Melatonin Supplementation Attenuates the Pro-Inflammatory Adipokines Expression in Visceral Fat from Obese Mice Induced by A High-Fat Diet



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Melatonin Supplementation
2.2. Glucose and Insulin Tolerance Tests
2.3. Adipocyte Isolation
2.4. Blood Measurements
2.5. RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.6. Adipokine Measurements
2.7. Statistical Analysis
3. Results
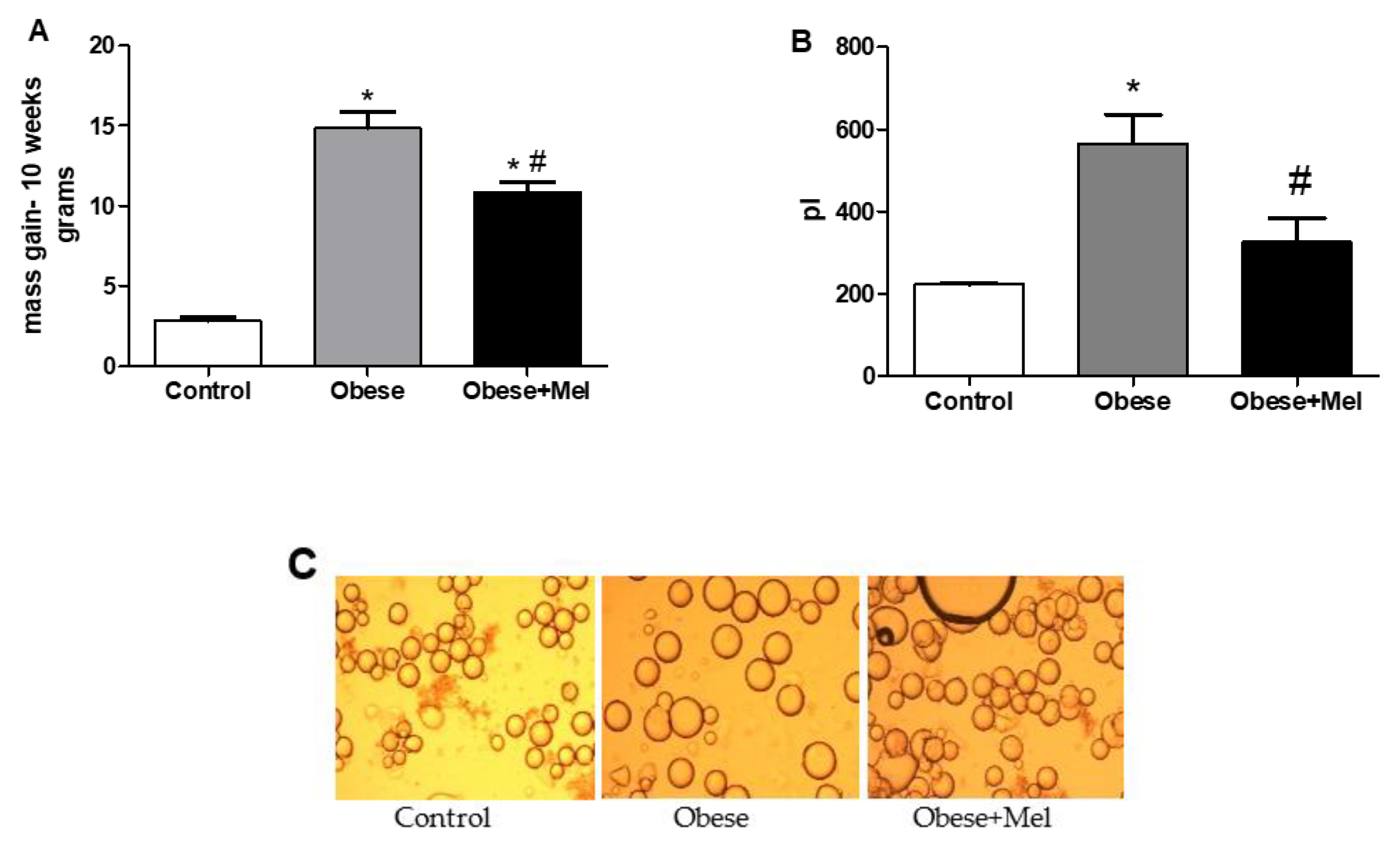
3.1. Melatonin Supplementation Decreased Body Mass (BM), Adipose Depots Mass, Adipocytes Hypertrophy and Blood Biochemical Parameters Triggered by HFD-Induced Obesity
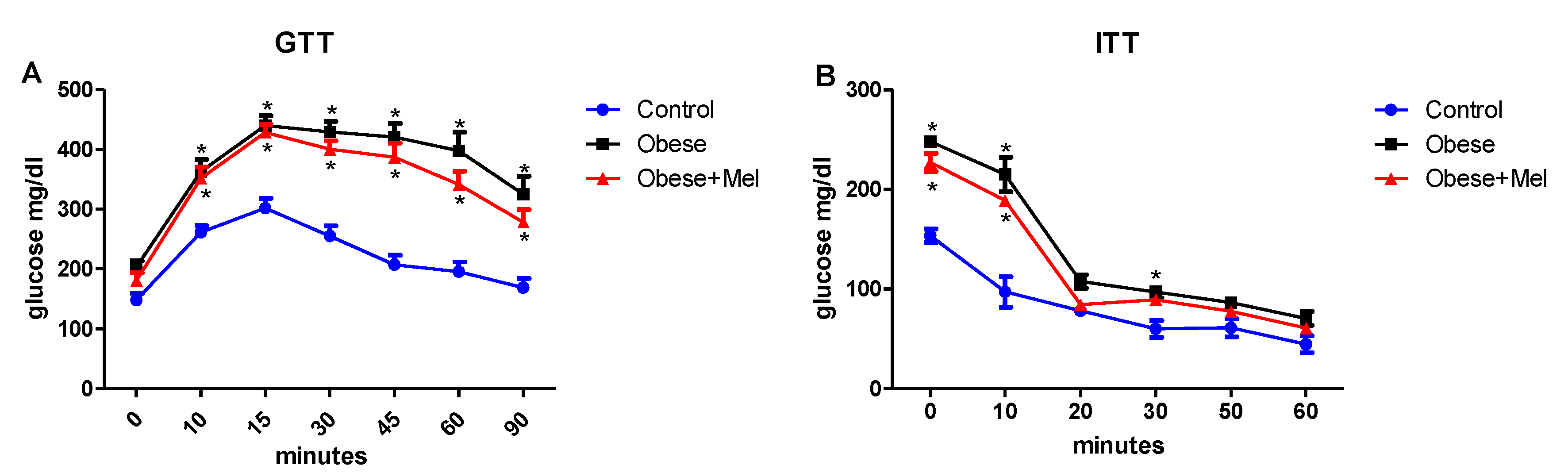
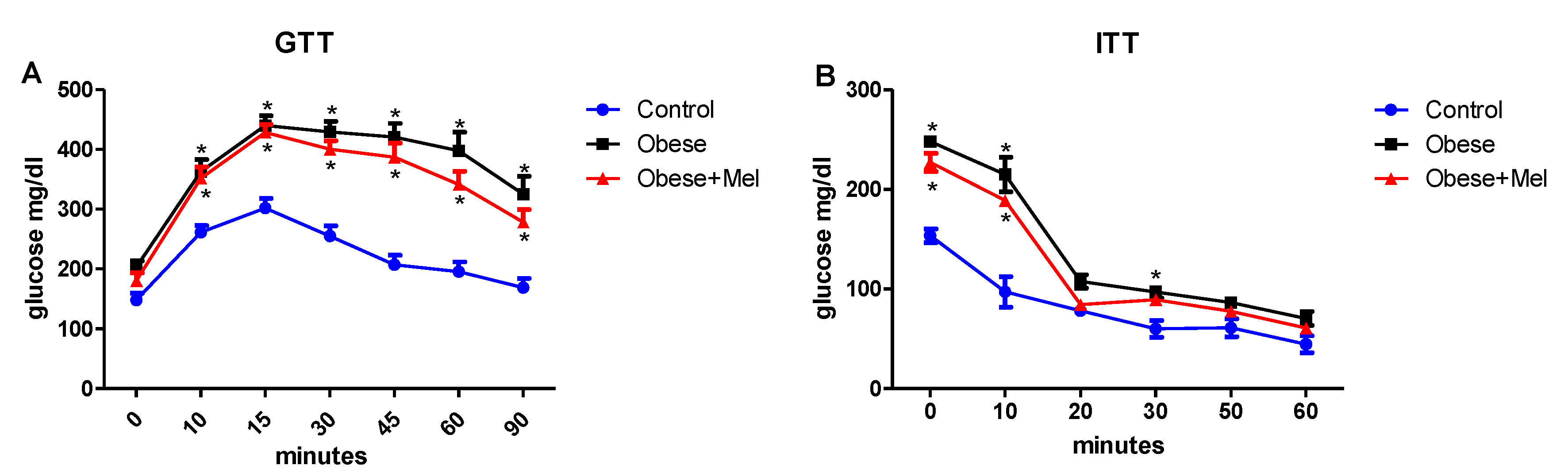
3.2. Melatonin Supplementation Did Not Alter Glycemic Curve after Glucose and Insulin Tolerance Test (GTT and ITT Test)
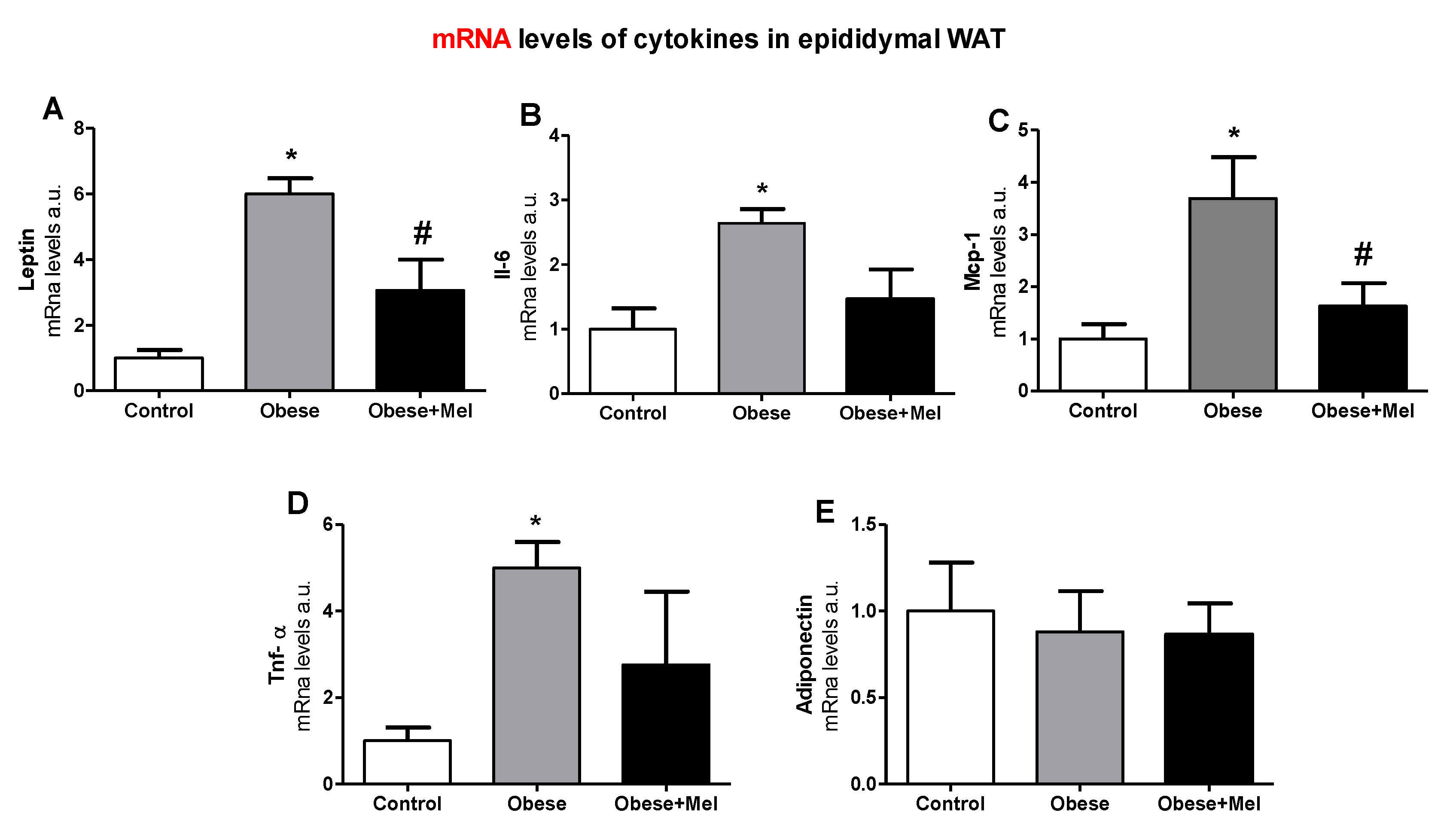
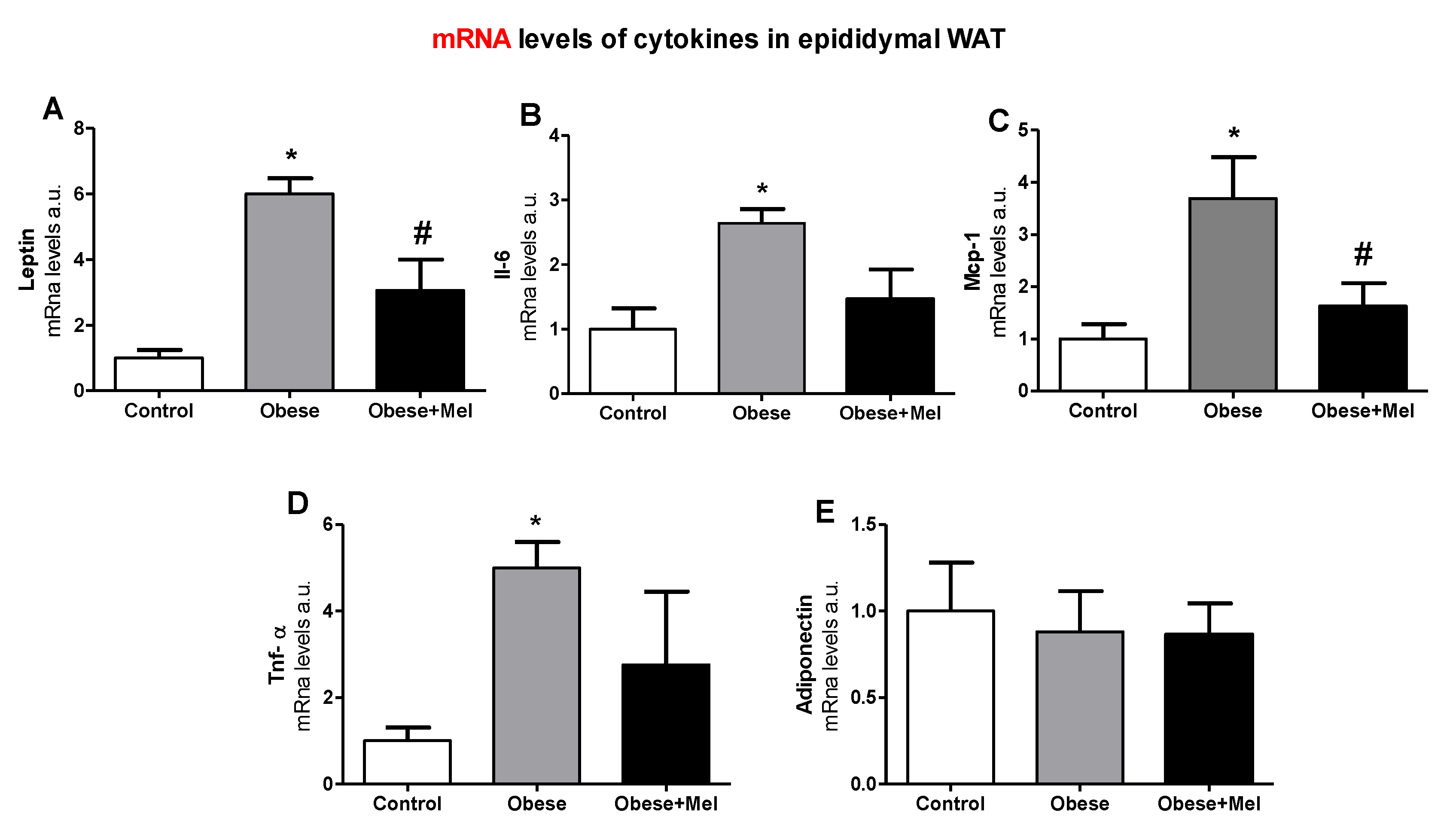
3.3. Melatonin Supplementation Decreased the Gene Expression of Inflammatory Cytokines on EPI Depot
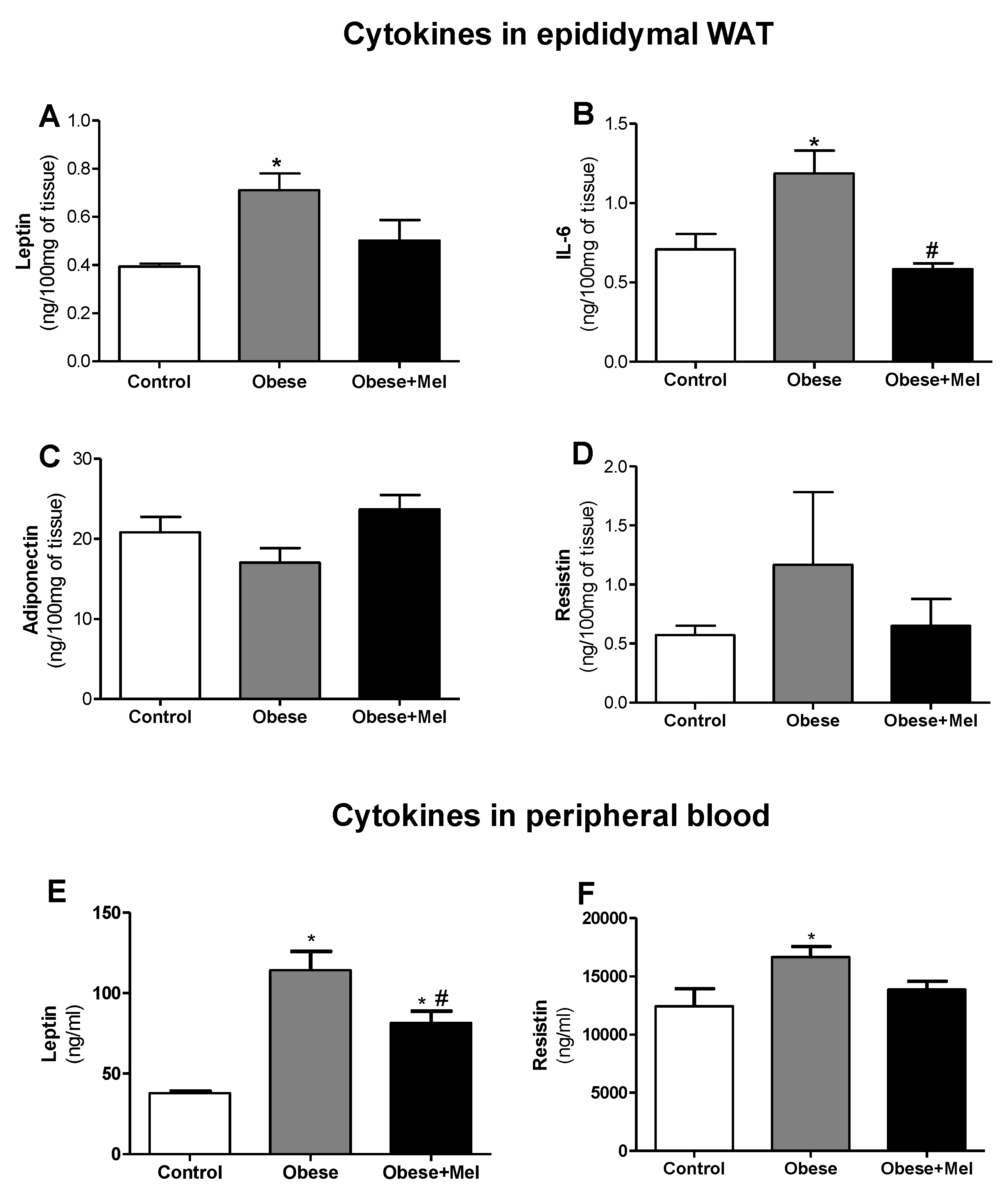
3.4. Melatonin Supplementation Reduced the Protein Expression of Inflammatory Cytokines on EPI Depot and Peripheral Bood
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef]
- Xu, H.; Barnes, G.T.; Yang, Q.; Tan, G.; Yang, D.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A.; et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Investig. 2003, 112, 1821–1830. [Google Scholar] [CrossRef]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef]
- Lancha, A.; Frühbeck, G.; Gómez-Ambrosi, J. Peripheral signalling involved in energy homeostasis control. Nutr. Res. Rev. 2012, 25, 223–248. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Ambrosi, J.; Catalán, V.; Rodríguez, A.; Andrada, P.; Ramírez, B.; Ibanez, P.; Vilà, N.; Romero, S.; Margall, M.A.; Gil, M.J.; et al. Increased Cardiometabolic Risk Factors and Inflammation in Adipose Tissue in Obese Subjects Classified as Metabolically Healthy. Diabetes Care 2014, 37, 2813–2821. [Google Scholar] [CrossRef] [Green Version]
- Fasshauer, M.; Blüher, M. Adipokines in health and disease. Trends Pharmacol Sci. 2015, 36, 461–470. [Google Scholar] [CrossRef]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef]
- Crewe, C.; An, Y.A.; Scherer, P.E. The ominous triad of adipose tissue dysfunction: inflammation, fibrosis, and impaired angiogenesis. J. Clin. Investig. 2017, 127, 74–82. [Google Scholar] [CrossRef] [Green Version]
- McNelis, J.C.; Olefsky, J.M. Macrophages, Immunity, and Metabolic Disease. Immunity 2014, 41, 36–48. [Google Scholar] [CrossRef] [Green Version]
- Szewczyk-Golec, K.; Wozniak, A.; Reiter, R.J. Inter-relationships of the chronobiotic, melatonin, with leptin and adiponectin: implications for obesity. J. Pineal Res. 2015, 59, 277–291. [Google Scholar] [CrossRef]
- Yadav, A.; Kataria, M.A.; Saini, V.; Yadav, A. Role of leptin and adiponectin in insulin resistance. Clin. Chim. Acta 2013, 417, 80–84. [Google Scholar] [CrossRef]
- López-Jaramillo, P.; Gomez-Arbelaez, D.; López-López, J.; Lopez-Lopez, C.; Martínez-Ortega, J.; Gómez-Rodríguez, A.; Triana-Cubillos, S. The role of leptin/adiponectin ratio in metabolic syndrome and diabetes. Horm. Mol. Boil. Clin. Investig. 2014, 18. [Google Scholar] [CrossRef]
- Bastard, J.-P.; Maachi, M.; Lagathu, C.; Kim, M.J.; Caron, M.; Vidal, H.; Capeau, J.; Feve, B. Recent advances in the relationship between obesity, inflammation, and insulin resistance. Eur. Cytokine Netw. 2006, 17. [Google Scholar]
- Greenberg, A.S.; Obin, M.S. Obesity and the role of adipose tissue in inflammation and metabolism. Am. J. Clin. Nutr. 2006, 83, 461S–465S. [Google Scholar] [CrossRef] [Green Version]
- Lima, F.B.; Matsushita, D.H.; Hell, N.S.; Dolnikoff, M.S.; Okamoto, M.M.; Neto, J.C. The regulation of insulin action in isolated adipocytes. Role of the periodicity of food intake, time of day and melatonin. Braz. J. Med Boil. Res. 1994, 27. [Google Scholar]
- Lima, F.B.; Machado, U.F.; Bartol, I.; Seraphim, P.M.; Sumida, D.H.; Moraes, S.M.F.; Hell, N.S.; Okamoto, M.M.; Saad, M.J.A.; Carvalho, C.R.O.; et al. Pinealectomy causes glucose intolerance and decreases adipose cell responsiveness to insulin in rats. Am. J. Physiol. Metab. 1998, 275. [Google Scholar] [CrossRef]
- Zanquetta, M.M.; Corrêa-Giannella, M.L.; Monteiro, M.B.; Villares, S.M. Body weight, metabolism and clock genes. Diabetol. Metab. Syndr. 2010, 2, 53. [Google Scholar] [CrossRef]
- Picinato, M.C.; Haber, E.P.; Carpinelli, A.R.; Cipolla-Neto, J.; Cipolla-Neto, J. Daily rhythm of glucose-induced insulin secretion by isolated islets from intact and pinealectomized rat. J. Pineal Res. 2002, 33, 172–177. [Google Scholar] [CrossRef]
- Alonso-Vale, M.I.C.; Andreotti, S.; Peres, S.B.; Anhê, G.F.; Borges-Silva, C.D.N.; Neto, J.C.; Lima, F.B.; Alonso-Vale, M.I.C. Melatonin enhances leptin expression by rat adipocytes in the presence of insulin. Am. J. Physiol. Metab. 2005, 288, E805–E812. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Vale, M.I.; Borges-Silva, C.N.; Anhe, G.F.; Andreotti, S.; Machado, M.A.; Cipolla-Neto, J.; Lima, F.B. Light/Dark Cycle-dependent Metabolic Changes in Adipose Tissue of Pinealectomized Rats. Horm. Metab. Res. 2004, 36, 474–479. [Google Scholar] [CrossRef]
- Takada, J.; Peres, S.B.; Andreotti, S.; Lima, F.B.; Borges-Silva, C.D.N.; Alonso-Vale, M.I.C.; Fonseca-Alaniz, M.H.; Cipolla-Neto, J.; Pithon-Curi, T.C.; Borges-Silva, C.D.N.; et al. Pinealectomy reduces hepatic and muscular glycogen content and attenuates aerobic power adaptability in trained rats. J. Pineal Res. 2007, 43, 96–103. [Google Scholar]
- She, M.; Deng, X.; Guo, Z.; Laudon, M.; Hu, Z.; Liao, D.; Hu, X.; Luo, Y.; Shen, Q.; Su, Z. NEU-P11, a novel melatonin agonist, inhibits weight gain and improves insulin sensitivity in high-fat/high-sucrose-fed rats. Pharmacol. Res. 2009, 59, 248–253. [Google Scholar] [CrossRef]
- Alamdari, N.M.; Mahdavi, R.; Roshanravan, N.; Yaghin, N.L.; Ostadrahimi, A.R.; Faramarzi, E. A double-blind, placebo-controlled trial related to the effects of melatonin on oxidative stress and inflammatory parameters of obese women. Horm Metab Res. 2015, 47, 504–508. [Google Scholar] [CrossRef]
- Farias, T.D.S.M.D.; Chimin, P.; De Oliveira, A.C.; De Proença, A.R.G.; Leal, F.L.T.; Andreotti, S.; Lopes, A.B.; De Sousa, A.H.; Amaral, F.G.; Sertié, R.A.L.; et al. Pinealectomy interferes with the circadian clock genes expression in white adipose tissue. J. Pineal Res. 2015, 58, 251–261. [Google Scholar] [CrossRef]
- de Sá, R.D.; Crisma, A.R.; Cruz, M.M.; Martins, A.R.; Masi, L.N.; do Amaral, C.L.; Curi, R.; Alonso-Vale, M.I. Fish oil prevents changes induced by a high-fat diet on metabolism and adipokine secretion in mice subcutaneous and visceral adipocytes. J. Physiol. 2016, 594, 6301–6317. [Google Scholar] [CrossRef]
- De Oliveira, A.C.; Andreotti, S.; Farias, T.D.S.; Torres-Leal, F.L.; De Proença, A.R.; Campaña, A.B.; De Souza, A.H.; Sertié, R.A.; Carpinelli, A.R.; Cipolla-Neto, J.; et al. Metabolic Disorders and Adipose Tissue Insulin Responsiveness in Neonatally STZ-Induced Diabetic Rats Are Improved by Long-Term Melatonin Treatment. Endocrinology 2012, 153, 2178–2188. [Google Scholar] [CrossRef] [Green Version]
- Rodbell, M. Metabolism of Isolated Fat Cells. I. Effects of Hormones on Glucose Metabolism and Lipolysis. J. Biol. Chem. 1964, 239, 375–380. [Google Scholar]
- Bolsoni-Lopes, A.; Festuccia, W.T.; Farias, T.S.M.; Chimin, P.; Torres-Leal, F.L.; Derogis, P.B.M.; De Andrade, P.B.; Miyamoto, S.; Lima, F.B.; Curi, R.; et al. Palmitoleic acid (n-7) increases white adipocyte lipolysis and lipase content in a PPARα-dependent manner. Am. J. Physiol. Metab. 2013, 305, E1093–E1102. [Google Scholar] [CrossRef]
- Bucolo, G.; David, H. Quantitative determination of serum triglycerides by the use of enzymes. Clin. Chem. 1973, 19, 476–482. [Google Scholar]
- Postiglione, L.; Oriente, P.; Nastasi, L.; Cimmino, F.; Di Fraia, G.; Spanò, A. Determination of serum and lipoprotein phospholipids by a new enzymatic colorimetric method: characterization of a "reference range" in a sample of clinically healthy people. Boll. della Soc. Ital. di Boil. Sper. 1982, 58, 612–617. [Google Scholar]
- Grillo, F.; Izzo, C.; Mazzotti, G.; Murador, E. Improved method for determination of high-density-lipoprotein cholesterol II. Enzymic determination of cholesterol in high-density lipoprotein fractions with a sensitive reagent. Clin. Chem. 1981, 27, 371–374. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)). Method. Methods. 2001, 25, 402–408. [Google Scholar]
- Favero, G.; Stacchiotti, A.; Castrezzati, S.; Bonomini, F.; Albanese, M.; Rezzani, R.; Rodella, L.F. Melatonin reduces obesity and restores adipokine patterns and metabolism in obese ( ob/ob ) mice. Nutr. Res. 2015, 35, 891–900. [Google Scholar] [CrossRef]
- Ríos-Lugo, M.J.; Cano, P.; Jiménez-Ortega, V.; Fernández-Mateos, M.P.; Scacchi, P.A.; Cardinali, D.P.; Esquifino, A.I.; Rios-Lugo, M.J.; Jimenez-Ortega, V.; Fernández-Mateos, M.P.; et al. Melatonin effect on plasma adiponectin, leptin, insulin, glucose, triglycerides and cholesterol in normal and high fat-fed rats. J. Pineal Res. 2010, 49, 342–348. [Google Scholar] [CrossRef]
- Halpern, B.; Mancini, M.C.; Bueno, C.; Barcelos, I.P.; De Melo, M.E.; Lima, M.S.; Carneiro, C.G.; Sapienza, M.T.; Buchpiguel, C.A.; Amaral, F.G.D.; et al. Melatonin Increases Brown Adipose Tissue Volume and Activity in Patients With Melatonin Deficiency: A Proof-of-Concept Study. Diabetes 2019, 68, 947–952. [Google Scholar] [CrossRef] [Green Version]
- Nduhirabandi, F.; Huisamen, B.; Strijdom, H.; Blackhurst, D.; Lochner, A. Short-term melatonin consumption protects the heart of obese rats independent of body weight change and visceral adiposity. J. Pineal Res. 2014, 57, 317–332. [Google Scholar] [CrossRef]
- Nogueira, T.C.; Lellis-Santos, C.; Jesus, D.S.; Taneda, M.; Rodrigues, S.C.; Amaral, F.G.; Lopes, A.M.S.; Cipolla-Neto, J.; Bordin, S.; Anhê, G.F. Absence of Melatonin Induces Night-Time Hepatic Insulin Resistance and Increased Gluconeogenesis Due to Stimulation of Nocturnal Unfolded Protein Response. Endocrinology 2011, 152, 1253–1263. [Google Scholar] [CrossRef] [Green Version]
- Zalatan, F.; Krause, J.A.; Blask, D.E. Inhibition of Isoproterenol-Induced Lipolysis in Rat Inguinal Adipocytes in Vitro by Physiological Melatonin via a Receptor-Mediated Mechanism. Endocrinology 2001, 142, 3783–3790. [Google Scholar] [CrossRef]
- Ha, E.; Yim, S.-V.; Chung, J.-H.; Yoon, K.-S.; Kang, I.; Cho, Y.H.; Baik, H.H. Melatonin stimulates glucose transport via insulin receptor substrate-1/phosphatidylinositol 3-kinase pathway in C2C12 murine skeletal muscle cells. J. Pineal Res. 2006, 41, 67–72. [Google Scholar] [CrossRef]
- Owino, S.; Sánchez-Bretaño, A.; Tchio, C.; Cecon, E.; Karamitri, A.; Dam, J.; Jockers, R.; Piccione, G.; Noh, H.L.; Kim, T.; et al. Nocturnal activation of melatonin receptor type 1 signaling modulates diurnal insulin sensitivity via regulation of PI3K activity. J. Pineal Res. 2018, 64, e12462. [Google Scholar] [CrossRef]
- Loloei, S.; Sepidarkish, M.; Heydarian, A.; Tahvilian, N.; Khazdouz, M.; Heshmati, J.; Pouraram, H. The effect of melatonin supplementation on lipid profile and anthropometric indices: A systematic review and meta-analysis of clinical trials. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 1901–1910. [Google Scholar] [CrossRef]
- Hoyos, M.; Guerrero, J.M.; Perez-Cano, R.; Olivan, J.; Fabiani, F.; Garcia-Pergañeda, A.; Osuna, C. Serum cholesterol and lipid peroxidation are decreased by melatonin in diet-induced hypercholesterolemic rats. J. Pineal Res. 2000, 28, 150–155. [Google Scholar] [CrossRef]
- Hussein, M.R.; Ahmed, O.G.; Hassan, A.F.; Ahmed, M.A. Intake of melatonin is associated with amelioration of physiological changes, both metabolic and morphological pathologies associated with obesity: An animal model. Int. J. Exp. Pathol. 2007, 88, 19–29. [Google Scholar] [CrossRef]
- Pan, M.; Song, Y.-L.; Xu, J.-M.; Gan, H.-Z.; Song, Y.; Xu, J.; Gan, H. Melatonin ameliorates nonalcoholic fatty liver induced by high-fat diet in rats. J. Pineal Res. 2006, 41, 79–84. [Google Scholar] [CrossRef]
- Hadjzadeh, M.A.R.; Alikhani, V.; Hosseinian, S.; Zarei, B.; Keshavarzi, Z. The Effect Of Melatonin Against Gastric Oxidative Stress And Dyslipidemia In Streptozotocin-Induced Diabetic Rats. Acta Endocrinol. (Buchar.) 2018, 14, 453–458. [Google Scholar] [CrossRef]
- Sun, H.; Wang, X.; Chen, J.; Song, K.; Gusdon, A.M.; Li, L.; Bu, L.; Qu, S. Melatonin improves non-alcoholic fatty liver disease via MAPK-JNK/P38 signaling in high-fat-diet-induced obese mice. Lipids Heal. Dis. 2016, 15, 202. [Google Scholar] [CrossRef]
- Franssen, R.; Monajemi, H.; Stroes, E.S.; Kastelein, J.J. Obesity and dyslipidemia. Med. Clin. North Am. 2011, 95, 893–902. [Google Scholar] [CrossRef]
- Graham, I.; Cooney, M.-T.; Bradley, D.; Dudina, A.; Reiner, Z. Dyslipidemias in the Prevention of Cardiovascular Disease: Risks and Causality. Curr. Cardiol. Rep. 2012, 14, 709–720. [Google Scholar] [CrossRef]
- Holmes, S.W.; Sugden, D. Proceedings: The effect of melatonin on pinealectomy-induced hypertension in the rat. Br. J. Pharmacol. 1976, 56, 360P–361P. [Google Scholar]
- Zanoboni, A.; Forni, A.; Zanoboni-Muciaccia, W.; Zanussi, C. Effect of pinealectomy on arterial blood pressure and food and water intake in the rat. J. Endocrinol. Investig. 1978, 1, 125–130. [Google Scholar] [CrossRef]
- Ayonrinde, O.T.; Olynyk, J.; Beilin, L.J.; Mori, T.A.; Pennell, C.E.; De Klerk, N.; Oddy, W.H.; Shipman, P.; Adams, L.A. Gender-specific differences in adipose distribution and adipocytokines influence adolescent nonalcoholic fatty liver disease. Hepatol. 2011, 53, 800–809. [Google Scholar] [CrossRef]
- Fox, C.S.; Massaro, J.M.; Hoffmann, U.; Pou, K.; Maurovich-Horvat, P.; Liu, C. Abdominal visceral and subcutaneous adipose tissue compartments: Association with metabolic risk factors in the Framingham Heart Study. J. Allergy Clin. Immunol. 2007, 116, 39–48. [Google Scholar] [CrossRef]
- Fontana, L.; Eagon, J.C.; Trujillo, M.E.; Scherer, P.E.; Klein, S. Visceral Fat Adipokine Secretion Is Associated With Systemic Inflammation in Obese Humans. Diabetes 2007, 56, 1010–1013. [Google Scholar] [CrossRef] [Green Version]
- Cancello, R.; Tordjman, J.; Poitou, C.; Guilhem, G.; Bouillot, J.L.; Hugol, D.; Coussieu, C.; Basdevant, A.; Hen, A.B.; Bedossa, P.; et al. Increased Infiltration of Macrophages in Omental Adipose Tissue Is Associated With Marked Hepatic Lesions in Morbid Human Obesity. Diabetes 2006, 55, 1554–1561. [Google Scholar] [CrossRef] [Green Version]
- Cuesta, S.; Kireev, R.; Forman, K.; García, C.; Escames, G.; Ariznavarreta, C.; Vara, E.; Tresguerres, J.A. Melatonin improves inflammation processes in liver of senescence-accelerated prone male mice (SAMP8). Exp. Gerontol. 2010, 45, 950–956. [Google Scholar] [CrossRef]
- Kohsaka, A.; Laposky, A.D.; Ramsey, K.M.; Estrada, C.; Joshu, C.; Kobayashi, Y.; Turek, F.W.; Bass, J. High-Fat Diet Disrupts Behavioral and Molecular Circadian Rhythms in Mice. Cell Metab. 2007, 6, 414–421. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, K.; Yamada, T.; Tsukita, S.; Takahashi, K.; Ishigaki, Y.; Oka, Y.; Katagiri, H. Obesity alters circadian expressions of molecular clock genes in the brainstem. Brain Res. 2009, 1263, 58–68. [Google Scholar] [CrossRef]
- Engin, A. Circadian Rhythms in Diet-Induced Obesity. Results Probl. Cell Differ. 2017, 960, 19–52. [Google Scholar]
- Pfeffer, M.; Korf, H.-W.; Wicht, H. Synchronizing effects of melatonin on diurnal and circadian rhythms. Gen. Comp. Endocrinol. 2018, 258, 215–221. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Obese | Obese + Mel | |
|---|---|---|---|
| Initial BM (g) | 22.70 ± 0.25 | 23.47 ± 0.29 | 22.56 ± 0.31 |
| Final BM (g) | 25.56 ± 0.30 | 38.22 ± 1.15* | 33.44 ± 0.66*# |
| Water intake (ml/day/mice) | 3.97 ± 0.20 | 3.08 ± 0.11* | 3.20 ± 0.08* |
| Food Intake (g/day/mice) | 4.76 ± 0.30 | 2.45 ± 0.20* | 2.34 ± 0.22* |
| Calories intake | 18.09 ± 0.32 | 13.08 ± 0.30* | 12.51 ± 0.34* |
| Fat intake (g/day/mice) | 0.43 ± 0.03 | 1.44 ± 0.12* | 1.38 ± 0.13* |
| Relative ING weight (g/100g BM) | 1.52 ± 0.07 | 4.67 ± 0.21* | 3.86 ± 0.16*# |
| Relative EPI weight (g/100g BM) | 2.04 ± 0.13 | 6.51 ± 0.20* | 5.54 ± 0.17*# |
| Relative RP weight (g/100g BM) | 0.45 ± 0.04 | 1.84 ± 0.05* | 1.70 ± 0.04* |
| Relative BAT weight (g/100g BM) | 0.25 ± 0.01 | 0.34 ± 0.02* | 0.31 ± 0.01* |
| Fasting blood glucose (mg/dl) | 175.64 ± 12.72 | 243.53 ± 8.20* | 202.08 ± 13.61# |
| Total Cholesterol (mg/dl) | 173.57 ± 13.19 | 265.50 ± 18.65* | 209.30 ± 18.38 |
| Triglycerides (mg/dl) | 83.04 ±5.34 | 129.12 ±14.55* | 94.87 ± 9.08 |
| HDL (mg/dl) | 69.56 ± 6.40 | 82.28 ±15.24 | 84.13 ± 11.62 |
| LDL (mg/dl) | 92.29 ± 9.74 | 148.34 ± 16.90* | 114.22 ± 17.92 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farias, T.d.S.M.d.; Paixao, R.I.d.; Cruz, M.M.; de Sa, R.D.C.d.C.; Simão, J.d.J.; Antraco, V.J.; Alonso-Vale, M.I.C. Melatonin Supplementation Attenuates the Pro-Inflammatory Adipokines Expression in Visceral Fat from Obese Mice Induced by A High-Fat Diet. Cells 2019, 8, 1041. https://doi.org/10.3390/cells8091041
Farias TdSMd, Paixao RId, Cruz MM, de Sa RDCdC, Simão JdJ, Antraco VJ, Alonso-Vale MIC. Melatonin Supplementation Attenuates the Pro-Inflammatory Adipokines Expression in Visceral Fat from Obese Mice Induced by A High-Fat Diet. Cells. 2019; 8(9):1041. https://doi.org/10.3390/cells8091041
Chicago/Turabian StyleFarias, Talita da Silva Mendes de, Regislane Ino da Paixao, Maysa Mariana Cruz, Roberta Dourado Cavalcante da Cunha de Sa, Jussara de Jesus Simão, Vitor Jaco Antraco, and Maria Isabel Cardoso Alonso-Vale. 2019. "Melatonin Supplementation Attenuates the Pro-Inflammatory Adipokines Expression in Visceral Fat from Obese Mice Induced by A High-Fat Diet" Cells 8, no. 9: 1041. https://doi.org/10.3390/cells8091041