
Limnospira indica PCC 8005 or Lacticaseibacillus rhamnosus GG Dietary Supplementation Modulate the Gut Microbiome in Mice

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
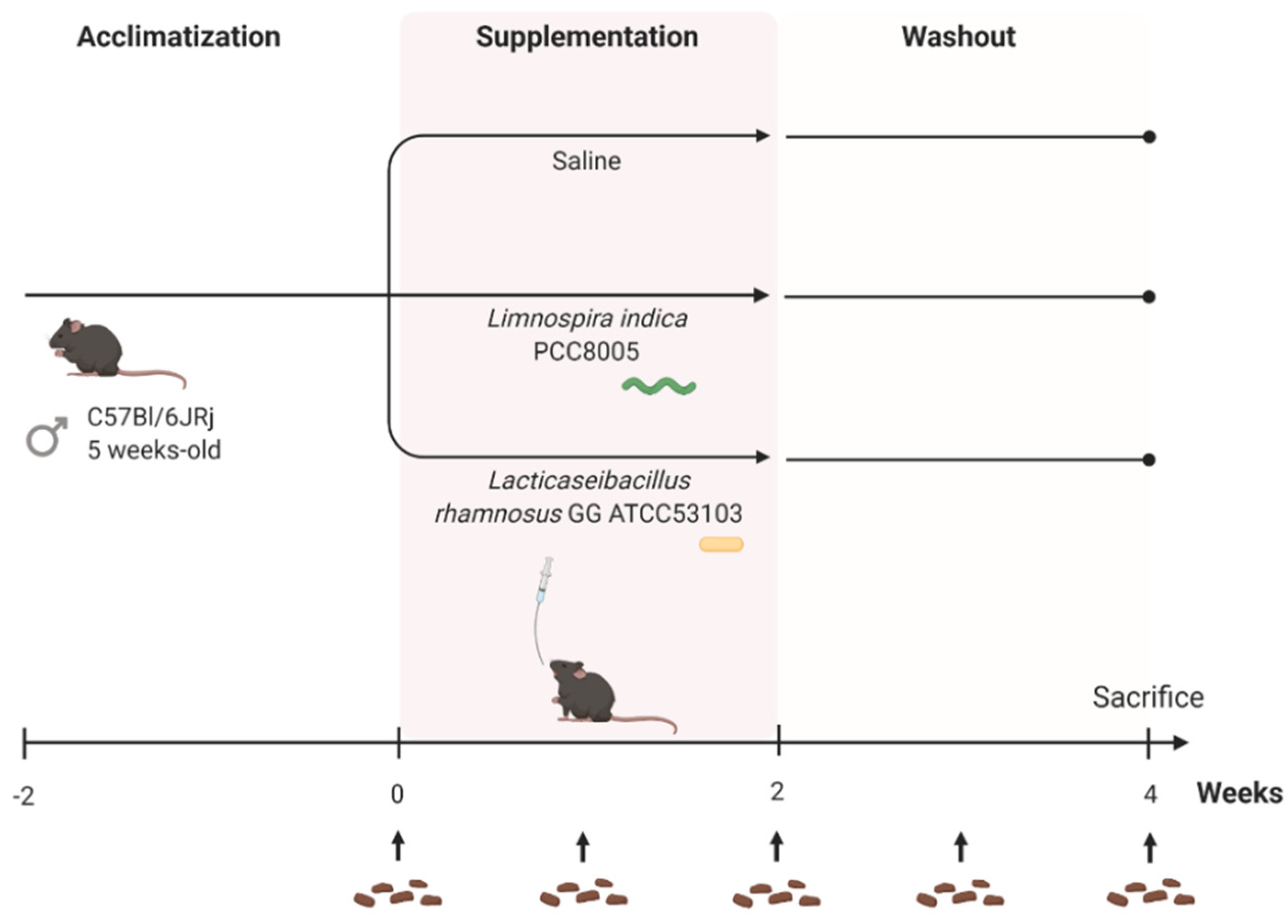
2.1. Mice
2.2. Bacterial Strains and Growth Conditions
2.3. Supplementation Protocol
2.4. Fecal DNA Extraction and 16S rRNA Gene Sequencing
2.5. Sequencing Data Processing and Analyses
2.6. Statistical Analysis
3. Results
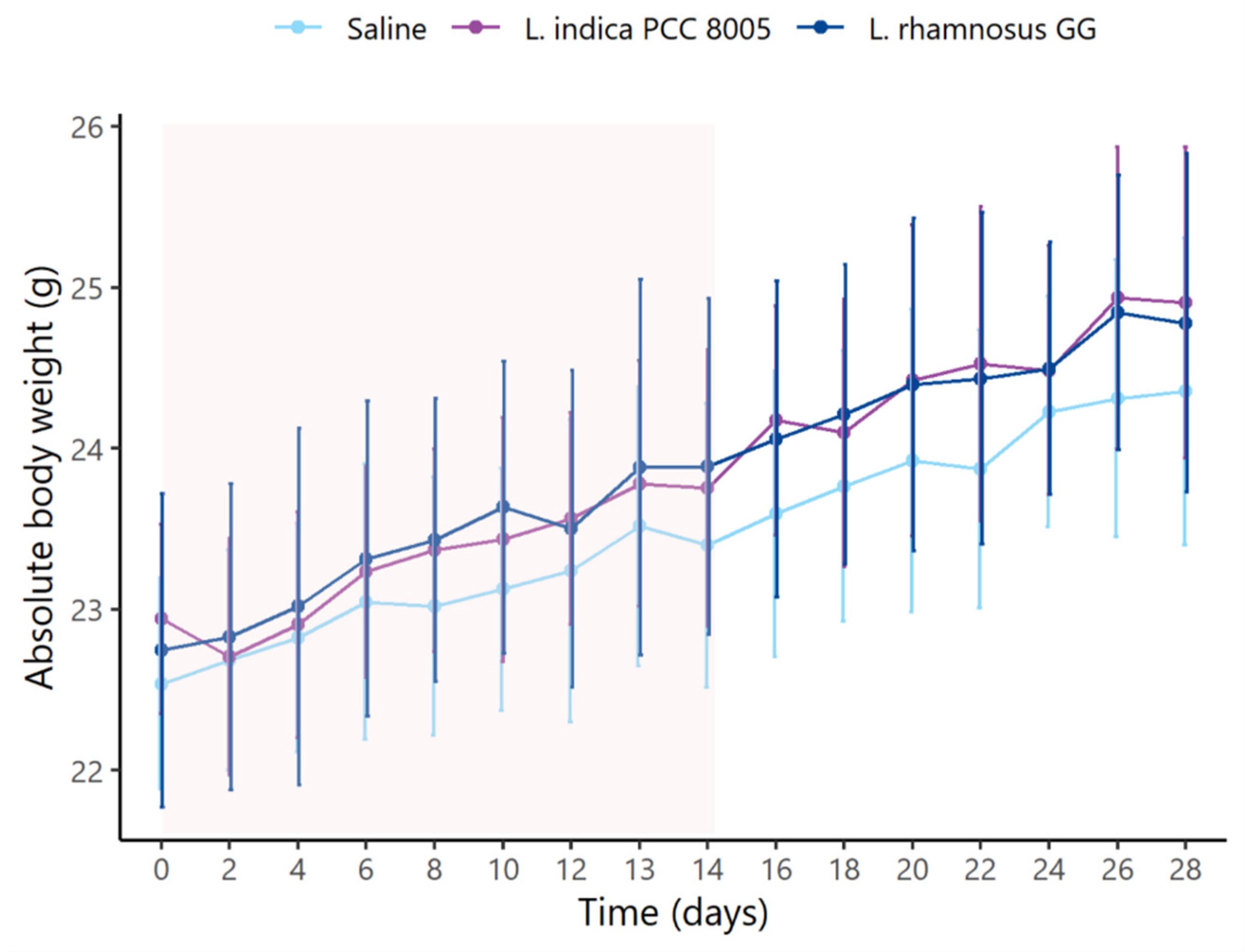
3.1. L. indica PCC 8005 and L. rhamnosus GG Supplementation Does Not Affect Body Weight
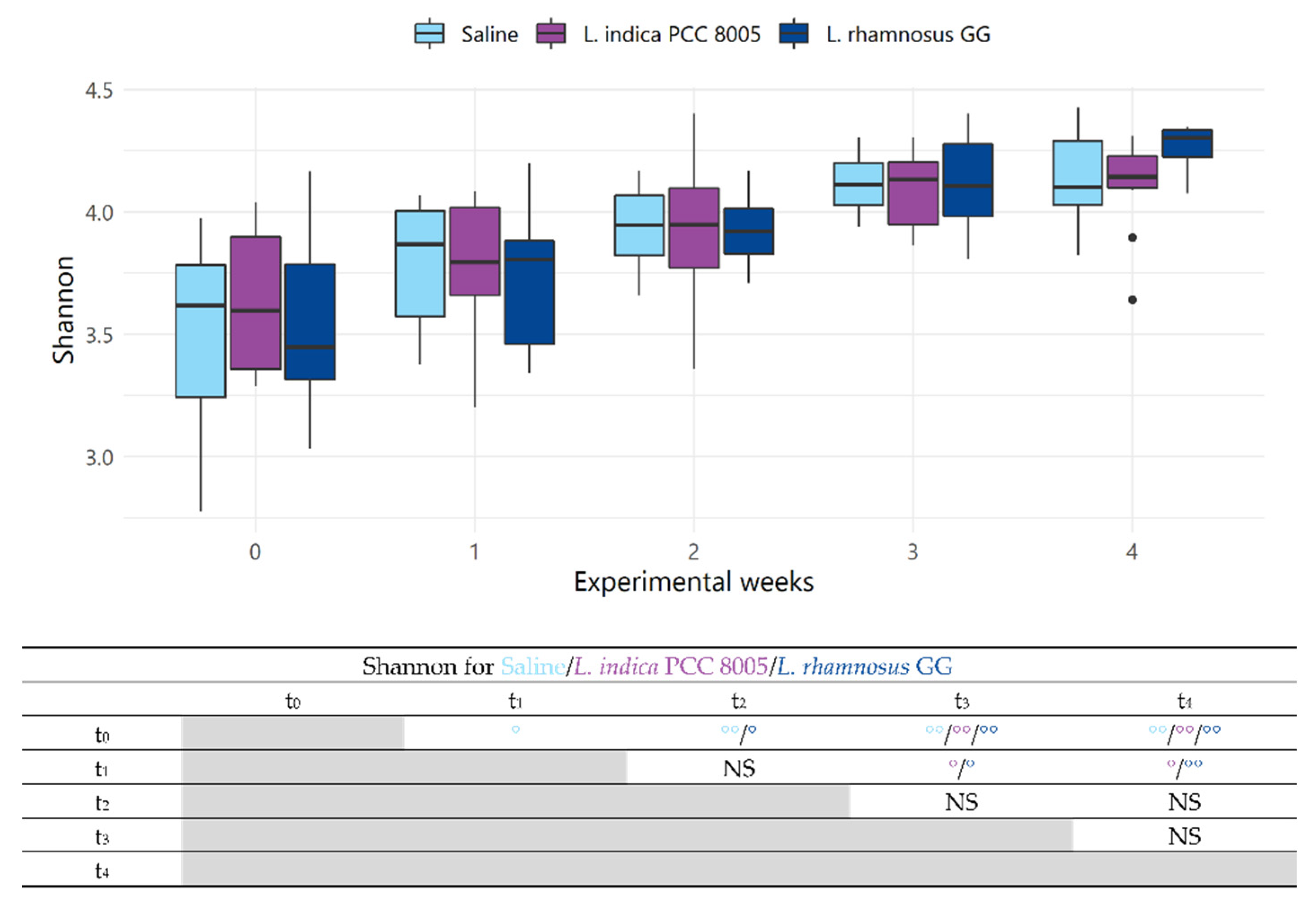
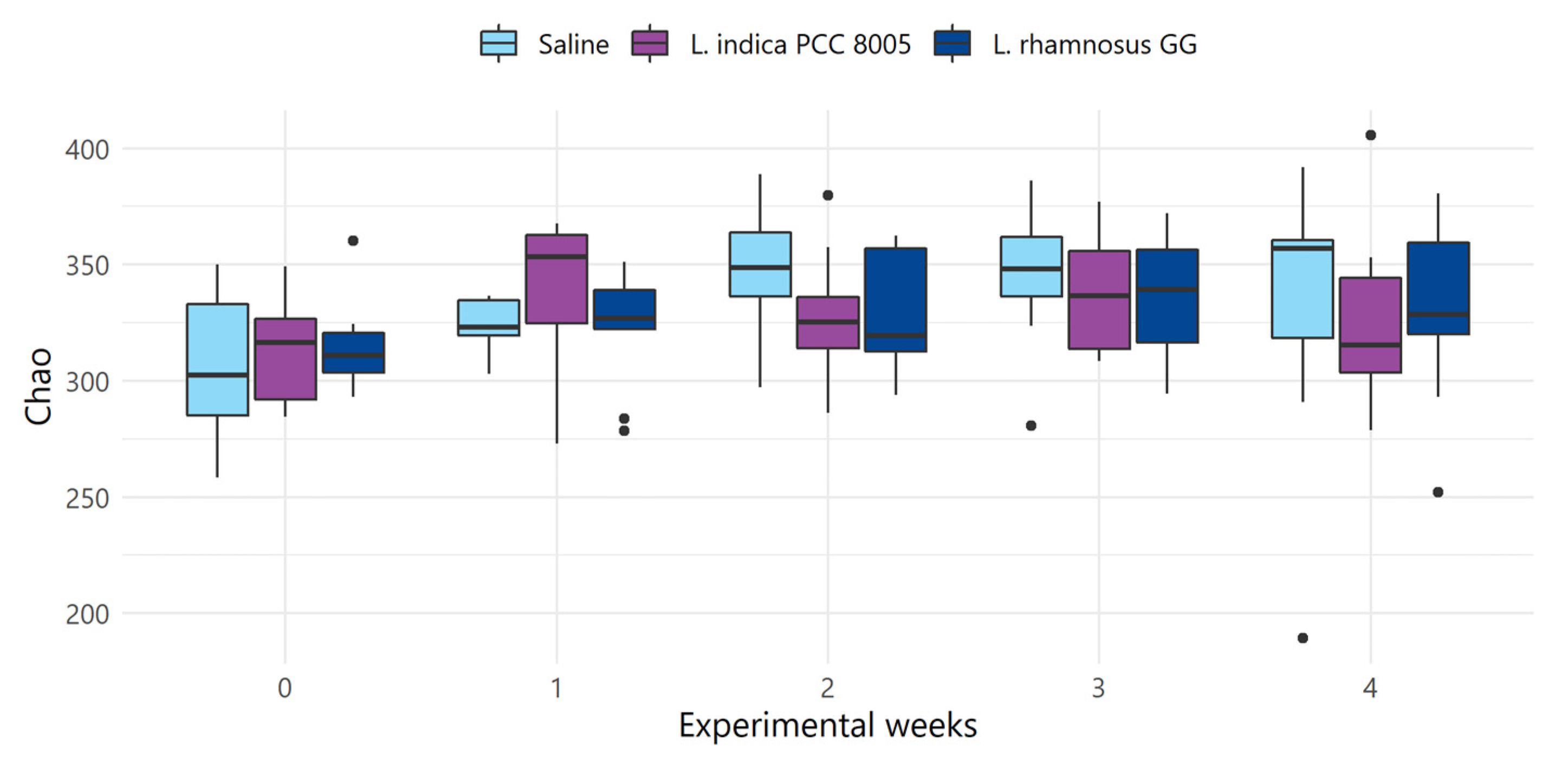
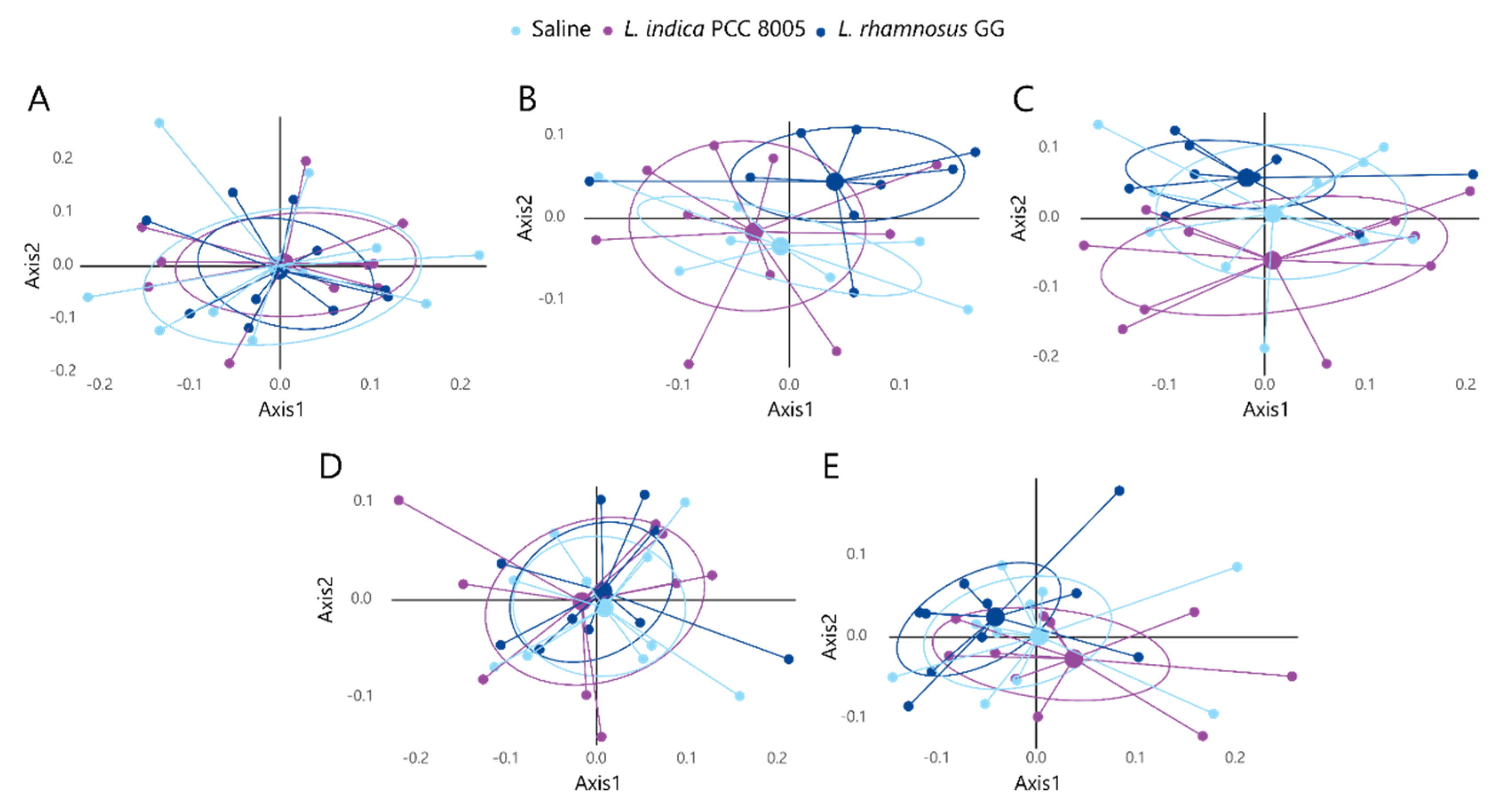
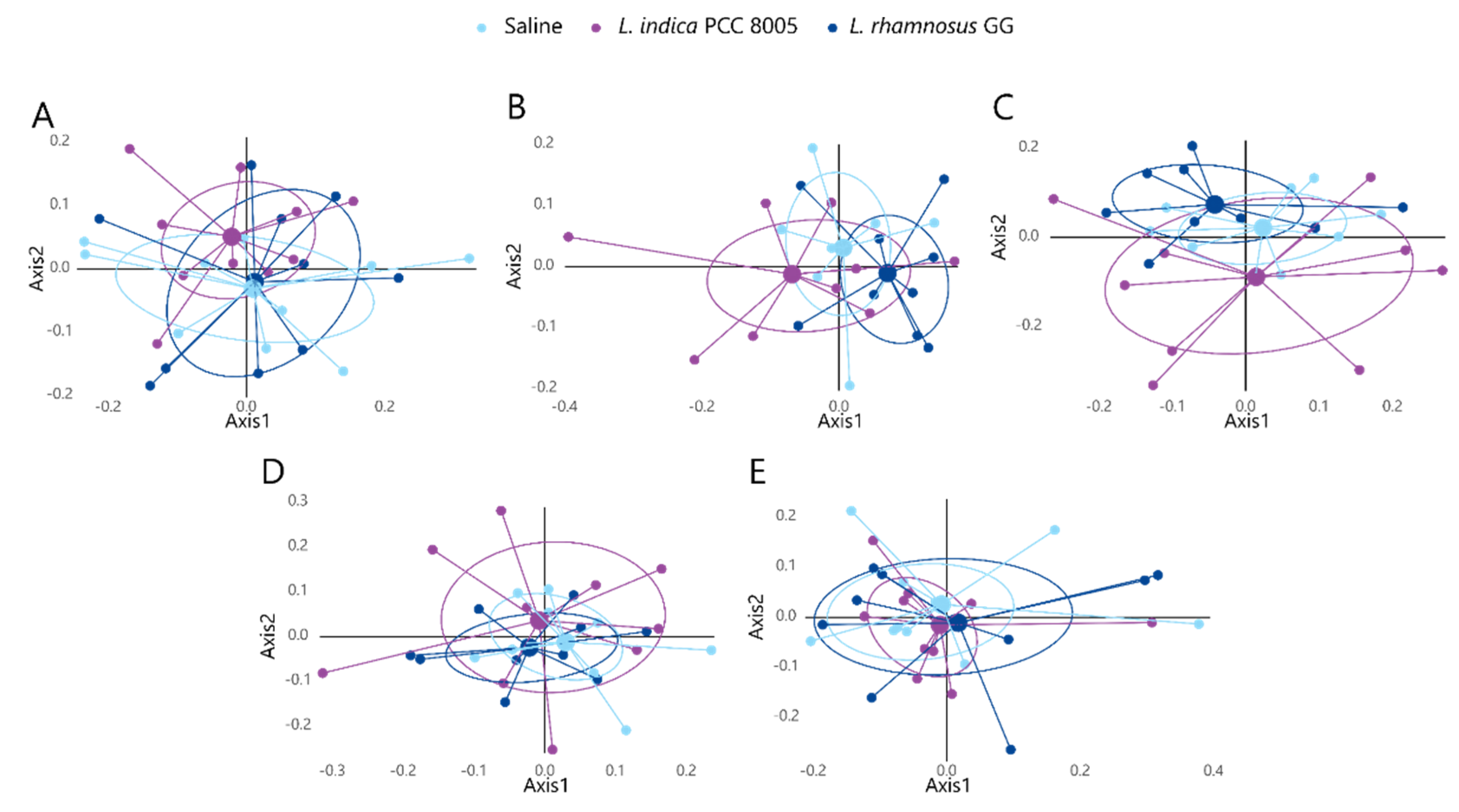
3.2. The Intestinal Microbial Community Temporarily Changes following L. indica PCC 8005 and L. rhamnosus GG Supplementation
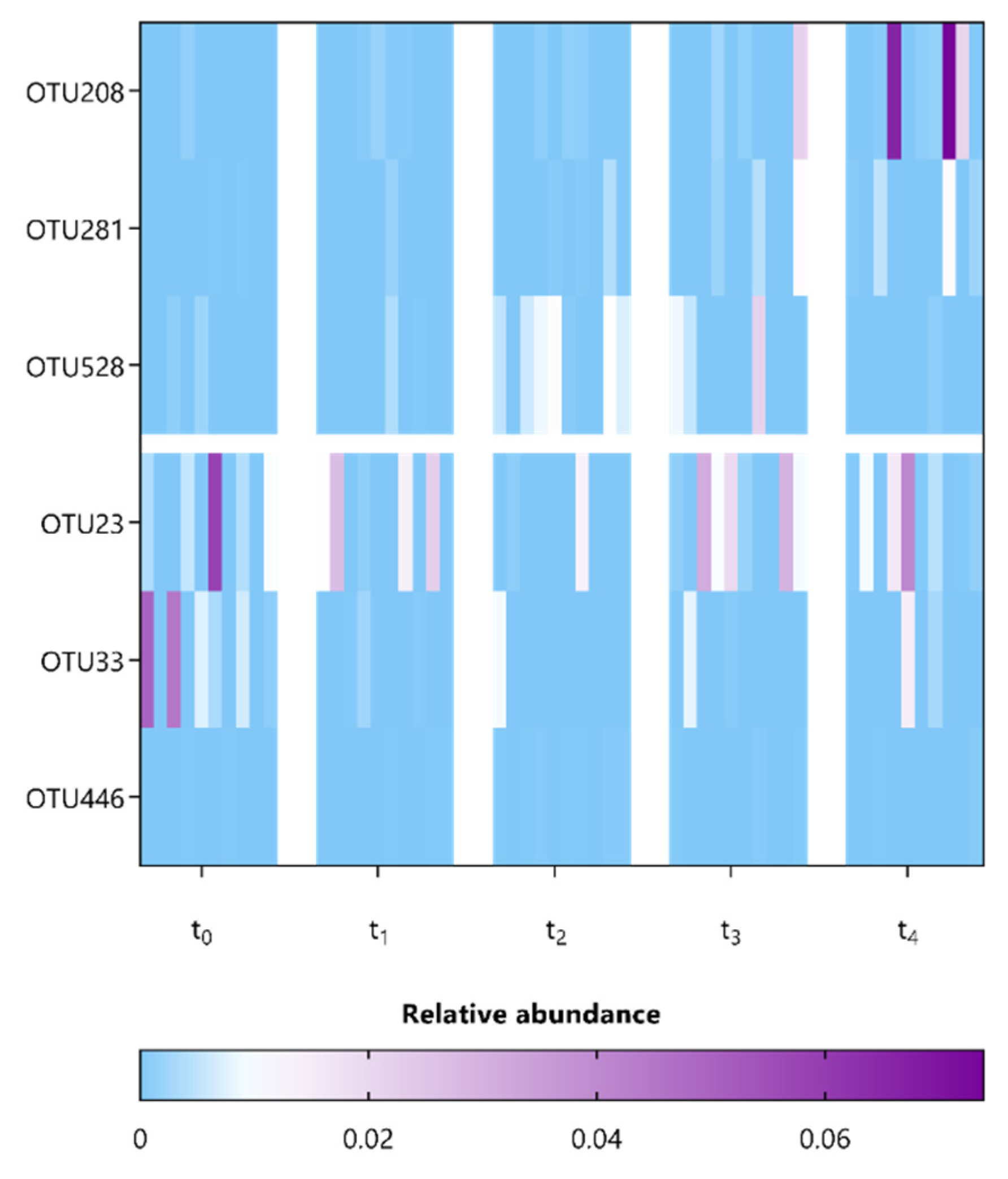
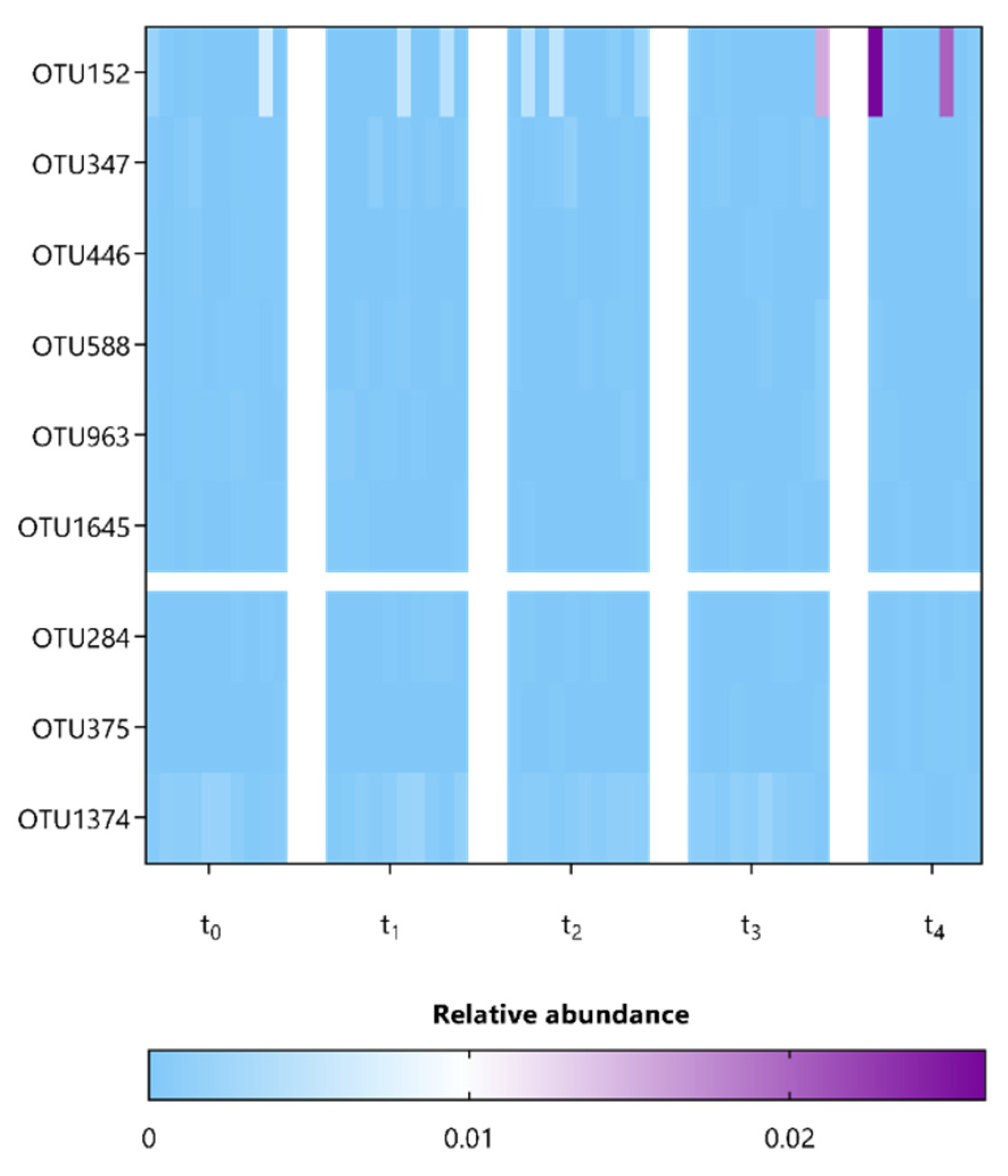
3.3. L. indica PCC 8005 and L. rhamnosus GG Supplementation Affect Members Belonging to the Lachnospiraceae and Porphyromonadaceae Families
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Vergès, M.C.; Charles, T.; Chen, X.; Cocolin, L.; Eversole, K.; Corral, G.H.; et al. Microbiome Definition Re-Visited: Old Concepts and New Challenges. Microbiome 2020, 8, 103. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.M.; Pereira-Marques, J.; Pinto-Ribeiro, I.; Costa, J.L.; Carneiro, F.; MacHado, J.C.; Figueiredo, C. Gastric Microbial Community Profiling Reveals a Dysbiotic Cancer-Associated Microbiota. Gut 2018, 67, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Ghavami, S.B.; Rostami, E.; Sephay, A.A.; Shahrokh, S.; Balaii, H.; Aghdaei, H.A.; Zali, M.R. Alterations of the Human Gut Methanobrevibacter Smithii as a Biomarker for Inflammatory Bowel Diseases. Microb. Pathog. 2018, 117, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Segers, C.; Verslegers, M.; Baatout, S.; Leys, N.; Lebeer, S.; Mastroleo, F. Food Supplements to Mitigate Detrimental Effects of Pelvic Radiotherapy. Microorganisms 2019, 7, 97. [Google Scholar] [CrossRef] [PubMed]
- Kleerebezem, M.; Binda, S.; Bron, P.A.; Gross, G.; Hill, C.; van Hylckama Vlieg, J.E.; Lebeer, S.; Satokari, R.; Ouwehand, A.C. Understanding Mode of Action Can Drive the Translational Pipeline towards More Reliable Health Benefits for Probiotics. Curr. Opin. Biotechnol. 2019, 56, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Wittouck, S.; Wuyts, S.; Meehan, C.J.; van Noort, V.; Lebeer, S. A Genome-Based Species Taxonomy of the Lactobacillus Genus Complex. mSystems 2019, 4, e00264-19. [Google Scholar] [CrossRef]
- Ciorba, M.A.; Riehl, T.E.; Rao, M.S.; Moon, C.; Ee, X.; Nava, G.M.; Walker, M.R.; Marinshaw, J.M.; Stappenbeck, T.S.; Stenson, W.F. Lactobacillus Probiotic Protects Intestinal Epithelium from Radiation Injury in a TLR-2/Cyclo-Oxygenase-2-Dependent Manner. Gut 2012, 61, 829–838. [Google Scholar] [CrossRef]
- Riehl, T.E.; Alvarado, D.; Ee, X.; Zuckerman, A.; Foster, L.; Kapoor, V.; Thotala, D.; Ciorba, M.A.; Stenson, W.F. Lactobacillus rhamnosus GG Protects the Intestinal Epithelium from Radiation Injury through Release of Lipoteichoic Acid, Macrophage Activation and the Migration of Mesenchymal Stem Cells. Gut 2018, 68, 1003–1013. [Google Scholar] [CrossRef]
- Österlund, P.; Ruotsalainen, T.; Korpela, R.; Saxelin, M.; Ollus, A.; Valta, P.; Kouri, M.; Elomaa, I.; Joensuu, H. Lactobacillus Supplementation for Diarrhoea Related to Chemotherapy of Colorectal Cancer: A Randomised Study. Br. J. Cancer 2007, 97, 1028–1034. [Google Scholar] [CrossRef]
- Baù, M.; Moretti, A.; Bertoni, E.; Vazzoler, V.; Luini, C.; Agosti, M.; Salvatore, S. Risk and Protective Factors for Gastrointestinal Symptoms Associated with Antibiotic Treatment in Children: A Population Study. Pediatr. Gastroenterol. Hepatol. Nutr. 2020, 23, 35–48. [Google Scholar] [CrossRef]
- Esposito, C.; Roberti, A.; Turrà, F.; Cerulo, M.; Severino, G.; Settimi, A.; Escolino, M. Frequency of Antibiotic-Associated Diarrhea and Related Complications in Pediatric Patients Who Underwent Hypospadias Repair: A Comparative Study Using Probiotics vs Placebo. Probiotics Antimicrob. Proteins 2018, 10, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Gamallat, Y.; Meyiah, A.; Kuugbee, E.D.; Hago, A.M.; Chiwala, G.; Awadasseid, A.; Bamba, D.; Zhang, X.; Shang, X.; Luo, F.; et al. Lactobacillus rhamnosus Induced Epithelial Cell Apoptosis, Ameliorates Inflammation and Prevents Colon Cancer Development in an Animal Model. Biomed. Pharmacother. 2016, 83, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Escamilla, J.; Lane, M.A.; Maitin, V. Cell-Free Supernatants from Probiotic Lactobacillus Casei and Lactobacillus rhamnosus GG Decrease Colon Cancer Cell Invasion in Vitro. Nutr. Cancer 2012, 64, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Yoda, K.; Miyazawa, K.; Hosoda, M.; Hiramatsu, M.; Yan, F.; He, F. Lactobacillus GG-Fermented Milk Prevents DSS-Induced Colitis and Regulates Intestinal Epithelial Homeostasis through Activation of Epidermal Growth Factor Receptor. Eur. J. Nutr. 2014, 53, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Claes, I.J.J.; Lebeer, S.; Shen, C.; Verhoeven, T.L.A.; Dilissen, E.; De Hertogh, G.; Bullens, D.M.A.; Ceuppens, J.L.; Van Assche, G.; Vermeire, S.; et al. Impact of Lipoteichoic Acid Modification on the Performance of the Probiotic Lactobacillus rhamnosus GG in Experimental Colitis. Clin. Exp. Immunol. 2010, 162, 306–314. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M.; Farouk, S.M.; Madkour, F.F.; Azab, S.S. Anti-Inflammatory and Immunomodulatory Effects of Spirulina platensis in Comparison to Dunaliella salina in Acetic Acid-Induced Rat Experimental Colitis. Immunopharmacol. Immunotoxicol. 2015, 37, 126–139. [Google Scholar] [CrossRef]
- Morsy, M.A.; Gupta, S.; Nair, A.B.; Venugopala, K.N.; Greish, K.; El-Daly, M. Protective Effect of Spirulina platensis Extract against Dextran-Sulfate-Sodium-Induced Ulcerative Colitis in Rats. Nutrients 2019, 11, 2309. [Google Scholar] [CrossRef]
- Yu, T.; Wang, Y.; Chen, X.; Xiong, W.; Tang, Y.; Lin, L. Spirulina platensis Alleviates Chronic Inflammation with Modulation of Gut Microbiota and Intestinal Permeability in Rats Fed a High-Fat Diet. J. Cell. Mol. Med. 2020, 24, 8603–8613. [Google Scholar] [CrossRef]
- Hu, J.; Li, Y.; Pakpour, S.; Wang, S.; Pan, Z.; Liu, J.; Wei, Q.; She, J.; Cang, H.; Zhang, R.X. Dose Effects of Orally Administered Spirulina Suspension on Colonic Microbiota in Healthy Mice. Front. Cell. Infect. Microbiol. 2019, 9, 243. [Google Scholar] [CrossRef]
- Najdenski, H.M.; Gigova, L.G.; Iliev, I.I.; Pilarski, P.S.; Lukavský, J.; Tsvetkova, I.V.; Ninova, M.S.; Kussovski, V.K. Antibacterial and Antifungal Activities of Selected Microalgae and Cyanobacteria. Int. J. Food Sci. Technol. 2013, 48, 1533–1540. [Google Scholar] [CrossRef]
- Alwaleed, E.A.; El-Sheekh, M.; Abdel-Daim, M.M.; Saber, H. Effects of Spirulina platensis and Amphora coffeaeformis as Dietary Supplements on Blood Biochemical Parameters, Intestinal Microbial Population, and Productive Performance in Broiler Chickens. Environ. Sci. Pollut. Res. 2020, 28, 1801–1811. [Google Scholar] [CrossRef] [PubMed]
- Papalia, T.; Sidari, R.; Panuccio, M.R. Impact of Different Storage Methods on Bioactive Compounds in Arthrospira platensis Biomass. Molecules 2019, 24, 2810. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, J.; Leone, V.; Nobutani, K.; Musch, M.W.; Martinez-Guryn, K.; Wang, Y.; Miyoshi, S.; Bobe, A.M.; Murat Eren, A.; Chang, E.B. Minimizing Confounders and Increasing Data Quality in Murine Models for Studies of the Gut Microbiome. PeerJ 2018, 6, e5166. [Google Scholar] [CrossRef] [PubMed]
- Cogne, G.; Lehmann, B.; Dussap, C.-G.; Gros, J.-B. Uptake of Macrominerals and Trace Elements by the Cyanobacterium Spirulina platensis (Arthrospira platensis PCC 8005) under Photoautotrophic Conditions: Culture Medium Optimization. Biotechnol. Bioeng. 2003, 81, 588–593. [Google Scholar] [CrossRef]
- Doron, S.; Snydman, D.R.; Gorbach, S.L. Lactobacillus GG: Bacteriology and Clinical Applications. Gastroenterol. Clin. N. Am. 2005, 34, 483–498. [Google Scholar] [CrossRef]
- Gorbach, S.L. The Discovery of Lactobacillus GG. Nutr. Today 1996, 31, 2S–4S. [Google Scholar] [CrossRef]
- De Man, J.C.; Rogosa, M.; Sharpe, M.E. A Medium for the Cultivation of Lactobacilli. J. Appl. Bacteriol. 1960, 23, 130–135. [Google Scholar] [CrossRef]
- Gamallat, Y.; Ren, X.; Walana, W.; Meyiah, A.; Xinxiu, R.; Zhu, Y.; Li, M.; Song, S.; Xie, L.; Jamalat, Y.; et al. Probiotic Lactobacillus rhamnosus Modulates the Gut Microbiome Composition Attenuates Preneoplastic Colorectal Aberrant Crypt Foci. J. Funct. Foods 2019, 53, 146–156. [Google Scholar] [CrossRef]
- Ki, Y.; Kim, W.; Cho, H.; Ahn, K.; Choi, Y.; Kim, D. The Effect of Probiotics for Preventing Radiation-Induced Morphological Changes in Intestinal Mucosa of Rats. J. Korean Med. Sci. 2014, 29, 1372–1378. [Google Scholar] [CrossRef]
- Li, X.; Hu, D.; Tian, Y.; Song, Y.; Hou, Y.; Sun, L.; Zhang, Y.; Man, C.; Zhang, W.; Jiang, Y. Protective Effects of a Novel Lactobacillus rhamnosus Strain with Probiotic Characteristics against Lipopolysaccharide-Induced Intestinal Inflammation In Vitro and In Vivo. Food Funct. 2020, 11, 5799–5814. [Google Scholar] [CrossRef]
- Mittal, A.; Suresh Kumar, P.V.; Banerjee, S.; Rao, A.R.; Kumar, A. Modulatory Potential of Spirulina fusiformis on Carcinogen Metabolizing Enzymes in Swiss Albino Mice. Phyther. Res. 1999, 13, 111–114. [Google Scholar] [CrossRef]
- Chamorro-Cevallos, G.; Garduño-Siciliano, L.; Barrón, B.L.; Madrigal-Bujaidar, E.; Cruz-Vega, D.E.; Pages, N. Chemoprotective Effect of Spirulina (Arthrospira) against Cyclophosphamide-Induced Mutagenicity in Mice. Food Chem. Toxicol. 2008, 46, 567–574. [Google Scholar] [CrossRef]
- El Bialy, B.E.; El-Boraey, N.G.; Hamouda, R.A.; Abdel-Daim, M.M. Comparative Protective Effects of Spirulina and Spirulina Supplemented with Thiamine against Sub-Acute Carbon Tetrachloride Toxicity in Rats. Biomed. Pharmacol. J. 2019, 12, 511–525. [Google Scholar] [CrossRef]
- Sharma, M.K.; Sharma, A.; Kumar, A.; Kumar, M. Spirulina fusiformis Provides Protection against Mercuric Chloride Induced Oxidative Stress in Swiss Albino Mice. Food Chem. Toxicol. 2007, 45, 2412–2419. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Rebolledo, G.A.; Galar-Martínez, M.; García-Rodríguez, R.V.; Chamorro-Cevallos, G.A.; Hernández-Reyes, A.G.; Martínez-Galero, E. Antioxidant Effect of Spirulina (Arthrospira) Maxima on Chronic Inflammation Induced by Freund’s Complete Adjuvant in Rats. J. Med. Food 2015, 18, 865–871. [Google Scholar] [CrossRef]
- Segers, C.; Mysara, M.; Claesen, J.; Baatout, S.; Leys, N.; Lebeer, S.; Verslegers, M.; Mastroleo, F. Intestinal Mucositis Precedes Dysbiosis in a Mouse Model for Pelvic Irradiation. ISME Commun. 2021, 1, 1–10. [Google Scholar] [CrossRef]
- Chao, A.; Jost, L. Coverage-Based Rarefaction and Extrapolation: Standardizing Samples by Completeness Rather than Size. Ecology 2012, 93, 2533–2547. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly Accurate OTU Sequences from Microbial Amplicon Reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Mandal, S.; Van Treuren, W.; White, R.A.; Eggesbø, M.; Knight, R.; Peddada, S.D. Analysis of Composition of Microbiomes: A Novel Method for Studying Microbial Composition. Microb. Ecol. Health Dis. 2015, 26, 27663. [Google Scholar] [CrossRef]
- Arantes-Rodrigues, R.; Henriques, A.; Pinto-Leite, R.; Faustino-Rocha, A.; Pinho-Oliveira, J.; Teixeira-Guedes, C.; Eixas, F.; Gama, A.; Colaço, B.; Colaço, A. The Effects of Repeated Oral Gavage on the Health of Male CD-1 Mice. Nature 2012, 41, 129–134. [Google Scholar] [CrossRef]
- Hutadilok-Towatana, N.; Reanmongkol, W.; Satitit, S.; Panichayupakaranant, P.; Ritthisunthorn, P. A Subchronic Toxicity Study of Spirulina platensis. Food Sci. Technol. Res. 2008, 14, 351–358. [Google Scholar] [CrossRef]
- Kim, S.W.; Park, K.Y.; Kim, B.; Kim, E.; Hyun, C.K. Lactobacillus rhamnosus GG Improves Insulin Sensitivity and Reduces Adiposity in High-Fat Diet-Fed Mice through Enhancement of Adiponectin Production. Biochem. Biophys. Res. Commun. 2013, 431, 258–263. [Google Scholar] [CrossRef]
- Ritze, Y.; Bárdos, G.; Claus, A.; Ehrmann, V.; Bergheim, I.; Schwiertz, A.; Bischoff, S.C. Lactobacillus rhamnosus GG Protects against Non-Alcoholic Fatty Liver Disease in Mice. PLoS ONE 2014, 9, e80169. [Google Scholar] [CrossRef]
- Lutgendorff, F.; Akkermans, L.; Soderholm, J. The Role of Microbiota and Probiotics in Stress-Induced Gastrointestinal Damage. Curr. Mol. Med. 2008, 8, 282–298. [Google Scholar] [CrossRef] [PubMed]
- Karl, P.J.; Hatch, A.M.; Arcidiacono, S.M.; Pearce, S.C.; Pantoja-Feliciano, I.G.; Doherty, L.A.; Soares, J.W. Effects of Psychological, Environmental and Physical Stressors on the Gut Microbiota. Front. Microbiol. 2018, 9, 2013. [Google Scholar] [CrossRef] [PubMed]
- Vandeputte, D.; Falony, G.; Vieira-Silva, S.; Tito, R.Y.; Joossens, M.; Raes, J. Stool Consistency Is Strongly Associated with Gut Microbiota Richness and Composition, Enterotypes and Bacterial Growth Rates. Gut 2016, 65, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.W.; Cheng, M.Y.; Yang, X.; Lu, Y.Y.; Yin, H.D.; Zeng, Y.; Wang, R.Y.; Jiang, Y.L.; Yang, W.T.; Wang, J.Z.; et al. Probiotic Lactobacillus rhamnosus GG Promotes Mouse Gut Microbiota Diversity and T Cell Differentiation. Front. Microbiol. 2020, 11, 607735. [Google Scholar] [CrossRef]
- Vital, M.; Karch, A.; Pieper, D.H. Colonic Butyrate-Producing Communities in Humans: An Overview Using Omics Data. mSystems 2017, 2, e00130-17. [Google Scholar] [CrossRef]
- Guilloteau, P.; Martin, L.; Eeckhaut, V.; Ducatelle, R.; Zabielski, R.; Van Immerseel, F. From the Gut to the Peripheral Tissues: The Multiple Effects of Butyrate. Nutr. Res. Rev. 2010, 23, 366–384. [Google Scholar] [CrossRef]
- Hamer, H.M.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, F.J.; Brummer, R.-J. Review Article: The Role of Butyrate on Colonic Function. Aliment. Pharmacol. Ther. 2008, 27, 104–119. [Google Scholar] [CrossRef]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The Role of Short-Chain Fatty Acids in Health and Disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar] [PubMed]
- Huang, C.; Song, P.; Fan, P.; Hou, C.; Thacker, P.; Ma, X. Dietary Sodium Butyrate Decreases Postweaning Diarrhea by Modulating Intestinal Permeability and Changing the Bacterial Communities in Weaned Piglets. J. Nutr. 2015, 145, 2774–2780. [Google Scholar] [CrossRef] [PubMed]
- Vernia, P.; Fracasso, P.L.; Casale, V.; Villotti, G.; Marcheggiano, A.; Stigliano, V.; Pinnaro, P.; Bagnardi, V.; Caprilli, R. Topical Butyrate for Acute Radiation Proctitis: Randomised, Crossover Trial. Lancet 2000, 356, 1232–1235. [Google Scholar] [CrossRef]
- Maggio, A.; Magli, A.; Rancati, T.; Fiorino, C.; Valvo, F.; Fellin, G.; Ricardi, U.; Munoz, F.; Cosentino, D.; Cazzaniga, L.F.; et al. Daily Sodium Butyrate Enema for the Prevention of Radiation Proctitis in Prostate Cancer Patients Undergoing Radical Radiation Therapy: Results of a Multicenter Randomized Placebo-Controlled Dose-Finding Phase 2 Study. Int. J. Radiat. Oncol. 2018, 89, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Haas, K.N.; Blanchard, J.L. Kineothrix alysoides, Gen. Nov., Sp. Nov., a Saccharolytic Butyrate-Producer within the Family Lachnospiraceae. Int. J. Syst. Evol. Microbiol. 2017, 67, 402–410. [Google Scholar] [CrossRef]
- Chen, Z.; Xie, Y.; Zhou, F.; Zhang, B.; Wu, J.; Zhang, X.; Xu, H.; Ren, J. Featured Gut Microbiomes Associated With the Progression of Chronic Hepatitis B Disease. Front Microbiol. 2020, 11, 383. [Google Scholar] [CrossRef]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C.J. Genes and Molecules of Lactobacilli Supporting Probiotic Action. Microbiol. Mol. Biol. Rev. 2008, 72, 728–764. [Google Scholar] [CrossRef]
- Machado, D.; Andrejev, S.; Tramontano, M.; Patil, K.R. Fast Automated Reconstruction of Genome-Scale Metabolic Models for Microbial Species and Communities. Nucleic Acids Res. 2018, 46, 7542–7553. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, Y.-J.; Yoon, J.Y.; Kemmitt, J.; Wright, C.; Schneider, K.; Sphabmixay, P.; Hernandez-Gordillo, V.; Holcomb, S.J.; Bhushan, B.; et al. Primary Human Colonic Mucosal Barrier Crosstalk with Super Oxygen-Sensitive Faecalibacterium Prausnitzii in Continuous Culture. Med 2021, 2, 74–98.e9. [Google Scholar] [CrossRef]
- Lebeer, S.; Claes, I.J.J.; Verhoeven, T.L.A.; Shen, C.; Lambrichts, I.; Ceuppens, J.L.; Vanderleyden, J.; De Keersmaecker, S.C.J. Impact of LuxS and Suppressor Mutations on the Gastrointestinal Transit of Lactobacillus rhamnosus GG. Appl. Environ. Microbiol. 2008, 74, 4711–4718. [Google Scholar] [CrossRef] [Green Version]
- Leeming, E.R.; Johnson, A.J.; Spector, T.D.; Roy, C.I.L. Effect of Diet on the Gut Microbiota: Rethinking Intervention Duration. Nutrients 2019, 11, 2862. [Google Scholar] [CrossRef] [PubMed]
- Meance, S.; Cayuela, C.; Raimondi, A.; Turchet, P.; Lucas, C.; Antoine, J.M. Recent Advances in the Use of Functional Foods: Effects of the Commercial Fermented Milk with Bifidobacterium Animalis Strain DN-173 010 and Yoghurt Strains on Gut Transit Time in the Elderly. Microb. Ecol. Health Dis. 2003, 15, 15–22. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Saline Group | L. indica PCC 8005 Group | L. rhamnosus GG Group | |
|---|---|---|---|
| Weight at start | 22.54 ± 0.62 | 22.94 ± 0.56 | 22.75 ± 0.92 |
| Weight gain during supplementation | 0.86 ± 0.43 | 0.81 ± 0.40 | 1.15 ± 0.39 a,b |
| Weight after supplementation | 23.40 ± 0.84 | 23.75 ± 0.81 | 23.89 ± 0.99 |
| Weight gain during washout | 0.66 ± 0.31 | 0.99 ± 0.32 | 0.73 ± 0.35 |
| Weight after washout | 24.35 ± 0.90 | 24.91 ± 0.92 | 24.78 ± 1.00 |
| Weight gain overall | 1.48 ± 0.35 | 1.80 ± 0.36 | 1.88 ± 0.24 a |
| Taxonomic Classification (Following Ribosomal Database Project) | ANCOM Biomarkers’ Effect Size and W-Statistic | Highest NCBI Blast Hit (% Identity) |
|---|---|---|
| Erysipelotrichaceae_OTU208 | 1.12–W = 0.9 (t4) | NA |
| Lachnospiraceae_OTU281 | 1.02–W = 0.9 (t4) | Kineothrix alysoides (~97% identity) |
| Lachnospiraceae_OTU528 | 1.35–W = 0.9 (t2) | NA |
| Lachnospiraceae_OTU23 | −1.26–W = 0.9 (t2) | NA |
| Turicibacter_OTU33 | −1.54–W = 0.9 (t2 and t4) | Turicibacter sanguinis (~97% identity) |
| Porphyromonadaceae_OTU446 | −1.14–W = 0.9 (t2) | NA |
| Taxonomic Classification (Following Ribosomal Database Project) | ANCOM Biomarkers’ Effect Size and W-Statistic | Highest NCBI Blast Hit (% Identity) |
|---|---|---|
| Lachnospiraceae_OTU152 | 1.06–W = 0.9 (t4) | NA |
| Porphyromonadaceae_OTU347 | 1.14–W = 0.9 (t4) | NA |
| Porphyromonadaceae_OTU446 | 1.04–W = 0.9 (t2 and t4) | NA |
| Porphyromonadaceae_OTU588 | 1.26–W = 0.9 (t4) | NA |
| Porphyromonadaceae_OTU963 | 1.09–W = 0.9 (t2) | NA |
| Porphyromonadaceae_OTU1645 | 1.05–W = 0.9 (t4) | NA |
| Anaerofustis_OTU284 | −1.06–W = 0.9 (t2) | Anaerofustis stercorihominis (~98% identity) |
| Lachnospiraceae_OTU375 | −1.33–W = 0.9 (t4) | NA |
| Bacteroidales_OTU1374 | −1.07–W = 0.8 (t4) | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Segers, C.; Mysara, M.; Coolkens, A.; Baatout, S.; Leys, N.; Lebeer, S.; Verslegers, M.; Mastroleo, F. Limnospira indica PCC 8005 or Lacticaseibacillus rhamnosus GG Dietary Supplementation Modulate the Gut Microbiome in Mice. Appl. Microbiol. 2022, 2, 636-650. https://doi.org/10.3390/applmicrobiol2030049
Segers C, Mysara M, Coolkens A, Baatout S, Leys N, Lebeer S, Verslegers M, Mastroleo F. Limnospira indica PCC 8005 or Lacticaseibacillus rhamnosus GG Dietary Supplementation Modulate the Gut Microbiome in Mice. Applied Microbiology. 2022; 2(3):636-650. https://doi.org/10.3390/applmicrobiol2030049
Chicago/Turabian StyleSegers, Charlotte, Mohamed Mysara, Amelie Coolkens, Sarah Baatout, Natalie Leys, Sarah Lebeer, Mieke Verslegers, and Felice Mastroleo. 2022. "Limnospira indica PCC 8005 or Lacticaseibacillus rhamnosus GG Dietary Supplementation Modulate the Gut Microbiome in Mice" Applied Microbiology 2, no. 3: 636-650. https://doi.org/10.3390/applmicrobiol2030049
APA StyleSegers, C., Mysara, M., Coolkens, A., Baatout, S., Leys, N., Lebeer, S., Verslegers, M., & Mastroleo, F. (2022). Limnospira indica PCC 8005 or Lacticaseibacillus rhamnosus GG Dietary Supplementation Modulate the Gut Microbiome in Mice. Applied Microbiology, 2(3), 636-650. https://doi.org/10.3390/applmicrobiol2030049