
Exploring Protein Kinase CK2 Substrate Recognition and the Dynamic Response of Substrate Phosphorylation to Kinase Modulation




Abstract
:1. Introduction
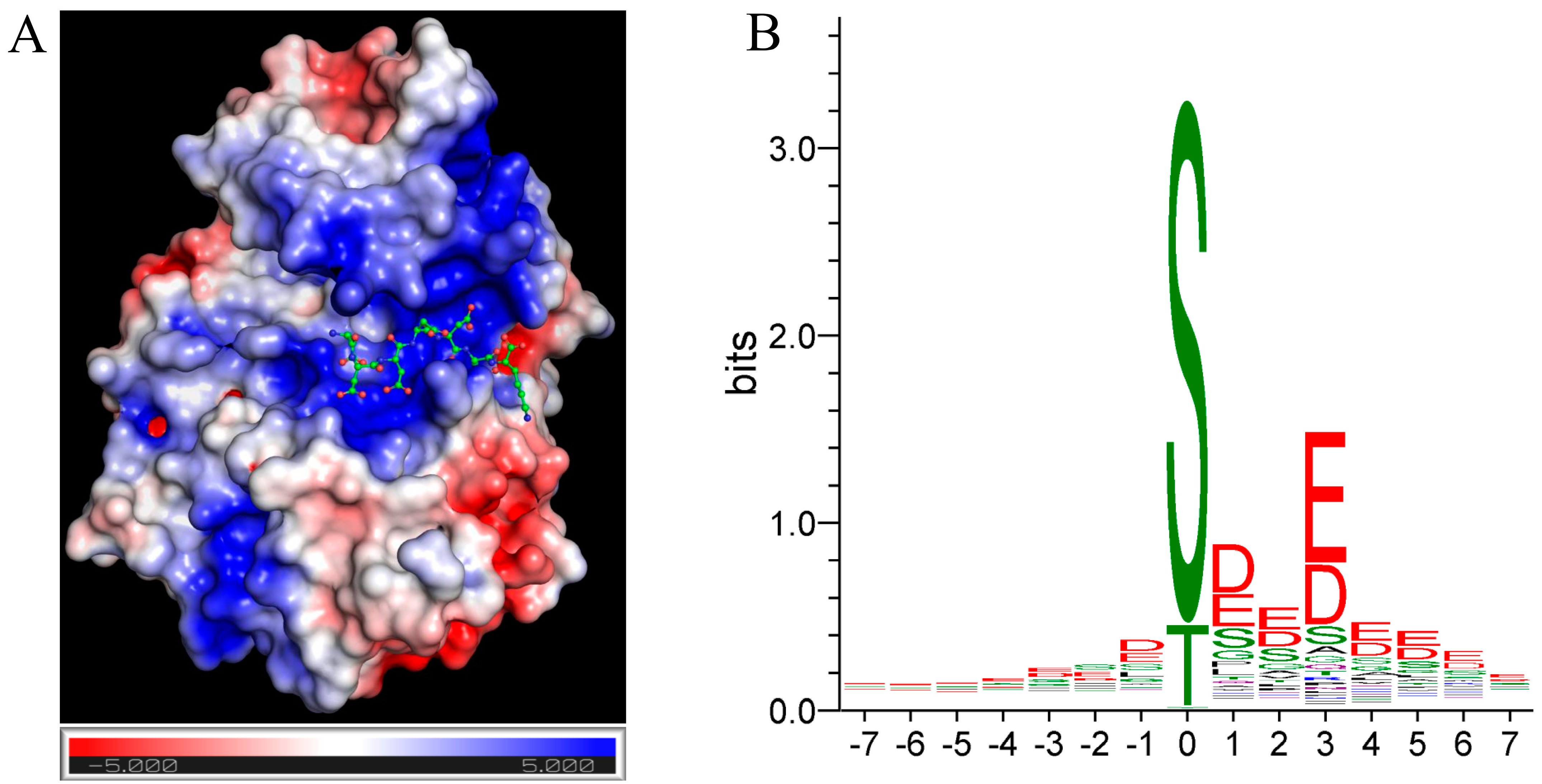
2. CK2 Consensus Sequence
3. Is a Phosphopeptide Containing the CK2 Consensus Sequence a True CK2 Substrate?
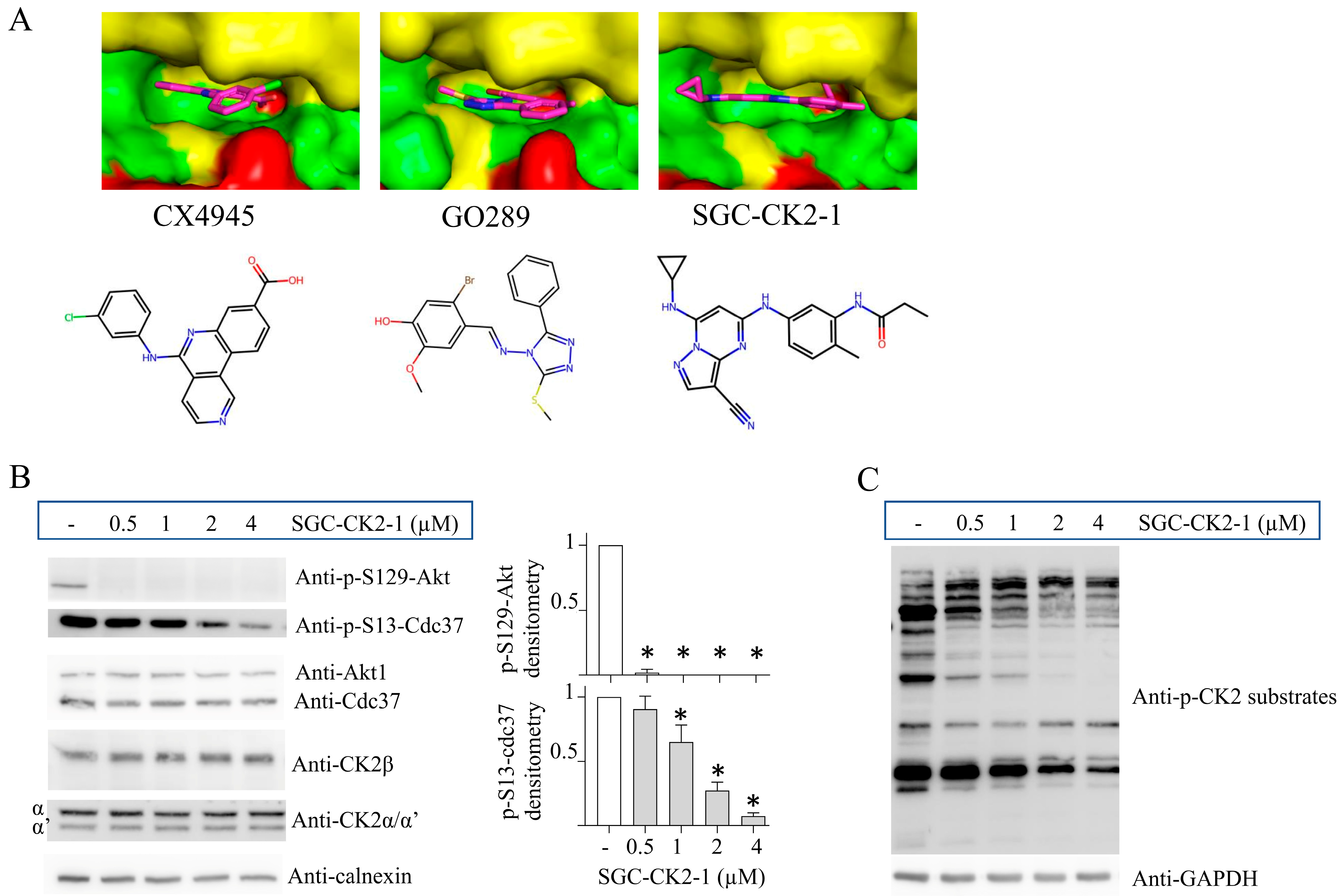
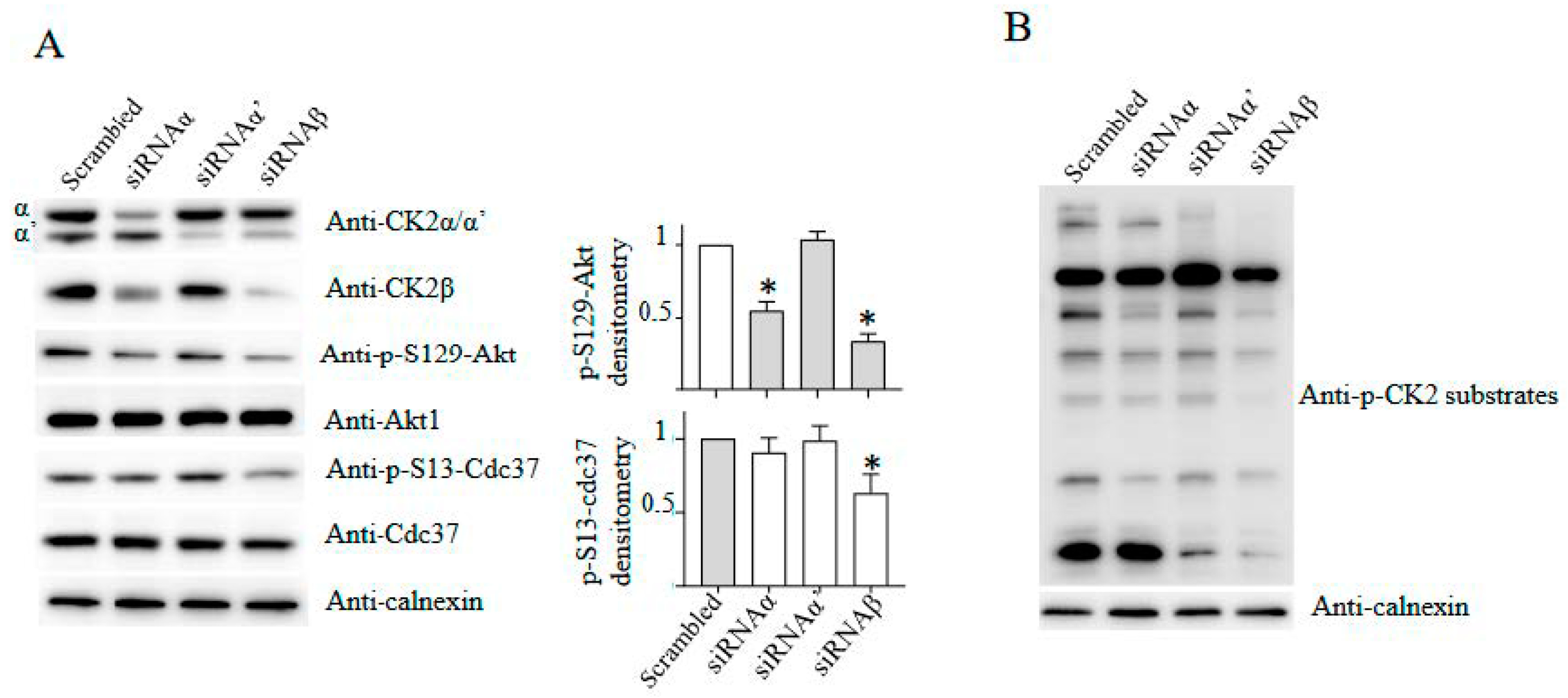
4. CK2 Knockdown, Knockout and Inhibition: Advantages and Disadvantages
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hornbeck, P.V.; Zhang, B.; Murray, B.; Kornhauser, J.M.; Latham, V.; Skrzypek, E. PhosphoSitePlus, 2014: Mutations, PTMs and Recalibrations. Nucleic Acids Res. 2015, 43, D512–D520. [Google Scholar] [CrossRef] [PubMed]
- D’Amore, C.; Salizzato, V.; Borgo, C.; Cesaro, L.; Pinna, L.A.; Salvi, M. A Journey through the Cytoskeleton with Protein Kinase CK2. Curr. Protein Pept. Sci. 2019, 20, 547–562. [Google Scholar] [CrossRef] [PubMed]
- Hashemolhosseini, S. The Role of Protein Kinase CK2 in Skeletal Muscle: Myogenesis, Neuromuscular Junctions, and Rhabdomyosarcoma. Neurosci. Lett. 2020, 729, 135001. [Google Scholar] [CrossRef]
- Montenarh, M. Protein Kinase CK2 and Angiogenesis. Adv. Clin. Exp. Med. 2014, 23, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Nuñez de Villavicencio-Diaz, T.; Rabalski, A.J.; Litchfield, D.W. Protein Kinase CK2: Intricate Relationships within Regulatory Cellular Networks. Pharmaceuticals 2017, 10, 27. [Google Scholar] [CrossRef]
- Montenarh, M.; Götz, C. Protein Kinase CK2 and Ion Channels (Review). Biomed. Rep. 2020, 13, 55. [Google Scholar] [CrossRef]
- St-Denis, N.A.; Litchfield, D.W. Protein Kinase CK2 in Health and Disease: From Birth to Death: The Role of Protein Kinase CK2 in the Regulation of Cell Proliferation and Survival. Cell. Mol. Life Sci. 2009, 66, 1817–1829. [Google Scholar] [CrossRef] [PubMed]
- Götz, C.; Montenarh, M. Protein Kinase CK2 in Development and Differentiation. Biomed. Rep. 2017, 6, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Kanki, T.; Kurihara, Y.; Jin, X.; Goda, T.; Ono, Y.; Aihara, M.; Hirota, Y.; Saigusa, T.; Aoki, Y.; Uchiumi, T.; et al. Casein Kinase 2 Is Essential for Mitophagy. EMBO Rep. 2013, 14, 788–794. [Google Scholar] [CrossRef]
- Borgo, C.; D’Amore, C.; Sarno, S.; Salvi, M.; Ruzzene, M. Protein Kinase CK2: A Potential Therapeutic Target for Diverse Human Diseases. Signal Transduct. Target. Ther. 2021, 6, 183. [Google Scholar] [CrossRef] [PubMed]
- Quezada Meza, C.P.; Ruzzene, M. Protein Kinase CK2 and SARS-CoV-2: An Expected Interplay Story. Kinases Phosphatases 2023, 1, 141–150. [Google Scholar] [CrossRef]
- Baier, A.; Szyszka, R. CK2 and Protein Kinases of the CK1 Superfamily as Targets for Neurodegenerative Disorders. Front. Mol. Biosci. 2022, 9, 916063. [Google Scholar] [CrossRef] [PubMed]
- White, A.; McGlone, A.; Gomez-Pastor, R. Protein Kinase CK2 and Its Potential Role as a Therapeutic Target in Huntington’s Disease. Biomedicines 2022, 10, 1979. [Google Scholar] [CrossRef] [PubMed]
- Halloran, D.; Pandit, V.; Nohe, A. The Role of Protein Kinase CK2 in Development and Disease Progression: A Critical Review. J. Dev. Biol. 2022, 10, 31. [Google Scholar] [CrossRef]
- Ampofo, E.; Nalbach, L.; Menger, M.D.; Montenarh, M.; Götz, C. Protein Kinase CK2-A Putative Target for the Therapy of Diabetes Mellitus? Int. J. Mol. Sci. 2019, 20, 4398. [Google Scholar] [CrossRef]
- Trembley, J.H.; Kren, B.T.; Afzal, M.; Scaria, G.A.; Klein, M.A.; Ahmed, K. Protein Kinase CK2—Diverse Roles in Cancer Cell Biology and Therapeutic Promise. Mol. Cell. Biochem. 2023, 478, 899–926. [Google Scholar] [CrossRef] [PubMed]
- Strum, S.W.; Gyenis, L.; Litchfield, D.W. CSNK2 in Cancer: Pathophysiology and Translational Applications. Br. J. Cancer 2022, 126, 994–1003. [Google Scholar] [CrossRef] [PubMed]
- Castello, J.; Ragnauth, A.; Friedman, E.; Rebholz, H. CK2-An Emerging Target for Neurological and Psychiatric Disorders. Pharmaceuticals 2017, 10, 7. [Google Scholar] [CrossRef]
- Unni, P.; Friend, J.; Weinberg, J.; Okur, V.; Hochscherf, J.; Dominguez, I. Predictive Functional, Statistical and Structural Analysis of CSNK2A1 and CSNK2B Variants Linked to Neurodevelopmental Diseases. Front. Mol. Biosci. 2022, 9, 851547. [Google Scholar] [CrossRef]
- Hong, H.; Benveniste, E.N. The Immune Regulatory Role of Protein Kinase CK2 and Its Implications for Treatment of Cancer. Biomedicines 2021, 9, 1932. [Google Scholar] [CrossRef] [PubMed]
- Salvi, M.; Sarno, S.; Cesaro, L.; Nakamura, H.; Pinna, L.A. Extraordinary Pleiotropy of Protein Kinase CK2 Revealed by Weblogo Phosphoproteome Analysis. Biochim. Biophys. Acta 2009, 1793, 847–859. [Google Scholar] [CrossRef] [PubMed]
- Cesaro, L.; Pinna, L.A. Prevalence and Significance of the Commonest Phosphorylated Motifs in the Human Proteome: A Global Analysis. Cell Mol. Life Sci. 2020, 77, 5281–5298. [Google Scholar] [CrossRef] [PubMed]
- Salvi, M.; Cesaro, L.; Tibaldi, E.; Pinna, L.A. Motif Analysis of Phosphosites Discloses a Potential Prominent Role of the Golgi Casein Kinase (GCK) in the Generation of Human Plasma Phospho-Proteome. J. Proteome Res. 2010, 9, 3335–3338. [Google Scholar] [CrossRef] [PubMed]
- Tagliabracci, V.S.; Wiley, S.E.; Guo, X.; Kinch, L.N.; Durrant, E.; Wen, J.; Xiao, J.; Cui, J.; Nguyen, K.B.; Engel, J.L.; et al. A Single Kinase Generates the Majority of the Secreted Phosphoproteome. Cell 2015, 161, 1619–1632. [Google Scholar] [CrossRef]
- Filhol, O.; Nueda, A.; Martel, V.; Gerber-Scokaert, D.; Benitez, M.J.; Souchier, C.; Saoudi, Y.; Cochet, C. Live-Cell Fluorescence Imaging Reveals the Dynamics of Protein Kinase CK2 Individual Subunits. Mol. Cell. Biol. 2003, 23, 975–987. [Google Scholar] [CrossRef]
- Niefind, K.; Guerra, B.; Ermakowa, I.; Issinger, O.G. Crystal Structure of Human Protein Kinase CK2: Insights into Basic Properties of the CK2 Holoenzyme. EMBO J. 2001, 20, 5320–5331. [Google Scholar] [CrossRef]
- Chua, M.M.J.; Lee, M.; Dominguez, I. Cancer-Type Dependent Expression of CK2 Transcripts. PLoS ONE 2017, 12, e0188854. [Google Scholar] [CrossRef]
- Salizzato, V.; Zanin, S.; Borgo, C.; Lidron, E.; Salvi, M.; Rizzuto, R.; Pallafacchina, G.; Donella-Deana, A. Protein Kinase CK2 Subunits Exert Specific and Coordinated Functions in Skeletal Muscle Differentiation and Fusogenic Activity. FASEB J. 2019, 33, 10648–10667. [Google Scholar] [CrossRef]
- Guerra, B.; Siemer, S.; Boldyreff, B.; Issinger, O.-G. Protein Kinase CK2: Evidence for a Protein Kinase CK2β Subunit Fraction, Devoid of the Catalytic CK2α Subunit, in Mouse Brain and Testicles. FEBS Lett. 1999, 462, 353–357. [Google Scholar] [CrossRef]
- Bibby, A.C.; Litchfield, D.W. The Multiple Personalities of the Regulatory Subunit of Protein Kinase CK2: CK2 Dependent and CK2 Independent Roles Reveal a Secret Identity for CK2β. Int. J. Biol. Sci. 2005, 1, 67–79. [Google Scholar] [CrossRef]
- Guerra, B.; Issinger, O.-G.; Wang, J.Y.J. Modulation of Human Checkpoint Kinase Chk1 by the Regulatory Beta-Subunit of Protein Kinase CK2. Oncogene 2003, 22, 4933–4942. [Google Scholar] [CrossRef]
- Ubersax, J.A.; Ferrell, J.E. Mechanisms of Specificity in Protein Phosphorylation. Nat. Rev. Mol. Cell. Biol. 2007, 8, 530–541. [Google Scholar] [CrossRef]
- Pinna, L.A.; Ruzzene, M. How Do Protein Kinases Recognize Their Substrates? Biochim. Biophys. Acta 1996, 1314, 191–225. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, P.S.L.; Ferraz, F.A.N.; Pena, D.A.; Pramio, D.T.; Morais, F.A.; Schechtman, D. Revisiting Protein Kinase–Substrate Interactions: Toward Therapeutic Development. Sci. Signal. 2016, 9, re3. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.L.; Yaron, T.M.; Huntsman, E.M.; Kerelsky, A.; Song, J.; Regev, A.; Lin, T.-Y.; Liberatore, K.; Cizin, D.M.; Cohen, B.M.; et al. An Atlas of Substrate Specificities for the Human Serine/Threonine Kinome. Nature 2023, 613, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Salvi, M.; Cesaro, L.; Pinna, L.A. Variable Contribution of Protein Kinases to the Generation of the Human Phosphoproteome: A Global Weblogo Analysis. Biomol. Concepts 2010, 1, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Borgo, C.; D’Amore, C.; Cesaro, L.; Sarno, S.; Pinna, L.A.; Ruzzene, M.; Salvi, M. How Can a Traffic Light Properly Work If It Is Always Green? The Paradox of CK2 Signaling. Crit. Rev. Biochem. Mol. Biol. 2021, 56, 321–359. [Google Scholar] [CrossRef] [PubMed]
- Pinna, L.A. Protein Kinase CK2: A Challenge to Canons. J. Cell Sci. 2002, 115, 3873–3878. [Google Scholar] [CrossRef]
- Borgo, C.; Franchin, C.; Cesaro, L.; Zaramella, S.; Arrigoni, G.; Salvi, M.; Pinna, L.A. A Proteomics Analysis of CK2β(-/-) C2C12 Cells Provides Novel Insights into the Biological Functions of the Non-Catalytic β Subunit. FEBS J. 2019, 286, 1561–1575. [Google Scholar] [CrossRef]
- Schneider, T.D.; Stephens, R.M. Sequence Logos: A New Way to Display Consensus Sequences. Nucleic Acids Res. 1990, 18, 6097–6100. [Google Scholar] [CrossRef]
- Vacic, V.; Iakoucheva, L.M.; Radivojac, P. Two Sample Logo: A Graphical Representation of the Differences between Two Sets of Sequence Alignments. Bioinformatics 2006, 22, 1536–1537. [Google Scholar] [CrossRef] [PubMed]
- Jurrus, E.; Engel, D.; Star, K.; Monson, K.; Brandi, J.; Felberg, L.E.; Brookes, D.H.; Wilson, L.; Chen, J.; Liles, K.; et al. Improvements to the APBS Biomolecular Solvation Software Suite. Protein Sci. 2018, 27, 112–128. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, E.; Kinoshita-Kikuta, E.; Takiyama, K.; Koike, T. Phosphate-Binding Tag, a New Tool to Visualize Phosphorylated Proteins. Mol. Cell. Proteom. 2006, 5, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.; Steinberg, T.H.; Cooley, L.A.; Gee, K.R.; Beechem, J.M.; Patton, W.F. Quantitative Analysis of Protein Phosphorylation Status and Protein Kinase Activity on Microarrays Using a Novel Fluorescent Phosphorylation Sensor Dye. Proteomics 2003, 3, 1244–1255. [Google Scholar] [CrossRef]
- Urban, J. A Review on Recent Trends in the Phosphoproteomics Workflow. From Sample Preparation to Data Analysis. Anal. Chim. Acta 2022, 1199, 338857. [Google Scholar] [CrossRef]
- Borgo, C.; Ruzzene, M. Protein Kinase CK2 Inhibition as a Pharmacological Strategy. Adv. Protein Chem. Struct. Biol. 2021, 124, 23–46. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, Y.; Wang, J.; Zhou, Z.; Cao, S.; Zhang, J. Strategies of Targeting CK2 in Drug Discovery: Challenges, Opportunities, and Emerging Prospects. J. Med. Chem. 2023, 66, 2257–2281. [Google Scholar] [CrossRef]
- Pierre, F.; Chua, P.C.; O’Brien, S.E.; Siddiqui-Jain, A.; Bourbon, P.; Haddach, M.; Michaux, J.; Nagasawa, J.; Schwaebe, M.K.; Stefan, E.; et al. Discovery and SAR of 5-(3-Chlorophenylamino)Benzo[c][2,6]Naphthyridine-8-Carboxylic Acid (CX-4945), the First Clinical Stage Inhibitor of Protein Kinase CK2 for the Treatment of Cancer. J. Med. Chem. 2011, 54, 635–654. [Google Scholar] [CrossRef]
- Bogusz, J.; Zrubek, K.; Rembacz, K.P.; Grudnik, P.; Golik, P.; Romanowska, M.; Wladyka, B.; Dubin, G. Structural Analysis of PIM1 Kinase Complexes with ATP-Competitive Inhibitors. Sci. Rep. 2017, 7, 13399. [Google Scholar] [CrossRef]
- Grygier, P.; Pustelny, K.; Nowak, J.; Golik, P.; Popowicz, G.M.; Plettenburg, O.; Dubin, G.; Menezes, F.; Czarna, A. Silmitasertib (CX-4945), a Clinically Used CK2-Kinase Inhibitor with Additional Effects on GSK3β and DYRK1A Kinases: A Structural Perspective. J. Med. Chem. 2023, 66, 4009–4024. [Google Scholar] [CrossRef]
- Kim, H.; Lee, K.-S.; Kim, A.-K.; Choi, M.; Choi, K.; Kang, M.; Chi, S.-W.; Lee, M.-S.; Lee, J.-S.; Lee, S.-Y.; et al. A Chemical with Proven Clinical Safety Rescues Down-Syndrome-Related Phenotypes in through DYRK1A Inhibition. Dis. Model Mech. 2016, 9, 839–848. [Google Scholar] [CrossRef]
- Kim, H.; Choi, K.; Kang, H.; Lee, S.-Y.; Chi, S.-W.; Lee, M.-S.; Song, J.; Im, D.; Choi, Y.; Cho, S. Identification of a Novel Function of CX-4945 as a Splicing Regulator. PLoS ONE 2014, 9, e94978. [Google Scholar] [CrossRef]
- Lertsuwan, J.; Lertsuwan, K.; Sawasdichai, A.; Tasnawijitwong, N.; Lee, K.Y.; Kitchen, P.; Afford, S.; Gaston, K.; Jayaraman, P.-S.; Satayavivad, J. CX-4945 Induces Methuosis in Cholangiocarcinoma Cell Lines by a CK2-Independent Mechanism. Cancers 2018, 10, 283. [Google Scholar] [CrossRef]
- D’Amore, C.; Moro, E.; Borgo, C.; Itami, K.; Hirota, T.; Pinna, L.A.; Salvi, M. “Janus” Efficacy of CX-5011: CK2 Inhibition and Methuosis Induction by Independent Mechanisms. Biochim. Biophys. Acta BBA Mol. Cell Res. 2020, 1867, 118807. [Google Scholar] [CrossRef]
- Zandomeni, R.O. Kinetics of Inhibition by 5,6-Dichloro-1-β-d-Ribofuranosylbenzimidazole on Calf Thymus Casein Kinase II. Biochem. J. 1989, 262, 469–473. [Google Scholar] [CrossRef]
- Andrzejewska, M.; Pagano, M.A.; Meggio, F.; Brunati, A.M.; Kazimierczuk, Z. Polyhalogenobenzimidazoles: Synthesis and Their Inhibitory Activity against Casein Kinases. Bioorganic Med. Chem. 2003, 11, 3997–4002. [Google Scholar] [CrossRef]
- Cozza, G.; Girardi, C.; Ranchio, A.; Lolli, G.; Sarno, S.; Orzeszko, A.; Kazimierczuk, Z.; Battistutta, R.; Ruzzene, M.; Pinna, L.A. Cell-Permeable Dual Inhibitors of Protein Kinases CK2 and PIM-1: Structural Features and Pharmacological Potential. Cell. Mol. Life Sci. 2014, 71, 3173–3185. [Google Scholar] [CrossRef]
- Pagano, M.A.; Andrzejewska, M.; Ruzzene, M.; Sarno, S.; Cesaro, L.; Bain, J.; Elliott, M.; Meggio, F.; Kazimierczuk, Z.; Pinna, L.A. Optimization of Protein Kinase CK2 Inhibitors Derived from 4,5,6,7-Tetrabromobenzimidazole. J. Med. Chem. 2004, 47, 6239–6247. [Google Scholar] [CrossRef]
- McCarty, M.F.; Assanga, S.I.; Lujan, L.L. Flavones and Flavonols May Have Clinical Potential as CK2 Inhibitors in Cancer Therapy. Med. Hypotheses 2020, 141, 109723. [Google Scholar] [CrossRef]
- Urolithin as a Converging Scaffold Linking Ellagic Acid and Coumarin Analogues: Design of Potent Protein Kinase CK2 Inhibitors—Cozza—2011—ChemMedChem—Wiley Online Library. Available online: https://chemistry-europe.onlinelibrary.wiley.com/doi/10.1002/cmdc.201100338 (accessed on 20 September 2023).
- Yim, H.; Lee, Y.H.; Lee, C.H.; Lee, S.K. Emodin, an Anthraquinone Derivative Isolated from the Rhizomes of Rheum Palmatum, Selectively Inhibits the Activity of Casein Kinase II as a Competitive Inhibitor. Planta Medica 1999, 65, 9–13. [Google Scholar] [CrossRef]
- Meggio, F.; Pagano, M.A.; Moro, S.; Zagotto, G.; Ruzzene, M.; Sarno, S.; Cozza, G.; Bain, J.; Elliott, M.; Deana, A.D.; et al. Inhibition of Protein Kinase CK2 by Condensed Polyphenolic Derivatives. An in Vitro and in Vivo Study. Biochemistry 2004, 43, 12931–12936. [Google Scholar] [CrossRef]
- Cozza, G.; Mazzorana, M.; Papinutto, E.; Bain, J.; Elliott, M.; di Maira, G.; Gianoncelli, A.; Pagano, M.A.; Sarno, S.; Ruzzene, M.; et al. Quinalizarin as a Potent, Selective and Cell-Permeable Inhibitor of Protein Kinase CK2. Biochem. J. 2009, 421, 387–395. [Google Scholar] [CrossRef]
- Wells, C.I.; Drewry, D.H.; Pickett, J.E.; Tjaden, A.; Krämer, A.; Müller, S.; Gyenis, L.; Menyhart, D.; Litchfield, D.W.; Knapp, S.; et al. Development of a Potent and Selective Chemical Probe for the Pleiotropic Kinase CK2. Cell Chem. Biol. 2021, 28, 546–558. [Google Scholar] [CrossRef]
- Vangrevelinghe, E.; Zimmermann, K.; Schoepfer, J.; Portmann, R.; Fabbro, D.; Furet, P. Discovery of a Potent and Selective Protein Kinase CK2 Inhibitor by High-Throughput Docking. J. Med. Chem. 2003, 46, 2656–2662. [Google Scholar] [CrossRef]
- Pagano, M.A.; Poletto, G.; Di Maira, G.; Cozza, G.; Ruzzene, M.; Sarno, S.; Bain, J.; Elliott, M.; Moro, S.; Zagotto, G.; et al. Tetrabromocinnamic Acid (TBCA) and Related Compounds Represent a New Class of Specific Protein Kinase CK2 Inhibitors. Chembiochem 2007, 8, 129–139. [Google Scholar] [CrossRef]
- Battistutta, R.; Cozza, G.; Pierre, F.; Papinutto, E.; Lolli, G.; Sarno, S.; O’Brien, S.E.; Siddiqui-Jain, A.; Haddach, M.; Anderes, K.; et al. Unprecedented Selectivity and Structural Determinants of a New Class of Protein Kinase CK2 Inhibitors in Clinical Trials for the Treatment of Cancer. Biochemistry 2011, 50, 8478–8488. [Google Scholar] [CrossRef]
- Oshima, T.; Niwa, Y.; Kuwata, K.; Srivastava, A.; Hyoda, T.; Tsuchiya, Y.; Kumagai, M.; Tsuyuguchi, M.; Tamaru, T.; Sugiyama, A.; et al. Cell-Based Screen Identifies a New Potent and Highly Selective CK2 Inhibitor for Modulation of Circadian Rhythms and Cancer Cell Growth. Sci. Adv. 2019, 5, eaau9060. [Google Scholar] [CrossRef]
- Menyhart, D.; Gyenis, L.; Jurcic, K.; Roffey, S.E.; Puri, A.; Jovanovic, P.; Szkop, K.J.; Pittock, P.; Lajoie, G.; Axtman, A.D.; et al. Comparison of CX-4945 and SGC-CK2-1 as Inhibitors of CSNK2 Using Quantitative Phosphoproteomics: Triple SILAC in Combination with Inhibitor-Resistant CSNK2. Curr. Res. Chem. Biol. 2023, 3, 100041. [Google Scholar] [CrossRef]
- Cohen, P. Guidelines for the Effective Use of Chemical Inhibitors of Protein Function to Understand Their Roles in Cell Regulation. Biochem. J. 2009, 425, 53–54. [Google Scholar] [CrossRef]
- Birus, R.; El-Awaad, E.; Ballentin, L.; Alchab, F.; Aichele, D.; Ettouati, L.; Götz, C.; Le Borgne, M.; Jose, J. 4,5,7-Trisubstituted Indeno[1,2-b]Indole Inhibits CK2 Activity in Tumor Cells Equivalent to CX-4945 and Shows Strong Anti-Migratory Effects. FEBS Open Bio 2022, 12, 394–411. [Google Scholar] [CrossRef]
- Marin, O.; Meggio, F.; Pinna, L.A. Design and Synthesis of Two New Peptide Substrates for the Specific and Sensitive Monitoring of Casein Kinases-1 and -2. Biochem. Biophys. Res. Commun. 1994, 198, 898–905. [Google Scholar] [CrossRef] [PubMed]
- Ruzzene, M.; Penzo, D.; Pinna, L.A. Protein Kinase CK2 Inhibitor 4,5,6,7-Tetrabromobenzotriazole (TBB) Induces Apoptosis and Caspase-Dependent Degradation of Haematopoietic Lineage Cell-Specific Protein 1 (HS1) in Jurkat Cells. Biochem. J. 2002, 364, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Zanin, S.; Borgo, C.; Girardi, C.; O’Brien, S.E.; Miyata, Y.; Pinna, L.A.; Donella-Deana, A.; Ruzzene, M. Effects of the CK2 Inhibitors CX-4945 and CX-5011 on Drug-Resistant Cells. PLoS ONE 2012, 7, e49193. [Google Scholar] [CrossRef] [PubMed]
- Borgo, C.; Cesaro, L.; Hirota, T.; Kuwata, K.; D’Amore, C.; Ruppert, T.; Blatnik, R.; Salvi, M.; Pinna, L.A. Comparing the Efficacy and Selectivity of Ck2 Inhibitors. A Phosphoproteomics Approach. Eur. J. Med. Chem. 2021, 214, 113217. [Google Scholar] [CrossRef]
- Borgo, C.; Franchin, C.; Scalco, S.; Bosello-Travain, V.; Donella-Deana, A.; Arrigoni, G.; Salvi, M.; Pinna, L.A. Generation and Quantitative Proteomics Analysis of CK2α/α’(-/-) Cells. Sci. Rep. 2017, 7, 42409. [Google Scholar] [CrossRef] [PubMed]
- Borgo, C.; D’Amore, C.; Cesaro, L.; Itami, K.; Hirota, T.; Salvi, M.; Pinna, L.A. A N-Terminally Deleted Form of the CK2α’ Catalytic Subunit Is Sufficient to Support Cell Viability. Biochem. Biophys. Res. Commun. 2020, 531, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Franchin, C.; Borgo, C.; Cesaro, L.; Zaramella, S.; Vilardell, J.; Salvi, M.; Arrigoni, G.; Pinna, L.A. Re-Evaluation of Protein Kinase CK2 Pleiotropy: New Insights Provided by a Phosphoproteomics Analysis of CK2 Knockout Cells. Cell. Mol. Life Sci. 2018, 75, 2011–2026. [Google Scholar] [CrossRef]
- Miyata, Y.; Nishida, E. CK2 Binds, Phosphorylates, and Regulates Its Pivotal Substrate Cdc37, an Hsp90-Cochaperone. Mol. Cell. Biochem. 2005, 274, 171–179. [Google Scholar] [CrossRef]
- Feliciano, A.; Castellvi, J.; Artero-Castro, A.; Leal, J.A.; Romagosa, C.; Hernández-Losa, J.; Peg, V.; Fabra, A.; Vidal, F.; Kondoh, H.; et al. MiR-125b Acts as a Tumor Suppressor in Breast Tumorigenesis via Its Novel Direct Targets ENPEP, CK2-α, CCNJ, and MEGF9. PLoS ONE 2013, 8, e76247. [Google Scholar] [CrossRef]
- Liang, Y.; Xu, J.; Wang, Y.; Tang, J.-Y.; Yang, S.-L.; Xiang, H.-G.; Wu, S.-X.; Li, X.-J. Inhibition of MiRNA-125b Decreases Cerebral Ischemia/Reperfusion Injury by Targeting CK2α/NADPH Oxidase Signaling. Cell. Physiol. Biochem. 2018, 45, 1818–1826. [Google Scholar] [CrossRef]
- Lee, Y.-H.; Kim, S.Y.; Bae, Y.-S. Upregulation of MiR-760 and MiR-186 Is Associated with Replicative Senescence in Human Lung Fibroblast Cells. Mol. Cells 2014, 37, 620–627. [Google Scholar] [CrossRef]
- Song, J.; Bae, Y.-S. CK2 Down-Regulation Increases the Expression of Senescence-Associated Secretory Phenotype Factors through NF-ΚB Activation. Int. J. Mol. Sci. 2021, 22, 406. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Bae, Y.-S. Long Non-Coding RNA KCNQ1OT1 Regulates Protein Kinase CK2 Via MiR-760 in Senescence and Calorie Restriction. Int. J. Mol. Sci. 2022, 23, 1888. [Google Scholar] [CrossRef] [PubMed]
- Naeli, P.; Winter, T.; Hackett, A.P.; Alboushi, L.; Jafarnejad, S.M. The Intricate Balance between MicroRNA-Induced MRNA Decay and Translational Repression. FEBS J. 2023, 290, 2508–2524. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Lee, Y.-H.; Bae, Y.-S. MiR-186, MiR-216b, MiR-337-3p, and MiR-760 Cooperatively Induce Cellular Senescence by Targeting α Subunit of Protein Kinase CKII in Human Colorectal Cancer Cells. Biochem. Biophys. Res. Commun. 2012, 429, 173–179. [Google Scholar] [CrossRef]
- Borgo, C.; Cesaro, L.; Hirota, T.; Kuwata, K.; D’Amore, C.; Ruppert, T.; Blatnik, R.; Salvi, M.; Pinna, L.A. Analysis of the Phosphoproteome of CK2α(-/-)/Δα’ C2C12 Myoblasts Compared to the Wild-Type Cells. Open Biol. 2023, 13, 220220. [Google Scholar] [CrossRef]
- Lettieri, A.; Borgo, C.; Zanieri, L.; D’Amore, C.; Oleari, R.; Paganoni, A.; Pinna, L.A.; Cariboni, A.; Salvi, M. Protein Kinase CK2 Subunits Differentially Perturb the Adhesion and Migration of GN11 Cells: A Model of Immature Migrating Neurons. Int. J. Mol. Sci. 2019, 20, 5951. [Google Scholar] [CrossRef]
- Di Maira, G.; Gentilini, A.; Pastore, M.; Caligiuri, A.; Piombanti, B.; Raggi, C.; Rovida, E.; Lewinska, M.; Andersen, J.B.; Borgo, C.; et al. The Protein Kinase CK2 Contributes to the Malignant Phenotype of Cholangiocarcinoma Cells. Oncogenesis 2019, 8, 61. [Google Scholar] [CrossRef]
- D’Amore, C.; Borgo, C.; Bosello-Travain, V.; Vilardell, J.; Salizzato, V.; Pinna, L.A.; Venerando, A.; Salvi, M. Deciphering the Role of Protein Kinase CK2 in the Maturation/Stability of F508del-CFTR. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2020, 1866, 165611. [Google Scholar] [CrossRef]
- Schmitt, B.M.; Boewe, A.S.; Götz, C.; Philipp, S.E.; Urbschat, S.; Oertel, J.; Menger, M.D.; Laschke, M.W.; Ampofo, E. CK2 Activity Mediates the Aggressive Molecular Signature of Glioblastoma Multiforme by Inducing Nerve/Glial Antigen (NG)2 Expression. Cancers 2021, 13, 1678. [Google Scholar] [CrossRef]
- Vilk, G.; Saulnier, R.B.; St Pierre, R.; Litchfield, D.W. Inducible Expression of Protein Kinase CK2 in Mammalian Cells. Evidence for Functional Specialization of CK2 Isoforms. J. Biol. Chem. 1999, 274, 14406–14414. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Toselli, P.A.; Russell, L.D.; Seldin, D.C. Globozoospermia in Mice Lacking the Casein Kinase II Alpha’ Catalytic Subunit. Nat. Genet. 1999, 23, 118–121. [Google Scholar] [CrossRef]
- Lou, D.Y.; Dominguez, I.; Toselli, P.; Landesman-Bollag, E.; O’Brien, C.; Seldin, D.C. The Alpha Catalytic Subunit of Protein Kinase CK2 Is Required for Mouse Embryonic Development. Mol. Cell. Biol. 2008, 28, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Prudent, R.; Cochet, C. New Protein Kinase CK2 Inhibitors: Jumping out of the Catalytic Box. Chem. Biol. 2009, 16, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Iegre, J.; Atkinson, E.L.; Brear, P.D.; Cooper, B.M.; Hyvönen, M.; Spring, D.R. Chemical Probes Targeting the Kinase CK2: A Journey Outside the Catalytic Box. Org. Biomol. Chem. 2021, 19, 4380–4396. [Google Scholar] [CrossRef]
- Ballardin, D.; Cruz-Gamero, J.M.; Bienvenu, T.; Rebholz, H. Comparing Two Neurodevelopmental Disorders Linked to CK2: Okur-Chung Neurodevelopmental Syndrome and Poirier-Bienvenu Neurodevelopmental Syndrome-Two Sides of the Same Coin? Front. Mol. Biosci. 2022, 9, 850559. [Google Scholar] [CrossRef]
- Dominguez, I.; Cruz-Gamero, J.M.; Corasolla, V.; Dacher, N.; Rangasamy, S.; Urbani, A.; Narayanan, V.; Rebholz, H. Okur-Chung Neurodevelopmental Syndrome-Linked CK2α Variants Have Reduced Kinase Activity. Hum. Genet. 2021, 140, 1077–1096. [Google Scholar] [CrossRef]
- Caefer, D.M.; Phan, N.Q.; Liddle, J.C.; Balsbaugh, J.L.; O’Shea, J.P.; Tzingounis, A.V.; Schwartz, D. The Okur-Chung Neurodevelopmental Syndrome Mutation CK2K198R Leads to a Rewiring of Kinase Specificity. Front. Mol. Biosci. 2022, 9, 850661. [Google Scholar] [CrossRef]
- Werner, C.; Gast, A.; Lindenblatt, D.; Nickelsen, A.; Niefind, K.; Jose, J.; Hochscherf, J. Structural and Enzymological Evidence for an Altered Substrate Specificity in Okur-Chung Neurodevelopmental Syndrome Mutant CK2αLys198Arg. Front. Mol. Biosci. 2022, 9, 831693. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| ATP-Competitive Inhibitors | Ki or IC50 | References | |
|---|---|---|---|
| Polyhalogenated benzimidazole and benzotriazole derivates | DRB | Ki = 23 μM | [55] |
| TBB | IC50 = 1.6 μM | [56] | |
| TDB | IC50 = 0.032 μM | [57] | |
| TBI | IC50 = 0.5 μM | [58] | |
| DMAT | IC50 = 0.14 μM | [58] | |
| Flavonoids | Quercetin | IC50 = 0.55 μM | [59] |
| Apigenin | IC50 = 0.8 μM | [59] | |
| Fisetin | IC50 = 0.35 μM | [59] | |
| Coumarins | Ellagic acid | IC50 = 40 nM | [60] |
| Antraquinones | Emodin | Ki = 1.85 μM | [61] |
| MNX | Ki = 0.8 μM | [62] | |
| MNA | Ki = 0.78 μM | [62] | |
| Quinalizarin | Ki = 0.06 μM | [63] | |
| Pyrazolo[1,5-a]pyrimidine derivatives | SGC-CK2-1 | IC50 = 36 nM (CK2α) IC50 = 16 nM (CK2α′) | [64] |
| Carboxyl acid derivatives | IQA | IC50 = 0.39 μM | [65] |
| TBCA | IC50 = 110 nM | [66] | |
| CX-5011 | IC50 = 2.3 nM | [67] | |
| CX-4945 | IC50 = 1 nM | [48] | |
| Other compounds | GO289 | IC50 = 7 nM | [68] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cesaro, L.; Zuliani, A.M.; Bosello Travain, V.; Salvi, M. Exploring Protein Kinase CK2 Substrate Recognition and the Dynamic Response of Substrate Phosphorylation to Kinase Modulation. Kinases Phosphatases 2023, 1, 251-264. https://doi.org/10.3390/kinasesphosphatases1040015
Cesaro L, Zuliani AM, Bosello Travain V, Salvi M. Exploring Protein Kinase CK2 Substrate Recognition and the Dynamic Response of Substrate Phosphorylation to Kinase Modulation. Kinases and Phosphatases. 2023; 1(4):251-264. https://doi.org/10.3390/kinasesphosphatases1040015
Chicago/Turabian StyleCesaro, Luca, Angelica Maria Zuliani, Valentina Bosello Travain, and Mauro Salvi. 2023. "Exploring Protein Kinase CK2 Substrate Recognition and the Dynamic Response of Substrate Phosphorylation to Kinase Modulation" Kinases and Phosphatases 1, no. 4: 251-264. https://doi.org/10.3390/kinasesphosphatases1040015
APA StyleCesaro, L., Zuliani, A. M., Bosello Travain, V., & Salvi, M. (2023). Exploring Protein Kinase CK2 Substrate Recognition and the Dynamic Response of Substrate Phosphorylation to Kinase Modulation. Kinases and Phosphatases, 1(4), 251-264. https://doi.org/10.3390/kinasesphosphatases1040015