


Zat12 Gene Ameliorates Temperature Stress in Wheat Transgenics by Modulating the Antioxidant Defense System

Abstract
1. Introduction
2. Results
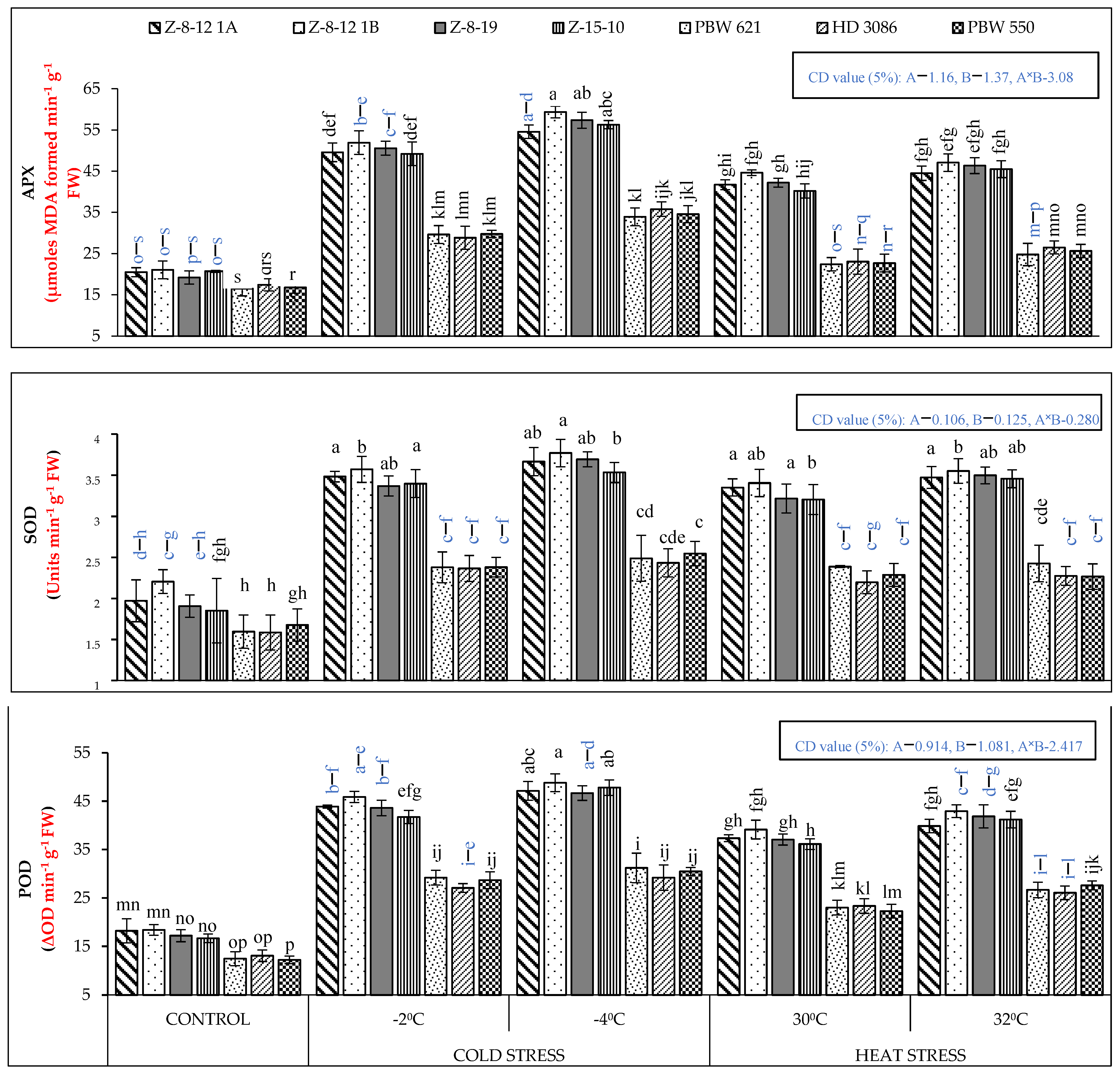
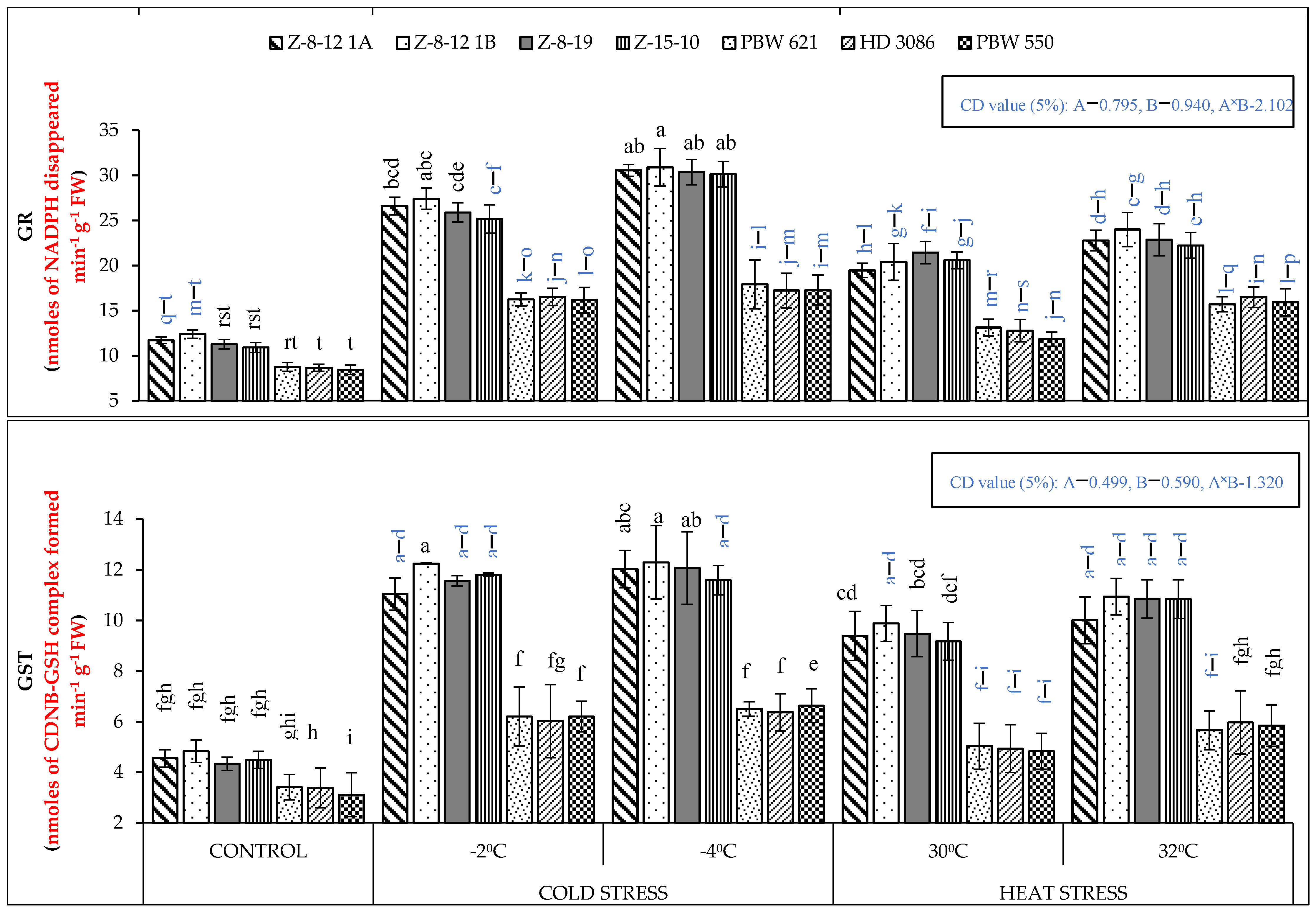
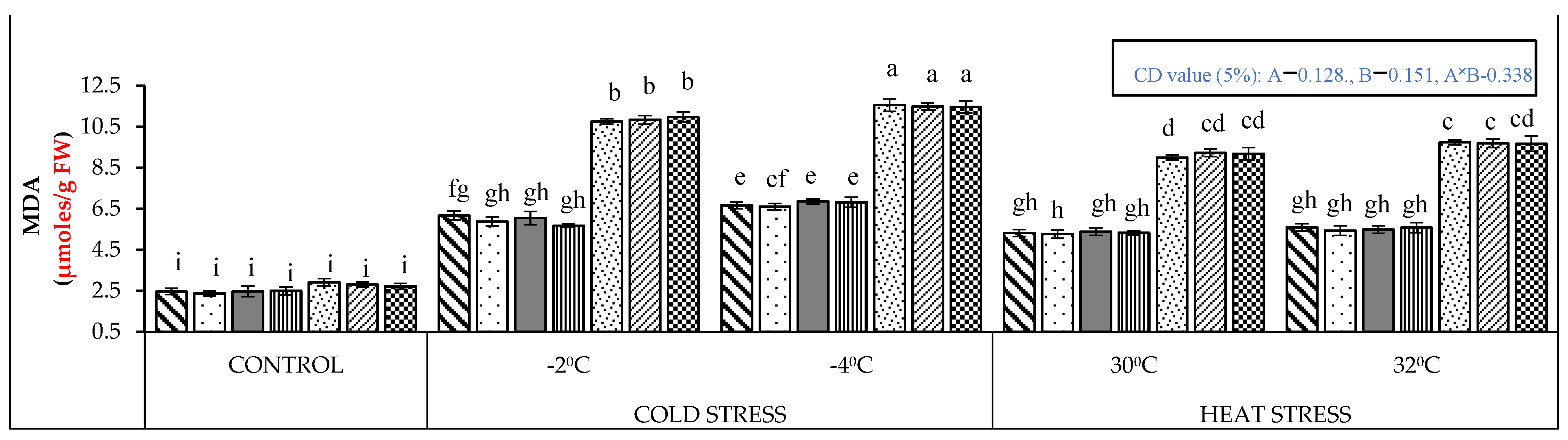
2.1. Biochemical Analysis
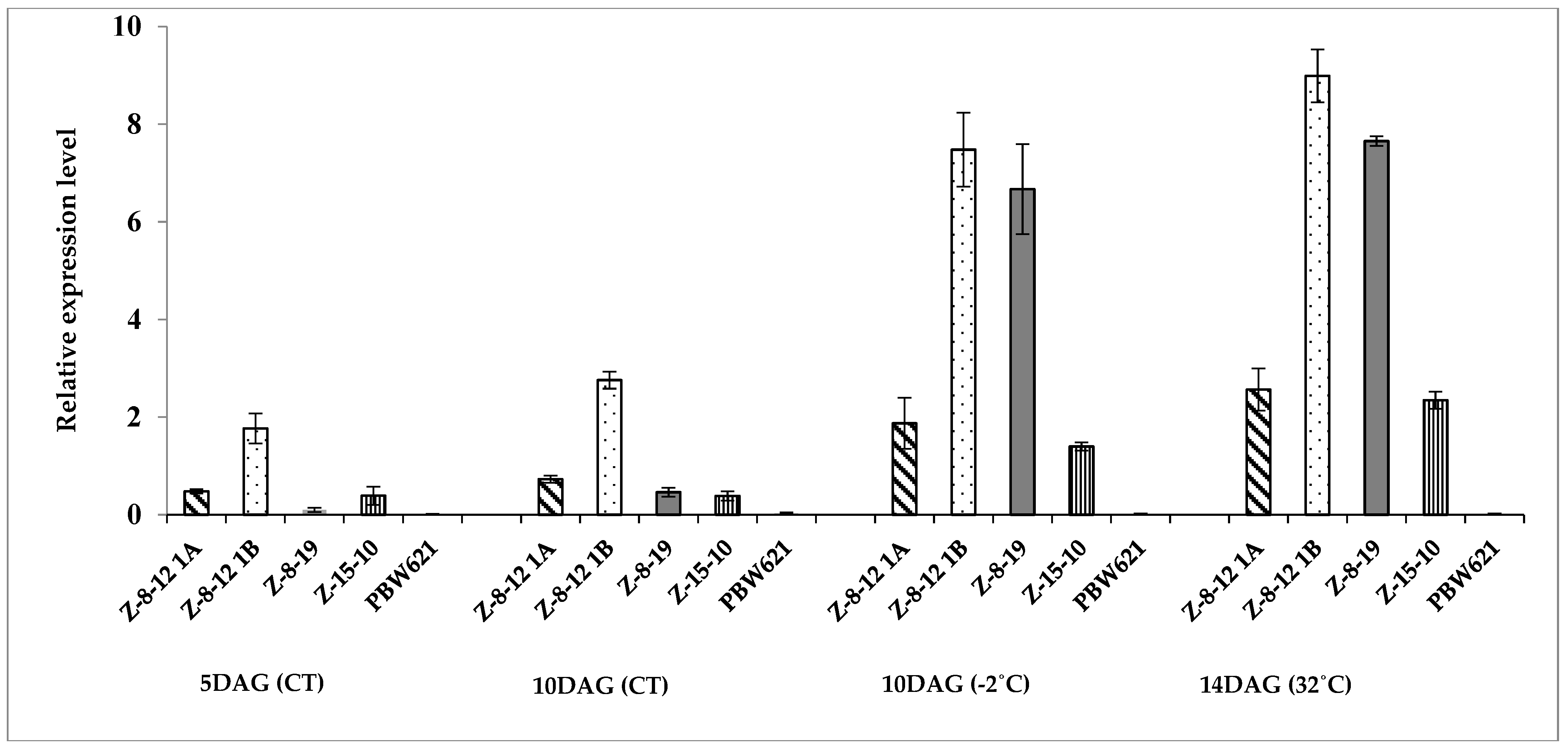
2.2. Molecular Analysis
2.3. Growth Parameter Analysis
3. Discussion
4. Materials and Methods
4.1. Material
4.2. Biochemical Analysis
4.3. Molecular Analysis
4.3.1. Genomic DNA Extraction
4.3.2. RT-PCR Analysis
4.4. Analysis of Growth Parameters
4.5. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Crop Prospects and Food Situation—Quarterly Global Report No. 2; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Poole, N.; Donovan, J.; Erenstein, O. Agri-nutrition research: Revisiting the contribution of maize and wheat to human nutrition and health. Food Policy 2021, 100, 101976. [Google Scholar] [CrossRef]
- Huang, M.; Mheni, N.; Brown-Guedira, G.; McKendry, A.; Grifey, C.; Van Sanford, D.; Costa, J.; Sneller, C. Genetic analysis of heading date in winter and spring wheat. Euphytica 2018, 214, 128. [Google Scholar] [CrossRef]
- Liu, B.; Martre, P.; Ewert, F.; Porter, J.R.; Challinor, A.J.; Müller, C.; Ruane, A.C.; Waha, K.; Thorburn, P.J.; Aggarwal, P.K.; et al. Global wheat production with 1.5 and 2.0 °C above pre-industrial warming. Glob. Chang. Biol. 2019, 4, 1428–1444. [Google Scholar] [CrossRef]
- Zheng, D.; Yang, X.; Mínguez, M.I.; Mu, C.; He, Q.; Wu, X. Effect of freezing temperature and duration on winter survival and grain yield of winter wheat. Agri. For. Meteorol. 2018, 260, 1–8. [Google Scholar] [CrossRef]
- Gupta, K.; Dey, A.; Gupta, B. Plant polyamines in abiotic stress responses. Acta Physiol. Plant. 2013, 35, 2015–2036. [Google Scholar] [CrossRef]
- Eulgem, T.; Rushton, P.; Robatzek, S.; Somssich, I. The WRKY superfamily of plant transcription factors. Trends Plant Sci. 2000, 5, 199–206. [Google Scholar] [CrossRef]
- Cham, L.T.T.; Thang, V.N.; Tuan, T.A. Expression of zinc finger protein ZAT12 from Arabidopsis thaliana in Escherichia coli. VJAS 2020, 3, 504–511. [Google Scholar] [CrossRef]
- Fowler, S.; Thomashow, M. Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway. Plant Cell 2002, 14, 1675–1690. [Google Scholar] [CrossRef]
- Bielen, A.; Remans, T.; Vangronsveld, J.; Cuypers, A. The role of ascorbate during cadmium-induced oxidative stress in Arabidopsis thaliana. Int. J. STD AIDS 2013, 14, 431–436. [Google Scholar]
- Iida, A.; Kazuoka, T.; Torikai, S.; Kikuchi, H.; Oeda, K. A zinc finger protein RHL41 mediates the light acclimatization response in Arabidopsis. Plant J. 2000, 24, 191–203. [Google Scholar] [CrossRef]
- Klok, E.J.; Wilson, I.W.; Wilson, D.; Chapman, S.C.; Ewing, R.M.; Somerville, S.C.; Peacock, W.J.; Dolferus, R.; Dennis, E.S. Expression profile analysis of the low-oxygen response in Arabidopsis root cultures. Plant Cell 2002, 14, 2481–2494. [Google Scholar] [CrossRef]
- Ciftci-Yilmaz, S.; Mittler, R. The zinc finger network of plants. Cell Mol. Life Sci. 2008, 65, 1150–1160. [Google Scholar] [CrossRef]
- Mittler, R.; Kim, Y.; Song, L.; Coutu, J.; Coutu, A.; Ciftci-Yilmaz, S.; Lee, H.; Stevenson, B.; Zhu, J. Gain- and loss-of-function mutations in Zat10 enhance the tolerance of plants to abiotic stress. FEBS Lett. 2006, 580, 6537–6542. [Google Scholar] [CrossRef]
- Davletova, S.; Rizhsky, L.; Liang, H.; Shengqiang, Z.; Oliver, D.J.; Coutu, J.; Shulaev, V.; Schlauch, K.; Mittler, R. Cytosolic ascorbate peroxidase 1 is a central component of the reactive oxygen gene network of Arabidopsis. Plant Cell 2005, 17, 268–281. [Google Scholar] [CrossRef]
- Vogel, J.T.; Zarka, D.G.; Van Buskirk, H.A.; Fowler, S.G.; Thomashow, M.F. Roles of the CBF2 and ZAT12 transcription factors in configuring the low-temperature transcriptome of Arabidopsis. Plant J. 2005, 41, 195–211. [Google Scholar] [CrossRef]
- Rai, A.C.; Singh, M.; Shah, K. Engineering drought tolerant tomato plant over-expressing BcZAT12 gene encoding a C2H2 zinc finger transcription factor. Phytochemistry 2013, 85, 44–50. [Google Scholar] [CrossRef]
- Zörb, C.; Geilfus, C.M.; Dietz, K.J. Salinity and crop yield. Plant Biol. 2019, 21, 31–38. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Fujita, M. Extreme temperature responses, oxidative stress and antioxidant defense in plants. In Abiotic Stress-Plant Responses and Applications in Agriculture; IntechOpen: London, UK, 2013; Volume 13, pp. 169–205. [Google Scholar] [CrossRef]
- Low, L.Y.; Yang, S.K.; Kok, D.X.; Ong-Abdullah, J.; Tan, N.P.; Lai, K.S. Transgenic Plants: Gene constructs, vector and transformation method. In New Visions in Plant Science; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef]
- Farooq, M.; Bramley, H.; Palta, J.A.; Siddique, K.H. Heat stress in wheat during reproductive and grain-filling phases. Crit. Rev. Plant Sci. 2011, 30, 491–507. [Google Scholar] [CrossRef]
- Anjum, N.A.; Duarte, A.C.; Pereira, E.; Ahmad, I. Juncus maritimus root biochemical assessment for its mercury stabilization potential in Ria de Aveiro coastal lagoon (Portugal). Environ. Sci. Pollut. Res. 2015, 22, 2231–2238. [Google Scholar] [CrossRef]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.J. The role of plant antioxidant system in drought tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef]
- Andrés, C.M.C.; Pérez de la Lastra, J.M.; Andrés Juan, C.; Plou, F.J.; Pérez-Lebeña, E. Superoxide anion chemistry—Its role at the core of the innate immunity. Int. J. Mol. Sci. 2023, 24, 1841. [Google Scholar] [CrossRef]
- Yap, C.K.; Tan, W.S.; Wong, K.W.; Ong, G.H.; Cheng, W.H.; Nulit, R.; Ibrahim, M.H.; Chew, W.; Edward, F.B.; Okamura, H.; et al. Antioxidant enzyme activities as biomarkers of Cu and Pb stress in Centella asiatica. Stresses 2021, 1, 253–265. [Google Scholar] [CrossRef]
- Bozca, F.D.; Leblebici, S. Interactive effect of boric acid and temperature stress on phenological characteristics and antioxidant system in Helianthus annuus L. S. Afr. J. Bot. 2022, 147, 391–399. [Google Scholar] [CrossRef]
- Guan, Q.; Wang, Z.; Wang, X.; Takano, T.; Liu, S. A peroxisomal APX from Puccinellia tenuiflora improves the abiotic stress tolerance of transgenic Arabidopsis thaliana through decreasing of H2O2 accumulation. J. Plant Physiol. 2015, 175, 183–191. [Google Scholar] [CrossRef]
- Ghosh, A.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. A glutathione responsive rice glyoxalase II, OsGLYII-2, function in salinity adaptation by maintaining better photosynthesis efficiency and anti-oxidant pool. Plant J. 2014, 80, 93–105. [Google Scholar] [CrossRef]
- Gill, S.S.; Anjum, N.A.; Hasanuzzaman, M.; Gill, R.; Trivedi, D.K.; Ahmad, I.; Pereira, E.; Tuteja, N. Glutathione and glutathione reductase: A boon in disguise for plant abiotic stress defense operations. Plant Physiol. Biochem. 2013, 70, 204–212. [Google Scholar] [CrossRef]
- Sairam, R.K.; Saxena, D.C. Oxidative stress and antioxidants in wheat genotypes: Possible mechanism of water stress tolerance. J. Agron. Crop Sci. 2000, 184, 55–61. [Google Scholar] [CrossRef]
- Sylvestre-Gonon, E.; Law, S.R.; Schwartz, M.; Robe, K.; Keech, O.; Didierjean, C.; Dubos, C.; Rouhier, N.; Hecker, A. Functional, structural and biochemical features of plant serinyl-glutathione transferases. Front. Plant Sci. 2019, 10, 608. [Google Scholar] [CrossRef]
- Gao, H.; Yu, C.; Liu, R.; Li, X.; Huang, H.; Wang, X.; Zhang, C.; Jiang, N.; Li, X.; Cheng, S.; et al. The glutathione s-transferase PtGSTF1 improves biomass production and salt tolerance through regulating xylem cell proliferation, ion homeostasis and reactive oxygen species scavenging in Poplar. Int. J. Mol. Sci. 2022, 23, 11288. [Google Scholar] [CrossRef]
- Hosseinifard, M.; Stefaniak, S.; Ghorbani Javid, M.; Soltani, E.; Wojtyla, Ł.; Garnczarska, M. Contribution of exogenous proline to abiotic stresses tolerance in plants: A Review. Int. J. Mol. Sci. 2022, 23, 5186. [Google Scholar] [CrossRef]
- Gęgotek, A.; Skrzydlewska, E. Antioxidative and anti-Inflammatory activity of ascorbic acid. Antioxidants 2022, 11, 1993. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.; Agnihotri, R.; Sharma, M.P.; Reddy, V.R.; Jajoo, A. Effect of high-temperature stress on plant physiological traits and mycorrhizal symbiosis in maize plants. J. Fungi 2021, 7, 867. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.M.; Doherty, S.J.; Croy, R.R.D. Biphasic superoxide generation in potato tubers: A self-amplifying response to stress. Plant Physiol. 2003, 131, 1440–1449. [Google Scholar] [CrossRef]
- Xu, J.; Zhou, Y.; Xu, Z.; Chen, Z.; Duan, L. Combining physiological and metabolic analysis to unravel the regulations of coronatine alleviating water stress in tobacco (Nicotiana tobacum L.). Biomolecules 2020, 10, 99. [Google Scholar] [CrossRef]
- Kumar, S.; Beena, A.S.; Awana, M.; Singh, A. Physiological, biochemical, epigenetic and molecular analyses of wheat (Triticum aestivum) genotypes with contrasting salt tolerance. Front. Plant Sci. 2017, 8, 1151. [Google Scholar] [CrossRef] [PubMed]
- Elkelish, A.; Qari, S.H.; Mazrou, Y.S.A.; Abdelaal, K.A.A.; Hafez, Y.M.; Abu-Elsaoud, A.M.; Batiha, G.E.-S.; El-Esawi, M.A.; El Nahhas, N. Exogenous ascorbic acid-induced chilling tolerance in tomato plants through modulating metabolism, osmolytes, antioxidants, and transcriptional regulation of catalase and heat shock proteins. Plants 2020, 9, 431. [Google Scholar] [CrossRef] [PubMed]
- Davletova, S.; Schlauch, K.; Coutu, J.; Mittler, R. The zinc-finger protein Zat12 plays a central role in reactive oxygen and abiotic stress signaling in Arabidopsis. Plant Physiol. 2005, 139, 847–856. [Google Scholar] [CrossRef]
- Rizhsky, L.; Liang, H.; Shuman, J.; Shulaev, V.; Davletova, S.; Mittler, R. When defense pathways collide: The response of Arabidopsis to a combination of drought and heat stress. Plant Physiol. 2004, 134, 1683–1696. [Google Scholar] [CrossRef]
- Negrao, S.; Schmockel, S.M.; Tester, M. Evaluating physiological responses of plants to salinity stress. Ann. Bot. 2017, 119, 1–11. [Google Scholar] [CrossRef]
- Kaur, R. Genetic Transformation of Bread Wheat (Triticum aestivum L.) by Particle Gun and Agrobacterium-Mediated Approaches. Ph.D. Dissertation, Punjab Agricultural University, Ludhiana, India, 2014. [Google Scholar]
- Gamborg, O.L.; Murashige, T.; Thorpe, T.A.; Vasil, I.K. Plant tissue culture media. In Vitro 1976, 12, 473–478. [Google Scholar] [CrossRef]
- Nankano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate peroxidase in spinach chloroplasts. Plant Cell Physiol. 1974, 22, 867–880. [Google Scholar]
- Markuland, S.; Marklund, G. Involvement of superoxide anion radical in the autooxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 169–174. [Google Scholar]
- Shannon, L.; Kay, E.; Law, J. Peroxidase isozymes from horseradish roots: Isolation and physical properties. J. Biol. Chem. 1966, 241, 2166–2172. [Google Scholar] [CrossRef]
- Goyal, M.; Asthir, B. Polyamine catabolism influences antioxidative defense mechanism in shoots and roots of five wheat genotypes under high-temperature stress. J. Plant Growth Regul. 2010, 60, 13–25. [Google Scholar] [CrossRef]
- Habig, W.; Pabst, M.; Jakoby, W. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef] [PubMed]
- Bates, L. Rapid determination of free proline content for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Law, M.; Charles, S.; Halliwell, B. Glutathione and ascorbic acid on spinach (Spinacia oleracea) Chloroplasts: The effect of hydrogen peroxide and paraquat. J. Biochem. 1983, 210, 889–903. [Google Scholar] [CrossRef]
- Patterson, B.D. Estimation of hydrogen peroxide in plant extracts using titanium (IV). Anal. Biochem. 1984, 139, 487–492. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Saghai-Maroof, M.A.; Biyashev, R.M.; Yang, G.R.; Zhang, Q.; Allard, R.W. Extraordinarily polymorphic microsatellite DNA in barley species diversity, chromosomal locations and population dynamics. Proc. Natl. Acad. Sci. USA 1984, 91, 5466–5547. [Google Scholar] [CrossRef]
- Wilkins, T.; Smart, L. Isolation of RNA from plant tissue. A Lab. Guide RNA Isol. Anal. Synth. 1996, 43, 21–42. [Google Scholar]
- Sheoran, O.P.; Tonk, D.S.; Kaushik, L.S.; Hasija, R.C.; Pannu, R.S. Statistical Software Package for Agricultural Research Workers. Recent Advances in Information Theory, Statistics & Computer Applications; Hooda, D.S., Hasija, R.C., Eds.; Department of Mathematics Statistics, CCS HAU: Hisar, India, 1998; pp. 139–143. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | APX | SOD | POD | GR | GST | PRO | ASC | H2O2 | MDA |
|---|---|---|---|---|---|---|---|---|---|
| Factor A (Temperature) | |||||||||
| 20 °C | 18.882 | 3.104 | 35.810 | 21.518 | 9.156 | 1.948 | 62.732 | 1.013 | 4.864 |
| −2 °C | 41.361 | 3.273 | 39.343 | 23.488 | 10.081 | 2.158 | 66.667 | 1.048 | 5.393 |
| −4 °C | 47.402 | 2.935 | 33.981 | 20.137 | 8.882 | 1.828 | 61.278 | 0.963 | 4.891 |
| 30 °C | 33.852 | 2.297 | 25.629 | 15.025 | 5.595 | 1.123 | 51.852 | 1.635 | 9.407 |
| 32 °C | 37.182 | 2.234 | 24.371 | 14.083 | 5.360 | 1.087 | 50.201 | 1.509 | 8.953 |
| CD (5%) | 1.166 | 0.106 | 0.914 | 0.795 | 0.499 | 0.154 | 1.761 | 0.042 | 0.128 |
| Factor B (Lines/Cultivar) | |||||||||
| Z-8-12 1A | 42.165 | 2.764 | 32.280 | 18.889 | 7.767 | 1.576 | 58.321 | 1.273 | 6.882 |
| Z-8-12 1B | 44.808 | 2.702 | 31.387 | 18.501 | 7.577 | 1.658 | 57.801 | 1.264 | 6.998 |
| Z-8-19 | 43.137 | 2.937 | 33.400 | 20.185 | 8.585 | 1.773 | 60.540 | 1.148 | 5.882 |
| Z-15-10 | 42.379 | 2.721 | 31.533 | 18.581 | 7.435 | 1.582 | 58.636 | 1.263 | 7.012 |
| PBW 621 | 25.452 | 2.837 | 32.627 | 18.995 | 8.151 | 1.693 | 59.419 | 1.153 | 6.183 |
| HD 3086 | 26.323 | 2.802 | 31.120 | 18.620 | 7.718 | 1.570 | 58.395 | 1.269 | 6.932 |
| PBW 550 | 25.894 | 2.681 | 30.440 | 18.180 | 7.472 | 1.549 | 56.708 | 1.265 | 7.022 |
| CD (5%) | 1.379 | 0.125 | 1.081 | 0.940 | 0.590 | 0.175 | 2.084 | 0.050 | 0.151 |
| Treatment | RL | SL | RFW | SFW | RDW | SDW |
|---|---|---|---|---|---|---|
| Factor A (Temperature) | ||||||
| 20 °C | 12.343 | 14.570 | 146.833 | 184.467 | 29.940 | 32.347 |
| 30 °C | 11.753 | 13.700 | 140.827 | 176.067 | 27.933 | 31.860 |
| 32 °C | 10.940 | 12.539 | 135.433 | 165.533 | 24.540 | 30.233 |
| CD (5%) | 0.188 | 0.108 | 1.171 | 1.083 | 0.656 | 0.862 |
| Factor B (Lines/Cultivar) | ||||||
| Z-8-12 1A | 11.694 | 13.967 | 142.544 | 178.278 | 29.217 | 32.667 |
| Z-8-12 1B | 11.667 | 13.650 | 142.333 | 177.389 | 27.811 | 31.578 |
| Z-8-19 | 11.522 | 13.564 | 141.389 | 176.500 | 27.389 | 30.989 |
| Z-15-10 | 11.489 | 13.244 | 138.111 | 169.722 | 26.556 | 31.111 |
| PBW 621 | 12.022 | 13.589 | 140.778 | 174.889 | 26.383 | 31.056 |
| CD (5%) | 0.243 | 0.139 | 1.512 | 1.398 | 0.847 | 1.113 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaur, M.; Asthir, B.; Kaur, R.; Chaudhary, A. Zat12 Gene Ameliorates Temperature Stress in Wheat Transgenics by Modulating the Antioxidant Defense System. Stresses 2023, 3, 316-330. https://doi.org/10.3390/stresses3010023
Kaur M, Asthir B, Kaur R, Chaudhary A. Zat12 Gene Ameliorates Temperature Stress in Wheat Transgenics by Modulating the Antioxidant Defense System. Stresses. 2023; 3(1):316-330. https://doi.org/10.3390/stresses3010023
Chicago/Turabian StyleKaur, Manpreet, Bavita Asthir, Ramandeep Kaur, and Ankur Chaudhary. 2023. "Zat12 Gene Ameliorates Temperature Stress in Wheat Transgenics by Modulating the Antioxidant Defense System" Stresses 3, no. 1: 316-330. https://doi.org/10.3390/stresses3010023
APA StyleKaur, M., Asthir, B., Kaur, R., & Chaudhary, A. (2023). Zat12 Gene Ameliorates Temperature Stress in Wheat Transgenics by Modulating the Antioxidant Defense System. Stresses, 3(1), 316-330. https://doi.org/10.3390/stresses3010023