

Assessment of Different Lactic Acid Bacteria Isolated from Agro-Industrial Residues: First Report of the Potential Role of Weissella soli for Lactic Acid Production from Milk Whey



 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Lactic Acid Bacteria
2.2. LAB Growth Curves
2.3. Metabolite Production
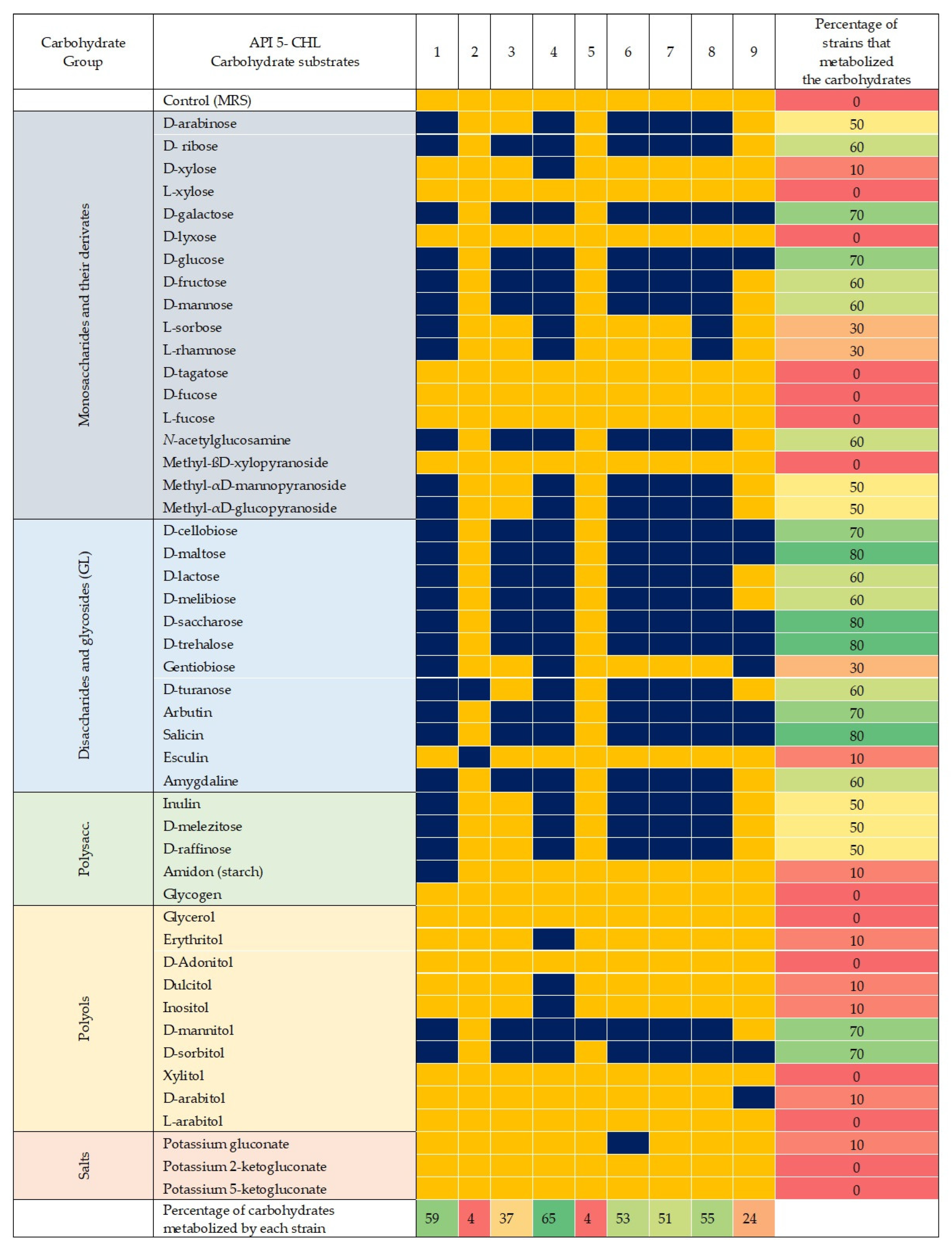
2.4. Metabolic Profile
2.5. Bioreactor Fermentation
2.6. Data Analysis
3. Results
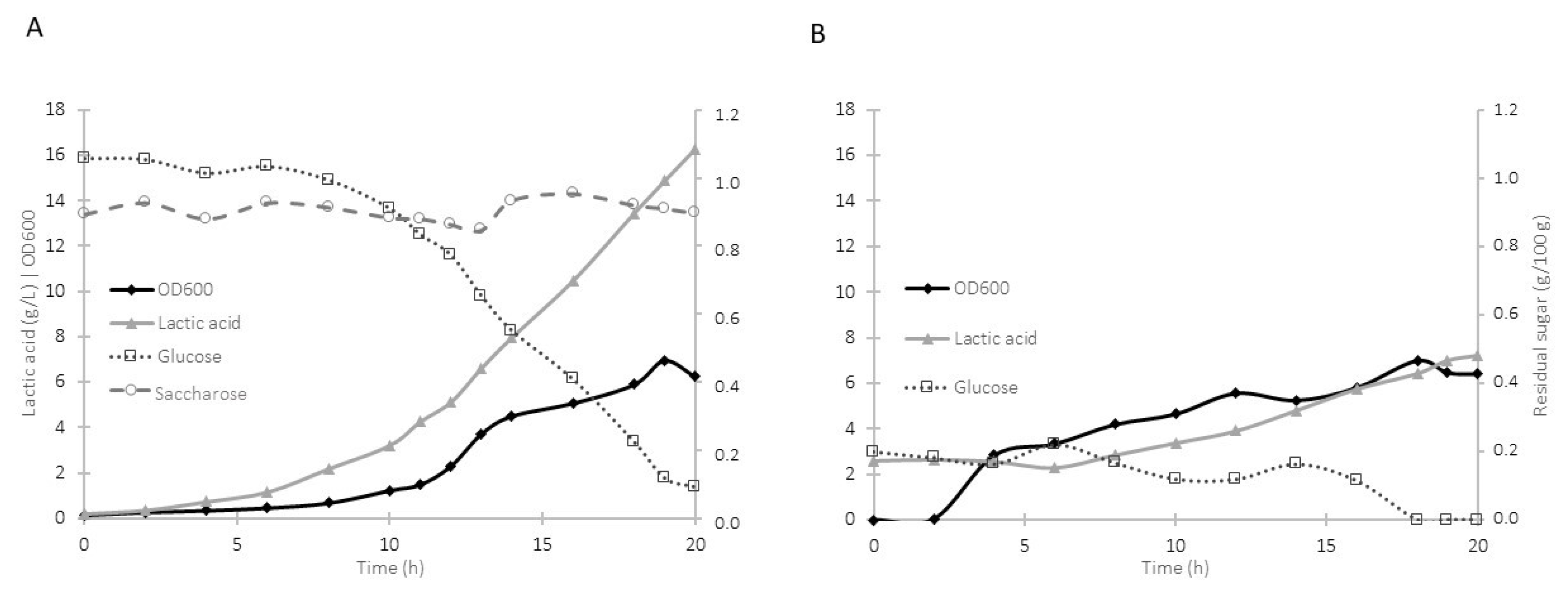
3.1. Growth Kinetics and Bioreactor Fermentation
3.2. Growth Kinetics and Microbioreactor Fermentation
3.3. Sugar Metabolism
3.4. Biorreactor Fermentation Using Synthetic Medium and Milk Whey
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mazzoli, R.; Bosco, F.; Mizrahi, I.; Bayer, E.A.; Pessione, E. Towards lactic acid bacteria-based biorefineries. Biotechnol. Adv. 2014, 32, 1216–1236. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chan, S.H.J.; Chen, J.; Solem, C.; Jensen, P.R. Systems biology—A guide for understanding and developing improved strains of lactic acid bacteria. Front. Microbiol. 2019, 10, 876. [Google Scholar] [CrossRef] [PubMed]
- Mora-Villalobos, J.A.; Montero-Zamora, J.; Barboza, N.; Rojas-Garbanzo, C.; Usaga, J.; Redondo-Solano, M.; Schroedter, L.; Olszewska-Widdrat, A.; López-Gómez, J.P. Multi-product lactic acid bacteria fermentations: A review. Fermentation 2020, 6, 23. [Google Scholar] [CrossRef]
- Champagne, C.P. The lactic acid bacteria. Int. Dairy J. 1994, 4, 665–666. [Google Scholar] [CrossRef]
- Sakr, E.A.E.; Massoud, M.I.; Ragaee, S. Food wastes as natural sources of lactic acid bacterial exopolysaccharides for the functional food industry: A review. Int. J. Biol. Macromol. 2021, 189, 232–241. [Google Scholar] [CrossRef]
- Börner, R.A.; Kandasamy, V.; Axelsen, A.M.; Nielsen, A.T.; Bosma, E.F. Genome editing of lactic acid bacteria: Opportunities for food, feed, pharma and biotech. FEMS Microbiol. Lett. 2019, 366, fny291. [Google Scholar] [CrossRef]
- Wen Fang Wu Wu, J.; Redondo-Solano, M.; Uribe, L.; WingChing-Jones, R.; Usaga, J.; Barboza, N. First characterization of the probiotic potential of lactic acid bacteria isolated from Costa Rican pineapple silages. PeerJ 2021, 9, e12437. [Google Scholar] [CrossRef]
- Eş, I.; Mousavi Khaneghah, A.; Barba, F.J.; Saraiva, J.A.; Sant’Ana, A.S.; Hashemi, S.M.B. Recent advancements in lactic acid production—A review. Food Res. Int. 2018, 107, 763–770. [Google Scholar] [CrossRef]
- Ibrahim, S.A.; Ayivi, R.D.; Zimmerman, T.; Siddiqui, S.A.; Altemimi, A.B.; Fidan, H.; Esatbeyoglu, T.; Bakhshayesh, R.V. Lactic acid bacteria as antimicrobial agents: Food safety and microbial food spoilage prevention. Foods 2021, 10, 3131. [Google Scholar] [CrossRef]
- Baiano, A. Recovery of biomolecules from food wastes—A review. Molecules 2014, 19, 14821–14842. [Google Scholar] [CrossRef]
- Tsegaye, B.; Jaiswal, S.; Jaiswal, A.K. Food waste biorefinery: Pathway towards circular bioeconomy. Foods 2021, 10, 1174. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Banat, F.; Taher, H. A review on the lactic acid fermentation from low-cost renewable materials: Recent developments and challenges. Environ. Technol. Inno. 2020, 20, 101138. [Google Scholar] [CrossRef]
- Pires, A.F.; Marnotes, N.G.; Rubio, O.D.; Garcia, A.C.; Pereira, C.D. Dairy By-Products: A review on the valorization of whey and second cheese whey. Foods 2021, 10, 1067. [Google Scholar] [CrossRef]
- Chen, G.Q.; Leong, T.S.H.; Kentish, S.E.; Ashokkumar, M.; Martin, G.J.O. Membrane separations in the dairy industry. In Separation of Functional Molecules in Food by Membrane Technology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 267–304. ISBN 978-0-12-815056-6. [Google Scholar]
- León-López, A.; Pérez-Marroquín, X.A.; Campos-Lozada, G.; Campos-Montiel, R.G.; Aguirre-Álvarez, G. Characterization of whey-based fermented beverages supplemented with hydrolyzed collagen: Antioxidant activity and bioavailability. Foods 2020, 9, 1106. [Google Scholar] [CrossRef]
- Barba, F.J. An integrated approach for the valorization of cheese whey. Foods 2021, 10, 564. [Google Scholar] [CrossRef]
- Edwards, U.; Rogall, T.; Blöcker, H.; Emde, M.; Böttger, E.C. Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucl Acids Res. 1989, 17, 7843–7853. [Google Scholar] [CrossRef] [PubMed]
- Hayek, S.A.; Ibrahim, S.A. Current limitations and challenges with lactic acid bacteria: A review. Food Nutr. Sci. 2013, 4, 73–87. [Google Scholar] [CrossRef]
- Montero-Zamora, J.; Cortés-Muñoz, M.; Esquivel, P.; Mora-Villalobos, J.; Velázquez, C. Growth conditions and survival kinetics during storage of Lactobacillus rhamnosus GG for the design of a sustainable probiotic whey-based beverage containing costa rican guava fruit pulp. J. Food Sci. 2020, 85, 3478–3486. [Google Scholar] [CrossRef]
- Olszewska-Widdrat, A.; Alexandri, M.; López-Gómez, J.P.; Schneider, R.; Venus, J. Batch and continuous lactic acid fermentation based on a multi-substrate approach. Microorganisms 2020, 8, 1084. [Google Scholar] [CrossRef]
- Soares, M.; Christen, P.; Pandey, A.; Soccol, C.R. Fruity flavour production by ceratocystis fimbriata grown on coffee husk in solid-state fermentation. Process Biochem. 2000, 35, 857–861. [Google Scholar] [CrossRef]
- Cox, R.; Narisetty, V.; Nagarajan, S.; Agrawal, D.; Ranade, V.V.; Salonitis, K.; Venus, J.; Kumar, V. High-level fermentative production of lactic acid from bread waste under non-sterile conditions with a circular biorefining approach and zero waste discharge. Fuel 2022, 313, 122976. [Google Scholar] [CrossRef]
- López-Gómez, J.P.; Alexandri, M.; Schneider, R.; Venus, J. A review on the current developments in continuous lactic acid fermentations and case studies utilising inexpensive raw materials. Process Biochem. 2019, 79, 1–10. [Google Scholar] [CrossRef]
- Tian, X.; Liu, X.; Zhang, Y.; Chen, Y.; Hang, H.; Chu, J.; Zhuang, Y. Metabolic engineering coupled with adaptive evolution strategies for the efficient production of high-quality L-lactic acid by Lactobacillus paracasei. Bioresour. Technol. 2021, 323, 124549. [Google Scholar] [CrossRef]
- Fessard, A.; Remize, F. Why are Weissella spp. not used as commercial starter cultures for food fermentation? Fermentation 2017, 3, 38. [Google Scholar] [CrossRef]
- Teixeira, C.G.; Fusieger, A.; Milião, G.L.; Martins, E.; Drider, D.; Nero, L.A.; de Carvalho, A.F. Weissella: An emerging bacterium with promising health benefits. Probiotics Antimicro. Prot. 2021, 13, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.D.; Samelis, J.; Metaxopoulos, J.; Wallbanks, S. Taxonomic studies on some Leuconostoc-like organisms from fermented sausages: Description of a new genus Weissella for the Leuconostoc paramesenteroides group of species. J. Appl. Bacteriol. 1993, 75, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Micanquer-Carlosama, A.; Cortés-Rodríguez, M.; Serna-Cock, L. Formulation of a fermentation substrate from pineapple and sacha inchi wastes to grow Weissella cibaria. Heliyon 2020, 6, e03790. [Google Scholar] [CrossRef]
- Micanquer-Carlosama, A.; Cortés-Rodríguez, M.; Correa-Londoño, G.; Orozco-Sánchez, F.; Serna-Cock, L. Optimization of the reproduction of Weissella cibaria in a fermentation substrate formulated with agroindustrial waste. Biotechnol. Rep. 2021, 32, e00671. [Google Scholar] [CrossRef]
- Zannini, E.; Jeske, S.; Lynch, K.M.; Arendt, E.K. Development of novel quinoa-based yoghurt fermented with dextran producer Weissella cibaria MG1. Int. J. Food Microbiol. 2018, 268, 19–26. [Google Scholar] [CrossRef]
- Dedenaro, G.; Costa, S.; Rugiero, I.; Pedrini, P.; Tamburini, E. Valorization of agri-food waste via fermentation: Production of L-lactic acid as a building block for the synthesis of biopolymers. Appl. Sci. 2016, 6, 379. [Google Scholar] [CrossRef]
- Araya-Cloutier, C.; Rojas-Garbanzo, C.; Velázquez-Carrillo, C. Effect of initial sugar concentration on the production of L (+) lactic acid by simultaneous enzymatic hydrolysis and fermentation of an agro-industrial waste product of pineapple (Ananas comosus) using Lactobacillus casei subspecies rhamnosus. Int. J. Biotech Well Ind. 2012, 1, 91–100. [Google Scholar] [CrossRef][Green Version]
- Alvarez, M.M.; Aguirre-Ezkauriatza, E.J.; Ramírez-Medrano, A.; Rodríguez-Sánchez, A. Kinetic analysis and mathematical modeling of growth and lactic acid production of Lactobacillus casei var. rhamnosus in milk whey. J. Dairy Sci. 2010, 93, 5552–5560. [Google Scholar] [CrossRef] [PubMed]
- Serna, L.; Rengifo, C.A.; Rojas, M.A. The use of earthworm flour for lactic acid biomass production. Afr. J. Biotechnol. 2013, 12, 5962–5967. [Google Scholar]
- Zhang, Z.; Tsapekos, P.; Alvarado-Morales, M.; Kovalovszki, A.; Yang, X.; Zhu, X.; Angelidaki, I. Improving lactic acid production via bio-augmentation with acid-tolerant isolates from source-sorted organic household waste. Biomass Conv. Bioref. 2022. [Google Scholar] [CrossRef]
- Xiang, W.-L.; Zhang, N.-D.; Lu, Y.; Zhao, Q.-H.; Xu, Q.; Rao, Y.; Liu, L.; Zhang, Q. Effect of Weissella cibaria co-inoculation on the quality of sichuan pickle fermented by Lactobacillus plantarum. LWT 2020, 121, 108975. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Strain (GenBank Accession Number) | Source | Maximal OD600 | Growth Rate (h−1) * |
|---|---|---|---|
| Lactiplantibacillus pentosus_70-1 (ON763307) | Orange waste residuals | 1.662 ± 0.011 a | 0.139 ± 0.013 i,j,k,l |
| Lactiplantibacillus pentosus_19-2 (ON763302) | Coffee brush | 1.563 ± 0.091 a,b,c | 0.267 ± 0.016 a,b |
| Lactiplantibacillus pentosus_71-6 (ON763308) | Orange waste residuals | 1.554 ± 0.051 a,b,c | 0.238 ± 0.011 b,c,d,e |
| Schleiferilactobacillus harbinensis_P7 (ON763331) | MFC with coffee syrup waters | 1.542 ± 0.023 a,b,c | 0.196 ± 0.008 d,e,f,g,h |
| Lacticaseibacillus paracasei subsp. tolerans_P6 (ON763290) | MFC with coffee syrup waters | 1.510 ± 0.014 a,b,c | 0.182 ± 0.003 e,f,g,h,i,j |
| Weissella soli_31 (ON763315) | Carrot waste residues | 1.504 ± 0.059 a,b,c | 0.256 ± 0.021 a,b,c |
| Weissella soli_29 (ON763313) | Carrot waste residues | 1.500 ± 0.045 a,b,c | 0.229 ± 0.014 b,c,d,e,f |
| Lactiplantibacillus pentosus_17-2 (ON763301) | Coffee brush | 1.492 ± 0.039 a,b,c | 0.224 ± 0.013 b,c,d,e,f |
| Lacticaseibacillus paracasei subsp. tolerans_P8 (ON763291) | MFC with coffee syrup waters | 1.490 ± 0.057 a,b,c | 0.194 ± 0.023 d,e,f,g,h,i |
| Lacticaseibacillus paracasei subsp. tolerans_P2 (ON763288) | MFC with coffee syrup waters | 1.488 ± 0.104 a,b,c | 0.176 ± 0.034 f,g,h,i,j,k |
| Leuconostoc falkenbergense_66-2 (ON763311) | Orange waste residuals | 1.477 ± 0.058 a,b,c | 0.225 ± 0.049 b,c,d,e,f |
| Lacticaseibacillus paracasei subsp. tolerans_P10 (ON763293) | MFC with coffee syrup waters | 1.471 ± 0.028 a,b,c | 0.201 ± 0.033 c,d,e,f,g |
| Lacticaseibacillus paracasei subsp. tolerans_I-C2 (ON763285) | MFC with coffee syrup waters | 1.470 ± 0.061 a,b,c | 0.189 ± 0.014 e,f,g,h,i,j |
| Lactiplantibacillus pentosus_68-3 (ON763305) | Orange waste residuals | 1.469 ± 0.035 a,b,c | 0.231 ± 0.044 b,c,d,e,f |
| Lactiplantibacillus pentosus_69 (ON763306) | Orange waste residuals | 1.457 ± 0.086 a,b,c | 0.225 ± 0.019 b,c,d,e,f |
| Lacticaseibacillus paracasei subsp. tolerans_II-C2-C (ON763287) | MFC with coffee syrup waters | 1.453 ± 0.118 a,b,c | 0.191 ± 0.008 d,e,f,g,h,i,j |
| Lacticaseibacillus paracasei subsp. tolerans_P9 (ON763292) | MFC with coffee syrup waters | 1.448 ± 0.137 a,b,c | 0.157 ± 0.031 g,h,i,j,k |
| Lacticaseibacillus paracasei subsp. tolerans_IA2-P (ON763283) | MFC with coffee syrup waters | 1.448 ± 0.040 a,b,c | 0.197 ± 0.022 d,e,f,g,h |
| Lacticaseibacillus paracasei subsp. tolerans_2A2-B (ON763280) | MFC with coffee syrup waters | 1.427 ± 0.031 a,b,c | 0.167 ± 0.044 g,h,i,j,k |
| Lactiplantibacillus paraplantarum_19-1 (ON763312) | Coffee brush | 1.420 ± 0.091 b,c | 0.247 ± 0.032 b,c,d |
| Leuconostoc pseudomesenteroides_18 (ON763310) | Coffee brush | 1.413 ± 0.121 c | 0.163 ± 0.087 g,h,i,j,k |
| Lacticaseibacillus paracasei subsp. tolerans_II-C1-B (ON763286) | MFC with coffee syrup waters | 1.412 ± 0.101 c | 0.157 ± 0.020 g,h,i,j,k |
| Lacticaseibacillus paracasei subsp. tolerans_P13 (ON763294) | MFC with coffee syrup waters | 1.399 ± 0.105 c | 0.183 ± 0.029 e,f,g,h,i,j |
| Lactiplantibacillus pentosus_16 (ON763300) | Coffee brush | 1.389 ± 0.131 c | 0.210 ± 0.020 c,d,e,f,g |
| Levilactobacillus brevis_68-1 (ON763329) | Orange waste residuals | 1.383 ± 0.202 c | 0.162 ± 0.057 g,h,i,j,k |
| Lacticaseibacillus paracasei subsp. tolerans_P4 (ON763289) | MFC with coffee syrup waters | 1.352 ± 0.019 c,d | 0.137 ± 0.012 j,k,l |
| Lactiplantibacillus argentoratensis_70-2 (ON763327) | Orange waste residuals | 1.332 ± 0.105 c,d | 0.160 ± 0.021 g,h,i,j,k |
| Lacticaseibacillus paracasei subsp. tolerans_II-C1-C (ON763282) | MFC with coffee syrup waters | 1.116 ± 0.666 d,e | 0.115 ± 0.081 k,l |
| Leuconostoc pseudomesenteroides_17-1 (ON763309) | Coffee brush | 0.970 ± 0.066 e | 0.142 ± 0.011 h,i,j,k,l |
| Weissella soli_30-2 (ON763314) | Carrot waste residues | 0.401 ± 0.226 f | 0.048 ± 0.024 m |
| Strain | Lactic Acid (g/L) | Acetic Acid (g/L) | Residual Glucose (g/100 g) |
|---|---|---|---|
| Weissella soli_29 | 20.833 ± 2.731 a | 4.023 ± 0.680 a,b | 0.000 ± 0.000 c,d,e,f |
| Lactiplantibacillus pentosus_70-1 | 20.100 ± 2.754 a,b | 4.367 ± 0.626 a | 0.000 ± 0.000 e,f |
| Lactiplantibacillus pentosus_69 | 20.100 ± 2.562 a | 4.115 ± 0.660 a,b | 0.069 ± 0.072 c,d |
| Lactiplantibacillus pentosus_17-2 | 19.867 ± 2.757 a | 3.965 ± 0.553 a,b | 0.000 ± 0.000 d |
| Lacticaseibacillus paracasei subsp. tolerans_P10 | 19.850 ± 2.829 a | 3.868 ± 0.492 a,b | 0.183 ± 0.193 c,d |
| Weissella soli_31 | 19.350 ± 3.336 a | 3.490 ± 0.595 b | 0.000 ± 0.000 d |
| Lactiplantibacillus pentosus_68-3 | 19.133 ± 2.553 a,b | 3.530 ± 0.470 a,b | 0.000 ± 0.000 d,e,f |
| Leuconostoc falkenbergense_66-2 | 18.633 ± 2.666 a,b | 3.888 ± 0.698 a,b | 0.000 ± 0.000 d |
| Lactiplantibacillus pentosus_19-2 | 18.550 ± 2.533 a,b | 3.858 ± 0.586 a,b | 0.062 ± 0.065 c,d |
| MRS | 0.000 ± 0.000 d | 4.233 ± 0.646 a | 1.998 ± 0.273 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montero-Zamora, J.; Fernández-Fernández, S.; Redondo-Solano, M.; Mazón-Villegas, B.; Mora-Villalobos, J.A.; Barboza, N. Assessment of Different Lactic Acid Bacteria Isolated from Agro-Industrial Residues: First Report of the Potential Role of Weissella soli for Lactic Acid Production from Milk Whey. Appl. Microbiol. 2022, 2, 626-635. https://doi.org/10.3390/applmicrobiol2030048
Montero-Zamora J, Fernández-Fernández S, Redondo-Solano M, Mazón-Villegas B, Mora-Villalobos JA, Barboza N. Assessment of Different Lactic Acid Bacteria Isolated from Agro-Industrial Residues: First Report of the Potential Role of Weissella soli for Lactic Acid Production from Milk Whey. Applied Microbiology. 2022; 2(3):626-635. https://doi.org/10.3390/applmicrobiol2030048
Chicago/Turabian StyleMontero-Zamora, Jéssica, Silvia Fernández-Fernández, Mauricio Redondo-Solano, Beatriz Mazón-Villegas, José Aníbal Mora-Villalobos, and Natalia Barboza. 2022. "Assessment of Different Lactic Acid Bacteria Isolated from Agro-Industrial Residues: First Report of the Potential Role of Weissella soli for Lactic Acid Production from Milk Whey" Applied Microbiology 2, no. 3: 626-635. https://doi.org/10.3390/applmicrobiol2030048
APA StyleMontero-Zamora, J., Fernández-Fernández, S., Redondo-Solano, M., Mazón-Villegas, B., Mora-Villalobos, J. A., & Barboza, N. (2022). Assessment of Different Lactic Acid Bacteria Isolated from Agro-Industrial Residues: First Report of the Potential Role of Weissella soli for Lactic Acid Production from Milk Whey. Applied Microbiology, 2(3), 626-635. https://doi.org/10.3390/applmicrobiol2030048