
Protein Kinase CK2 and SARS-CoV-2: An Expected Interplay Story



{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. CK2 General Features, Structure, and Regulation
1.2. CK2 in Diseases
1.3. CK2 in Viral Infections
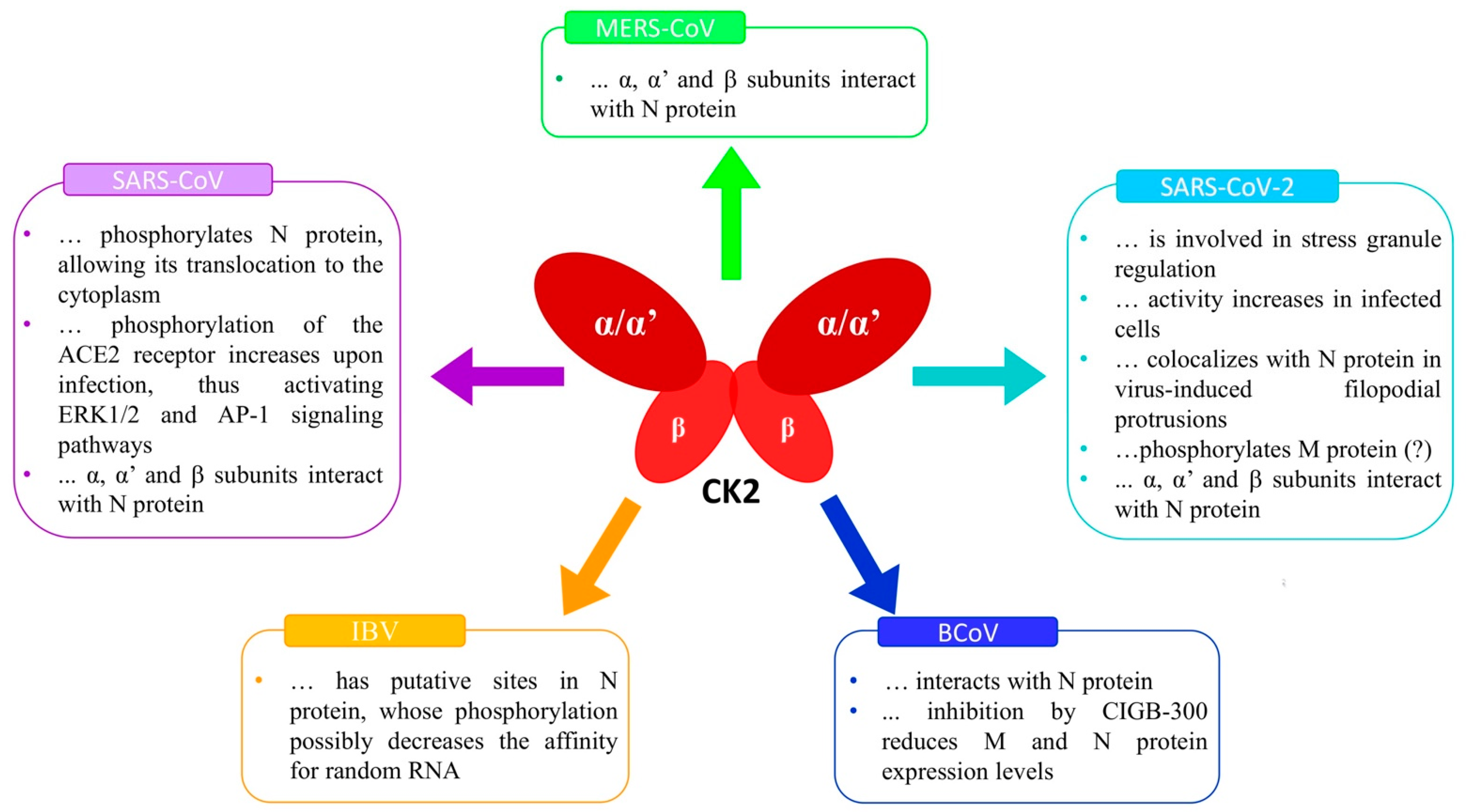
2. CK2 and Coronaviruses
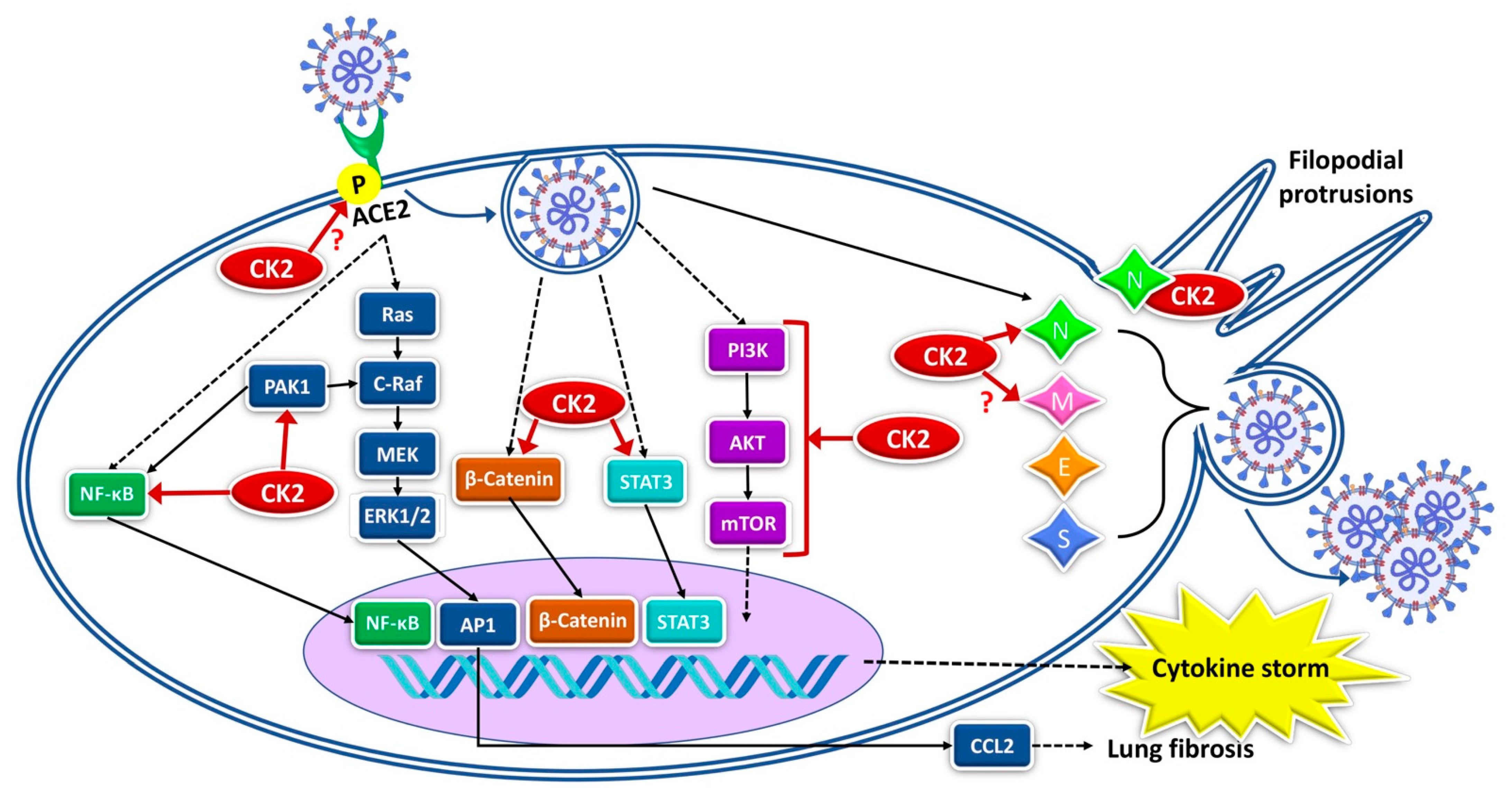
3. CK2 and SARS-CoV-2
4. CK2 Targeting as Anti-COVID-19 Strategy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Filhol, O.; Cochet, C. Protein Kinase CK2 in Health and Disease: Cellular Functions of Protein Ki-nase CK2: A Dynamic Affair. Cell. Mol. Life Sci. CMLS 2009, 66, 1830–1839. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, F.; Allende, C.C.; Allende, J.E. Protein Kinase Casein Kinase 2 Holoenzyme Produced Ectopi-cally in Human Cells Can Be Exported to the External Side of the Cellular Membrane. Proc. Natl. Acad. Sci. USA 2005, 102, 4718–4723. [Google Scholar] [CrossRef] [PubMed]
- Venerando, A.; Ruzzene, M.; Pinna, L.A. Casein Kinase: The Triple Meaning of a Misnomer. Biochem. J. 2014, 460, 141–156. [Google Scholar] [CrossRef]
- Meggio, F.; Pinna, L.A. One-Thousand-and-One Substrates of Protein Kinase CK2? FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2003, 17, 349–368. [Google Scholar] [CrossRef]
- Dominguez, I.; Degano, I.R.; Chea, K.; Cha, J.; Toselli, P.; Seldin, D.C. CK2α Is Essential for Embryonic Morphogenesis. Mol. Cell. Biochem. 2011, 356, 209–216. [Google Scholar] [CrossRef]
- Guerra, B.; Issinger, O.G. Protein Kinase CK2 and Its Role in Cellular Proliferation, Development and Pathology. Electrophoresis 1999, 20, 391–408. [Google Scholar] [CrossRef]
- Issinger, O.G. Casein Kinases: Pleiotropic Mediators of Cellular Regulation. Pharmacol. Ther. 1993, 59, 1–30. [Google Scholar] [CrossRef]
- St-Denis, N.A.; Litchfield, D.W. Protein Kinase CK2 in Health and Disease: From Birth to Death: The Role of Protein Kinase CK2 in the Regulation of Cell Proliferation and Survival. Cell. Mol. Life Sci. CMLS 2009, 66, 1817–1829. [Google Scholar] [CrossRef]
- Ruzzene, M.; Pinna, L.A. Addiction to Protein Kinase CK2: A Common Denominator of Diverse Cancer Cells? Biochim. Biophys. Acta 2010, 1804, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Firnau, M.-B.; Brieger, A. CK2 and the Hallmarks of Cancer. Biomedicines 2022, 10, 1987. [Google Scholar] [CrossRef] [PubMed]
- Strum, S.W.; Gyenis, L.; Litchfield, D.W. CSNK2 in Cancer: Pathophysiology and Translational Applications. Br. J. Cancer 2022, 126, 994–1003. [Google Scholar] [CrossRef] [PubMed]
- Trembley, J.H.; Kren, B.T.; Afzal, M.; Scaria, G.A.; Klein, M.A.; Ahmed, K. Protein Kinase CK2—Diverse Roles in Cancer Cell Biology and Therapeutic Promise. Mol. Cell. Biochem. 2022, 478, 899–926. [Google Scholar] [CrossRef]
- Borgo, C.; Ruzzene, M. Protein Kinase CK2 Inhibition as a Pharmacological Strategy. In Advances in Protein Chemistry and Structural Biology; Academic Press: Cambridge, MA, USA, 2021. [Google Scholar]
- Borgo, C.; Ruzzene, M. Role of Protein Kinase CK2 in Antitumor Drug Resistance. J. Exp. Clin. Cancer Res. CR 2019, 38, 287. [Google Scholar] [CrossRef] [PubMed]
- Buontempo, F.; McCubrey, J.A.; Orsini, E.; Ruzzene, M.; Cappellini, A.; Lonetti, A.; Evangelisti, C.; Chiarini, F.; Evangelisti, C.; Barata, J.T.; et al. Therapeutic Targeting of CK2 in Acute and Chronic Leukemias. Leukemia 2018, 32, 1–10. [Google Scholar] [CrossRef]
- Gowda, C.; Sachdev, M.; Muthusami, S.; Kapadia, M.; Petrovic-Dovat, L.; Hartman, M.; Ding, Y.; Song, C.; Payne, J.L.; Tan, B.-H.; et al. Casein Kinase II (CK2) as a Therapeutic Target for Hematological Malignancies. Curr. Pharm. Des. 2017, 23, 95–107. [Google Scholar] [CrossRef]
- Trembley, J.H.; Chen, Z.; Unger, G.; Slaton, J.; Kren, B.T.; Van Waes, C.; Ahmed, K. Emergence of Protein Kinase CK2 as a Key Target in Cancer Therapy. BioFactors Oxf. Engl. 2010, 36, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Borgo, C.; D’Amore, C.; Cesaro, L.; Sarno, S.; Pinna, L.A.; Ruzzene, M.; Salvi, M. How Can a Traffic Light Properly Work If It Is Always Green? The Paradox of CK2 Signaling. Crit. Rev. Biochem. Mol. Biol. 2021, 56, 321–359. [Google Scholar] [CrossRef]
- Roffey, S.E.; Litchfield, D.W. CK2 Regulation: Perspectives in 2021. Biomedicines 2021, 9, 1361. [Google Scholar] [CrossRef]
- Pinna, L.A. Protein Kinase CK2: A Challenge to Canons. J. Cell Sci. 2002, 115, 3873–3878. [Google Scholar] [CrossRef]
- Litchfield, D.W.; Bosc, D.G.; Canton, D.A.; Saulnier, R.B.; Vilk, G.; Zhang, C. Functional Specialization of CK2 Isoforms and Characterization of Isoform-Specific Binding Partners. Mol. Cell. Biochem. 2001, 227, 21–29. [Google Scholar] [CrossRef]
- Borgo, C.; D’Amore, C.; Sarno, S.; Salvi, M.; Ruzzene, M. Protein Kinase CK2: A Potential Therapeutic Target for Diverse Human Diseases. Signal Transduct. Target. Ther. 2021, 6, 183. [Google Scholar] [CrossRef]
- ole-MoiYoi, O.K. Casein Kinase II in Theileriosis. Science 1995, 267, 834–836. [Google Scholar] [CrossRef] [PubMed]
- Ortega, C.E.; Seidner, Y.; Dominguez, I. Mining CK2 in Cancer. PLoS ONE 2014, 9, e115609. [Google Scholar] [CrossRef]
- Chua, M.M.J.; Lee, M.; Dominguez, I. Cancer-Type Dependent Expression of CK2 Transcripts. PLoS ONE 2017, 12, e0188854. [Google Scholar] [CrossRef]
- Hong, H.; Benveniste, E.N. The Immune Regulatory Role of Protein Kinase CK2 and Its Implications for Treatment of Cancer. Biomedicines 2021, 9, 1932. [Google Scholar] [CrossRef] [PubMed]
- Ampofo, E.; Nalbach, L.; Menger, M.D.; Montenarh, M.; Götz, C. Protein Kinase CK2-A Putative Target for the Therapy of Diabetes Mellitus? Int. J. Mol. Sci. 2019, 20, 4398. [Google Scholar] [CrossRef] [PubMed]
- Ljubimov, A.V.; Caballero, S.; Aoki, A.M.; Pinna, L.A.; Grant, M.B.; Castellon, R. Involvement of Protein Kinase CK2 in Angiogenesis and Retinal Neovascularization. Investig. Ophthalmol. Vis. Sci. 2004, 45, 4583–4591. [Google Scholar] [CrossRef]
- Morooka, S.; Hoshina, M.; Kii, I.; Okabe, T.; Kojima, H.; Inoue, N.; Okuno, Y.; Denawa, M.; Yoshida, S.; Fukuhara, J.; et al. Identification of a Dual Inhibitor of SRPK1 and CK2 That Attenuates Pathological Angiogenesis of Macular Degeneration in Mice. Mol. Pharmacol. 2015, 88, 316–325. [Google Scholar] [CrossRef]
- Hauck, L.; Harms, C.; An, J.; Rohne, J.; Gertz, K.; Dietz, R.; Endres, M.; von Harsdorf, R. Protein Kinase CK2 Links Extracellular Growth Factor Signaling with the Control of P27 Kip1 Stability in the Heart. Nat. Med. 2008, 14, 315–324. [Google Scholar] [CrossRef]
- Wadey, K.S.; Brown, B.A.; Sala-Newby, G.B.; Jayaraman, P.-S.; Gaston, K.; George, S.J. Protein Kinase CK2 Inhibition Suppresses Neointima Formation via a Proline-Rich Homeodomain-Dependent Mechanism. Vascul. Pharmacol. 2017, 99, 34–44. [Google Scholar] [CrossRef]
- Zhou, H.; Zhu, P.; Wang, J.; Zhu, H.; Ren, J.; Chen, Y. Pathogenesis of Cardiac Ischemia-Reperfusion Injury Is Associated with CK2α-Disturbed Mitochondrial Homeostasis via Suppression of FUNDC1-Related Mitophagy. Cell Death Differ. 2018, 25, 1080–1093. [Google Scholar] [CrossRef] [PubMed]
- Castello, J.; Ragnauth, A.; Friedman, E.; Rebholz, H. CK2-An Emerging Target for Neurological and Psychiatric Disorders. Pharmaceuticals 2017, 10, 7. [Google Scholar] [CrossRef]
- Mehta, A. Cystic Fibrosis as a Bowel Cancer Syndrome, and the Potential Role of CK2. Mol. Cell. Biochem. 2008, 316, 169–175. [Google Scholar] [CrossRef]
- Ballardin, D.; Cruz-Gamero, J.M.; Bienvenu, T.; Rebholz, H. Comparing Two Neurodevelopmental Disorders Linked to CK2: Okur-Chung Neurodevelopmental Syndrome and Poirier-Bienvenu Neurodevelopmental Syndrome-Two Sides of the Same Coin? Front. Mol. Biosci. 2022, 9, 850559. [Google Scholar] [CrossRef]
- Chatterjee, B.; Thakur, S.S. SARS-CoV-2 Infection Triggers Phosphorylation: Potential Target for Anti-COVID-19 Therapeutics. Front. Immunol. 2022, 13, 829474. [Google Scholar] [CrossRef]
- Gottardo, M.F.; Capobianco, C.S.; Sidabra, J.E.; Garona, J.; Perera, Y.; Perea, S.E.; Alonso, D.F.; Farina, H.G. Preclinical Efficacy of CIGB-300, an Anti-CK2 Peptide, on Breast Cancer Metastasic Colonization. Sci. Rep. 2020, 10, 14689. [Google Scholar] [CrossRef] [PubMed]
- Perea, S.E.; Reyes, O.; Baladron, I.; Perera, Y.; Farina, H.; Gil, J.; Rodriguez, A.; Bacardi, D.; Marcelo, J.L.; Cosme, K.; et al. CIGB-300, a Novel Proapoptotic Peptide That Impairs the CK2 Phosphorylation and Exhibits Anticancer Properties Both in Vitro and Vivo. Mol. Cell. Biochem. 2008, 316, 163–167. [Google Scholar] [CrossRef]
- Montenarh, M.; Grässer, F.A.; Götz, C. Protein Kinase CK2 and Epstein-Barr Virus. Biomedicines 2023, 11, 358. [Google Scholar] [CrossRef] [PubMed]
- Duncan, J.S.; Turowec, J.P.; Vilk, G.; Li, S.S.C.; Gloor, G.B.; Litchfield, D.W. Regulation of Cell Proliferation and Survival: Convergence of Protein Kinases and Caspases. Biochim. Biophys. Acta 2010, 1804, 505–510. [Google Scholar] [CrossRef]
- Majerciak, V.; Pripuzova, N.; Chan, C.; Temkin, N.; Specht, S.I.; Zheng, Z.-M. The Stability of Structured Kaposi’s Sarcoma-Associated Herpesvirus ORF57 Protein Is Regulated by Protein Phosphorylation and Homodimerization. J. Virol. 2015, 89, 3256–3274. [Google Scholar] [CrossRef]
- Piirsoo, A.; Piirsoo, M.; Kala, M.; Sankovski, E.; Lototskaja, E.; Levin, V.; Salvi, M.; Ustav, M. Activity of CK2alpha Protein Kinase Is Required for Efficient Replication of Some HPV Types. PLoS Pathog. 2019, 15, e1007788. [Google Scholar] [CrossRef]
- Du, M.; Liu, J.; Chen, X.; Xie, Y.; Yuan, C.; Xiang, Y.; Sun, B.; Lan, K.; Chen, M.; James, S.J.; et al. Casein Kinase II Controls TBK1/IRF3 Activation in IFN Response against Viral Infection. J. Immunol. 2015, 194, 4477–4488. [Google Scholar] [CrossRef]
- Sun, Z.; Ren, H.; Liu, Y.; Teeling, J.L.; Gu, J. Phosphorylation of RIG-I by Casein Kinase II Inhibits Its Antiviral Response. J. Virol. 2011, 85, 1036–1047. [Google Scholar] [CrossRef]
- Kohlstedt, K.; Shoghi, F.; Müller-Esterl, W.; Busse, R.; Fleming, I. CK2 Phosphorylates the Angiotensin-Converting Enzyme and Regulates Its Retention in the Endothelial Cell Plasma Membrane. Circ. Res. 2002, 91, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.-Y.; Chang, S.C.; Wu, H.-Y.; Yu, T.-C.; Wei, W.-C.; Lin, S.; Chien, C.-L.; Chang, M.-F. Upregulation of the Chemokine (C-C Motif) Ligand 2 via a Severe Acute Respiratory Syndrome Coronavirus Spike-ACE2 Signaling Pathway. J. Virol. 2010, 84, 7703–7712. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Dickmander, R.J.; Bayati, A.; Taft-Benz, S.A.; Smith, J.L.; Wells, C.I.; Madden, E.A.; Brown, J.W.; Lenarcic, E.M.; Yount, B.L.; et al. Host Kinase CSNK2 Is a Target for Inhibition of Pathogenic SARS-like β-Coronaviruses. ACS Chem. Biol. 2022, 17, 1937–1950. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 Spike Receptor-Binding Domain Bound to the ACE2 Receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef]
- Burkard, C.; Verheije, M.H.; Wicht, O.; van Kasteren, S.I.; van Kuppeveld, F.J.; Haagmans, B.L.; Pelkmans, L.; Rottier, P.J.M.; Bosch, B.J.; Haan, C.A.M. de Coronavirus Cell Entry Occurs through the Endo-/Lysosomal Pathway in a Proteolysis-Dependent Manner. PLoS Pathog. 2014, 10, e1004502. [Google Scholar] [CrossRef]
- Ramón, A.C.; Pérez, G.V.; Caballero, E.; Rosales, M.; Aguilar, D.; Vázquez-Blomquist, D.; Ramos, Y.; Rodríguez-Ulloa, A.; Falcón, V.; Rodríguez-Moltó, M.P.; et al. Targeting of Protein Kinase CK2 Elicits Antiviral Activity on Bovine Coronavirus Infection. Viruses 2022, 14, 552. [Google Scholar] [CrossRef]
- Perea, S.E.; Reyes, O.; Puchades, Y.; Mendoza, O.; Vispo, N.S.; Torrens, I.; Santos, A.; Silva, R.; Acevedo, B.; López, E.; et al. Antitumor Effect of a Novel Proapoptotic Peptide That Impairs the Phosphorylation by the Protein Kinase 2 (Casein Kinase 2). Cancer Res. 2004, 64, 7127–7129. [Google Scholar] [CrossRef]
- Vlasova, A.N.; Saif, L.J. Bovine Coronavirus and the Associated Diseases. Front. Vet. Sci. 2021, 8, 643220. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Gill, A.; Dove, B.K.; Emmett, S.R.; Kemp, C.F.; Ritchie, M.A.; Dee, M.; Hiscox, J.A. Mass Spectroscopic Characterization of the Coronavirus Infectious Bronchitis Virus Nucleoprotein and Elucidation of the Role of Phosphorylation in RNA Binding by Using Surface Plasmon Resonance. J. Virol. 2005, 79, 1164–1179. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Xu, L.; Huang, M.; Qisheng Li, F.; Liu, D.X. Identification of Two ATR-Dependent Phosphorylation Sites on Coronavirus Nucleocapsid Protein with Nonessential Functions in Viral Replication and Infectivity in Cultured Cells. Virology 2013, 444, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.A.; Ouchi, T. Cellular Commitment to Reentry into the Cell Cycle after Stalled DNA Is Determined by Site-Specific Phosphorylation of Chk1 and PTEN. Mol. Cancer Ther. 2008, 7, 2509–2516. [Google Scholar] [CrossRef]
- Gordon, D.E.; Hiatt, J.; Bouhaddou, M.; Rezelj, V.V.; Ulferts, S.; Braberg, H.; Jureka, A.S.; Obernier, K.; Guo, J.Z.; Batra, J.; et al. Comparative Host-Coronavirus Protein Interaction Networks Reveal Pan-Viral Disease Mechanisms. Science 2020, 370, eabe9403. [Google Scholar] [CrossRef]
- Surjit, M.; Kumar, R.; Mishra, R.N.; Reddy, M.K.; Chow, V.T.K.; Lal, S.K. The Severe Acute Respiratory Syndrome Coronavirus Nucleocapsid Protein Is Phosphorylated and Localizes in the Cytoplasm by 14-3-3-Mediated Translocation. J. Virol. 2005, 79, 11476–11486. [Google Scholar] [CrossRef]
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.; Obernier, K.; White, K.M.; O’Meara, M.J.; Rezelj, V.V.; Guo, J.Z.; Swaney, D.L.; et al. A SARS-CoV-2 Protein Interaction Map Reveals Targets for Drug Repurposing. Nature 2020, 583, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Bouhaddou, M.; Memon, D.; Meyer, B.; White, K.M.; Rezelj, V.V.; Correa Marrero, M.; Polacco, B.J.; Melnyk, J.E.; Ulferts, S.; Kaake, R.M.; et al. The Global Phosphorylation Landscape of SARS-CoV-2 Infection. Cell 2020, 182, 685–712.e19. [Google Scholar] [CrossRef]
- Shaath, H.; Alajez, N.M. Computational and Transcriptome Analyses Revealed Preferential Induction of Chemotaxis and Lipid Synthesis by SARS-CoV-2. Biology 2020, 9, 260. [Google Scholar] [CrossRef]
- Fatoki, T.H.; Ibraheem, O.; Ogunyemi, I.O.; Akinmoladun, A.C.; Ugboko, H.U.; Adeseko, C.J.; Awofisayo, O.A.; Olusegun, S.J.; Enibukun, J.M. Network Analysis, Sequence and Structure Dynamics of Key Proteins of Coronavirus and Human Host, and Molecular Docking of Selected Phytochemicals of Nine Medicinal Plants. J. Biomol. Struct. Dyn. 2020, 39, 6195–6217. [Google Scholar] [CrossRef]
- Ruzzene, M.; Bertacchini, J.; Toker, A.; Marmiroli, S. Cross-Talk between the CK2 and AKT Signaling Pathways in Cancer. Adv. Biol. Regul. 2017, 64, 1–8. [Google Scholar] [CrossRef]
- Basile, M.S.; Cavalli, E.; McCubrey, J.; Hernández-Bello, J.; Muñoz-Valle, J.F.; Fagone, P.; Nicoletti, F. The PI3K/Akt/mTOR Pathway: A Potential Pharmacological Target in COVID-19. Drug Discov. Today 2022, 27, 848–856. [Google Scholar] [CrossRef] [PubMed]
- Klann, K.; Bojkova, D.; Tascher, G.; Ciesek, S.; Münch, C.; Cinatl, J. Growth Factor Receptor Signaling Inhibition Prevents SARS-CoV-2 Replication. Mol. Cell 2020, 80, 164–174.e4. [Google Scholar] [CrossRef] [PubMed]
- Berretta, A.A.; Silveira, M.A.D.; Cóndor Captcha, J.M.; De Jong, D. Propolis and Its Potential against SARS-CoV-2 Infection Mechanisms and COVID-19 Disease: Running Title: Propolis against SARS-CoV-2 Infection and COVID-19. Biomed. Pharmacother. 2020, 131, 110622. [Google Scholar] [CrossRef]
- Su, C.-M.; Wang, L.; Yoo, D. Activation of NF-ΚB and Induction of Proinflammatory Cytokines Expressions Mediated by ORF7a Protein of SARS-CoV-2. Sci. Rep. 2021, 11, 13464. [Google Scholar] [CrossRef] [PubMed]
- Jafarzadeh, A.; Nemati, M.; Jafarzadeh, S. Contribution of STAT3 to the Pathogenesis of COVID-19. Microb. Pathog. 2021, 154, 104836. [Google Scholar] [CrossRef]
- Ponce, D.P.; Yefi, R.; Cabello, P.; Maturana, J.L.; Niechi, I.; Silva, E.; Galindo, M.; Antonelli, M.; Marcelain, K.; Armisen, R.; et al. CK2 Functionally Interacts with AKT/PKB to Promote the β-Catenin-Dependent Expression of Survivin and Enhance Cell Survival. Mol. Cell. Biochem. 2011, 356, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Song, D.H.; Sussman, D.J.; Seldin, D.C. Endogenous Protein Kinase CK2 Participates in Wnt Signaling in Mammary Epithelial Cells. J. Biol. Chem. 2000, 275, 23790–23797. [Google Scholar] [CrossRef]
- Miranda, J.; Bringas, R.; Fernandez-de-Cossio, J.; Perera-Negrin, Y. Targeting CK2 Mediated Signaling to Impair/Tackle SARS-CoV-2 Infection: A Computational Biology Approach. Mol. Med. 2021, 27, 161. [Google Scholar] [CrossRef]
- Cruz, L.R.; Baladrón, I.; Rittoles, A.; Díaz, P.A.; Valenzuela, C.; Santana, R.; Vázquez, M.M.; García, A.; Chacón, D.; Thompson, D.; et al. Treatment with an Anti-CK2 Synthetic Peptide Improves Clinical Response in COVID-19 Patients with Pneumonia. A Randomized and Controlled Clinical Trial. ACS Pharmacol. Transl. Sci. 2021, 4, 206–212. [Google Scholar] [CrossRef]
- Niefind, K.; Guerra, B.; Ermakowa, I.; Issinger, O.G. Crystal Structure of Human Protein Kinase CK2: Insights into Basic Properties of the CK2 Holoenzyme. EMBO J. 2001, 20, 5320–5331. [Google Scholar] [CrossRef] [PubMed]
- Meggio, F.; Marin, O.; Pinna, L.A. Substrate Specificity of Protein Kinase CK2. Cell. Mol. Biol. Res. 1994, 40, 401–409. [Google Scholar] [PubMed]
- Chitalia, V.C.; Munawar, A.H. A Painful Lesson from the COVID-19 Pandemic: The Need for Broad-Spectrum, Host-Directed Antivirals. J. Transl. Med. 2020, 18, 390. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quezada Meza, C.P.; Ruzzene, M. Protein Kinase CK2 and SARS-CoV-2: An Expected Interplay Story. Kinases Phosphatases 2023, 1, 141-150. https://doi.org/10.3390/kinasesphosphatases1020009
Quezada Meza CP, Ruzzene M. Protein Kinase CK2 and SARS-CoV-2: An Expected Interplay Story. Kinases and Phosphatases. 2023; 1(2):141-150. https://doi.org/10.3390/kinasesphosphatases1020009
Chicago/Turabian StyleQuezada Meza, Camila Paz, and Maria Ruzzene. 2023. "Protein Kinase CK2 and SARS-CoV-2: An Expected Interplay Story" Kinases and Phosphatases 1, no. 2: 141-150. https://doi.org/10.3390/kinasesphosphatases1020009
APA StyleQuezada Meza, C. P., & Ruzzene, M. (2023). Protein Kinase CK2 and SARS-CoV-2: An Expected Interplay Story. Kinases and Phosphatases, 1(2), 141-150. https://doi.org/10.3390/kinasesphosphatases1020009