
Enhancing the Thermal and Kinetic Stability of Ketol-Acid Reductoisomerase, a Central Catalyst of a Cell-Free Enzyme Cascade for the Manufacture of Platform Chemicals

 , , , , , ,
, , , , , ,  ,
,  , , , ,
, , , , _Schenk.png) and
and
Abstract
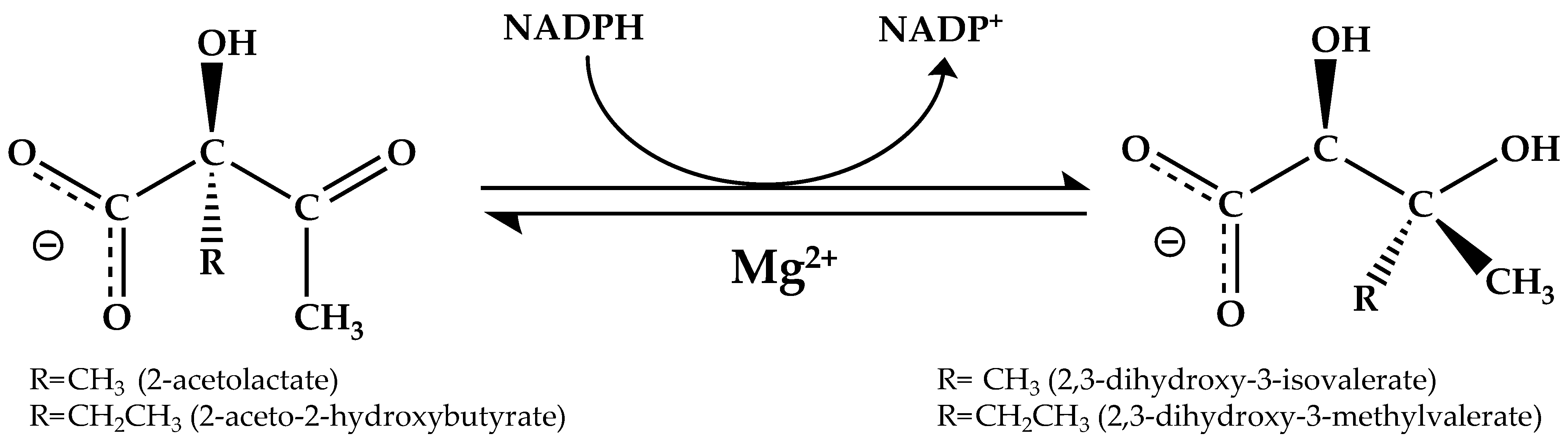
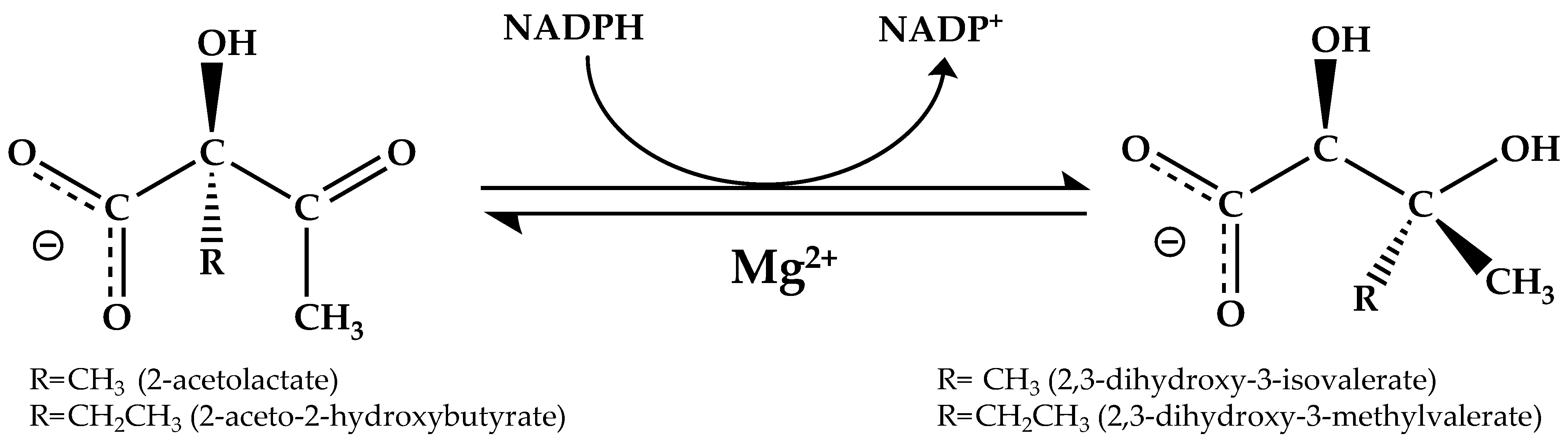
:1. Introduction
2. Materials and Methods
2.1. Expression and Purification of the Enzymes and Their Mutants
2.2. Preparation of the CjKARI for Cryo-EM Studies
2.3. Cryo-Electron Microscopy Studies
2.4. Cryo-EM Image Processing, Model Building, Refinement and Validation
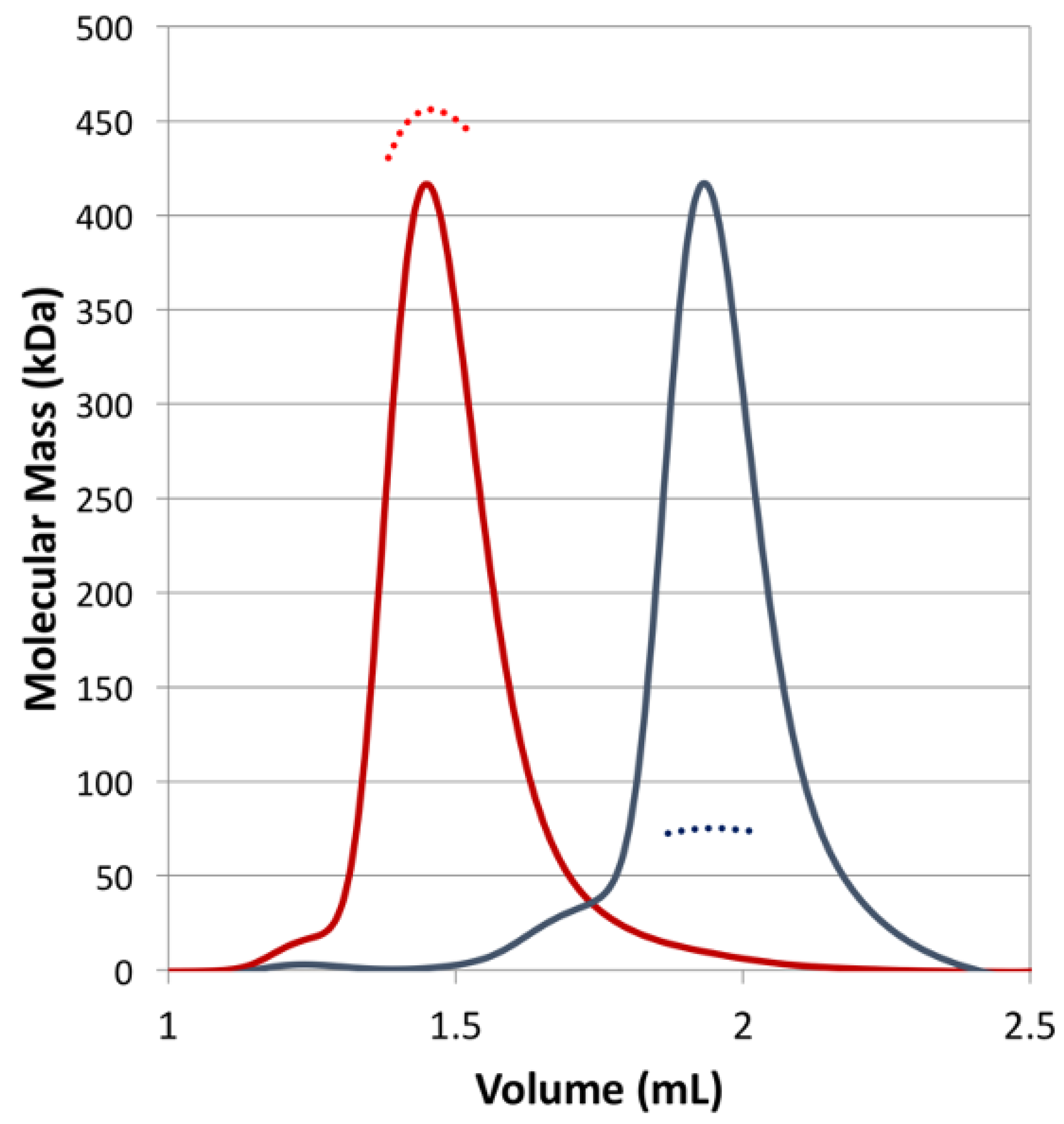
2.5. Oligomeric States Analysis
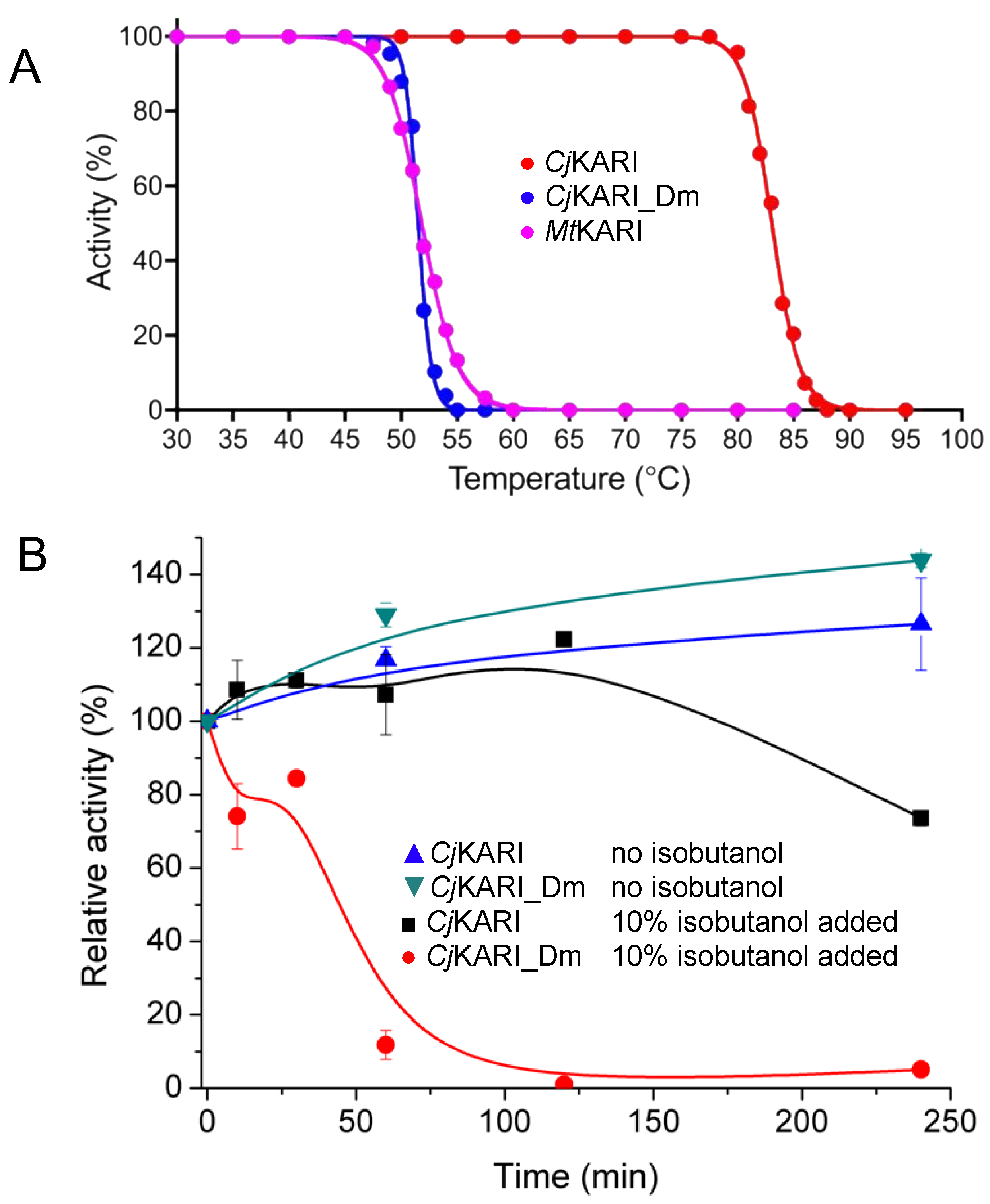
2.6. Enzyme Kinetics and Thermal Stability Studies
2.7. PROSS Design of EcKARI
3. Results and Discussion
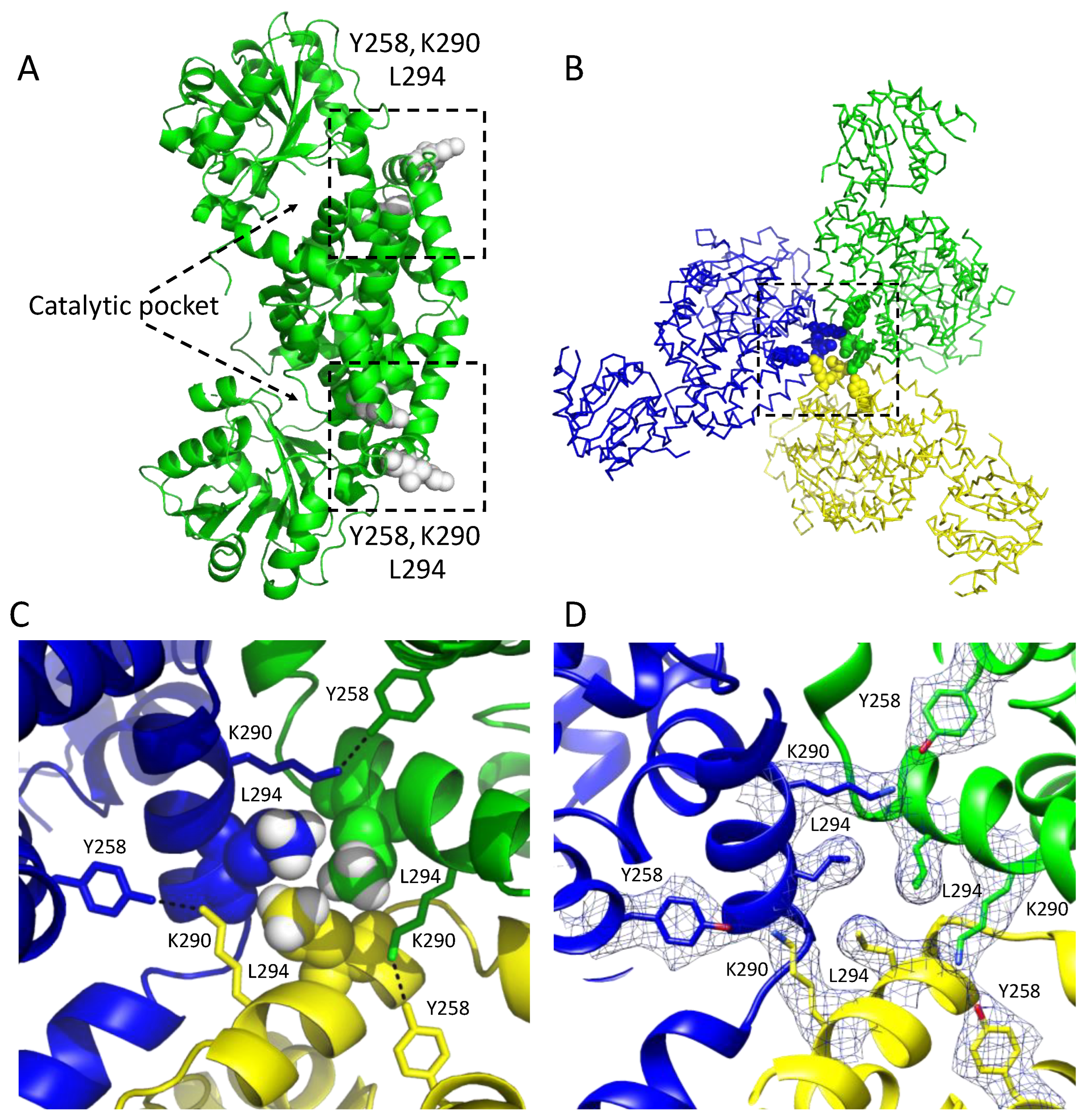
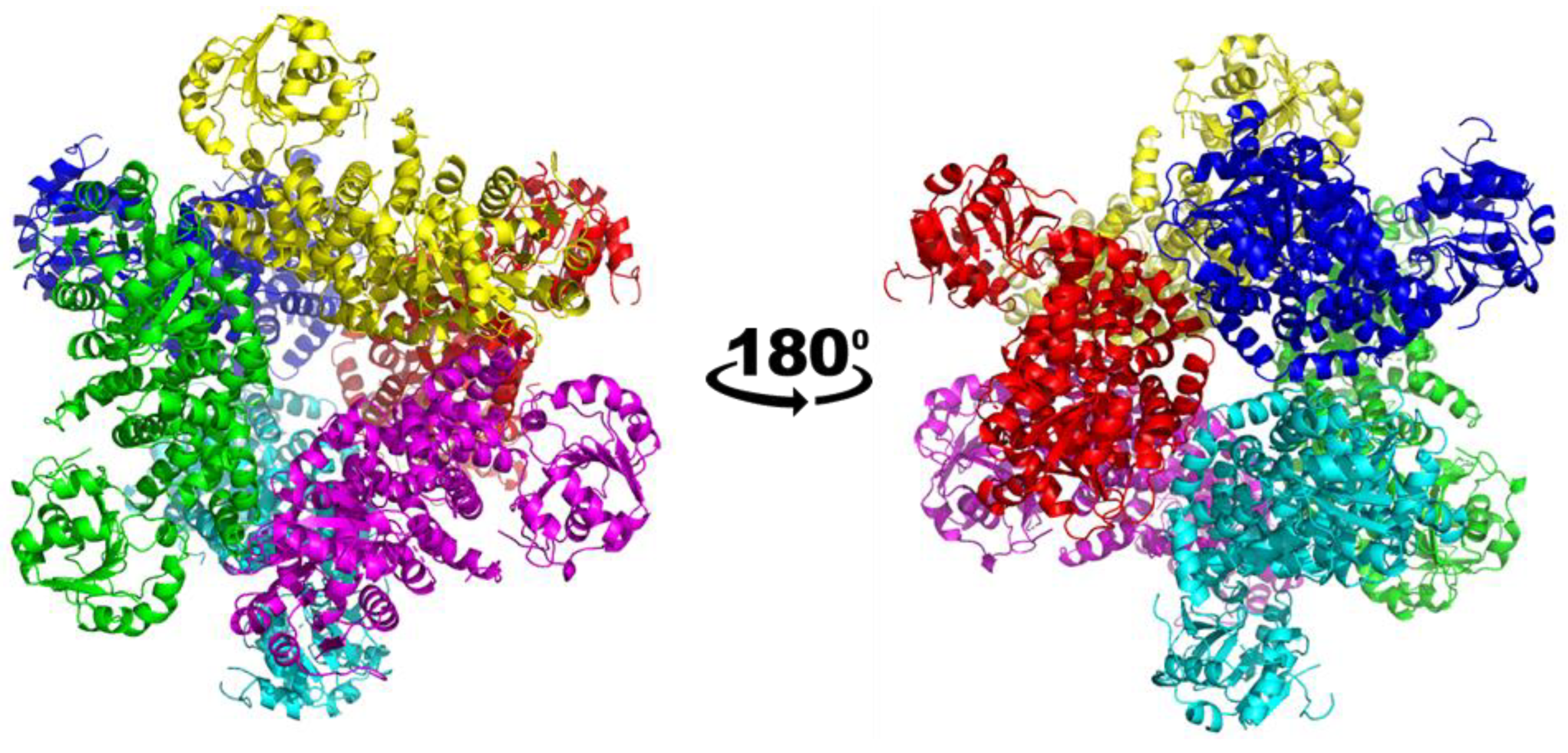
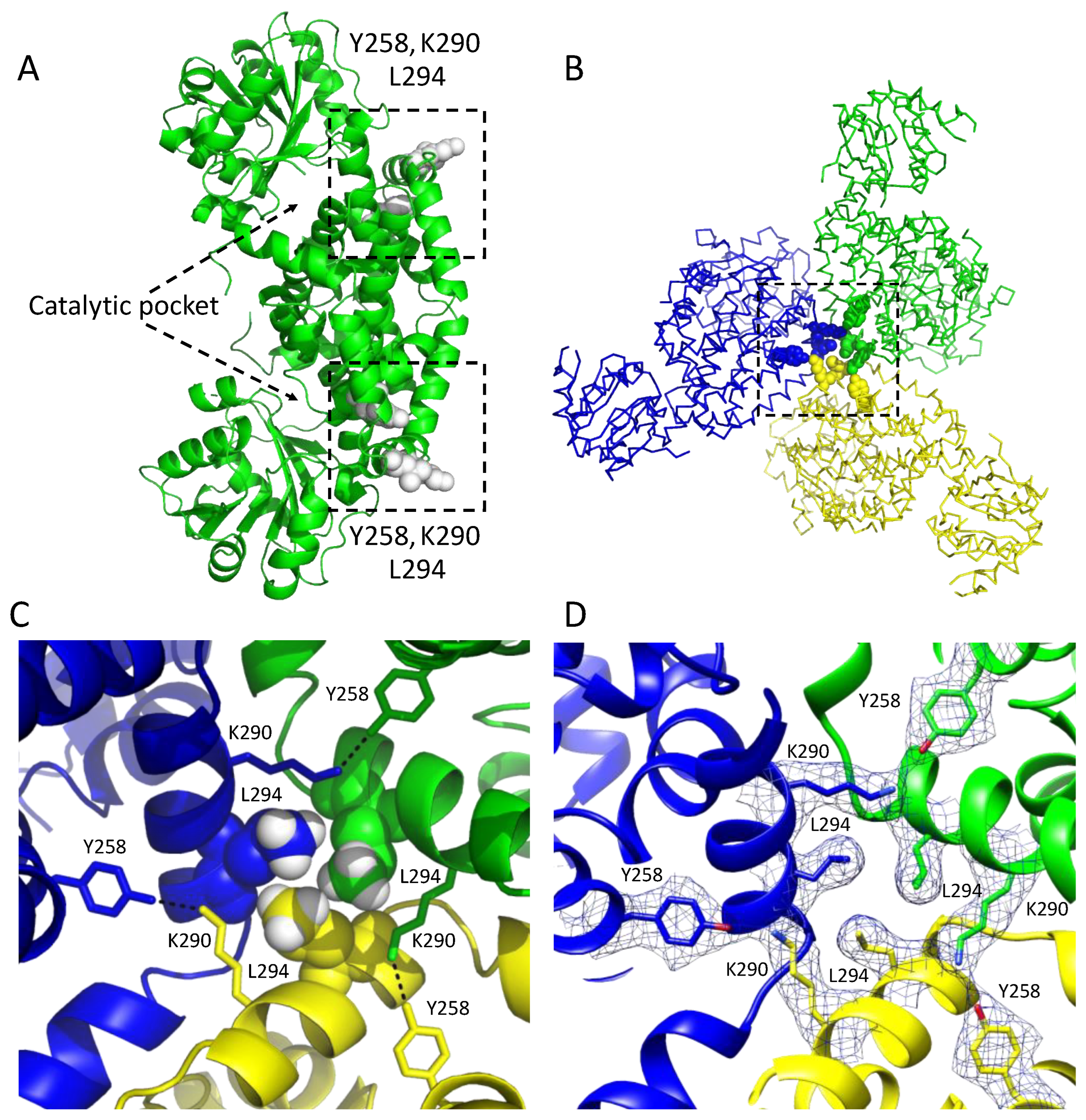
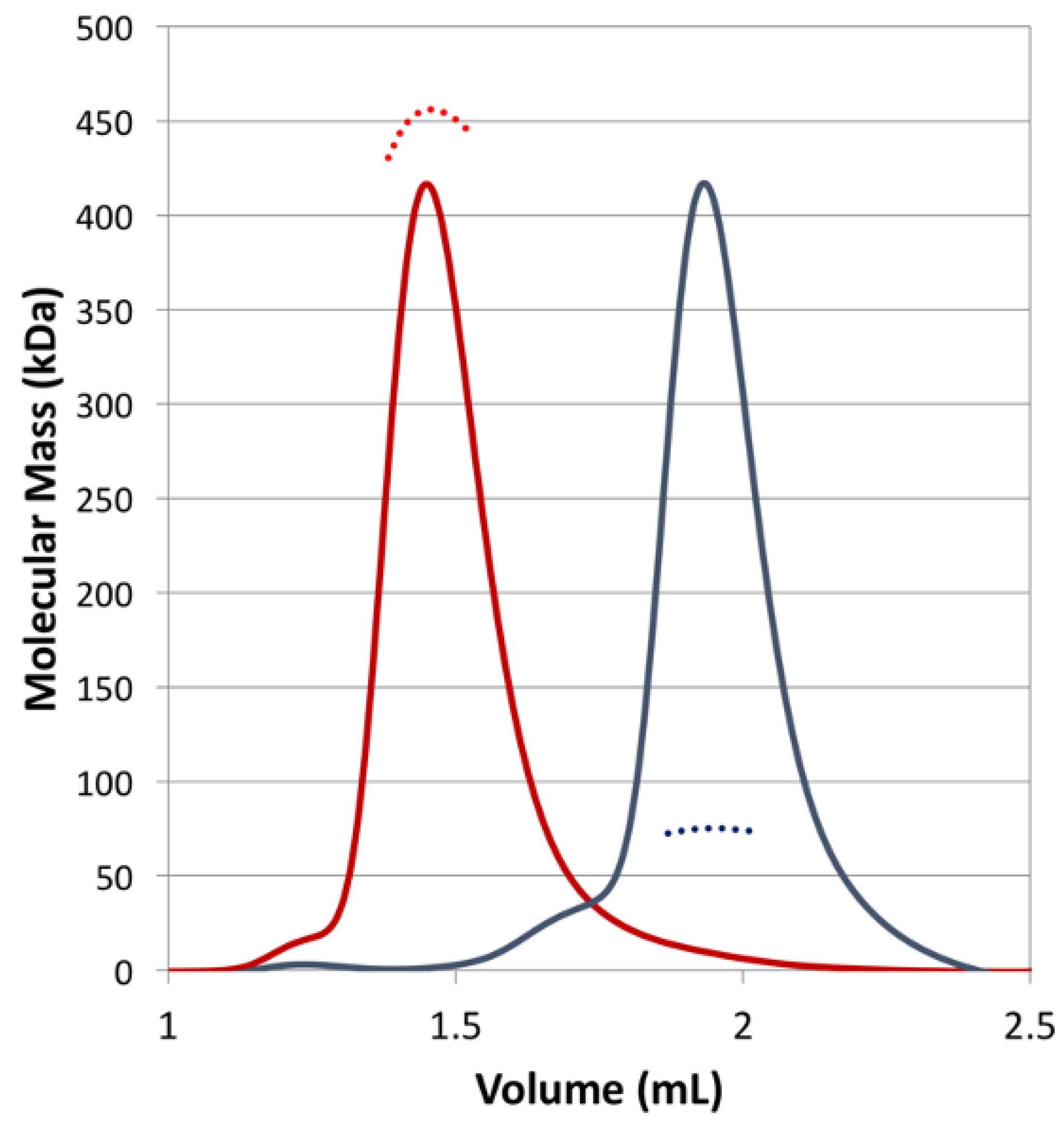
3.1. Overall Structure of the Class I CjKARI and the Relevance of the Oligomeric State on Enzyme Stability
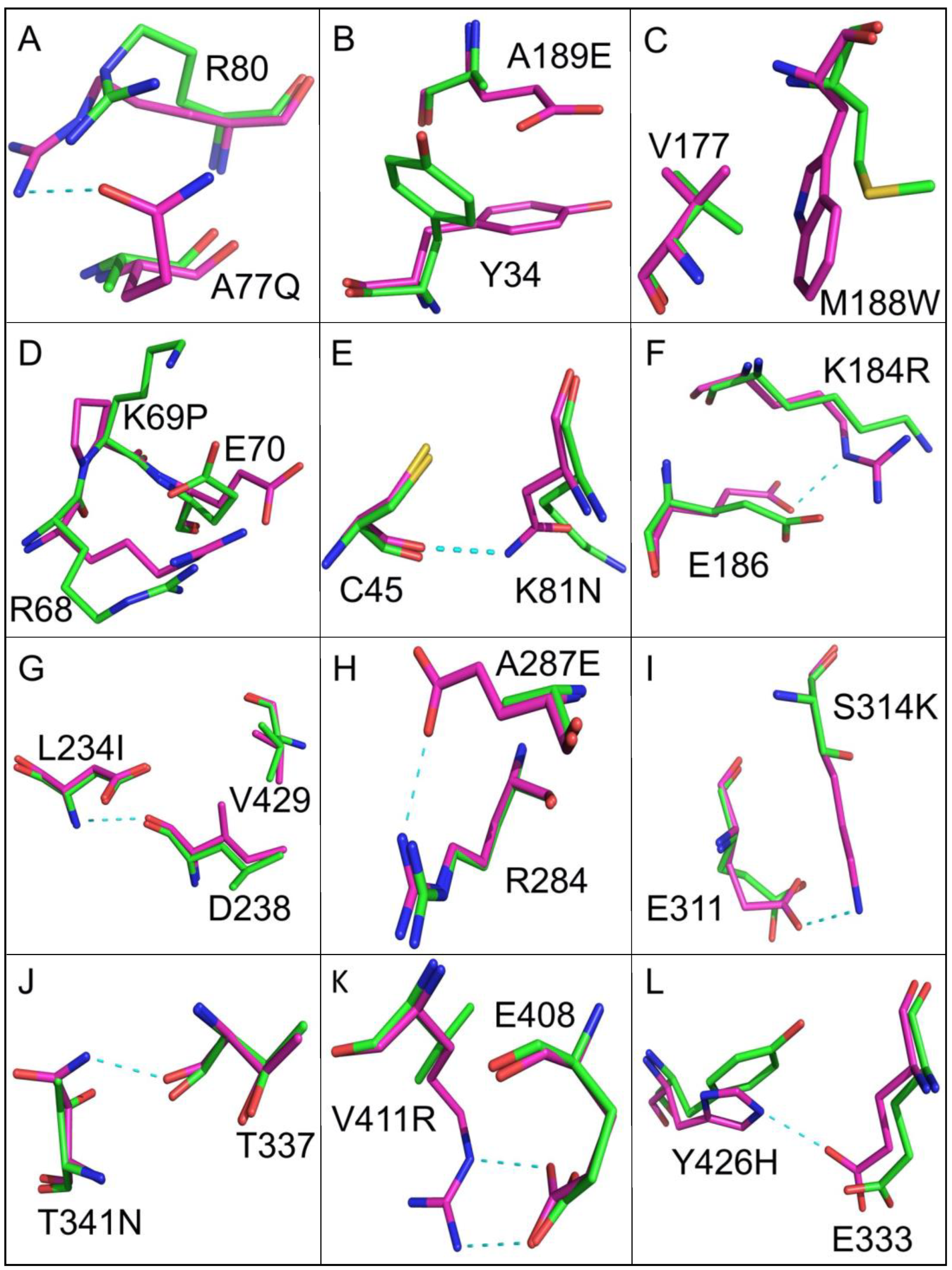
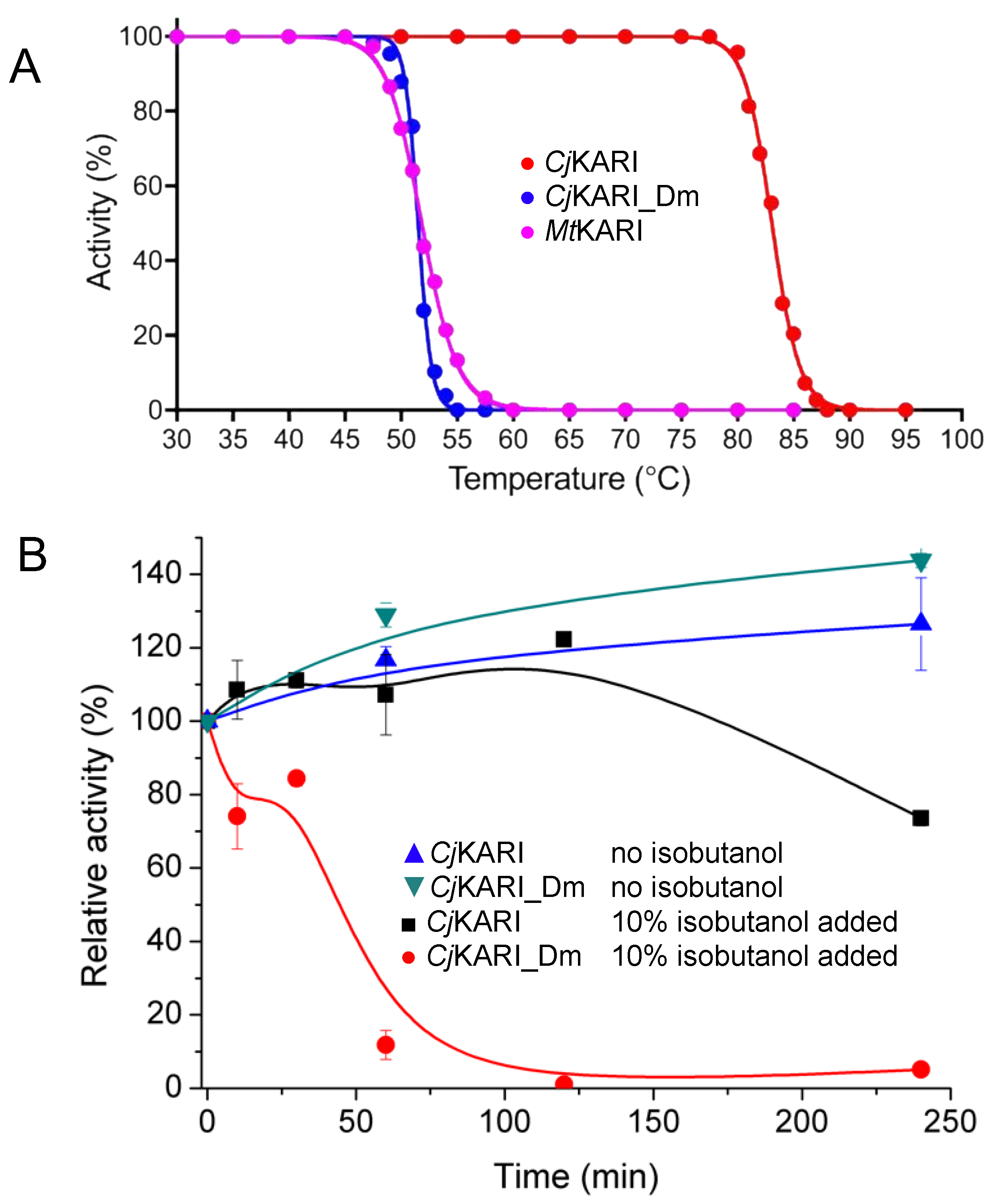
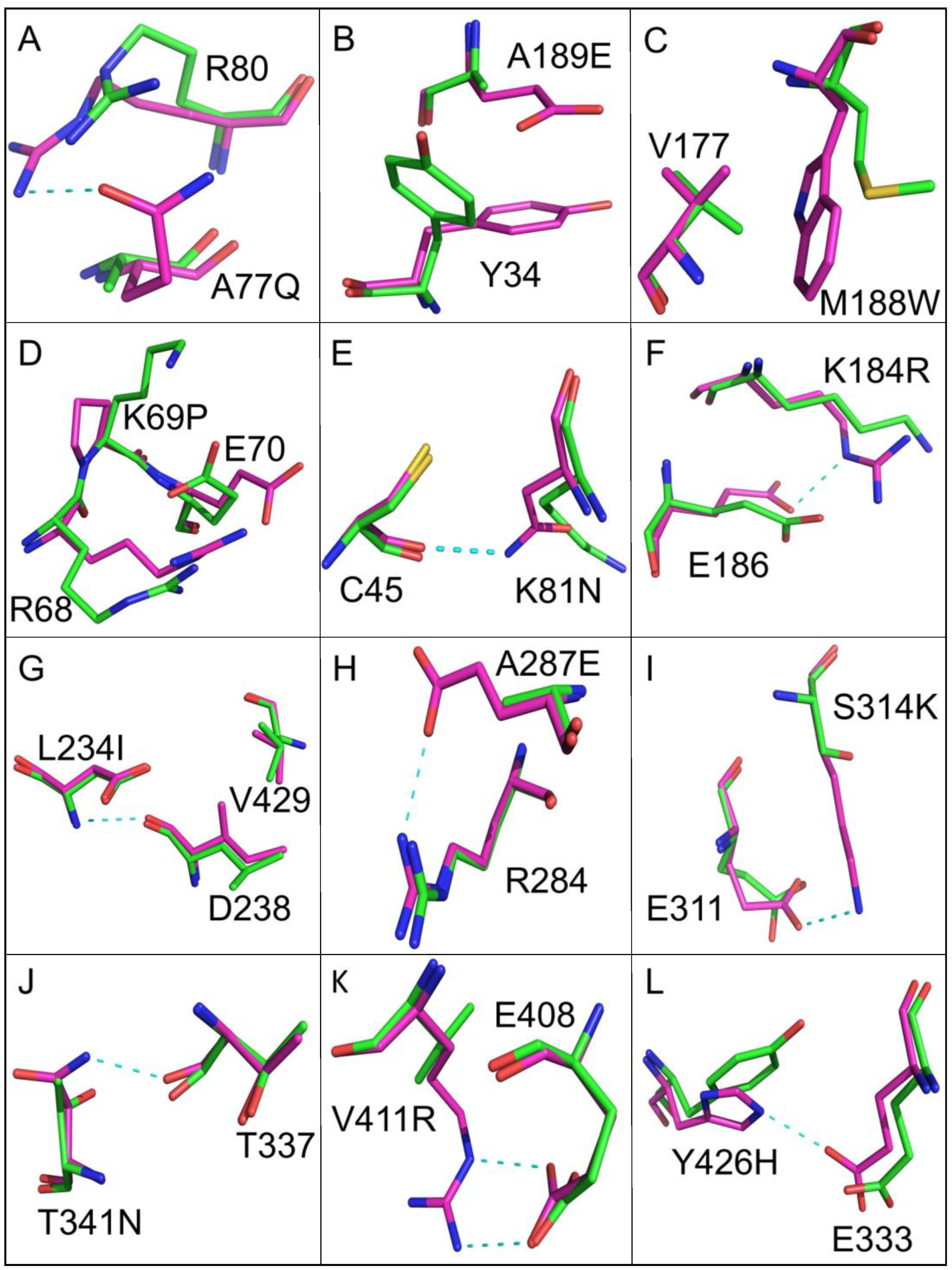
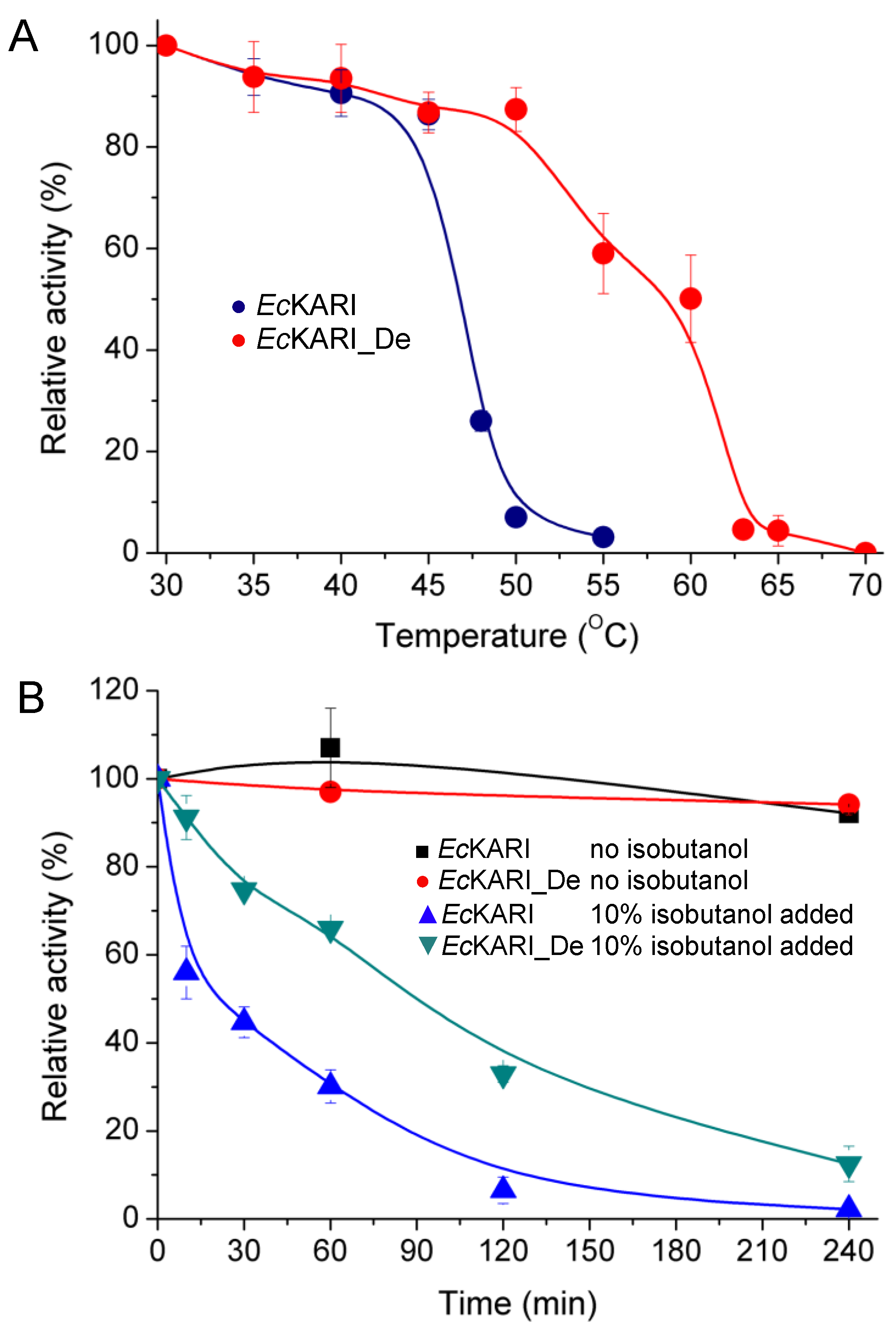
3.2. Improving Enzyme Stability through Molecular Design
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duggleby, R.G.; Pang, S.S. Acetohydroxyacid synthase. J. Biochem. Mol. Biol. 2000, 33, 1–36. [Google Scholar]
- Grandoni, J.A.; Marta, P.T.; Schloss, J.V. Inhibitors of branched-chain amino acid biosynthesis as potential antituberculosis agents. J. Antimicrob. Chemother. 1998, 4, 475–482. [Google Scholar] [CrossRef] [Green Version]
- Schulz, A.; Sponemann, P.; Kocher, H.; Wengenmayer, F. The herbicidally active experimental compound Hoe 704 is a potent inhibitor of the enzyme acetolactate reductoisomerase. FEBS Lett. 1988, 238, 375–378. [Google Scholar] [CrossRef] [Green Version]
- Epelbaum, S.; LaRossa, R.A.; VanDyk, T.K.; Elkayam, T.; Chipman, D.M.; Barak, Z. Branched-chain amino acid biosynthesis in Salmonella typhimurium: A quantitative analysis. J. Bacteriol. 1998, 180, 4056–4067. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.H.; Lonhienne, T.G.; Winzor, D.J.; Schenk, G.; Guddat, L.W. Bacterial and plant ketol-acid reductoisomerases have different mechanisms of induced fit during the catalytic cycle. J. Mol. Biol. 2012, 424, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Pue, N.; Guddat, L.W. Acetohydroxyacid synthase: A target for antimicrobial drug discovery. Curr. Pharm. Des. 2014, 20, 740–753. [Google Scholar] [CrossRef] [PubMed]
- Atsumi, S.; Hanai, T.; Liao, J.C. Non-fermentative pathways for synthesis of branched-chain higher alcohols as biofuels. Nature 2008, 451, 86–89. [Google Scholar] [CrossRef]
- Wang, B.W.; Shi, A.Q.; Tu, R.; Zhang, X.L.; Wang, Q.H.; Bai, F.W. Branched-chain higher alcohols. Adv. Biochem. Eng. Biotechnol. 2012, 128, 101–118. [Google Scholar]
- Guterl, J.K.; Garbe, D.; Carsten, J.; Steffler, F.; Sommer, B.; Reisse, S.; Philipp, A.; Haack, M.; Ruhmann, B.; Koltermann, A.; et al. Cell-free metabolic engineering: Production of chemicals by minimized reaction cascades. ChemSusChem 2012, 5, 2165–2172. [Google Scholar] [CrossRef]
- Gumulya, Y.; Baek, J.-M.; Wun, S.-J.; Thomson, R.E.S.; Harris, K.L.; Hunter, D.J.B.; Behrendorff, J.B.Y.H.; Kulig, J.; Zheng, S.; Wu, X.; et al. Engineering highly functional thermostable proteins using ancestral sequence reconstruction. Nature Catal. 2018, 1, 878–888. [Google Scholar] [CrossRef]
- Lonhienne, T.; Low, Y.S.; Garcia, M.D.; Croll, T.; Gao, Y.; Wang, Q.; Brillault, L.; Williams, C.M.; Fraser, J.A.; McGeary, R.P.; et al. Structures of fungal and plant acetohydroxyacid synthases. Nature 2020, 586, 317–321. [Google Scholar] [CrossRef]
- Flint, D.H.; Emptage, M.H.; Finnegan, M.G.; Fu, W.; Johnson, M.K. The role and properties of the iron-sulfur cluster in Escherichia coli dihydroxy-acid dehydratase. J. Biol. Chem. 1993, 268, 14732–14742. [Google Scholar] [CrossRef]
- Sutiono, S.; Teshima, M.; Beer, B.; Schenk, G.; Sieber, V. Enabling the direct enzymatic dehydration of D-glycerate to pyruvate as the key step in synthetic enzyme cascades used in the cell-free production of fine chemicals. ACS Catal. 2020, 10, 3110–3118. [Google Scholar] [CrossRef]
- Melse, O.; Sutiono, S.; Haslbeck, M.; Schenk, G.; Antes, I.; Sieber, V. Structure-guided modulation of the catalytic properties of dihydroxyacid dehydratases. ChemBioChem 2022, 23, e202200088. [Google Scholar] [CrossRef]
- Bayaraa, T.; Gaete, J.; Sutiono, S.; Kurz, J.; Lonhienne, T.; Harmer, J.R.; Bernhardt, P.V.; Sieber, V.; Guddat, L.W.; Schenk, G. Dihydroxy-acid dehydratases from pathogenic bacteria: Emerging drug targets to combat antibiotic resistance. Chem. Eur. J. 2022, 28, e202200927. [Google Scholar] [CrossRef] [PubMed]
- Tadrowski, S.; Pedroso, M.M.; Sieber, V.; Larrabee, J.A.; Guddat, L.W.; Schenk, G. Metal ions play an essential catalytic role in the mechanism of ketol-acid reductoisomerase. Chemistry 2016, 22, 7427–7436. [Google Scholar] [CrossRef] [PubMed]
- Dumas, R.; Biou, V.; Halgand, F.; Douce, R.; Duggleby, R.G. Enzymology, structure, and dynamics of acetohydroxy acid isomeroreductase. Acc. Chem. Res. 2001, 34, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, R.; Lee, Y.T.; Guddat, L.W.; Duggleby, R.G. Probing the mechanism of the bifunctional enzyme ketol-acid reductoisomerase by site-directed mutagenesis of the active site. FEBS J. 2005, 272, 593–602. [Google Scholar] [CrossRef]
- Tyagi, R.; Duquerroy, S.; Navaza, J.; Guddat, L.W.; Duggleby, R.G. The crystal structure of a bacterial class II ketol-acid reductolsomerase: Domain conservation and evolution. Protein Sci. 2005, 14, 3089–3100. [Google Scholar] [CrossRef] [Green Version]
- Lv, Y.; Kandale, A.; Wun, S.J.; McGeary, R.P.; Williams, S.J.; Kobe, B.; Sieber, V.; Schembri, M.A.; Schenk, G.; Guddat, L.W. Crystal structure of Mycobacterium tuberculosis ketol-acid reductoisomerase at 1.0 Å resolution-a potential target for anti-tuberculosis drug discovery. FEBS J. 2016, 283, 1184–1196. [Google Scholar] [CrossRef]
- Patel, K.; Teran, D.; Zheng, S.; Kandale, A.; Garcia, M.; Lv, Y.; Schembri, M.A.; McGeary, R.P.; Schenk, G.; Guddat, L.W. Crystal structures of Staphylococcus aureus ketol-acid reductoisomerase in complex with two transition state analogs that have biocidal activity. Chem. Eur. J. 2017, 23, 18289–18295. [Google Scholar] [CrossRef] [PubMed]
- Ahn, H.J.; Eom, S.J.; Yoon, H.J.; Lee, B.I.; Cho, H.J.; Suh, S.W. Crystal structure of class I acetohydroxy acid isomeroreductase from Pseudomonas aeruginosa. J. Mol. Biol. 2003, 328, 505–515. [Google Scholar] [CrossRef]
- Chen, C.Y.; Chang, Y.C.; Lin, B.L.; Lin, K.F.; Huang, C.H.; Hsieh, D.L.; Ko, T.P.; Tsai, M.D. Use of Cryo-EM to uncover structural bases of pH effect and cofactor bispecificity of ketol-acid reductoisomerase. J. Am. Chem. Soc. 2019, 141, 6136–6140. [Google Scholar] [CrossRef] [PubMed]
- Cahn, J.K.; Brinkmann-Chen, S.; Spatzal, T.; Wiig, J.A.; Buller, A.R.; Einsle, O.; Hu, Y.; Ribbe, M.W.; Arnold, F.H. Cofactor specificity motifs and the induced fit mechanism in class I ketol-acid reductoisomerases. Biochem. J. 2015, 468, 475–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, E.W.W.; Guddat, L.W. Conformational changes in a plant ketol-acid reductoisomerase upon Mg2+ and NADPH binding as revealed by two crystal structures. J. Mol. Biol. 2009, 389, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Kirk, O.; Borchert, T.V.; Fuglsang, C.C. Industrial enzyme applications. Curr. Opin. Biotechnol. 2002, 13, 345–351. [Google Scholar] [CrossRef]
- Van Beilen, J.B.; Li, Z. Enzyme technology: An overview. Curr. Opin. Biotechnol. 2002, 13, 338–344. [Google Scholar] [CrossRef]
- Ahuja, S.K.; Ferreira, G.M.; Moreira, A.R. Utilization of enzymes for environmental applications. Crit. Rev. Biotechnol. 2004, 24, 125–154. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A. Engineering a more sustainable world through catalysis and green chemistry. J. R. Soc. Interface 2016, 13. [Google Scholar] [CrossRef] [Green Version]
- Bornscheuer, U.T. Trends and challenges in enzyme technology. Adv. Biochem. Eng. Biotechnol. 2005, 100, 181–203. [Google Scholar]
- Brinkmann-Chen, S.; Flock, T.; Cahn, J.K.; Snow, C.D.; Brustad, E.M.; McIntosh, J.A.; Meinhold, P.; Zhang, L.; Arnold, F.H. General approach to reversing ketol-acid reductoisomerase cofactor dependence from NADPH to NADH. Proc. Natl. Acad. Sci. USA 2013, 110, 10946–10951. [Google Scholar] [CrossRef] [Green Version]
- Goldenzweig, A.; Goldsmith, M.; Hill, S.E.; Gertman, O.; Laurino, P.; Ashani, Y.; Dym, O.; Unger, T.; Albeck, S.; Prilusky, J.; et al. Automated structure- and sequence-based design of proteins for high bacterial expression and stability. Mol. Cell 2016, 63, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.C.; von Hippel, P.H. Calculation of protein extinction coefficients from amino acid sequence data. Anal. Biochem. 1989, 182, 319–326. [Google Scholar] [CrossRef]
- Punjani, A.; Rubinstein, J.L.; Fleet, D.J.; Brubaker, M.A. cryoSPARC: Algorithms for rapid unsupervised cryo-EM structure determination. Nat. Methods 2017, 14, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Grigorieff, N. Frealign: An exploratory tool for single-particle cryo-EM. Methods Enzym. 2016, 579, 191–226. [Google Scholar]
- Pintilie, G.D.; Zhang, J.; Goddard, T.D.; Chiu, W.; Gossard, D.C. Quantitative analysis of cryo-EM density map segmentation by watershed and scale-space filtering, and fitting of structures by alignment to regions. J. Struct. Biol. 2010, 170, 427–438. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [Green Version]
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.-W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Taylor, W.R. A deeply knotted protein structure and how it might fold. Nature 2000, 406, 916–919. [Google Scholar] [CrossRef]
- Atsumi, S.; Wu, T.Y.; Machado, I.M.; Huang, W.C.; Chen, P.Y.; Pellegrini, M.; Liao, J.C. Evolution, genomic analysis, and reconstruction of isobutanol tolerance in Escherichia coli. Mol. Syst. Biol. 2010, 6, 449. [Google Scholar] [CrossRef]
- Brynildsen, M.P.; Liao, J.C. An integrated network approach identifies the isobutanol response network of Escherichia coli. Mol. Syst. Biol. 2009, 5, 277. [Google Scholar] [CrossRef] [PubMed]
- Wessel, P.M.; Biou, V.; Douce, R.; Dumas, R. A loop deletion in the plant acetohydroxy acid isomeroreductase homodimer generates an active monomer with reduced stability and altered magnesium affinity. Biochemistry 1998, 37, 12753–12760. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann-Chen, S.; Cahn, J.K.B.; Arnold, F.H. Uncovering rare NADH-preferring ketol-acid reductoisomerases. Metab. Eng. 2014, 26, 17–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, K.D.; Tomii, K.; Katoh, K. Application of the MAFFT sequence alignment program to large data-reexamination of the usefulness of chained guide trees. Bioinformatics 2016, 32, 3246–3251. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Matthews, B.W.; Nicholson, H.; Becktel, W.J. Enhanced protein thermostability from site-directed mutations that decrease the entropy of unfolding. Proc. Natl. Acad. Sci. USA 1987, 84, 6663–6667. [Google Scholar] [CrossRef] [Green Version]
- Bayaraa, T.; Kurz, J.L.; Patel, K.M.; Hussein, W.M.; West, N.P.; Schenk, G.; McGeary, R.P.; Guddat, L.W. Discovery, synthesis and evaluation of a novel ketol-acid reductoisomerase inhibitor. Chem. Eur. J. 2020, 26, 8958–8968. [Google Scholar] [CrossRef]
- Wun, S.J.; Johnson, L.A.; McGeary, R.P.; Schenk, G.; Guddat, L.W. Inhibition studies of ketol-acid reductoisomerases from pathogenic microorganisms. Arch. Biochem. Biophys. 2020, 692, 108516. [Google Scholar] [CrossRef]
- Lin, X.; Kurz, J.L.; Patel, K.M.; Wun, S.J.; Hussein, W.M.; Lonhienne, T.; West, N.P.; McGeary, R.P.; Schenk, G.; Guddat, L.W. Discovery of a pyrimidine-dione derivative with potent inhibitory activity against Mycobacterium tuberculosis ketol-acid reductoisomerase. Chem. Eur. J. 2020, 27, 3130–3141. [Google Scholar] [CrossRef]
- Kandale, A.; Patel, K.; Waleed, H.; Wun, S.J.; Zheng, S.; Tan, L.; West, N.; Schenk, G.; Guddat, L.W.; McGeary, R.P. Analogs of the herbicide, N-hydroxy-N-isopropyloxamate (IpOHA), inhibit Mycobacterium tuberculosis ketol-acid reductoisomerase and their prodrugs are promising anti-TB drug leads. J. Med. Chem. 2021, 64, 1670–1684. [Google Scholar] [CrossRef]
- Valera, A.; Wang, S.; Carr, R.; Trembleau, L.; Deng, H. Characterization of a class II ketol-acid reductoisomerase from Mycobacterium tuberculosis. RSC Adv. 2022, 12, 10540–10544. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Collection and Processing | |
|---|---|
| PDB Entry | 8CY8 |
| EMDB Entry | EMD-27070 |
| EM equipment | FEI Titan Krios |
| Camera | K3 |
| Frames per micrograph | 40 |
| Magnification | 29,000 |
| Voltage (keV) | 300 |
| Electron exposure (e-/Å2) | 50 |
| Defocus range (µm) | −1.2 to −1.8 |
| Pixel size (Å) | 0.82 |
| Symmetry | T |
| Micrographs (no.) | 2247 |
| Initial particle images (no.) | 2,472,436 |
| Final particle images (no.) | 74,899 |
| Map resolution (Å) | 2.94 |
| FSC threshold | 0.143 |
| Map local resolution range (Å) | 2.5–4.2 |
| Local resolution FSC | 0.5 |
| Refinement | |
| Model composition | |
| Non-hydrogen atoms | 28,800 |
| Protein residues | 3744 |
| Metal | 0 |
| Cofactor | 0 |
| Inhibitor | 0 |
| Ligands | 12 |
| R.m.s. deviations | |
| Bond length (Å) | 0.010 |
| Bond angles (°) | 0.772 |
| Validation | |
| MolProbity score | 1.13 |
| Clash score | 3.38 |
| Poor rotamers (%) | 0.00 |
| Ramachandran plot (%) | |
| Favored (%) | 99.37 |
| Allowed (%) | 0.63 |
| Outliers (%) | 0.00 |
| B factors (Å2) | |
| Protein (min/max/mean) | 10.51/116.90/53.22 |
| Ligand (min/max/mean) | 33.80/36.87/35.02 |
| Polar or Charged Surface | Core Packing | Proline at Turn Region | Hydrogen Bonds/Ion Pairs | Molecular Force Unclear |
|---|---|---|---|---|
| G24D S33N Q36K A73D A77Q R116N D124S V139E A189E F256Y A297R G315T K326N E346D G456N | A32C E141M M188W L283I S425A | K69P E383P A469P | K81N K184R L234I A287E S314K T341N V411R Y426H E438T | T245V E251A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, Y.; Zheng, S.; Goldenzweig, A.; Liu, F.; Gao, Y.; Yang, X.; Kandale, A.; McGeary, R.P.; Williams, S.; Kobe, B.; et al. Enhancing the Thermal and Kinetic Stability of Ketol-Acid Reductoisomerase, a Central Catalyst of a Cell-Free Enzyme Cascade for the Manufacture of Platform Chemicals. Appl. Biosci. 2022, 1, 163-178. https://doi.org/10.3390/applbiosci1020011
Lv Y, Zheng S, Goldenzweig A, Liu F, Gao Y, Yang X, Kandale A, McGeary RP, Williams S, Kobe B, et al. Enhancing the Thermal and Kinetic Stability of Ketol-Acid Reductoisomerase, a Central Catalyst of a Cell-Free Enzyme Cascade for the Manufacture of Platform Chemicals. Applied Biosciences. 2022; 1(2):163-178. https://doi.org/10.3390/applbiosci1020011
Chicago/Turabian StyleLv, You, Shan Zheng, Adi Goldenzweig, Fengjiang Liu, Yan Gao, Xiuna Yang, Ajit Kandale, Ross P. McGeary, Simon Williams, Bostjan Kobe, and et al. 2022. "Enhancing the Thermal and Kinetic Stability of Ketol-Acid Reductoisomerase, a Central Catalyst of a Cell-Free Enzyme Cascade for the Manufacture of Platform Chemicals" Applied Biosciences 1, no. 2: 163-178. https://doi.org/10.3390/applbiosci1020011
APA StyleLv, Y., Zheng, S., Goldenzweig, A., Liu, F., Gao, Y., Yang, X., Kandale, A., McGeary, R. P., Williams, S., Kobe, B., Schembri, M. A., Landsberg, M. J., Wu, B., Brück, T. B., Sieber, V., Boden, M., Rao, Z., Fleishman, S. J., Schenk, G., & Guddat, L. W. (2022). Enhancing the Thermal and Kinetic Stability of Ketol-Acid Reductoisomerase, a Central Catalyst of a Cell-Free Enzyme Cascade for the Manufacture of Platform Chemicals. Applied Biosciences, 1(2), 163-178. https://doi.org/10.3390/applbiosci1020011