
Forage Availability and Quality, and Feeding Behaviour of Indigenous Goats Grazing in a Mediterranean Silvopastoral System

 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
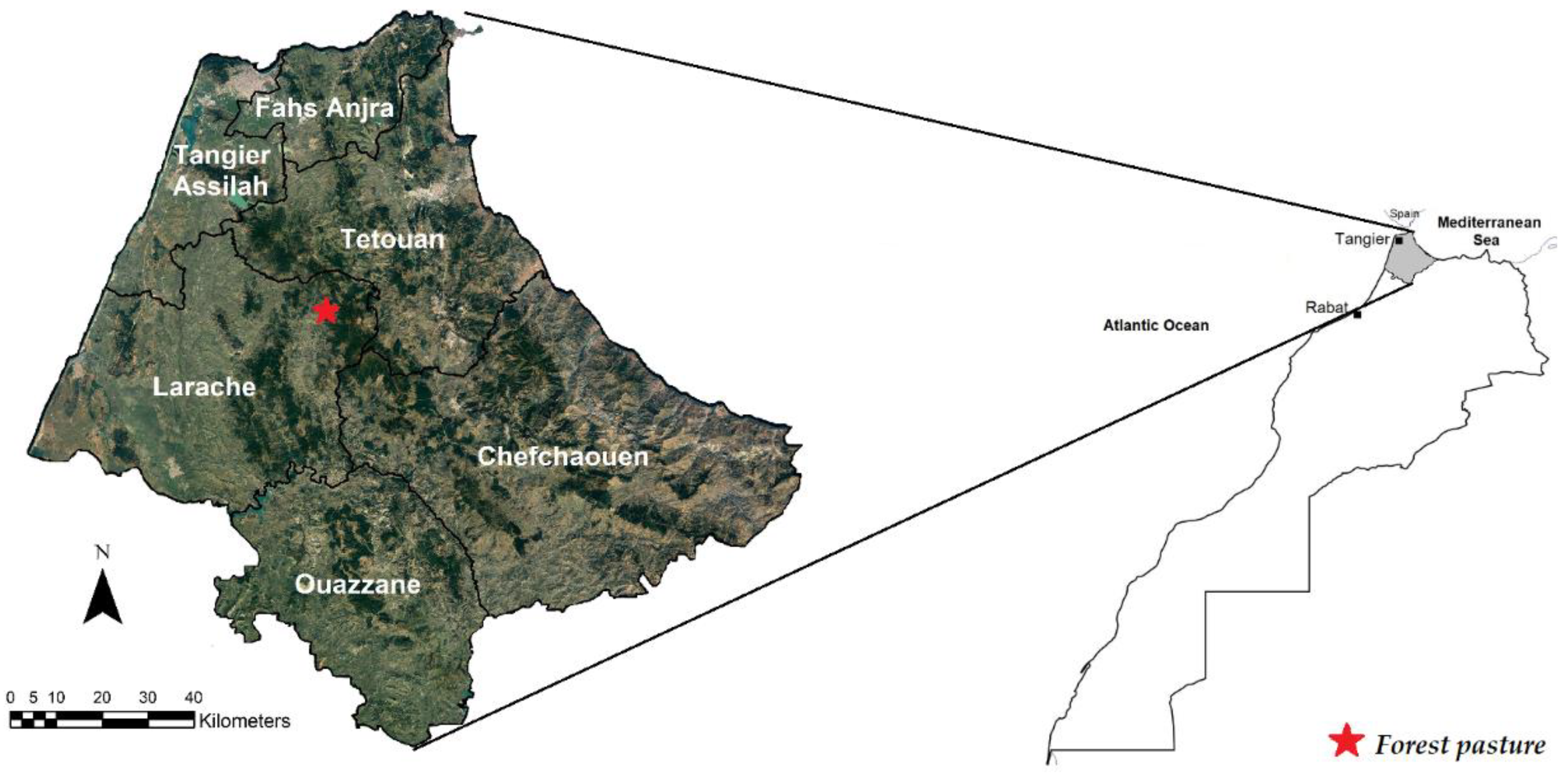
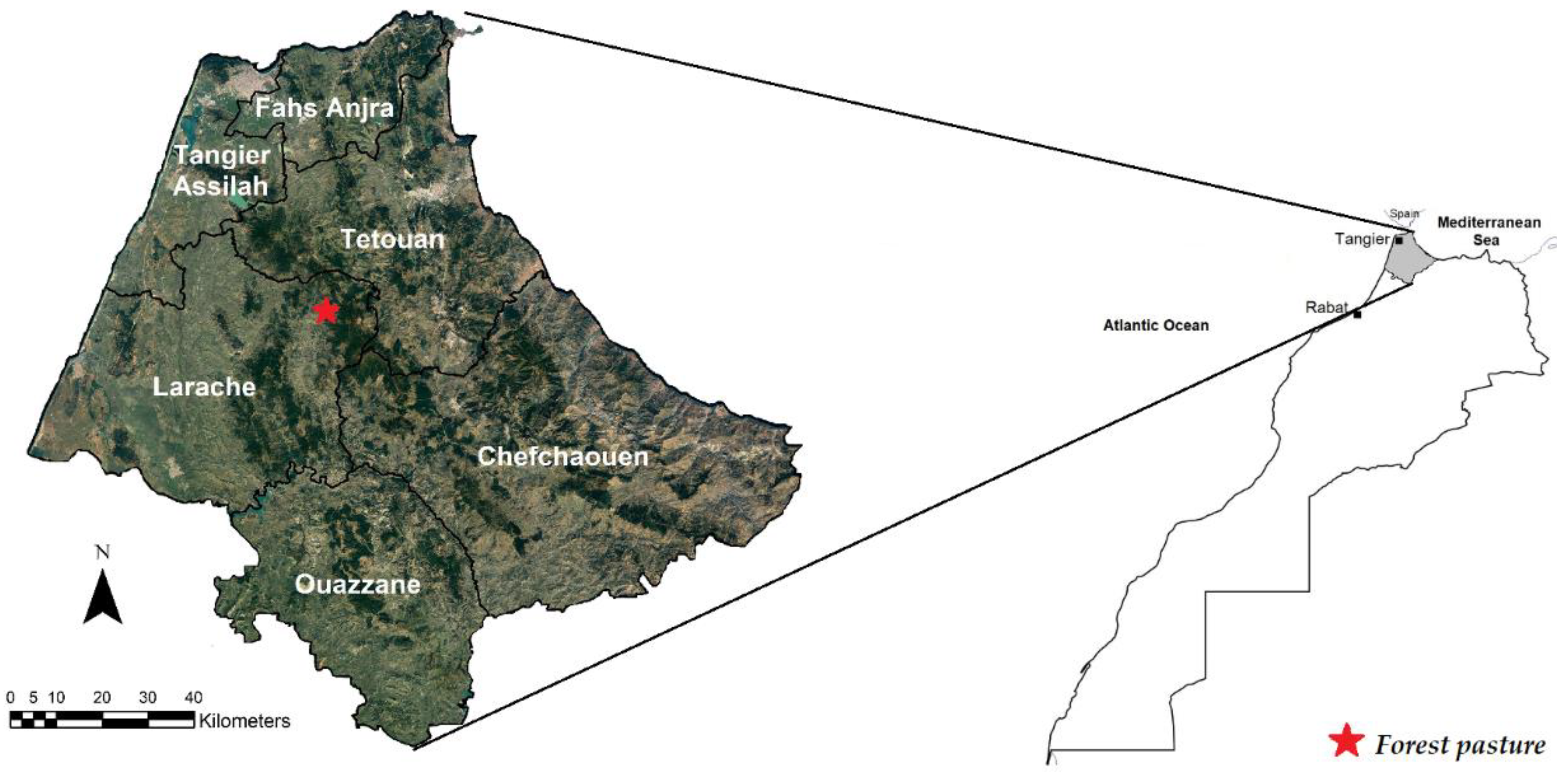
2.1. Experimental Forest Pasture
2.2. Experimental Animals and Feeding Behaviour
2.3. Forage Availability and Diet Selection
2.4. Forage Quality
2.4.1. Chemical Composition
2.4.2. Digestibility and Metabolizable Energy
2.5. Statistical Analysis
3. Results
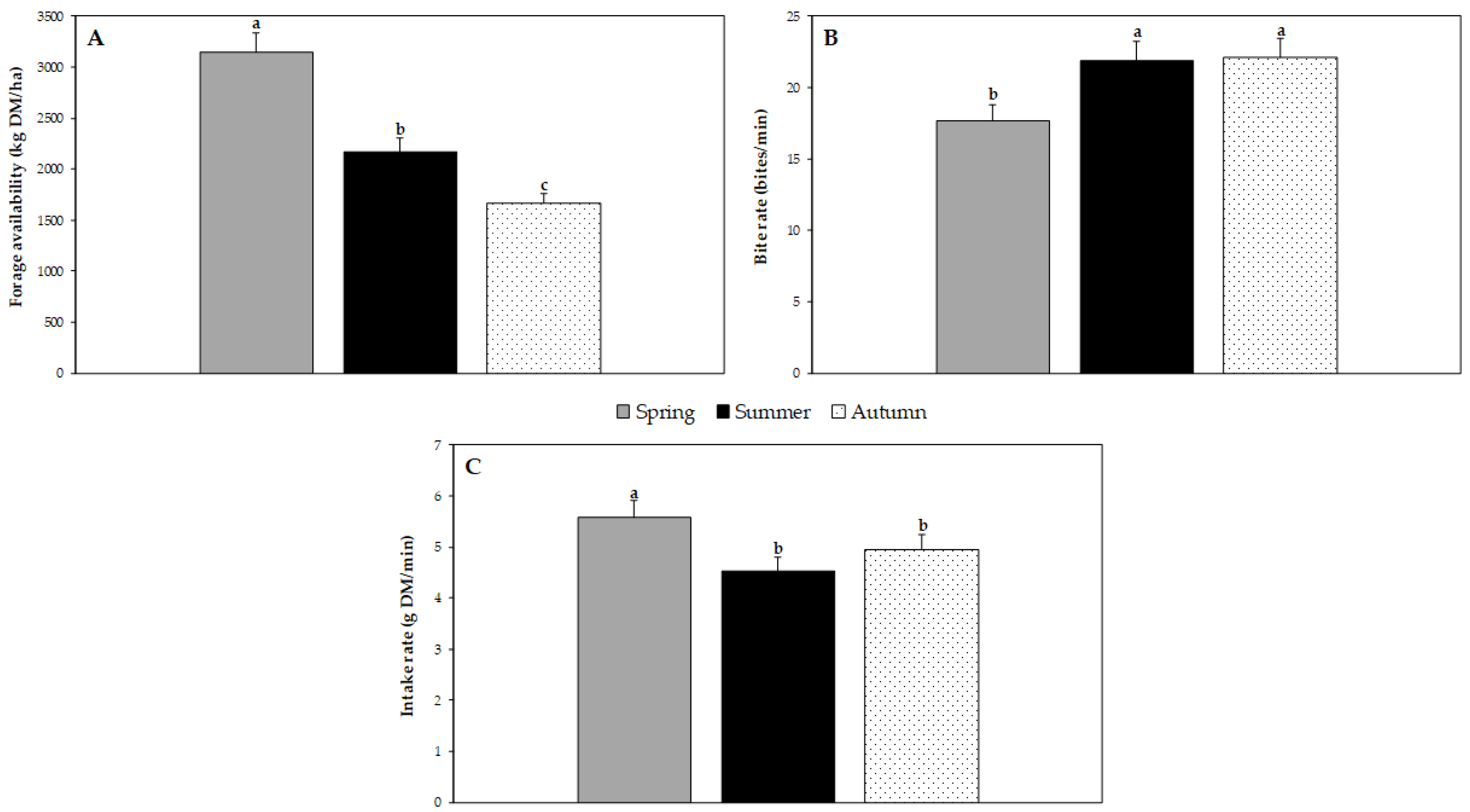
3.1. Forage Availability and Diet Selection
3.2. Feeding Behaviour
3.3. Forage Quality
4. Discussion
4.1. Forage Availability and Diet Selection
4.2. Feeding Behaviour
4.3. Forage Quality: Chemical Composition, Digestibility and Metabolizable Energy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chebli, Y.; El Otmani, S.; Elame, F.; Moula, N.; Chentouf, M.; Hornick, J.-L.; Cabaraux, J.-F. Silvopastoral System in Morocco: Focus on Their Importance, Strategic Functions, and Recent Changes in the Mediterranean Side. Sustainability 2021, 13, 10744. [Google Scholar] [CrossRef]
- Chentouf, M.; Boulanouar, B.; Bister, J.L. Elevage Caprin au Nord du Maroc; INRA-Editions Press: Rabat, Morocco, 2015; 168p. [Google Scholar]
- El Otmani, S.; Chebli, Y.; Taminiau, B.; Chentouf, M.; Hornick, J.-L.; Cabaraux, J.-F. Effect of Olive Cake and Cactus Cladodes Incorporation in Goat Kids’ Diet on the Rumen Microbial Community Profile and Meat Fatty Acid Composition. Biology 2021, 10, 1237. [Google Scholar] [CrossRef] [PubMed]
- Hilal, B.; Boujenane, I.; El Otmani, S.; Chentouf, M.; Piro, M. Genetic characterization of hamra goat population in two different locations of Morocco using microsatellite markers. Iran. J. Appl. Anim. Sci. 2016, 6, 901–907. [Google Scholar]
- El Otmani, S.; Chebli, Y.; Chentouf, M.; Hornick, J.L.; Caba-raux, J.F. Effects of Olive Cake and Cactus Cladodes as Alternative Feed Resources on Goat Milk Production and Quality. Agriculture 2021, 11, 3. [Google Scholar] [CrossRef]
- El Otmani, S.; Chebli, Y.; Hornick, J.L.; Cabaraux, J.F.; Chentouf, M. Growth performance, carcass characteristics and meat quality of male goat kids supplemented by alternative feed resources: Olive cake and cactus cladodes. Anim. Feed. Sci. Technol. 2021, 272, 114746. [Google Scholar] [CrossRef]
- Silanikove, N. The physiological basis of adaptation in goats to harsh environments. Small Rumin. Res. 2000, 35, 181–193. [Google Scholar] [CrossRef]
- Kababya, D.; Perevolotsky, A.; Bruckental, I.; Landau, S. Selection of diets by dual-purpose Mamber goats in Mediterranean woodland. J. Agric. Sci. 1998, 131, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Aharon, H.; Henkin, Z.; Ungar, E.; Kababya, D.; Baram, H.; Perevolotsky, A. Foraging behaviour of the newly introduced Boer goat breed in a Mediterranean woodland: A research observation. Small Rumin. Res. 2007, 69, 144–153. [Google Scholar] [CrossRef]
- Papachristou, T.; Dziba, L.; Villalba, J.J.; Provenza, F.D. Patterns of diet mixing by sheep offered foods varying in nutrients and plant secondary compounds. Appl. Anim. Behav. Sci. 2007, 108, 68–80. [Google Scholar] [CrossRef]
- Glasser, T.; Landau, S.; Ungar, E.; Perevolotsky, A.; Dvash, L.; Muklada, H.; Kababya, D.; Walker, J. Foraging selectivity of three goat breeds in a Mediterranean shrubland. Small Rumin. Res. 2012, 102, 7–12. [Google Scholar] [CrossRef]
- Koluman, N.; Boga, M.; Silanikove, N.; Gorgulu, M. Performance and eating behaviour of crossbred goats in Mediterranean climate of Turkey. Rev. Bras. Zootec. 2016, 45, 768–772. [Google Scholar] [CrossRef] [Green Version]
- Ventura-Cordero, J.; González-Pech, P.; Sandoval-Castro, C.A.; Torres-Acosta, J.F.J.; Tun-Garrido, J. Feed resource selection by Criollo goats browsing a tropical deciduous forest. Anim. Prod. Sci. 2017, 58, 2314–2320. [Google Scholar] [CrossRef]
- Manousidis, T.; Parissi, Z.; Kyriazopoulos, A.; Malesios, C.; Koutroubas, S.; Abas, Z. Relationships among nutritive value of selected forages, diet composition and milk quality in goats grazing in a Mediterranean woody rangeland. Livest. Sci. 2018, 218, 8–19. [Google Scholar] [CrossRef]
- Chebli, Y.; Chentouf, M.; Ozer, P.; Hornick, J.-L.; Cabaraux, J.-F. Forest and silvopastoral cover changes and its drivers in northern Morocco. Appl. Geogr. 2018, 101, 23–35. [Google Scholar] [CrossRef]
- Chebli, Y.; El Otmani, S.; Chentouf, M.; Hornick, J.-L.; Cabaraux, J.-F. Temporal Variations in Chemical Composition, In Vitro Digestibility, and Metabolizable Energy of Plant Species Browsed by Goats in Southern Mediterranean Forest Rangeland. Animals 2021, 11, 1441. [Google Scholar] [CrossRef]
- Manousidis, T.; Kyriazopoulos, A.; Parissi, Z.; Abraham, E.; Korakis, G.; Abas, Z. Grazing behavior, forage selection and diet composition of goats in a Mediterranean woody rangeland. Small Rumin. Res. 2016, 145, 142–153. [Google Scholar] [CrossRef]
- Agreil, C.; Meuret, M. An improved method for quantifying intake rate and ingestive behaviour of ruminants in diverse and variable habitats using direct observation. Small Rumin. Res. 2004, 54, 99–113. [Google Scholar] [CrossRef]
- Chebli, Y.; El Otmani, S.; Chentouf, M.; Hornick, J.-L.; Bindelle, J.; Cabaraux, J.-F.; Chebli, Y. Foraging Behavior of Goats Browsing in Southern Mediterranean Forest Rangeland. Animals. 2020, 10, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meuret, M.; Provenza, F.D. How French shepherds create meal sequences to stimulate intake and optimise use of forage diversity on rangeland. Anim. Prod. Sci. 2015, 55, 309–318. [Google Scholar] [CrossRef]
- González-Pech, P.G.; Torres-Acosta, J.F.D.J.; Sandoval-Castro, C.A.; Tun-Garrido, J.; de Jesús Torres-Acosta, J.F.; Sand-oval-Castro, C.A.; Tun-Garrido, J. Feeding behavior of sheep and goats in a deciduous tropical forest during the dry season: The same menu consumed differently. Small Rumin. Res. 2015, 133, 128–134. [Google Scholar] [CrossRef]
- Cook, C.W. Symposium on Nutrition of Forages and Pastures: Collecting Forage Samples Representative of Ingested Material of Grazing Animals for Nutritional Studies. J. Anim. Sci. 1964, 23, 265–270. [Google Scholar] [CrossRef]
- Barros, C.S.; Dittrich, J.R.; Rocha, C. Comportamento de caprinos em pasto de Brachiaria hibrida cv. mulato. Rev. Fac. Veterinária Zootec. Agron. 2007, 14, 187–206. [Google Scholar]
- Pontes, L.D.S.; Agreil, C.; Magda, D.; Gleizes, B.; Fritz, H. Feeding behaviour of sheep on shrubs in response to contrasting herbaceous cover in rangelands dominated by Cytisus scoparius L. Appl. Anim. Behav. Sci. 2010, 124, 35–44. [Google Scholar] [CrossRef]
- Chebli, Y.; Chentouf, M.; Hornick, J.L.; Cabaraux, J.F. Extensive goat production systems in northern Morocco: Production and use of pastoral resources. In Grassland Resources for Extensive Farming Systems in Marginal Lands: Major Drivers and Future scenarios; Porqueddu, A., Franca, C., Lombardi, A., Molle, G., Peratoner, G., Hopkins, G., Eds.; Wageningen Academic Publishers Press: Wageningen, The Netherlands, 2017; pp. 131–133. [Google Scholar]
- Lechowicz, M. The sampling characteristics of electivity indices. Oecologia 1982, 52, 22–30. [Google Scholar] [CrossRef]
- Ivlev, V.S. Experimental Ecology of the Feeding of Fishes; Yale University Press: New Haven, CT, USA, 1961; 302p. [Google Scholar]
- Barroso, F.G.; Alados, C.L.; Boza, J. Food selection by domestic goats in Mediterranean arid shrublands. J. Arid Environ. 1995, 31, 205–217. [Google Scholar] [CrossRef]
- Makkar, H.P.S.; Singh, B. Effect of drying conditions on tannin, fibre and lignin levels in mature oak (Quercus incana) leaves. J. Sci. Food Agric. 1991, 54, 323–328. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 16th ed.; Association of Analytical Chemists: Arlington, VA, USA, 1997. [Google Scholar]
- Mertens, D.R. Gravimetric determination of amylase-treated neutral detergent fiber in feeds with refluxing in beakers or crucibles: Collaborative study. J. AOAC Int. 2002, 85, 1217–1240. [Google Scholar]
- AOAC. Official Methods of Analysis, 15th ed.; Association of Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar]
- Robertson, J.B.; Van Soest, P.J. The Detergent System of Analysis; James, W.P.T., Theander, O., Eds.; The Analysis of dietary Fibre in Food; Marcel Dekker: New York, NY, USA, 1981; pp. 123–158. [Google Scholar]
- Porter, L.J.; Hrstich, L.N.; Chan, B.G. The conversion of procyanidins and prodelphinidins to cyanidin and delphinidin. Phytochemistry 1986, 25, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Mabjeesh, S.; Cohen, M.; Arieli, A. In Vitro Methods for Measuring the Dry Matter Digestibility of Ruminant Feedstuffs: Comparison of Methods and Inoculum Source. J. Dairy Sci. 2000, 83, 2289–2294. [Google Scholar] [CrossRef]
- Tassone, S.; Fortina, R.; Peiretti, P.G. In Vitro Techniques Using the Daisy II Incubator for the Assessment of Digestibility: A Review. Animals. 2020, 10, 775. [Google Scholar] [CrossRef]
- El Otmani, S.; Chentouf, M.; Hornick, J.L.; Cabaraux, J.F. Chemical composition and in vitro digestibility of alternative feed resources for ruminants in Mediterranean climates: Olive cake and cactus cladodes. J. Agric. Sci. 2019, 157, 260–271. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics, 2nd ed.; McGraw-Hill: New York, NY, USA, 1980. [Google Scholar]
- Littell, R.C.; Henry, P.R.; Ammerman, C.B. Statistical analysis of repeated measures data using SAS procedures. J. Anim. Sci. 1998, 76, 1216–1231. [Google Scholar] [CrossRef] [Green Version]
- Papachristou, T.; Platis, P.; Nastis, A. Foraging behaviour of cattle and goats in oak forest stands of varying coppicing age in Northern Greece. Small Rumin. Res. 2005, 59, 181–189. [Google Scholar] [CrossRef]
- Baraza, E.; Hódar, J.A.; Zamora, R. Consequences of plant–chemical diversity for domestic goat food preference in Mediterranean forests. Acta Oecol. 2009, 35, 117–127. [Google Scholar] [CrossRef]
- Papachristou, T. Foraging behaviour of goats and sheep on Mediterranean kermes oak shrublands. Small Rumin. Res. 1997, 24, 85–93. [Google Scholar] [CrossRef]
- Ferreira, L.M.M.; Celaya, R.; Benavides, R.; Jáuregui, B.M.; García, U.; Santos, A.S.; García, R.R.; Rodrigues, M.A.M.; Osoro, K. Foraging behaviour of domestic herbivore species grazing on heathlands associated with improved pasture areas. Livest. Sci. 2013, 155, 373–383. [Google Scholar] [CrossRef]
- Dziba, L.; Scogings, P.; Gordon, I.; Raats, J. Effects of season and breed on browse species intake rates and diet selection by goats in the False Thornveld of the Eastern Cape, South Africa. Small Rumin. Res. 2003, 47, 17–30. [Google Scholar] [CrossRef]
- Harris, S.E. Cyprus as a degraded landscape or resilient environment in the wake of colonial intrusion. Proc. Natl. Acad. Sci. USA 2012, 109, 3670–3675. [Google Scholar] [CrossRef] [Green Version]
- Lovreglio, R.; Sahar, O.; Leone, V. Goat grazing as a wildfire prevention tool: A basic review. IForest—Biogeosci. For. 2014, 7, 260–268. [Google Scholar] [CrossRef] [Green Version]
- Egea, A.; Allegretti, L.; Lama, S.P.; Grilli, D.; Sartor, C.; Fucili, M.; Guevara, J.; Passera, C. Selective behavior of Creole goats in response to the functional heterogeneity of native forage species in the central Monte desert, Argentina. Small Rumin. Res. 2014, 120, 90–99. [Google Scholar] [CrossRef]
- Fomum, S.W.; Scogings, P.F.; Dziba, L.; Nsahlai, I.V. Seasonal variations in diet selection of Nguni goats: Effects of physical and chemical traits of browse. Afr. J. Range Forage Sci. 2015, 32, 193–201. [Google Scholar] [CrossRef]
- Sanon, H.; Kaboré-Zoungrana, C.; Ledin, I. Behaviour of goats, sheep and cattle and their selection of browse species on natural pasture in a Sahelian area. Small Rumin. Res. 2007, 67, 64–74. [Google Scholar] [CrossRef]
- Egea, A.; Allegretti, L.; Lama, S.P.; Grilli, D.; Fucili, M.; Guevara, J.; Villalba, J. Diet mixing and condensed tannins help explain foraging preferences by Creole goats facing the physical and chemical diversity of native woody plants in the central Monte desert (Argentina). Anim. Feed. Sci. Technol. 2016, 215, 47–57. [Google Scholar] [CrossRef]
- Mero, R.N.; Udén, P. Promising tropical grasses and legumes as feed resources in central Tanzania. VI. Nitrogen balance in growing bulls consuming tropical herbaceous forage legumes. Anim. Feed. Sci. Technol. 1998, 72, 387–396. [Google Scholar] [CrossRef]
- Hassen, A.; Rethman, N.; Van Niekerk, W.; Tjelele, T. Influence of season/year and species on chemical composition and in vitro digestibility of five Indigofera accessions. Anim. Feed. Sci. Technol. 2007, 136, 312–322. [Google Scholar] [CrossRef]
- Safari, J.; Mushi, D.; Kifaro, G.; Mtenga, L.; Eik, L. Seasonal variation in chemical composition of native forages, grazing behaviour and some blood metabolites of Small East African goats in a semi-arid area of Tanzania. Anim. Feed. Sci. Technol. 2011, 164, 62–70. [Google Scholar] [CrossRef]
- Rawnsley, R.; Donaghy, D.J.; Fulkerson, W.J.; Lane, P.A. Changes in the physiology and feed quality of cocksfoot (Dactylis glomerata L.) during regrowth. Grass Forage Sci. 2002, 57, 203–211. [Google Scholar] [CrossRef] [Green Version]
- Minson, D.J. Forage in Ruminant Nutrition; Academic Press: New York, NY, USA, 1990. [Google Scholar]
- Min, B.; Barry, T.; Attwood, G.; McNabb, W. The effect of condensed tannins on the nutrition and health of ruminants fed fresh temperate forages: A review. Anim. Feed Sci. Technol. 2003, 106, 3–19. [Google Scholar] [CrossRef]
- Waghorn, G. Beneficial and detrimental effects of dietary condensed tannins for sustainable sheep and goat production—Progress and challenges. Anim. Feed Sci. Technol. 2008, 147, 116–139. [Google Scholar] [CrossRef]
- Dove, H. Ingestive behaviour, diet selection, and feed intake. In Goat Science and Production; Solaiman, S.G., Ed.; Wiley-Blackwell: Ames, IA, USA, 2010; pp. 179–192. [Google Scholar]
- Mkhize, N.R.; Scogings, P.F.; Nsahlai, I.V.; Dziba, L. Diet selection of goats depends on season: Roles of plant physical and chemical traits. Afr. J. Range Forage Sci. 2014, 31, 209–214. [Google Scholar] [CrossRef]
- Iason, G.R.; Villalba, J.J. Behavioral Strategies of Mammal Herbivores Against Plant Secondary Metabolites: The Avoidance–Tolerance Continuum. J. Chem. Ecol. 2006, 32, 1115–1132. [Google Scholar] [CrossRef]
- Rogosic, J.; Pfister, J.; Provenza, F.; Grbesa, D. Sheep and goat preference for and nutritional value of Mediterranean maquis shrubs. Small Rumin. Res. 2006, 64, 169–179. [Google Scholar] [CrossRef]
- Molina-Alcaide, E.; Yáñez-Ruiz, D. Potential use of olive by-products in ruminant feeding: A review. Anim. Feed Sci. Technol. 2008, 147, 247–264. [Google Scholar] [CrossRef]
- Paton, D. Elaboration of a multi-variate model for the determination of the metabolizable energy of Mediterranean bushes based on chemical parameters. J. Arid Environ. 2003, 53, 271–280. [Google Scholar] [CrossRef]
- Kozloski, G.; Perottoni, J.; Sanchez, L. Influence of regrowth age on the nutritive value of dwarf elephant grass hay (Pennisetum purpureum Schum. cv. Mott) consumed by lambs. Anim. Feed. Sci. Technol. 2005, 119, 1–11. [Google Scholar] [CrossRef]
- Van Soest, P.J. Nutritional Ecology of the Ruminant, 2nd ed.; Cornell University Press: Ithaca, NY, USA, 1994; p. 476. [Google Scholar] [CrossRef]
- Tainton, N. Veld Management in South Africa; University of Natal Press: Pietermaritzburg, South Africa, 1999; p. 472. [Google Scholar]
- Papachristou, T.G.; Nastis, A.S.; Mathur, R.; Hutchings, M.R. Effect of physical and chemical plant defences on herbivory: Implications for Mediterranean shrubland management. Basic Appl. Ecol. 2003, 4, 395–403. [Google Scholar] [CrossRef]
- Emmans, G.C. Diet selection by animals: Theory and experimental design. Proc. Nutr. Soc. 1991, 50, 59–64. [Google Scholar] [CrossRef] [Green Version]
- De Rosa, G.; Moio, L.; Napolitano, F.; Grasso, F.; Gubitosi, L.; Bordi, A. Influence of Flavor on Goat Feeding Preferences. J. Chem. Ecol. 2002, 28, 269–281. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Availability (%) | Selectivity Index | Bite Mass (g DM/Bite) | Total Bites (%) | Diet Composition (%) | |
|---|---|---|---|---|---|
| Spring | |||||
| Arbutus unedo | 31.4 c | −0.983 n | 0.119 j | 0.619 mno | 0.234 lmno |
| Cistus crispus | 12.9 de | 0.549 c | 0.340 a | 26.1 a | 25.9 a |
| Cistus monspeliensis | 13.9 de | 0.255 d | 0.323 b | 23.6 b | 23.9 b |
| Cistus salviifolius | 18.3 d | 0.045 e | 0.349 a | 21.8 cd | 24.3 b |
| Calicotome villosa | 1.55 ij | −0.919 lmn | 0.110 jk | 0.185 no | 0.064 no |
| Erica arborea | 16.6 e | −0.784 jkl | 0.222 ef | 4.43 l | 2.74 i |
| Lavandula stoechas | 3.45 ghi | 0.757 b | 0.319 b | 22.2 cd | 22.5 b |
| Myrtus communis | 0.033 kl | 0.650 b | 0.080 mn | 0.319 no | 0.078 no |
| Pistacia lentiscus | 1.72 ijk | −0.822 klmn | 0.091 lm | 0.505 no | 0.146 mno |
| Phillyrea media | 0.034 kl | 0.672 b | 0.070 n | 0.255 no | 0.057 no |
| Rubus ulmifolius | 0.010 l | −0.984 n | 0.050 o | 0.006 o | 0.001 o |
| SEM | 0.057 | 0.025 | 0.004 | 0.402 | 0.043 |
| Summer | |||||
| Arbutus unedo | 12.02 gh | −0.470 h | 0.220 ef | 12.8 i | 14.6 e |
| Cistus crispus | 10.5 ef | −0.904 lmn | 0.110 jk | 1.29 mno | 0.75 kl |
| Cistus monspeliensis | 26.8 c | −0.957 mn | 0.101 kl | 1.01 mno | 0.529 klm |
| Cistus salviifolius | 42.7 a | −0.961 mn | 0.110 jk | 1.32 mn | 0.760 k |
| Calicotome villosa | 2.11 hi | 0.618 bc | 0.170 gh | 9.33 j | 8.25 g |
| Erica arborea | 1.71 ijk | −0.139 de | 0.179 g | 16.7 gh | 15.7 e |
| Lavandula stoechas | 3.27 ghi | −0.810 jklm | 0.100 kl | 1.28 mno | 0.673 kl |
| Myrtus communis | 0.338 jkl | 0.974 a | 0.260 c | 17.5 g | 23.6 c |
| Pistacia lentiscus | 0.197 jkl | 0.734 b | 0.242 d | 18.7 f | 23.4 c |
| Phillyrea media | 0.040 jkl | 0.969 a | 0.147 i | 12.7 i | 7.26 g |
| Rubus ulmifolius | 0.329 jkl | 0.740 b | 0.120 j | 7.32 k | 4.54 h |
| SEM | 0.075 | 0.029 | 0.002 | 2.57 | 0.031 |
| Autumn | |||||
| Arbutus unedo | 30.7 bc | −0.200 fg | 0.215 f | 17.3 e | 18.9 d |
| Cistus crispus | 2.77 ghi | −0.890 lmn | 0.093 lm | 1.64 m | 0.788 kl |
| Cistus monspeliensis | 5.89 fg | −0.962 lm | 0.090 lm | 1.01 mno | 0.470 klmn |
| Cistus salviifolius | 12.5 de | −0.971 mn | 0.105 jkl | 1.09 mno | 0.552 klm |
| Calicotome villosa | 0.218 jkl | 0.600 bc | 0.165 gh | 0.902 mno | 0.725 klm |
| Erica arborea | 41.8 ab | −0.265 g | 0.167 gh | 20.3 bc | 18.0 d |
| Lavandula stoechas | 2.91 ghi | −0.518 hi | 0.090 lm | 3.66 l | 1.68 j |
| Myrtus communis | 0.150 jkl | 0.978 a | 0.242 d | 19.3 d | 23.5 c |
| Pistacia lentiscus | 3.00 ghi | 0.762 b | 0.233 de | 20.1 cd | 23.3 c |
| Phillyrea media | 0.078 jkl | 0.947 a | 0.163 h | 13.6 h | 11.2 f |
| Rubus ulmifolius | 0.037 kl | 0.093 e | 0.144 i | 1.13 nm | 0.831 kl |
| SEM | 0.069 | 0.028 | 0.002 | 0.328 | 0.035 |
| p-Value | |||||
| Season | 0.005 | <0.001 | <0.001 | <0.001 | 0.002 |
| Plant species | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Season × Plant species | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Availability (%) | Selectivity Index | Bite Mass (g DM/bite) | Total Bites (%) | Diet Composition (%) | |
|---|---|---|---|---|---|
| Spring | |||||
| Olea europaea | 9.48 c | −0.655 de | 0.071 g | 13.1 d | 23.6 b |
| Quercus coccifera | 20.9 b | −0.929 f | 0.071 g | 2.09 f | 3.52 e |
| Quercus ilex | 28.6 b | −0.793 ef | 0.060 h | 10.1 e | 16.0 cd |
| Quercus suber | 40.9 a | −0.464 d | 0.029 i | 74.8 a | 56.9 a |
| SEM | 4.95 | 0.027 | 0.002 | 2.66 | 0.271 |
| Summer | |||||
| Olea europaea | 9.46 c | 0.979 a | 0.191 c | 26.6 c | 20.6 b |
| Quercus coccifera | 19.1 b | 0.868 ab | 0.139 f | 10.8 e | 18.2 c |
| Quercus ilex | 25.2 b | 0.619 c | 0.139 f | 16.9 d | 9.60 d |
| Quercus suber | 46.2 a | 0.906 ab | 0.282 a | 45.6 b | 51.6 a |
| SEM | 4.00 | 0.013 | 0.004 | 0.928 | 1.01 |
| Autumn | |||||
| Olea europaea | 5.00 c | 0.714 bc | 0.205 b | 15.6 d | 14.9 cd |
| Quercus coccifera | 24.3 b | 0.572 c | 0.171 d | 18.6 cd | 15.5 cd |
| Quercus ilex | 24.5 b | 0.652 c | 0.153 e | 23.9 c | 18.2 c |
| Quercus suber | 46.2 a | 0.941 a | 0.289 a | 41.8 b | 51.3 a |
| SEM | 7.83 | 0.029 | 0.003 | 0.845 | 0.109 |
| p-Value | |||||
| Season | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Plant species | 0.008 | <0.001 | <0.001 | <0.001 | <0.001 |
| Season × Plant species | 0.956 | <0.001 | <0.001 | <0.001 | <0.001 |
| Availability (%) | Selectivity Index | Bite Mass (g DM/bite) | Total Bites (%) | Diet Composition (%) | |
|---|---|---|---|---|---|
| Spring | |||||
| Herbaceous | 8.49 c | 0.079 b | 0.361 a | 8.10 d | 10.7 d |
| Shrubs | 90.10 b | −0.383 d | 0.188 c | 88.9 a | 88.9 a |
| Trees | 1.41 de | −0.710 e | 0.058 f | 2.99 e | 0.37 f |
| SEM | 3.74 | 0.018 | 0.007 | 2.70 | 2.78 |
| Summer | |||||
| Herbaceous | 8.91 c | −0.466 d | 0.110 e | 3.12 e | 1.66 e |
| Shrubs | 89.02 b | −0.019 c | 0.160 d | 67.9 b | 63.6 b |
| Trees | 2.07 d | 0.843 a | 0.188 c | 28.9 c | 34.7 c |
| SEM | 3.66 | 0.038 | 0.002 | 1.84 | 1.74 |
| Autumn | |||||
| Herbaceous | 2.56 d | −0.037 c | 0.105 e | 5.56 de | 1.86 e |
| Shrubs | 96.84 a | −0.165 d | 0.155 d | 69.1 b | 67.7 b |
| Trees | 0.600 e | 0.720 a | 0.204 b | 25.4 c | 30.4 c |
| SEM | 4.12 | 0.029 | 0.003 | 0.467 | 1.87 |
| p-Value | |||||
| Season | 0.335 | <0.001 | <0.001 | <0.001 | <0.001 |
| Plant groups | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Season × Plant groups | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| DM | OM | CP | EE | NDF | ADF | ADL | CT | IVOMD | ME | |
|---|---|---|---|---|---|---|---|---|---|---|
| Spring | ||||||||||
| Herbaceous | 472 bc | 915 b | 169 a | 20.5 a | 529 ab | 344 ab | 61.3 c | 2.67 b | 823 a | 12.2 a |
| Shrubs | 466 bc | 940 ab | 109 b | 45.3 a | 402 c | 274 b | 145 ab | 65.4 ab | 575 d | 8.10 d |
| Trees | 548 ab | 959 a | 95.2 bc | 39.5 a | 511 ab | 349 ab | 164 a | 42.6 ab | 547 de | 7.86 d |
| SEM | 15.6 | 4.19 | 4.41 | 4.27 | 11.3 | 8.99 | 6.92 | 8.26 | 14.0 | 0.23 |
| Summer | ||||||||||
| Herbaceous | 628 a | 878 c | 86.6 bc | 23.1 a | 581 a | 382 a | 79.9 bc | 3.70 b | 652 c | 9.60 c |
| Shrubs | 557 ab | 947 ab | 89.7 bc | 47.8 a | 453 bc | 304 ab | 187 a | 86.6 a | 460 f | 6.29 e |
| Trees | 597 a | 947 ab | 77.1 bc | 47.9 a | 508 ab | 340 ab | 162 a | 55.7 ab | 459 f | 6.52 e |
| SEM | 13.7 | 5.24 | 4.54 | 4.69 | 12.9 | 10.3 | 10.8 | 9.14 | 10.4 | 0.18 |
| Autumn | ||||||||||
| Herbaceous | 537 abc | 847 d | 66.4 c | 21.4 a | 455 bc | 281 b | 64.2 c | 3.30 b | 724 b | 10.8 b |
| Shrubs | 429 c | 942 ab | 90.3 bc | 37.2 a | 449 bc | 311 ab | 179 a | 78.6 a | 458 f | 6.33 e |
| Trees | 563 ab | 951 a | 78.1 bc | 36.2 a | 489 b | 338 ab | 164 a | 49.7 ab | 496 ef | 7.15 de |
| SEM | 15.3 | 4.62 | 4.79 | 3.18 | 10.3 | 10.3 | 9.72 | 8.51 | 11.4 | 0.20 |
| p-Value | ||||||||||
| Season | 0.003 | 0.020 | <0.001 | 0.669 | 0.089 | 0.295 | 0.582 | 0.809 | <0.001 | <0.001 |
| Plant groups | <0.001 | <0.001 | 0.039 | 0.093 | <0.001 | 0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Season × Plant groups | 0.230 | 0.036 | 0.023 | 0.983 | 0.065 | 0.150 | 0.577 | 0.993 | 0.119 | 0.162 |
| DM | OM | CP | EE | NDF | ADF | ADL | CT | IVOMD | ME | |
|---|---|---|---|---|---|---|---|---|---|---|
| Spring | ||||||||||
| Arbutus unedo | 572 efg | 961 abcde | 77.8 jklmn | 71.8 b | 348 no | 248 klmnop | 112 hij | 92.5 hij | 638 b | 9.14 b |
| Cistus crispus | 415 pq | 954 abcde | 112 cde | 15.9 k | 303 p | 267 jklm | 107 hijk | 16.4 pq | 696 a | 9.92 a |
| Cistus monspeliensis | 583 def | 900 hi | 97.7 efgh | 55.9 c | 372 mn | 217 pqr | 205 de | 43.7 no | 593 cd | 8.38 cd |
| Cistus salviifolius | 489 m | 878 ij | 105 def | 23.2 ijk | 412 ijkl | 252 klmno | 150 fg | 24.3 op | 620 bc | 8.66 bc |
| Calicotome villosa | 227 t | 918 efghi | 188 a | 28.8 ghi | 529 bc | 429 bc | 104 hijk | 3.42 q | 553 de | 7.90 def |
| Erica arborea | 567 efgh | 979 ab | 82.3 hijkl | 96.1 a | 437 ghi | 337 fg | 244 c | 114 efg | 499 fghi | 6.74 ghij |
| Lavandula stoechas | 293 s | 947 abcdef | 105 def | 90.9 a | 413 ijkl | 256 klmno | 172 f | 2.73 q | 717 a | 10.4 a |
| Myrtus communis | 556 fghi | 956 abcde | 92.2 fghij | 44.3 de | 382 klmn | 229 pqr | 119 hij | 103 ghi | 506 fgh | 7.20 efgh |
| Pistacia lentiscus | 542 ghij | 956 abcde | 96.8 efghi | 28.6 ghi | 451 fgh | 293 hij | 124 ghi | 173 b | 533 ef | 7.42 efg |
| Phillyrea media | 522 jkl | 961 abcde | 110 de | 25.1 hijk | 398 jklm | 271 jkl | 177 ef | 3.23 q | 527 efg | 7.46 efg |
| Rubus ulmifolius | 363 r | 925 defgh | 135 b | 17.7 jk | 378 lmn | 214 qr | 79.9 klm | 144 c | 441 lmno | 5.81 klmn |
| SEM | 20.9 | 5.43 | 5.19 | 4.94 | 10.1 | 10.6 | 8.52 | 10.5 | 14.7 | 0.24 |
| Summer | ||||||||||
| Arbutus unedo | 662 ab | 967 abcd | 54.8 op | 93.6 a | 464 efg | 328 fg | 246 c | 133 cde | 556 de | 7.75 def |
| Cistus crispus | 537 hij | 952 abcdef | 62.8 nop | 24.9 hijk | 247 q | 236 mnopq | 216 cd | 69.5 klm | 443 klmno | 5.88 klmn |
| Cistus monspeliensis | 691 a | 943 bcdefg | 82.8 ghijkl | 90.4 a | 493 cde | 266 jklm | 171 f | 64.4 lm | 408 op | 5.33 mn |
| Cistus salviifolius | 440 op | 849 j | 81.5 hijklm | 51.9 cd | 502 bcd | 352 ef | 245 c | 85.8 ijk | 443 klmno | 6.03 jklm |
| Calicotome villosa | 496 ml | 987 a | 190 a | 25.8 hijk | 616 a | 464 a | 112 hij | 2.47 q | 470 hijkl | 6.29 ijkl |
| Erica arborea | 656 b | 969 abc | 53.3 p | 91.7 a | 536 b | 403 cd | 373 a | 119 defg | 360 q | 4.41 p |
| Lavandula stoechas | 476 mn | 944 bcdefg | 87.4 ghijk | 36.2 efg | 468 defg | 313 gh | 226 cd | 3.51 q | 501 fghi | 6.84 ghi |
| Myrtus communis | 535 ijk | 946 abcdef | 73.4 klmn | 39.7 ef | 366 mno | 233 nopq | 95.4 ijkl | 129 cdef | 566 de | 7.93 cde |
| Pistacia lentiscus | 619 c | 948 abcdef | 99.5 defg | 23.8 hijk | 486 def | 263 jklmn | 178 ef | 206 a | 454 jklmn | 6.53 hijk |
| Phillyrea media | 612 cd | 966 abcd | 85.7 ghijk | 27.0 ghij | 434 ghij | 268 jklm | 127 gh | 2.57 q | 417 nop | 5.80 klmn |
| Rubus ulmifolius | 405 q | 944 bcdefg | 115 cd | 20.8 ijk | 372 mn | 218 pqr | 72.1 lm | 138 cd | 438 lmno | 6.38 ijk |
| SEM | 16.4 | 6.42 | 6.45 | 5.03 | 16.6 | 13.2 | 14.7 | 11.2 | 10.5 | 0.17 |
| Autumn | ||||||||||
| Arbutus unedo | 447 no | 952 abcdef | 64.6 mnop | 72.1 b | 495 cde | 380 de | 232 cd | 124 cdef | 395 pq | 5.22 no |
| Cistus crispus | 340 r | 927 cdefgh | 74.7 klmn | 17.4 jk | 375 mn | 255 klmno | 173 f | 58.1 mn | 510 fgh | 7.16 fgh |
| Cistus monspeliensis | 369 r | 936 cdefgh | 66.7 lmnop | 86.0 a | 436 ghi | 307 ghi | 228 cd | 79.3 jkl | 508 fgh | 6.92 ghi |
| Cistus salviifolius | 348 r | 903 ghi | 67.8 lmnop | 39.6 ef | 485 def | 293 hij | 230 cd | 51.5 mn | 464 ijklm | 6.34 ijk |
| Calicotome villosa | 295 s | 955 abcde | 197 a | 24.8 hijk | 594 a | 446 ab | 126 gh | 2.09 q | 359 q | 4.57 op |
| Erica arborea | 506 klm | 968 abc | 68.0 lmnop | 42.9 def | 584 a | 459 ab | 323 b | 117 efg | 426 mnop | 5.56 lmn |
| Lavandula stoechas | 405 q | 912 fghi | 70.5 klmno | 33.7 fgh | 436 ghi | 294 hij | 211 d | 1.93 q | 526 efg | 7.40 efg |
| Myrtus communis | 434 opq | 946 abcdefg | 80.5 ijklm | 22.5 ijk | 334 op | 243 lmnopq | 92.1 jkl | 111 fgh | 492 ghij | 7.33 efg |
| Pistacia lentiscus | 591 cde | 952 abcdef | 98.4 defgh | 22.5 ijk | 419 hijk | 279 ijk | 168 f | 195 a | 483 hijk | 6.75 ghij |
| Phillyrea media | 575 ef | 963 abcd | 78.3 jklmn | 28.5 ghi | 425 hij | 266 jklm | 123 ghi | 2.93 q | 427 mnop | 5.93 klmn |
| Rubus ulmifolius | 411 pq | 944 bcdefg | 127 bc | 19.7 ijk | 361 no | 199 r | 61.2 m | 122 defg | 451 jklmn | 6.47 hijk |
| SEM | 16.1 | 3.80 | 6.77 | 3.80 | 14.3 | 14.0 | 12.8 | 10.5 | 9.01 | 0.16 |
| p-Value | ||||||||||
| Season | <0.001 | 0.071 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Plant species | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Season × Plant species | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| DM | OM | CP | EE | NDF | ADF | ADL | CT | IVOMD | ME | |
|---|---|---|---|---|---|---|---|---|---|---|
| Spring | ||||||||||
| Olea europaea | 466 g | 958 a | 78.7 cde | 97.4 b | 419 g | 317 d | 161 cd | 4.20 f | 511 c | 7.04 ef |
| Quercus coccifera | 567 f | 957 a | 105 a | 18.3 e | 489 f | 343 cd | 132 f | 18.4 e | 613 a | 8.93 a |
| Quercus ilex | 576 f | 952 a | 109 a | 18.7 e | 548 bc | 349 c | 191 a | 29.3 c | 515 c | 7.33 de |
| Quercus suber | 584 ef | 970 a | 88.4 bc | 23.4 de | 589 a | 386 ab | 174 abc | 119 b | 549 b | 8.13 b |
| SEM | 14.5 | 4.42 | 3.91 | 10.1 | 19.6 | 7.64 | 6.64 | 13.5 | 12.6 | 0.227 |
| Summer | ||||||||||
| Olea europaea | 436 h | 914 b | 91.6 b | 121 a | 444 g | 255 f | 150 de | 2.57 f | 525 bc | 8.08 b |
| Quercus coccifera | 693 a | 963 a | 66.6 e | 25.4 d | 552 b | 389 ab | 172 bc | 21.4 de | 429 ef | 5.74 h |
| Quercus ilex | 611 cd | 951 a | 71.2 de | 19.3 e | 532 bcd | 353 c | 182 ab | 61.2 c | 421 f | 5.83 h |
| Quercus suber | 648 b | 959 a | 79.0 cde | 27.0 d | 502 ef | 363 bc | 144 ef | 138 a | 460 de | 6.43 g |
| SEM | 29.4 | 5.97 | 2.93 | 2.93 | 12.5 | 15.6 | 4.98 | 15.6 | 12.6 | 0.285 |
| Autumn | ||||||||||
| Olea europaea | 422 h | 916 b | 81.1 bcd | 77.2 c | 443 g | 286 e | 179 abc | 2.77 f | 521 bc | 7.81 bc |
| Quercus coccifera | 629 bc | 967 a | 71.0 de | 23.0 de | 522 cde | 401 a | 175 abc | 16.8 e | 463 d | 6.50 g |
| Quercus ilex | 600 de | 963 a | 70.8 de | 18.8 e | 503 def | 347 c | 170 bc | 54.6 c | 477 d | 6.73 fg |
| Quercus suber | 602 de | 957 a | 89.4 bc | 25.9 d | 488 f | 317 d | 134 ef | 125 b | 525 bc | 7.58 cd |
| SEM | 24.9 | 6.49 | 2.58 | 7.21 | 8.78 | 13.0 | 5.63 | 14.3 | 8.40 | 0.168 |
| p-Value | ||||||||||
| Season | <0.001 | 0.023 | <0.001 | <0.001 | <0.001 | 0.016 | 0.507 | <0.001 | <0.001 | <0.001 |
| Plant species | <0.001 | <0.001 | 0.144 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Season × Plant species | <0.001 | 0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| EE | NDF | ADF | ADL | CP | CT | IVDMD | IVOMD | ||
|---|---|---|---|---|---|---|---|---|---|
| Intake rate | Spring | 0.029 | −0.42 *** | −0.39 *** | −0.10 ** | 0.11 ** | −0.38 *** | 0.52 *** | 0.57 *** |
| Summer | 0.036 | 0.04 | 0.02 | −0.02 | 0.21 ** | 0.58 *** | 0.09 ** | 0.07 * | |
| Autumn | −0.06 | −0.03 | 0.09 | 0.06 | 0.08 * | 0.63 *** | 0.08 ** | 0.11 | |
| Selection | Spring | 0.09 ** | −0.25 *** | −0.27 *** | −0.15 ** | 0.19 ** | −0.36 *** | 0.51 *** | 0.53 *** |
| Summer | −0.22 *** | 0.11 *** | −0.01 | −0.45 *** | 0.26 *** | 0.10 *** | 0.07 * | −0.03 | |
| Autumn | −0.24 *** | −0.120 *** | −0.11 *** | −0.36 *** | 0.07 * | 0.13 *** | 0.17 *** | 0.09 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chebli, Y.; El Otmani, S.; Hornick, J.-L.; Keli, A.; Bindelle, J.; Cabaraux, J.-F.; Chentouf, M. Forage Availability and Quality, and Feeding Behaviour of Indigenous Goats Grazing in a Mediterranean Silvopastoral System. Ruminants 2022, 2, 74-89. https://doi.org/10.3390/ruminants2010004
Chebli Y, El Otmani S, Hornick J-L, Keli A, Bindelle J, Cabaraux J-F, Chentouf M. Forage Availability and Quality, and Feeding Behaviour of Indigenous Goats Grazing in a Mediterranean Silvopastoral System. Ruminants. 2022; 2(1):74-89. https://doi.org/10.3390/ruminants2010004
Chicago/Turabian StyleChebli, Youssef, Samira El Otmani, Jean-Luc Hornick, Abdelhafid Keli, Jérôme Bindelle, Jean-François Cabaraux, and Mouad Chentouf. 2022. "Forage Availability and Quality, and Feeding Behaviour of Indigenous Goats Grazing in a Mediterranean Silvopastoral System" Ruminants 2, no. 1: 74-89. https://doi.org/10.3390/ruminants2010004
APA StyleChebli, Y., El Otmani, S., Hornick, J.-L., Keli, A., Bindelle, J., Cabaraux, J.-F., & Chentouf, M. (2022). Forage Availability and Quality, and Feeding Behaviour of Indigenous Goats Grazing in a Mediterranean Silvopastoral System. Ruminants, 2(1), 74-89. https://doi.org/10.3390/ruminants2010004