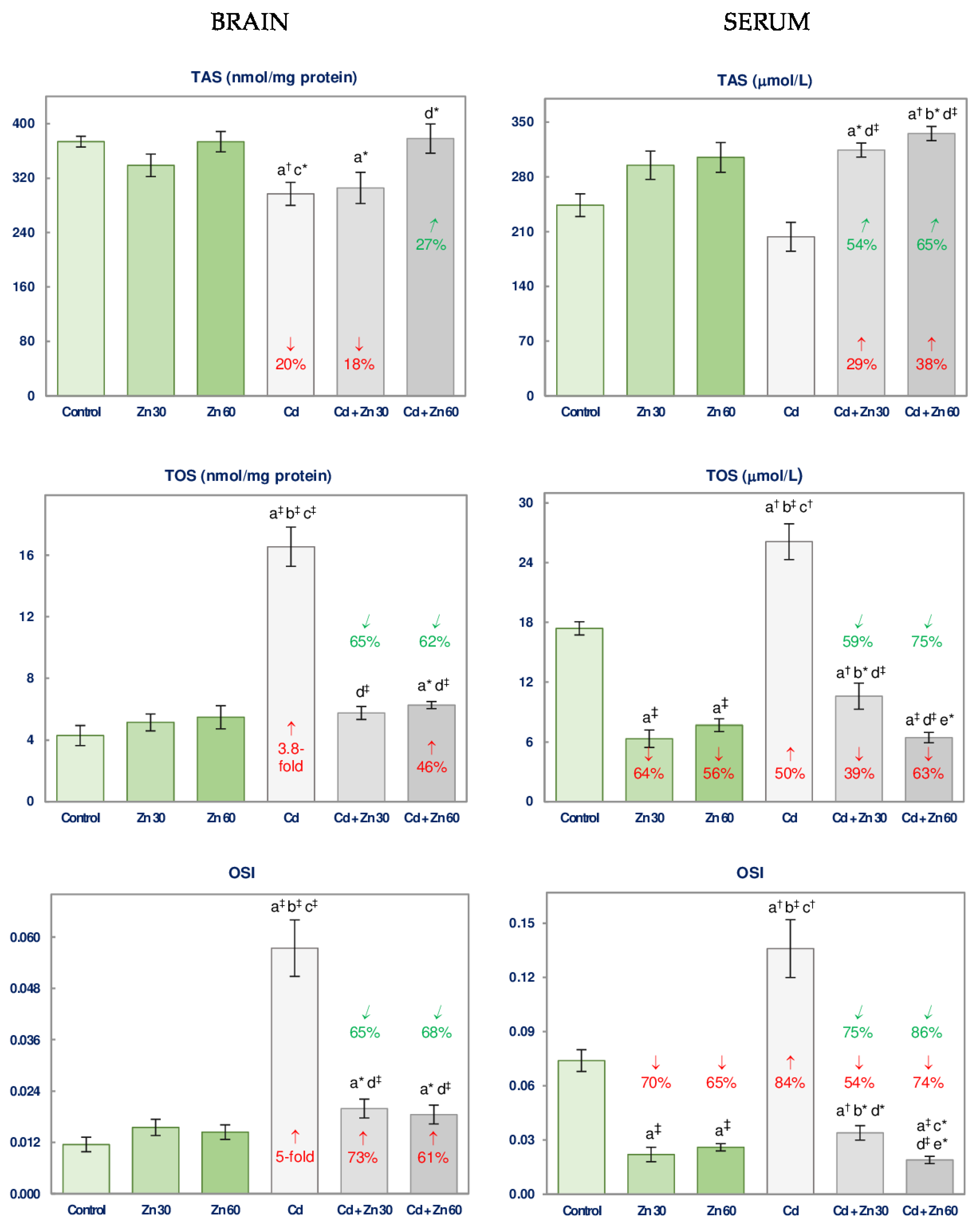
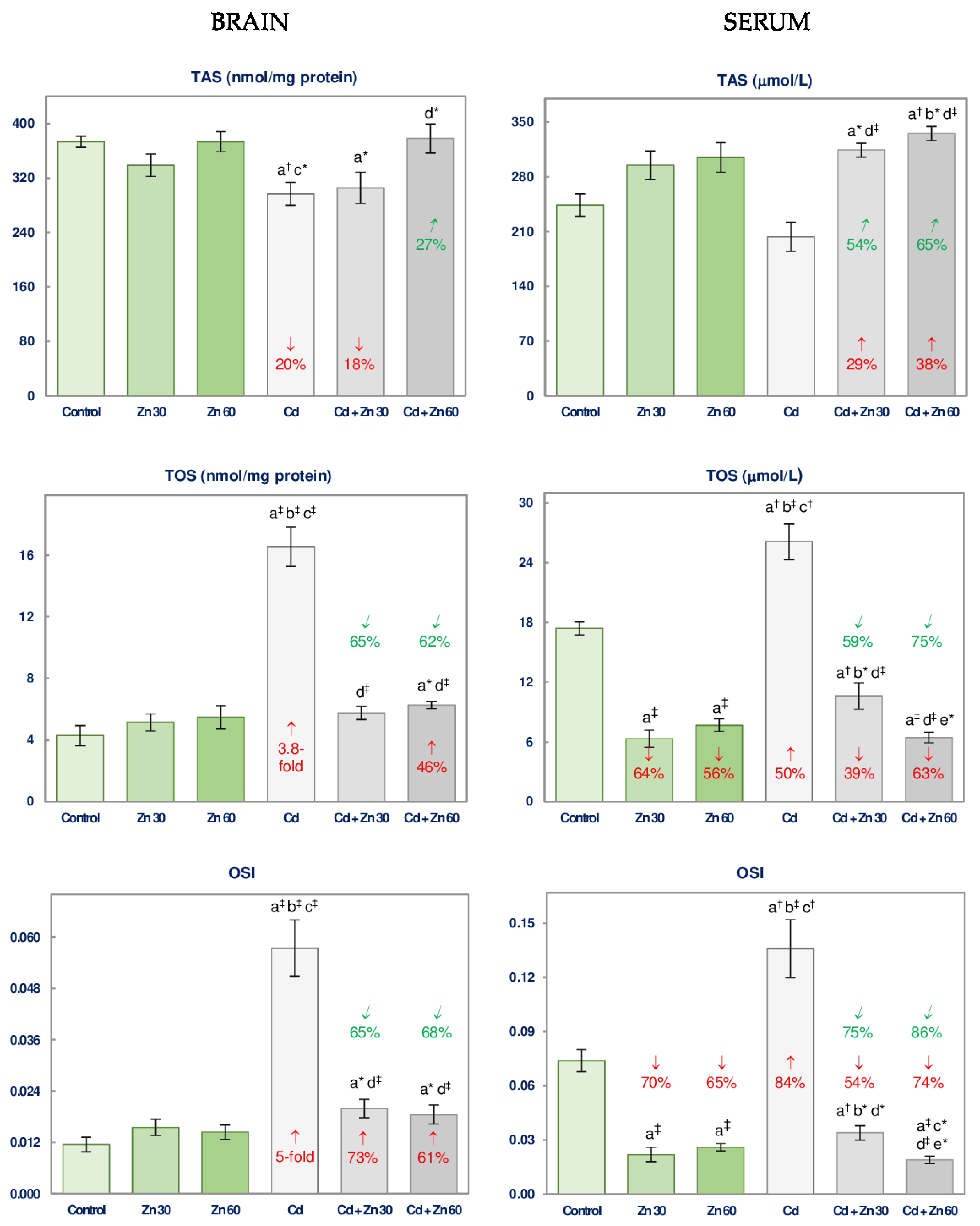
3.4. The Impact of Cd and/or Zn on TAS and TOS and the Severity of Oxidative Stress in the Brain and Serum
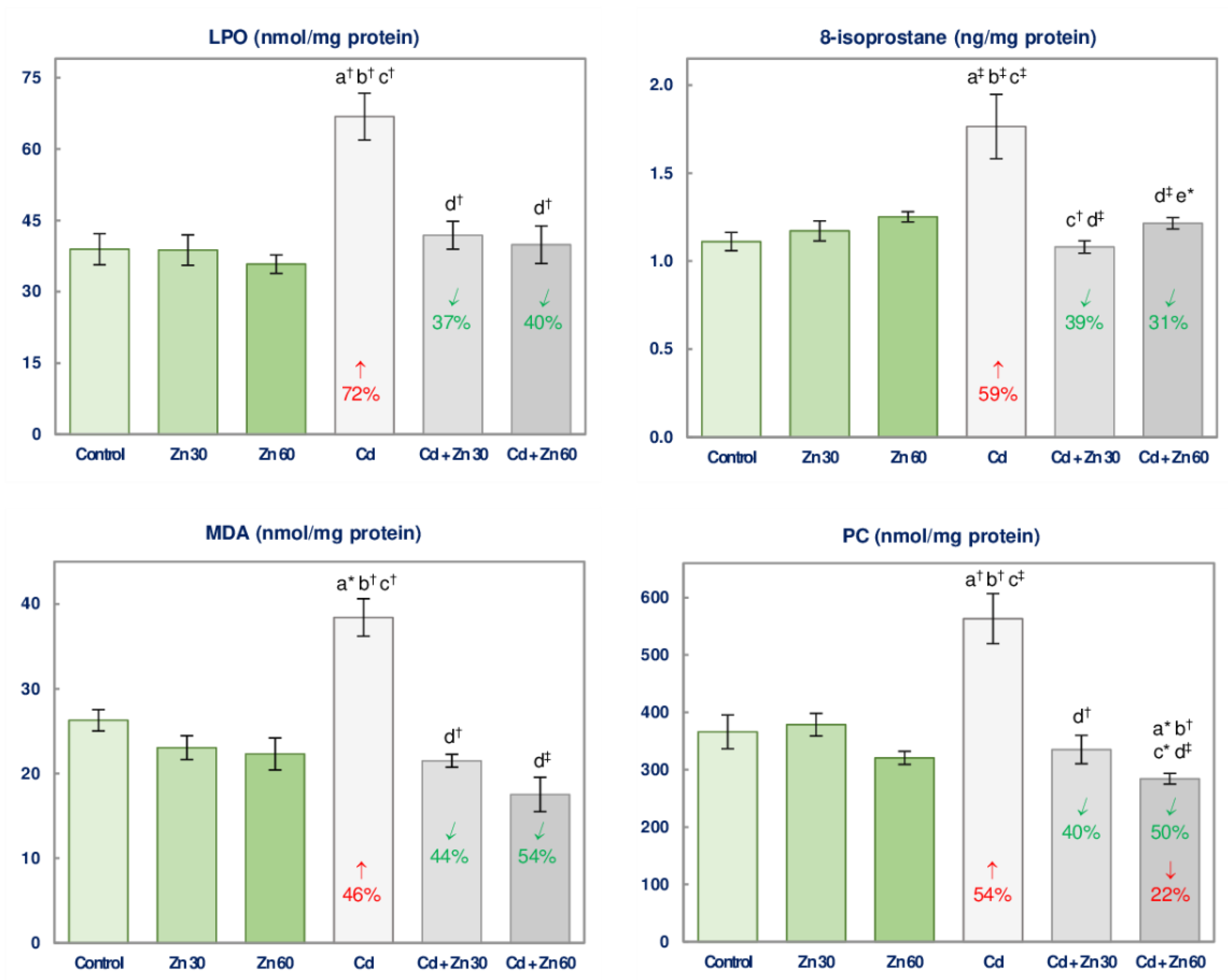
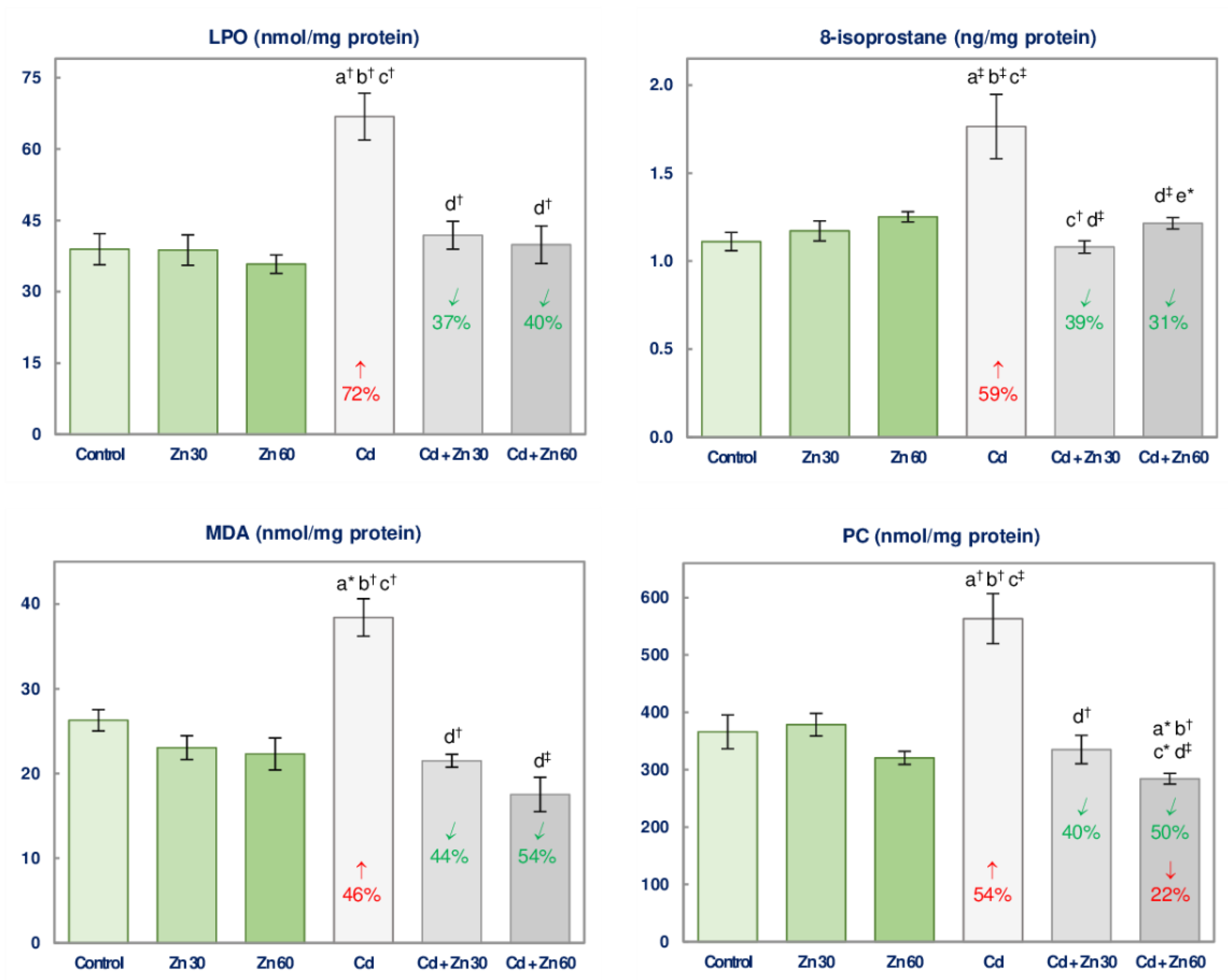
3.5. The Impact of Cd and/or Zn on the Oxidative Injury of Lipids and Proteins in the Brain
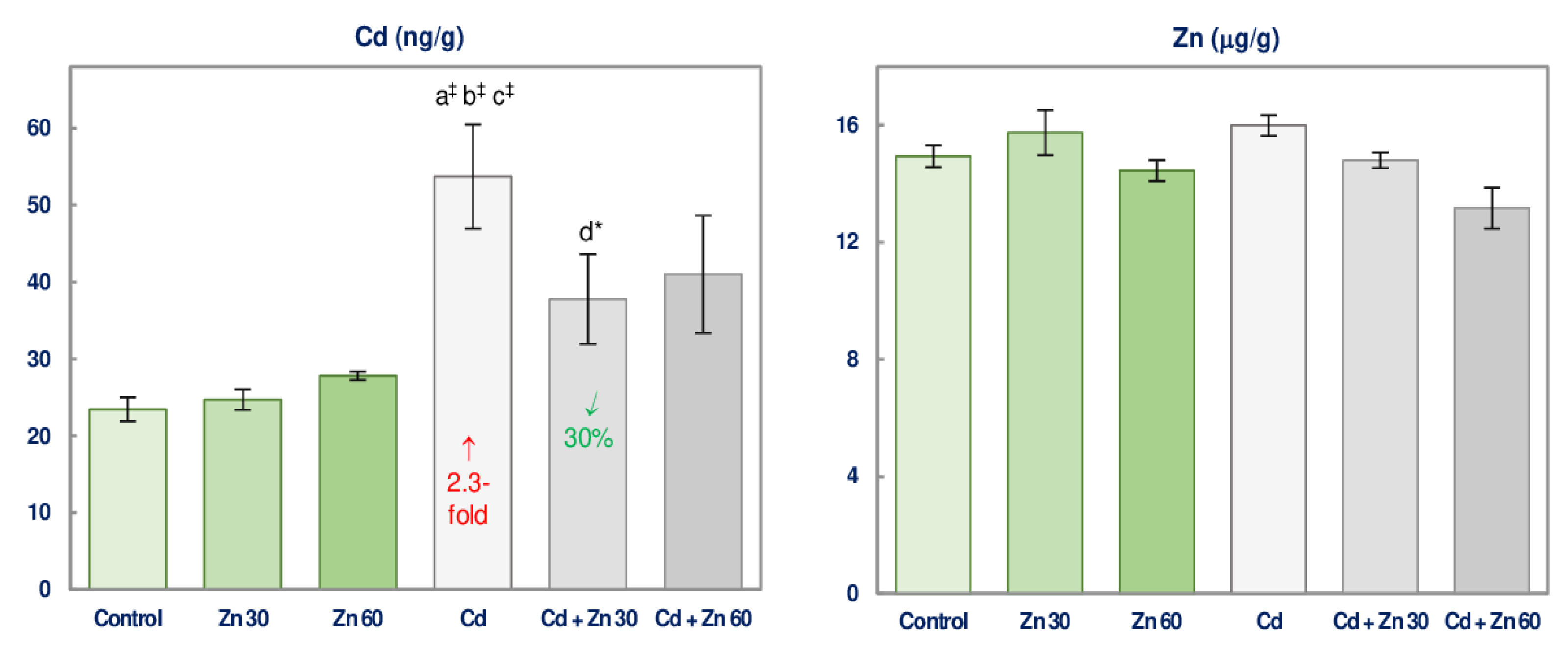
3.6. Cd and Zn Concentration in the Brain
3.7. Correlations
4. Discussion
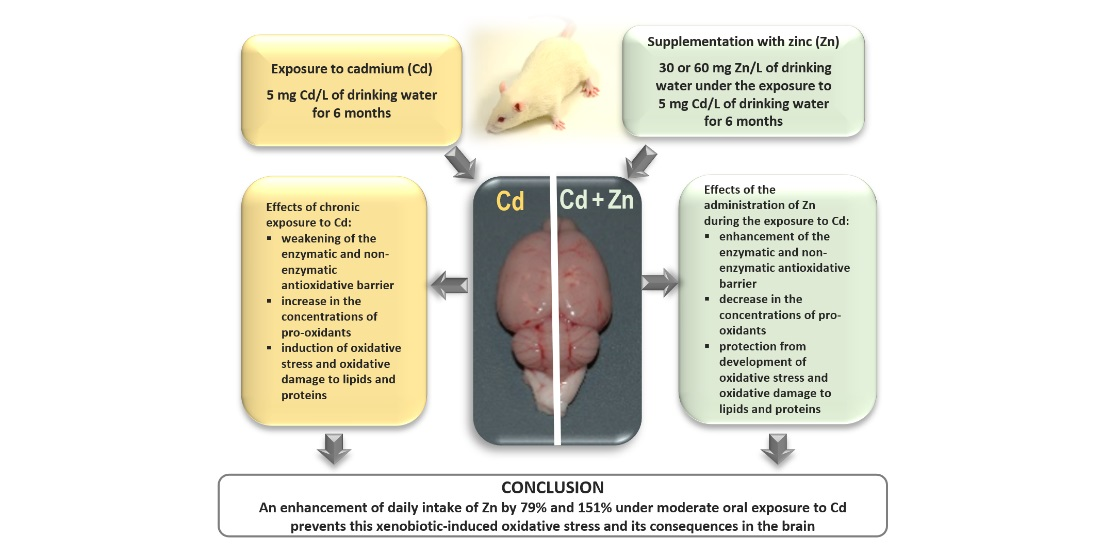
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AAS | atomic absorption spectrophotometry |
| ANOVA/MANOVA | two-way analysis of variance |
| b.w. | body weight |
| Ca | calcium |
| CAT | catalase |
| Cd | cadmium |
| Cd2+ | cadmium ion |
| CdCl2 | cadmium chloride |
| Cu | copper |
| CV | coefficient of variation |
| ELISA | enzyme-linked immunosorbent assay |
| Fe | iron |
| GPx | glutathione peroxidase |
| GR | glutathione reductase |
| GSH | reduced glutathione |
| GSSG | oxidized glutathione |
| H2O2 | hydrogen peroxide |
| i.p. | intraperitoneal, intraperitoneally |
| LPO | lipid peroxides |
| MDA | malondialdehyde |
| MPO | myeloperoxidase |
| MT | metallothionein |
| OSI | oxidative stress index |
| O2− | superoxide radicals |
| p | level of statistical significance |
| PC | protein carbonyls |
| PSH | protein thiol groups |
| r | correlation coefficient |
| ROS | reactive oxygen species |
| SE | standard error |
| SD | standard deviation |
| -SH | sulfhydryl group |
| SOD | superoxide dismutase |
| TAS | total antioxidative status |
| TOS | total oxidative status |
| TSH | total thiol groups |
| Zn | zinc |
| ZnCl2 | zinc chloride |
| Zn2+ | zinc ion |
References
- Roohani, N.; Hurrell, R.; Kelishadi, R.; Schulin, R. Zinc and its importance for human health: An integrative review. J. Res. Med. Sci. 2013, 18, 144–157. [Google Scholar] [PubMed]
- Jarosz, M.; Olbert, M.; Wyszogrodzka, G.; Młyniec, K.; Librowski, T. Antioxidant and anti-inflammatory effects of zinc. Zinc-dependent NF-κB signaling. Inflammopharmacology 2017, 25, 11–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.R. Critical role of zinc as either an antioxidant or a prooxidant in cellular systems. Oxid. Med. Cell. Longev. 2018, 2018, 9156285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfgang, M.; Sandstead, H.H. Zinc requirements and the risks and benefits of zinc supplementation. J. Trace Elem. Med. Biol. 2006, 20, 3–18. [Google Scholar] [CrossRef]
- Narayanan, S.E.; Rehuman, N.A.; Harilal, S.; Vincent, A.; Rajamma, R.G.; Behl, T.; Uddin, M.S.; Ashraf, G.M.; Mathew, B. Molecular mechanism of zinc neurotoxicity in Alzheimer’s disease. Environ. Sci. Pollut. Res. 2020, 27, 43542–43552. [Google Scholar] [CrossRef]
- Irving, J.A.; Mattman, A.; Lockitch, G.; Farrell, K.; Wadsworth, L.D. Element of caution: A case of reversible cytopenias associated with excessive zinc supplementation. CMAJ 2003, 169, 129–131. [Google Scholar]
- Prasad, A.S. Discovery of human zinc deficiency: Its impact on human health and disease. Adv. Nutr. 2013, 4, 176–190. [Google Scholar] [CrossRef]
- Kumssa, D.; Joy, E.; Ander, E.L.; Watts, M.J.; Young, S.D.; Walker, S.; Broadley, M.R. Dietary calcium and zinc deficiency risks are decreasing but remain prevalent. Sci. Rep. 2015, 5, 10974. [Google Scholar] [CrossRef] [Green Version]
- Terrin, G.; Boscarino, G.; Chiara, M.D.; Iacobelli, S.; Faccioli, F.; Greco, C.; Onestà, E.; Sabatini, G.; Pietravalle, A.; Oliva, S.; et al. Nutritional intake influences zinc levels in preterm newborns: An observational study. Nutrients 2020, 12, 529. [Google Scholar] [CrossRef] [Green Version]
- Brzóska, M.M.; Rogalska, J.; Gałażyn-Sidorczuk, M.; Jurczuk, M.; Roszczenko, A.; Kulikowska-Karpińska, E.; Moniuszko-Jakoniuk, J. Effect of zinc supplementation on bone metabolism in male rats chronically exposed to cadmium. Toxicology 2007, 237, 89–103. [Google Scholar] [CrossRef]
- Rogalska, J.; Brzóska, M.M.; Roszczenko, A.; Moniuszko-Jakoniuk, J. Enhanced zinc consumption prevents cadmium-induced alterations in lipid metabolism in male rats. Chem. Biol. Interact. 2009, 177, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Rogalska, J.; Piłat-Marcinkiewicz, B.; Brzóska, M.M. Protective effect of zinc against cadmium hepatotoxicity depends on this bioelement intake and level of cadmium exposure: A study in a rat model. Chem. Biol. Interact. 2011, 193, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Brzóska, M.M.; Roszczenko, A.; Gałażyn-Sidorczuk, M.; Majewska, K. Zinc supplementation can protect from enhanced risk of femoral neck fracture in male rats chronically exposed to cadmium. Exp. Toxicol. Pathol. 2011, 63, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Gałażyn-Sidorczuk, M.; Brzóska, M.M.; Rogalska, J.; Roszczenko, A.; Jurczuk, M. Effect of zinc supplementation on glutathione peroxidase activity and selenium concentration in the serum, liver and kidney of rats chronically exposed to cadmium. J. Trace Elem. Med. Biol. 2012, 26, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Brzóska, M.M.; Rogalska, J. Protective effect of zinc supplementation against cadmium-induced oxidative stress and the RANK/RANKL/OPG system imbalance in the bone tissue of rats. Toxicol. Appl. Pharmacol. 2013, 272, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Brzóska, M.M.; Moniuszko-Jakoniuk, J.; Jurczuk, M.; Gałażyn-Sidorczuk, M.; Rogalska, J. The effect of zinc supply on cadmium-induced changes in the tibia of rats. Food Chem. Toxicol. 2001, 39, 729–737. [Google Scholar] [CrossRef]
- Yu, H.T.; Zhen, J.; Leng, J.Y.; Cai, L.; Ji, H.L.; Keller, B.B. Zinc as a countermeasure for cadmium toxicity. Acta Pharmacol. Sin. 2020. [Google Scholar] [CrossRef]
- Mimouna, S.B.; Chemek, M.; Boughammoura, S.; Haouas, Z.; Messaoudi, I. Protective role of zinc against the neurotoxicity induced by exposure to cadmium during gestation and lactation periods on hippocampal volume of pups tested in early adulthood. Drug Chem. Toxicol. 2018, 41, 424–433. [Google Scholar] [CrossRef]
- Rahman, M.M.; Hossain, K.F.B.; Banik, S.; Sikder, M.T.; Akter, M.; Bondad, S.E.C.; Rahaman, M.S.; Hosokawa, T.; Saito, T.; Kurasaki, M. Selenium and zinc protections against metal-(loids)-induced toxicity and disease manifestations: A review. Ecotoxicol. Environ. Saf. 2019, 30, 146–163. [Google Scholar] [CrossRef]
- Wang, C.C.; Si, L.F.; Guo, S.N.; Zheng, J.L. Negative effects of acute cadmium on stress defense, immunity, and metal homeostasis in liver of zebrafish: The protective role of environmental zinc pre-exposure. Chemosphere 2019, 222, 91–97. [Google Scholar] [CrossRef]
- Kostecka-Sochoń, P.; Onopiuk, B.; Dąbrowska, E. Protective effect of increased zinc supply against oxidative damage of sublingual gland in chronic exposure to cadmium: Experimental study on rats. Oxid. Med. Cell. Longev. 2018, 2018, 3732842. [Google Scholar] [CrossRef] [PubMed]
- Bernotiene, R.; Ivanoviene, L.; Sadauskiene, I.; Liekis, A.; Ivanov, L. Effects of cadmium and zinc ions on mice brain lipid peroxidation and amounts of thiol-rich compounds. Trace Elem. Electrolytes 2016, 33, 83–88. [Google Scholar] [CrossRef] [Green Version]
- Mężyńska, M.; Brzóska, M.M. Environmental exposure to cadmium—A risk for health of the general population in industrialized countries and preventive strategies. Environ. Sci. Pollut. Res. 2018, 25, 3211–3232. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Melough, M.M.; Vance, T.M.; Noh, H.; Koo, S.I.; Chun, O.K. Dietary cadmium intake and sources in the US. Nutrients 2019, 11, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eom, S.Y.; Seo, M.N.; Lee, Y.S.; Park, K.S.; Hong, Y.S.; Sohn, S.J.; Kim, Y.D.; Choi, B.S.; Lim, J.A.; Kwon, H.J.; et al. Low-level environmental cadmium exposure induces kidney tubule damage in the general population of Korean adults. Arch. Environ. Contam. Toxicol. 2017, 73, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Hyder, O.; Chung, M.; Cosgrove, D.; Herman, J.M.; Li, Z.; Firoozmand, A.; Gurakar, A.; Koteish, A.; Pawlik, T.M. Cadmium exposure and liver disease among US adults. J. Gastrointest. Surg. 2013, 17, 1265–1273. [Google Scholar] [CrossRef]
- Fagerberg, B.; Barregard, L.; Sallsten, G.; Forsgard, N.; Ostling, G.; Persson, M.; Borné, Y.; Engström, G.; Hedblad, B. Cadmium exposure and atherosclerotic carotid plaques—Results from the Malmö diet and cancer study. Environ. Res. 2015, 136, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Djordjevic, V.R.; Wallace, D.R.; Schweitzer, A.; Boricic, N.; Knezevic, D.; Maticm, S.; Grubor, N.; Kerkez, M.; Radenkovic, D.; Bulat, Z.; et al. Environmental cadmium exposure and pancreatic cancer: Evidence from case control, animal and in vitro studies. Environ. Int. 2019, 128, 353–361. [Google Scholar] [CrossRef]
- Okuda, B.; Iwamoto, Y.; Tachibana, H.; Sugita, M. Parkinsonism after acute cadmium poisoning. Clin. Neurol. Neurosurg. 1997, 99, 263–265. [Google Scholar] [CrossRef]
- Min, J.; Min, K. Blood cadmium levels and Alzheimer’s disease mortality risk in older US adults. Environ. Health 2016, 15, 69. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Du, Y. Cadmium and its neurotoxic effects. Oxid. Med. Cell. Longev. 2013, 2013, 898034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mir, R.H.; Sawhney, G.; Pottoo, F.H.; Mohi-ud-din, R.; Madishetti, S.; Jachak, S.M.; Ahmed, Z.; Masoodi, M.H. Role of environmental pollutants in Alzheimer’s disease: A review. Environ. Sci. Pollut. Res. 2020, 27, 44724–44742. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Wakode, S.; Sharma, A.; Nair, N.; Dhobi, M.; Wani, M.A.; Pottoo, F.H. Effect of environmental toxicants on neuronal functions. Environ. Sci. Pollut. Res. 2020, 27, 44906–44921. [Google Scholar] [CrossRef] [PubMed]
- Braga, M.M.; Dick, T.; de Oliveira, D.L.; Scopel-Guerra, A.; Mussulini, B.H.M.; Souza, D.O.; Rocha, J.B.T. Evaluation of zinc effect on cadmium action in lipid peroxidation and metallothionein levels in the brain. Toxicol. Rep. 2015, 2, 858–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karri, V.; Schuhmacher, M.; Kumar, V. Heavy metals (Pb, Cd, As and MeHg) as risk factors for cognitive dysfunction: A general review of metal mixture mechanism in brain. Environ. Toxicol. Pharmacol. 2016, 48, 203–213. [Google Scholar] [CrossRef] [Green Version]
- Mimouna, S.M.; Boughammoura, S.; Chemek, M.; Haouas, Z.; Banni, M.; Messaoudi, I. Disruption of the zinc metabolism in rat fœtal brain after prenatal exposure to cadmium. Chem. Biol. Interact. 2018, 286, 88–95. [Google Scholar] [CrossRef]
- Carageorgiou, H.; Tzotzes, V.; Sideris, A.; Zarros, A.; Tsakiris, S. Cadmium effects on brain acetylcholinesterase activity and antioxidant status of adult rats: Modulation by zinc, calcium and L-cysteine co-administration. Basic Clin. Pharmacol. Toxicol. 2005, 97, 320–324. [Google Scholar] [CrossRef]
- Branca, J.; Fiorillo, C.; Carrino, D.; Paternostro, F.; Taddei, N.; Gulisano, M.; Pacini, A.; Becatti, M. Cadmium-induced oxidative stress: Focus on the central nervous system. Antioxidants 2020, 9, 492. [Google Scholar] [CrossRef]
- Agnihotri, S.K.; Agrawal, U.; Ghosh, I. Brain most susceptible to cadmium induced oxidative stress in mice. J. Trace Elem. Med. Biol. 2015, 30, 184–193. [Google Scholar] [CrossRef]
- El-Sokkary, R.; Awadalla, E. The protective role of vitamin C against cerebral and pulmonary damage induced by cadmium chloride in male adult albino rat. Open Neuroendocrinol. J. 2011, 411, 1–8. [Google Scholar] [CrossRef]
- Radhakrishnan, S. Protective role of silibinin in cadmium induced changes of acetylcholinesterase, ATPases and oxidative stress in brain of albino Wistar rats. J. Ecobiotechnol. 2011, 3, 34–39. [Google Scholar]
- Kumar, R.; Agarwal, A.K.; Seth, P.K. Oxidative stress-mediated neurotoxicity of cadmium. Toxicol. Lett. 1996, 89, 65–69. [Google Scholar] [CrossRef]
- Pari, L.; Ponnusamy, M. Diallyl tetrasulfide improves cadmium induced alterations of acetylcholinesterase, ATPases and oxidative stress in brain of rats. Toxicology 2007, 234, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Saleh, H.M.; El-Sayed, Y.S.; Naser, S.M.; Eltahawy, A.S.; Onoda, A.; Umezawa, M. Efficacy of α-lipoic acid against cadmium toxicity on metal ion and oxidative imbalance, and expression of metallothionein and antioxidant genes in rabbit brain. Environ. Sci. Pollut. Res. 2017, 24, 24593–24601. [Google Scholar] [CrossRef]
- Javorac, D.; Đorđević, A.B.; Anđelković, M.; Tatović, S.; Baralić, K.; Antonijević, E.; Kotur-Stevuljević, J.; Đukić-Ćosić, D.; Antonijević, B.; Bulat, Z. Redox and essential metal status in the brain of Wistar rats acutely exposed to a cadmium and lead mixture. Arh. Hig. Rada Toksikol. 2020, 71, 197–204. [Google Scholar] [CrossRef]
- Cobbina, S.J.; Chen, Y.; Zhou, Z.; Wu, X.; Feng, W.; Wang, W.; Li, Q.; Zhao, T.; Mao, G.; Wu, X.; et al. Interaction of four low dose toxic metals with essential metals in brain, liver and kidneys of mice on sub-chronic exposure. Environ. Toxicol. Pharmacol. 2015, 39, 280–291. [Google Scholar] [CrossRef]
- Tobwala, S.; Wang, H.-J.; Carey, J.W.; Banks, W.A.; Ercal, N. Effects of lead and cadmium on brain endothelial cell survival, monolayer permeability, and crucial oxidative stress markers in an in vitro model of the blood-brain barrier. Toxics 2014, 2, 258–275. [Google Scholar] [CrossRef] [Green Version]
- Branca, J.J.V.; Maresca, M.; Morucci, G.; Mello, T.; Becatti, M.; Pazzagli, L.; Colzi, I.; Gonnelli, C.; Carrino, D.; Paternostro, F.; et al. Effects of cadmium on ZO-1 tight junction integrity of the blood brain barrier. Int. J. Mol. Sci. 2019, 20, 6010. [Google Scholar] [CrossRef] [Green Version]
- Mężyńska, M.; Brzóska, M.M.; Rogalska, J.; Galicka, A. Extract from Aronia melanocarpa L. berries protects against cadmium-induced lipid peroxidation and oxidative damage to proteins and DNA in the liver: A study in a rat model of environmental human exposure to this xenobiotic. Nutrients 2019, 11, 758. [Google Scholar] [CrossRef] [Green Version]
- Leutner, S.; Eckert, A.; Müller, W.E. ROS generation, lipid peroxidation and antioxidant enzyme activities in the aging brain. J. Neural. Transm. 2001, 108, 955–967. [Google Scholar] [CrossRef]
- Aebi, H.E. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Meth. Enzymol. 1978, 52, 302–310. [Google Scholar]
- Reznick, A.Z.; Packer, L. Oxidative damage to proteins: Spectrophotometric method for carbonyl assay. Methods Enzymol. 1994, 233, 357–363. [Google Scholar]
- Kozłowska, M.; Brzóska, M.M.; Rogalska, J.; Galicka, A. The impact of a polyphenol-rich extract from the berries of Aronia melanocarpa L. on collagen metabolism in the liver: A study in an in vivo model of human environmental exposure to cadmium. Nutrients 2020, 12, 2766. [Google Scholar] [CrossRef]
- Brzóska, M.M.; Gałażyn-Sidorczuk, M.; Jurczuk, M.; Tomczyk, M. Protective effect of Aronia melanocarpa polyphenols on cadmium accumulation in the body: A study in a rat model of human exposure to this metal. Curr. Drug Targets 2015, 16, 1470–1487. [Google Scholar] [CrossRef]
- Mężyńska, M.; Brzóska, M.M.; Rogalska, J.; Piłat-Marcinkiewicz, B. Extract from Aronia melanocarpa L. berries prevents cadmium-induced oxidative stress in the liver: A study in a rat model of low-level and moderate lifetime human exposure to this toxic metal. Nutrients 2019, 11, 21. [Google Scholar] [CrossRef] [Green Version]
- Đukić-Ćosić, D.; Baralić, K.; Javorac, D.; Buha, A.; Bulat, Z. An overview of molecular mechanisms in cadmium toxicity. Curr. Opin. Toxicol. 2019, 19, 56–62. [Google Scholar] [CrossRef]
- Williams, L.R. Oxidative stress, age-related neurodegeneration, and the potential for neurotrophic treatment. Cerebrovasc. Brain Metab. Rev. 1995, 7, 55–73. [Google Scholar]
- Borowska, S.; Brzóska, M.M.; Gałażyn-Sidorczuk, M.; Rogalska, J. Effect of an extract from Aronia melanocarpa L. berries on the body status of zinc and copper under chronic exposure to cadmium: An in vivo experimental study. Nutrients 2017, 9, 1374. [Google Scholar] [CrossRef] [Green Version]
- GBD 2016 Neurology Collaborators. Global, regional, and national burden of neurological disorders, 1990-2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 459–480. [Google Scholar] [CrossRef] [Green Version]
- Singla, N.; Dhawan, D.K. Zinc improves cognitive and neuronal dysfunction during aluminium-induced neurodegeneration. Mol. Neurobiol. 2017, 54, 406–422. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.; Magdy, M.; Ahmed, E.; Omar, H. The protective effects of vitamin E and zinc supplementation against lithium-induced brain toxicity of male albino rats. Environ. Pollut. 2014, 4. [Google Scholar] [CrossRef] [Green Version]
- Prasanthi, R.J.; Devi, C.B.; Basha, D.C.; Reddy, N.S.; Reddy, G.R. Calcium and zinc supplementation protects lead (Pb)-induced perturbations in antioxidant enzymes and lipid peroxidation in developing mouse brain. Int. J. Dev. Neurosci. 2010, 28, 161–167. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
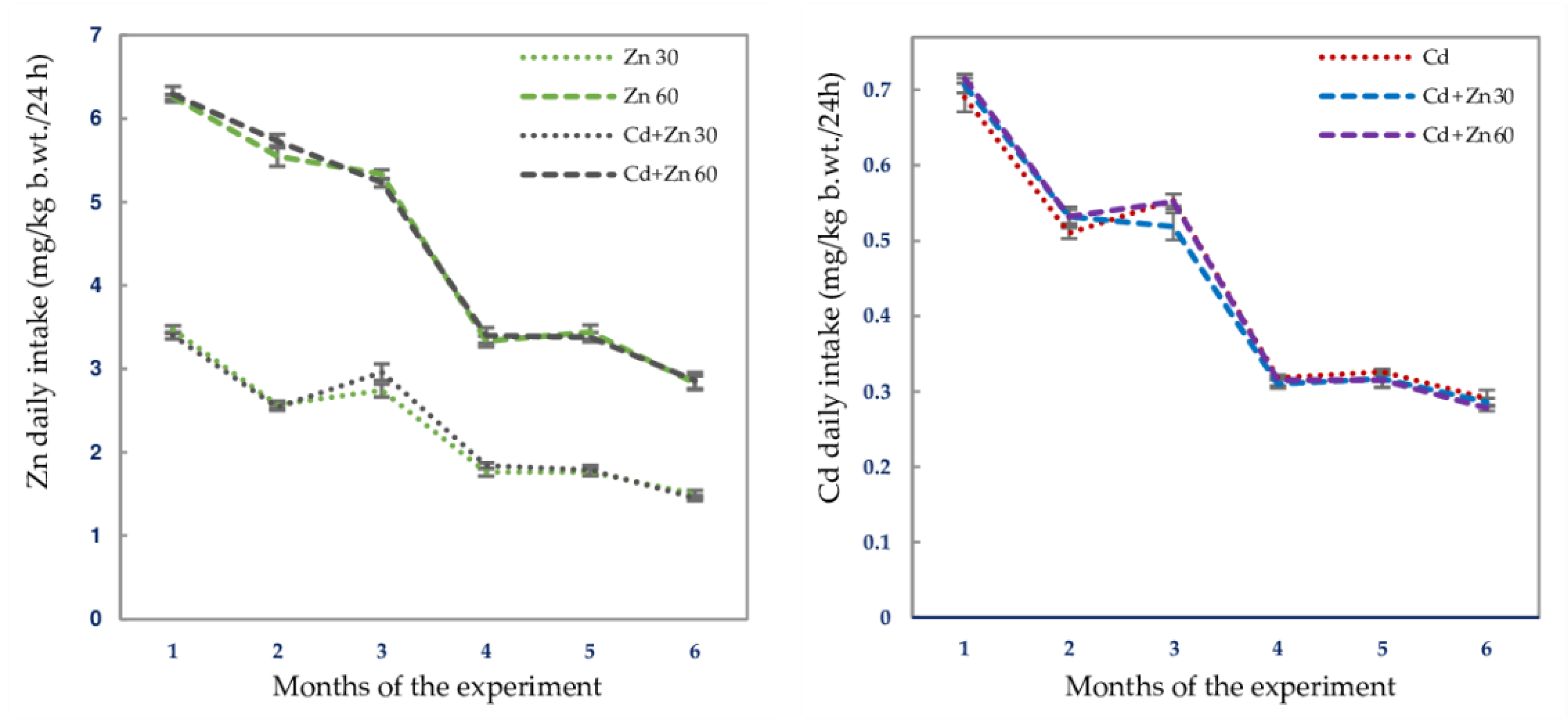
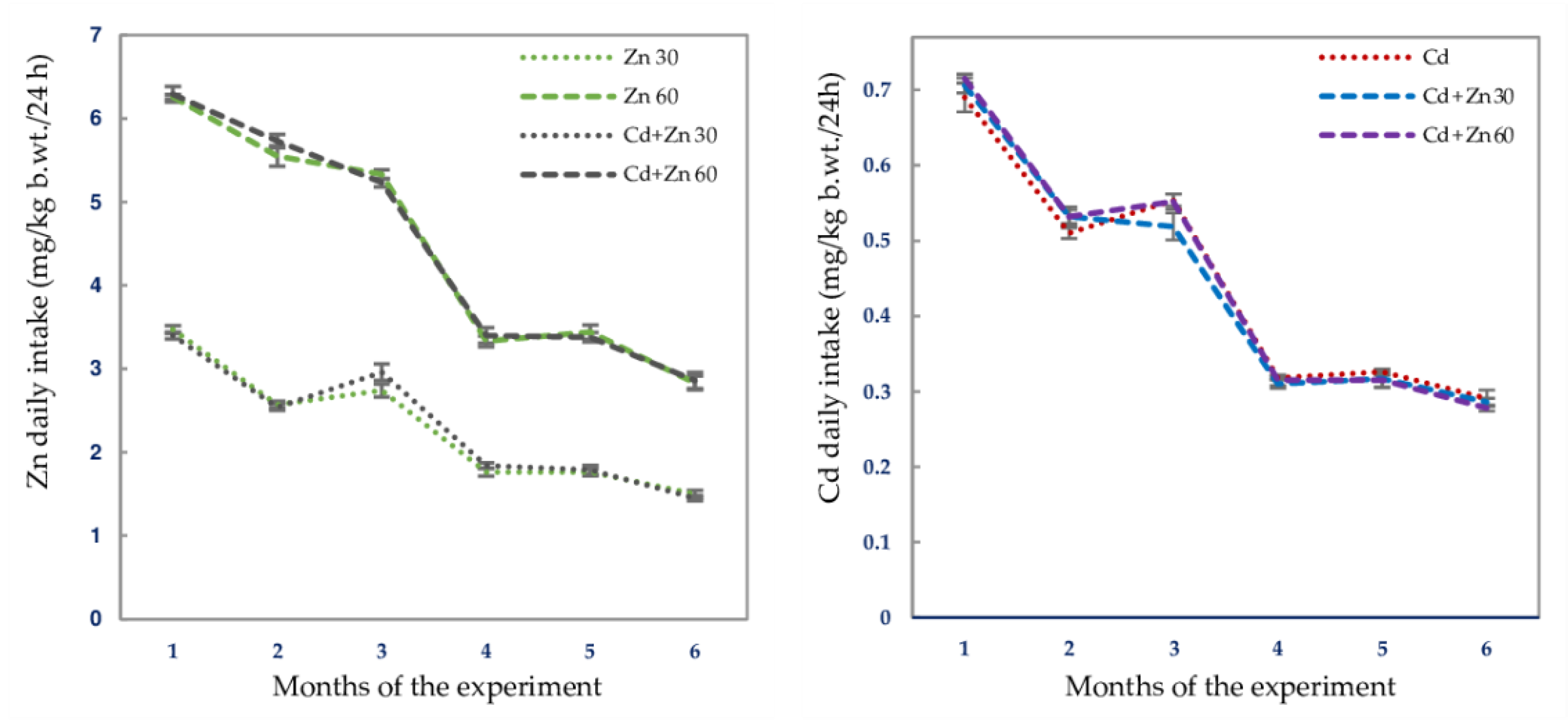
| Experimental Group | Intake of Zn Mean/Range (mg/kg b.w./24 h) | Intake of Cd Mean/Range (mg/kg b.w./24 h) |
|---|---|---|
| Control | 0 | 0 |
| Zn 30 | 2.210 ± 0.083/1.260–3.670 3 | 0 |
| Zn 60 | 4.460 ± 0.649/2.400–6.410 | 0 |
| Cd | 0 | 0.436 ± 0.019/0.222–0.731 |
| Cd + Zn 30 | 2.230 ± 0.085/1.330–3.570 | 0.423 ± 0.019/0.243–0.745 |
| Cd + Zn 60 | 4.490 ± 0.166/2.410–6.670 | 0.432 ± 0.020/0.260–0.740 |
| Parameter | 5 mg Cd/L + 30 mg Zn/L | 5 mg Cd/L + 60 mg Zn/L | ||||||
|---|---|---|---|---|---|---|---|---|
| Main Effect of Cd | Main Effect of Zn | Interactive Effect of Cd + Zn | Cd + Zn Effect vs. Cd Effect + Zn Effect Character of Cd-Zn Interaction | Main Effect of Cd | Main Effect of Zn | Interactive Effect of Cd + Zn | Cd + Zn Effect vs. Cd Effect + Zn Effect Character of Cd-Zn Interaction | |
| SOD | 17.47 ‡ | NS | NS | No interaction | NS | 19.68 ‡ | 10.17 † | +24 3 vs. −29 + 0 +24 vs. −29 4 |
| GPx | 8.048 † | NS | NS | No interaction | NS | 8.596 † | 5.412 * | 0 vs. −30 + 0 0 vs. −30 Antagonism |
| GR | 27.33 ‡ | 35.80 ‡ | 16.81 ‡ | 0 vs. +90 + 0 0 vs. +90 Antagonism | 4.274 * | 30.93 ‡ | 43.30 ‡ | −36 vs. +90 + 0 −36 vs. +90 4 |
| CAT | 13.99 ‡ | 7.307 * | 5.000 * | 0 vs. −36 + 0 0 vs. −36 Antagonism | 19.49 ‡ | 9.254 † | NS | No interaction |
| Parameter | 5 mg Cd/L + 30 mg Zn/L | 5 mg Cd/L + 60 mg Zn/L | ||||||
|---|---|---|---|---|---|---|---|---|
| Main Effect of Cd | Main Effect of Zn | Interactive Effect of Cd + Zn | Cd + Zn Effect vs. Cd Effect + Zn Effect Character of Cd-Zn Interaction | Main Effect of Cd | Main Effect of Zn | Interactive Effect of Cd + Zn | Cd + Zn Effect vs. Cd Effect + Zn Effect Character of Cd-Zn Interaction | |
| TSH | 69.10 ‡ | 31.31 ‡ | 49.57 ‡ | −48 3 vs. −53 + (−44) −48 vs. −97 Antagonism | 15.94 ‡ | 5.051 * | 49.70 ‡ | −30 vs. −53 + (−45) −30 vs. −98 Antagonism |
| GSH | 4.443 * | 4.468 * | 21.22 ‡ | −22 vs. −35 + (−35) −22 vs. −70 Antagonism | 27.35 ‡ | NS | NS | No interaction |
| GSSG | NS | 124.4 ‡ | 62.07 ‡ | −37 vs. +31 + (−12) −37 vs. +19 4 | 7.579 * | 24.69 ‡ | 10.20 † | 0 vs. +31 + 0 0 vs. +31 Antagonism |
| GSH/GSSG | NS | 21.98 ‡ | 102.3 ‡ | +24 vs. −51 + (−27) +24 vs. −78 4 | 29.43 ‡ | 8.425 † | 4.364 * | 0 vs. −51 + 0 0 vs. −51 Antagonism |
| Parameter | 5 mg Cd/L + 30 mg Zn/L | 5 mg Cd/L + 60 mg Zn/L | ||||||
|---|---|---|---|---|---|---|---|---|
| Main Effect of Cd | Main Effect of Zn | Interactive Effect of Cd + Zn | Cd + Zn Effect vs. Cd Effect + Zn Effect Character of Cd-Zn Interaction | Main Effect of Cd | Main Effect of Zn | Interactive Effect of Cd + Zn | Cd + Zn Effect vs. Cd Effect + Zn Effect Character of Cd-Zn Interaction | |
| H2O2 | 29.81 ‡ | 20.42 ‡ | 63.25 ‡ | 0 vs. +60 3 + 0 0 vs. +60 Antagonism | 50.14 ‡ | 10.95 † | 36.89 ‡ | 0 vs. +60 + 0 0 vs. +60 Antagonism |
| MPO | 19.59 ‡ | 71.50 ‡ | 51.54 ‡ | −13 vs. +37 + 0 −13 vs. +37 4 | NS | 73.27 ‡ | 75.84 ‡ | −27 vs. +37 + 0 −27 vs. +37 4 |
| Parameter | 5 mg Cd/L + 30 mg Zn/L | 5 mg Cd/L + 60 mg Zn/L | ||||||
|---|---|---|---|---|---|---|---|---|
| Main Effect of Cd | Main Effect of Zn | Interactive Effect of Cd + Zn | Cd + Zn Effect vs. Cd Effect + Zn Effect Character of Cd-Zn Interaction | Main Effect of Cd | Main Effect of Zn | Interactive Effect of Cd + Zn | Cd + Zn Effect vs. Cd Effect + Zn Effect Character of Cd-Zn Interaction | |
| BRAIN | ||||||||
| TAS | 10.55 † | NS | NS | No interaction | 5.021 * | 6.334 * | 6.379 * | 0 vs. −20 3 + 0 0 vs. −20 Antagonism |
| TOS | 65.67 ‡ | 39.45 ‡ | 53.86 ‡ | 0 vs. +3.8-fold + 0 0 vs. +3.8-fold Antagonism | 64.09 ‡ | 31.35 ‡ | 49.61 ‡ | +46 vs. +3.8-fold + 0 +46 vs. 3.8-fold Antagonism |
| OSI | 46.66 ‡ | 20.80 ‡ | 31.89 ‡ | +73 vs. +5-fold + 0 +73 vs. +5-fold Antagonism | 46.78 ‡ | 24.30 ‡ | 32.73 ‡ | +61 vs. +5-fold + 0 +61 vs. +5-fold Antagonism |
| SERUM | ||||||||
| TAS | NS | 27.32 ‡ | NS | No interaction | NS | 37.50 ‡ | 5.042 * | +38 vs. 0 + 0 +38 vs. 0 4 |
| TOS | 27.30 ‡ | 114.9 ‡ | NS | No interaction | 12.88 † | 198.6 ‡ | 22.74 ‡ | +63 vs. +50 + (−56) +63 vs. −6 4 |
| OSI | 17.56 ‡ | 74.10 ‡ | 8.103 † | −54 vs. +84 + (−70) −54 vs. +14 4 | 10.77 † | 91.43 ‡ | 16.16 ‡ | −74 vs. +84 + (−65) −74 vs. +19 4 |
| Parameter | 5 mg Cd/L + 30 mg Zn/L | 5 mg Cd/L + 60 mg Zn/L | ||||||
|---|---|---|---|---|---|---|---|---|
| Main Effect of Cd | Main Effect of Zn | Interactive Effect of Cd + Zn | Cd + Zn Effect vs. Cd Effect + Zn Effect Character of Cd-Zn Interaction | Main Effect of Cd | Main Effect of Zn | Interactive Effect of Cd + Zn | Cd + Zn Effect vs. Cd Effect + Zn Effect Character of Cd-Zn Interaction | |
| LPO | 17.81 ‡ | 11.69 † | 11.37 † | 0 vs. +72 3 + 0 0 vs. +72 Antagonism | 18.47 ‡ | 16.56 ‡ | 10.39 † | 0 vs. +72 + 0 0 vs. +72 Antagonism |
| MDA | 12.42 † | 45.01 ‡ | 20.75 ‡ | 0 vs. +46 + 0 0 vs. +46 Antagonism | 3.824 # | 43.68 ‡ | 20.19 ‡ | 0 vs. +46 + 0 0 vs. +46 Antagonism |
| 8-isoprostane | 7.857 † | 9.644 † | 13.79 ‡ | 0 vs. +59 + 0 0 vs. +59 Antagonism | 10.08 † | 4.457 * | 16.65 † | 0 vs. +59 + 0 0 vs. +59 Antagonism |
| PC | 6.302 * | 12.36 † | 15.39 ‡ | 0 vs. +54 + 0 0 vs. +54 Antagonism | 8.679 † | 35.31 ‡ | 18.31 ‡ | −22 vs. +54 + 0 −22 vs. +54 4 |
| Indices of Antioxidative Status | Indices of Oxidative Status | Markers of Oxidative Damage to the Cellular Macromolecules | |||
|---|---|---|---|---|---|
| Parameter | r p | Parameter | r p | Parameter | r p |
| SOD | NS | H2O2 | 0.344 * | LPO | NS |
| CAT | −0.407 † | MPO | NS | MDA | NS |
| GPx | NS | TOS | 0.525 ‡ | 8-isoprostane | 0.365 * |
| GR | NS | OSI | 0.388 † | PC | NS |
| TSH | −0.364 * | ||||
| PSH | NS | ||||
| GSH | NS | ||||
| GSSG | 0.356 * | ||||
| GSH/GSSG | −0.335 * | ||||
| TAS | NS | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brzóska, M.M.; Kozłowska, M.; Rogalska, J.; Gałażyn-Sidorczuk, M.; Roszczenko, A.; Smereczański, N.M. Enhanced Zinc Intake Protects against Oxidative Stress and Its Consequences in the Brain: A Study in an In Vivo Rat Model of Cadmium Exposure. Nutrients 2021, 13, 478. https://doi.org/10.3390/nu13020478
Brzóska MM, Kozłowska M, Rogalska J, Gałażyn-Sidorczuk M, Roszczenko A, Smereczański NM. Enhanced Zinc Intake Protects against Oxidative Stress and Its Consequences in the Brain: A Study in an In Vivo Rat Model of Cadmium Exposure. Nutrients. 2021; 13(2):478. https://doi.org/10.3390/nu13020478
Chicago/Turabian StyleBrzóska, Małgorzata M., Magdalena Kozłowska, Joanna Rogalska, Małgorzata Gałażyn-Sidorczuk, Alicja Roszczenko, and Nazar M. Smereczański. 2021. "Enhanced Zinc Intake Protects against Oxidative Stress and Its Consequences in the Brain: A Study in an In Vivo Rat Model of Cadmium Exposure" Nutrients 13, no. 2: 478. https://doi.org/10.3390/nu13020478