
Analyzing the Effects of Urban Photopollution on Photosynthetic Efficiency of Certain Trees through Chlorophyll Fluorescence OJIP Transient




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
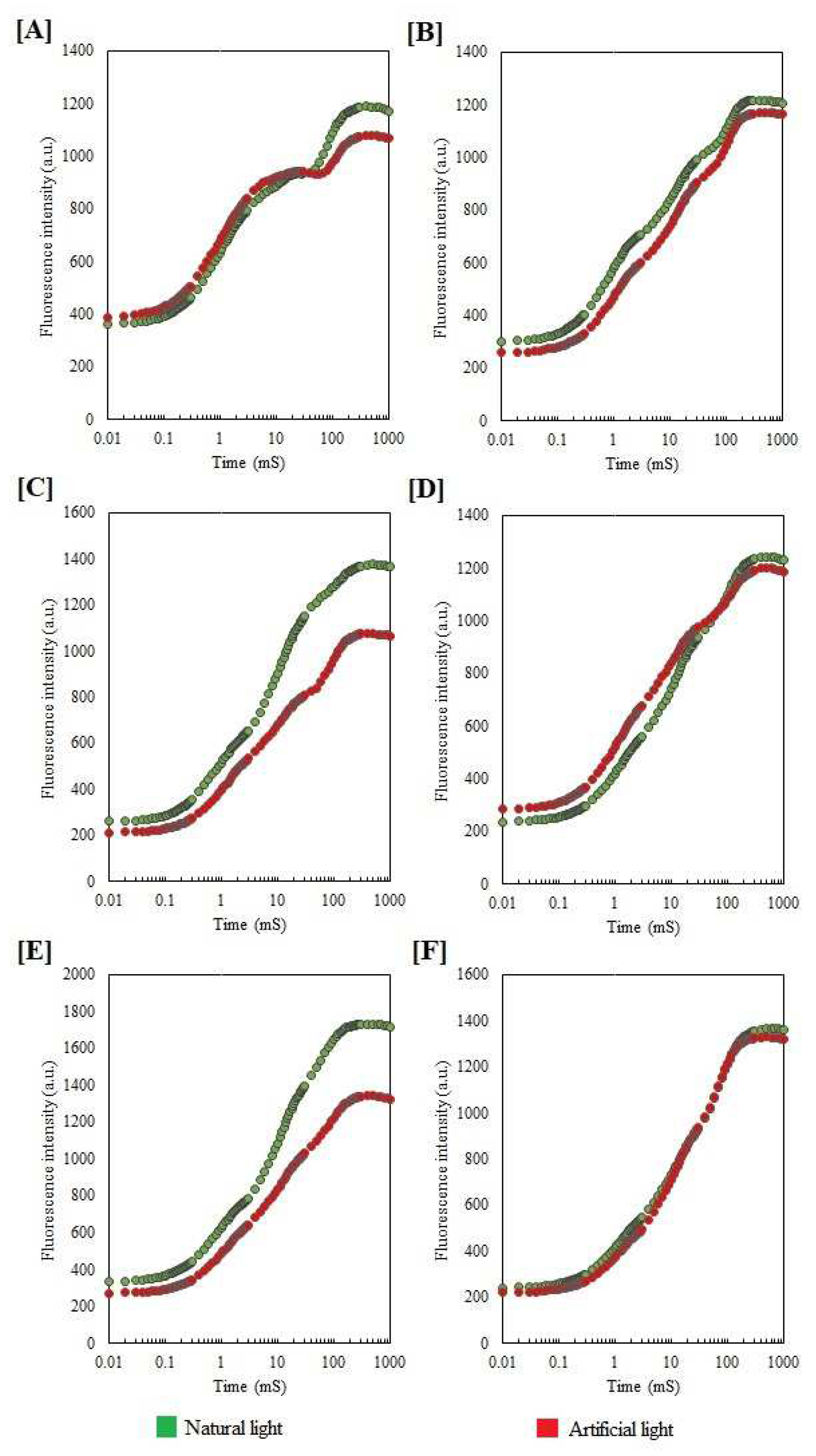
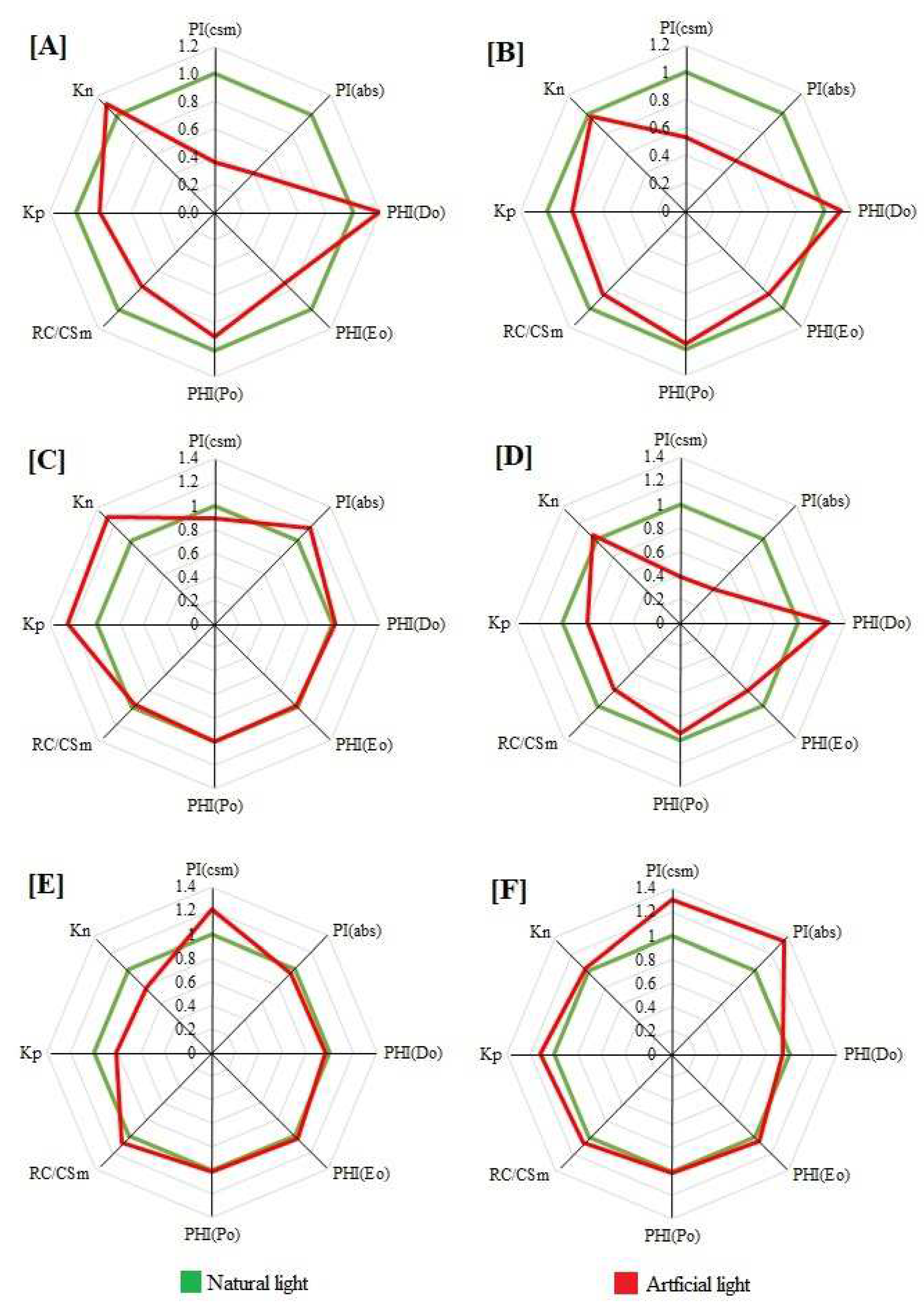
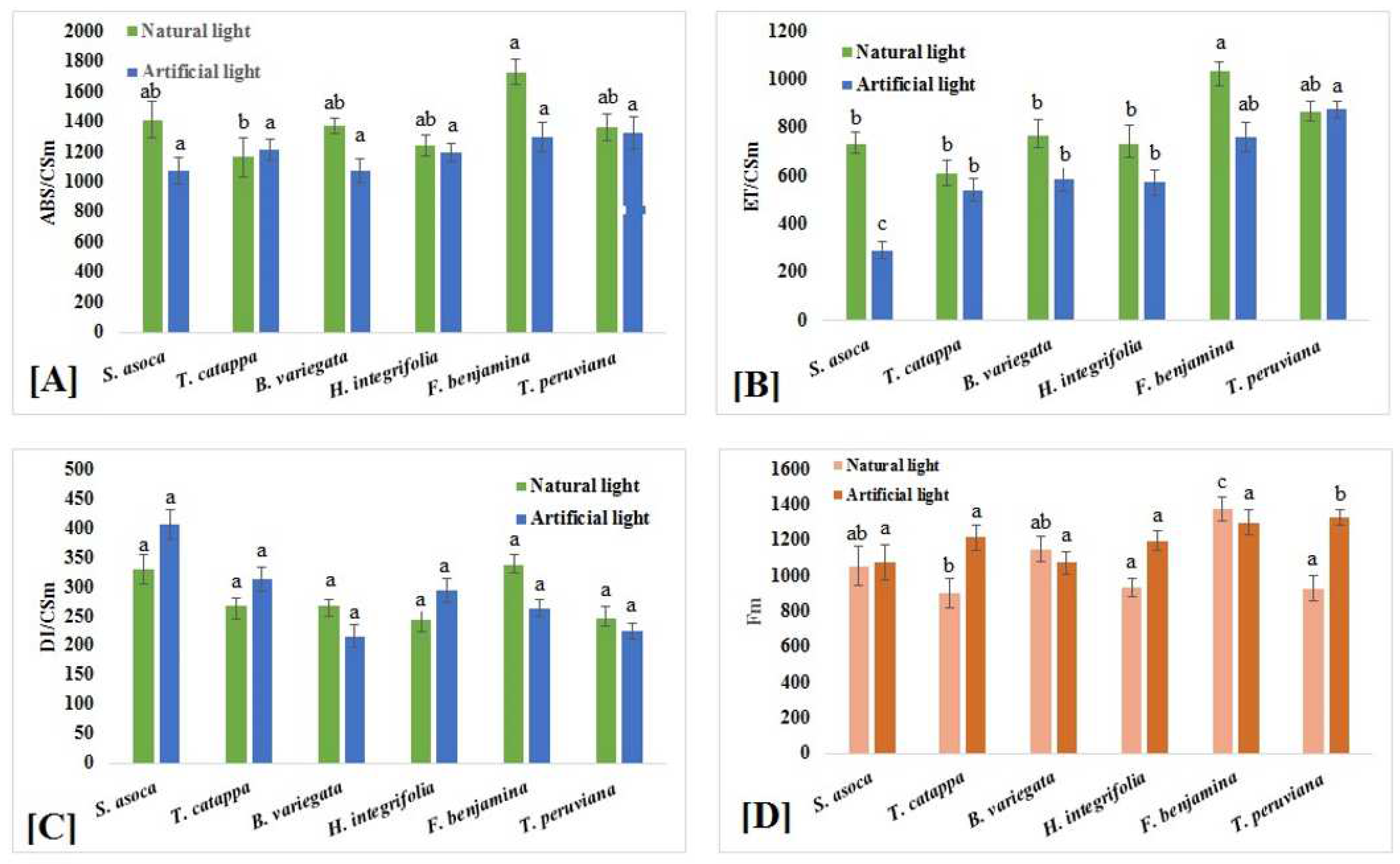
2. Results
2.1. Chlorophyll a Fluorescence
2.2. Density of Active Reaction Centers
2.3. Yield and Flux Ratio
2.4. Performance Index
2.5. Phenomenological Energy Fluxes
3. Discussion
4. Materials and Methods
4.1. Site Description
4.2. Plant Selection
4.3. Measurement of Chlorophyll a Fluorescence
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Butchart, S.H.M.; Walpole, M.; Collen, B.; Van Strien, A.; Scharlemann, J.P.W.; Almond, R.E.A.; Baillie, J.E.M.; Bomhard, B.; Brown, C.; Bruno, J.; et al. Global Biodiversity: Indicators of Recent Declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef] [PubMed]
- Lian, X.; Jiao, L.; Zhong, J.; Jia, Q.; Liu, J.; Liu, Z. Artificial Light Pollution Inhibits Plant Phenology Advance Induced by Climate Warming. Environ. Pollut. 2021, 291, 118110. [Google Scholar] [CrossRef] [PubMed]
- Longcore, T.; Rich, C. Ecological Light Pollution. Front. Ecol. Environ. 2004, 2, 191–198. [Google Scholar] [CrossRef]
- Hölker, F.; Wolter, C.; Perkin, E.K.; Tockner, K. Light Pollution as a Biodiversity Threat. Trends Ecol. Evol. 2010, 25, 681–682. [Google Scholar] [CrossRef] [PubMed]
- Orzechowska, A.; Trtílek, M.; Niewiadomska, E. Thermographic Study of Plant Response to Excessive Light. Acta Phys. Pol. Ser. A 2021, 139, 257–260. [Google Scholar] [CrossRef]
- Crump, M.C.; Brown, C.; Griffin-Nolan, R.J.; Angeloni, L.; Lemoine, N.P.; Seymoure, B.M. Effects of Low-Level Artificial Light at Night on Kentucky Bluegrass and an Introduced Herbivore. Front. Ecol. Evol. 2021, 9, 732959. [Google Scholar] [CrossRef]
- Dominoni, D.M.; Nelson, R.J. Artificial Light at Night as an Environmental Pollutant: An Integrative Approach across Taxa, Biological Functions, and Scientific Disciplines. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2018, 329, 387. [Google Scholar] [CrossRef]
- Meravi, N.; Prajapati, S.K. Effect Street Light Pollution on the Photosynthetic Efficiency of Different Plants. Biol. Rhythm. Res. 2020, 51, 67–75. [Google Scholar] [CrossRef]
- Wang, H.; Gu, M.; Cui, J.; Shi, K.; Zhou, Y.; Yu, J. Effects of Light Quality on CO2 Assimilation, Chlorophyll-Fluorescence Quenching, Expression of Calvin Cycle Genes and Carbohydrate Accumulation in Cucumis Sativus. J. Photochem. Photobiol. B Biol. 2009, 96, 30–37. [Google Scholar] [CrossRef]
- Zha, L.; Liu, W.; Zhang, Y.; Zhou, C.; Shao, M. Morphological and Physiological Stress Responses of Lettuce to Different Intensities of Continuous Light. Front. Plant Sci. 2019, 10, 1440. [Google Scholar] [CrossRef]
- Devlin, P.F.; Christie, J.M.; Terry, M.J. Many Hands Make Light Work. J. Exp. Bot. 2007, 58, 3071–3077. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Li, P.; Wu, Y. Effects of Different Light Intensities on Chlorophyll Fluorescence Characteristics and Yield in Lettuce. Sci. Hortic. 2012, 135, 45–51. [Google Scholar] [CrossRef]
- Kumar, D.; Singh, H.; Bhatt, U.; Soni, V.; Allakhverdiev, S. Effect of Continuous Light on Antioxidant Activity, Lipid Peroxidation, Proline and Chlorophyll Content in Vigna radiata L. Funct. Plant Biol. 2021, 49, 145–154. [Google Scholar] [CrossRef]
- Liu, P.; Cao, B.; Wang, Y.; Wei, Z.; Ye, J.; Wei, H. Spectral Effect of Streetlamps on Urban Trees: A Simulated Study on Tissue Water, Nitrogen, and Carbohydrate Contents in Maple and Oak. PLoS ONE 2021, 16, e0248463. [Google Scholar] [CrossRef] [PubMed]
- Müller, P.; Li, X.-P.; Niyogi, K.K. Non-Photochemical Quenching. A Response to Excess Light Energy. Plant Physiol. 2001, 125, 1558–1566. [Google Scholar] [CrossRef]
- Bennie, J.; Davies, T.W.; Cruse, D.; Gaston, K.J. Ecological Effects of Artificial Light at Night on Wild Plants. J. Ecol. 2016, 104, 611–620. [Google Scholar] [CrossRef]
- Briggs, W.R. Physiology of Plant Responses to Artificial Lighting. In Ecological Consequences of Artificial Night Lighting; Island Press: Washington, DC, USA, 2006; pp. 389–411. [Google Scholar]
- Massetti, L. Assessing the Impact of Street Lighting on Platanus x Acerifolia Phenology. Urban For. Urban Green. 2018, 34, 71–77. [Google Scholar] [CrossRef]
- Niyogi, K.K. Photoprotection Revisited: Genetic and Molecular Approaches. Annu. Rev. Plant Biol. 1999, 50, 333–359. [Google Scholar] [CrossRef]
- Vollsnes, A.V.; Eriksen, A.B.; Otterholt, E.; Kvaal, K.; Oxaal, U.; Futsaether, C.M. Visible Foliar Injury and Infrared Imaging Show that Daylength Affects Short-Term Recovery after Ozone Stress in Trifolium subterraneum. J. Exp. Bot. 2009, 60, 3677–3686. [Google Scholar] [CrossRef]
- Viera-Pérez, M.; Hernández-Calvento, L.; Hesp, P.A.; Santana-del Pino, A. Effects of Artificial Light on Flowering of Foredune Vegetation. Ecology 2019, 100, e02678. [Google Scholar] [CrossRef]
- Han, B.-H.; Kim, J.-Y.; Kwak, J.-I.; Choi, T.-Y. Correlation between the Illuminance and the Flowering and Leaf Growth of Trees at Night1a. Korean J. Environ. Ecol. 2015, 29, 441–453. [Google Scholar] [CrossRef]
- Pataki, D.E.; Alberti, M.; Cadenasso, M.L.; Felson, A.J.; McDonnell, M.J.; Pincetl, S.; Pouyat, R.V.; Setälä, H.; Whitlow, T.H. The Benefits and Limits of Urban Tree Planting for Environmental and Human Health. Front. Ecol. Evol. 2021, 9, 603757. [Google Scholar] [CrossRef]
- Gajdošik, M.S.; Vicić, A.; Gvozdić, V.; Galić, V.; Begović, L.; Mlinarić, S. Effect of Prolonged Photoperiod on Light-Dependent Photosynthetic Reactions in Cannabis. Int. J. Mol. Sci. 2022, 23, 9702. [Google Scholar] [CrossRef] [PubMed]
- Hey, M.H.; DiBiase, E.; Roach, D.A.; Carr, D.E.; Haynes, K.J. Interactions between Artificial Light at Night, Soil Moisture, and Plant Density Affect the Growth of a Perennial Wildflower. Oecologia 2020, 193, 503–510. [Google Scholar] [CrossRef]
- Anic, V.; Gaston, K.J.; Davies, T.W.; Bennie, J. Long-Term Effects of Artificial Nighttime Lighting and Trophic Complexity on Plant Biomass and Foliar Carbon and Nitrogen in a Grassland Community. Ecol. Evol. 2022, 12, e9157. [Google Scholar] [CrossRef]
- Segrestin, J.; Mondy, N.; Boisselet, C.; Guigard, L.; Lengagne, T.; Poussineau, S.; Secondi, J.; Puijalon, S. Effects of Artificial Light at Night on the Leaf Functional Traits of Freshwater Plants. Freshw. Biol. 2021, 66, 2264–2271. [Google Scholar] [CrossRef]
- Umar, M.; Uddin, Z.; Siddiqui, Z.S. Responses of Photosynthetic Apparatus in Sunflower Cultivars to Combined Drought and Salt Stress. Photosynthetica 2019, 57, 627–639. [Google Scholar] [CrossRef]
- Logan, B.A. Chlorophyll a Fluorescence: A Signature of Photosynthesis. J. Torrey Bot. Soc. 2005, 132, 650. [Google Scholar] [CrossRef]
- Lagorio, M.G. Chlorophyll Fluorescence Emission Spectra in Photosynthetic Organisms. In Chlorophyll: Structure, Production and Medicinal Uses; Nova Science Publishers: New York, NY, USA, 2011; pp. 115–150. [Google Scholar]
- Bucher, S.F.; Bernhardt-Römermann, M.; Römermann, C. Chlorophyll Fluorescence and Gas Exchange Measurements in Field Research: An Ecological Case Study. Photosynthetica 2018, 56, 1161–1170. [Google Scholar] [CrossRef]
- Moustakas, M.; Calatayud, Á.; Guidi, L. Chlorophyll Fluorescence Imaging Analysis in Biotic and Abiotic Stress. Front. Plant Sci. 2021, 12, 658500. [Google Scholar] [CrossRef]
- Tsimilli-Michael, M.; Strasser, R.J. In Vivo Assessment of Stress Impact on Plant’s Vitality: Applications in Detecting and Evaluating the Beneficial Role of Mycorrhization on Host Plants. In Mycorrhiza; Springer: Berlin/Heidelberg, Germany, 2008; pp. 679–703. [Google Scholar]
- Clark, A.J.; Landolt, W.; Bucher, J.B.; Strasser, R.J. Beech (Fagus sylvatica) Response to Ozone Exposure Assessed with a Chlorophyll a Fluorescence Performance Index. Environ. Pollut. 2000, 109, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Strasser, B.J. Donor Side Capacity of Photosystem II Probed by Chlorophyll a Fluorescence Transients. Photosynth. Res. 1997, 52, 147–155. [Google Scholar] [CrossRef]
- Tsimilli-Michael, M.; Strasser, R.J. The Energy Flux Theory 35 Years Later: Formulations and Applications. Photosynth. Res. 2013, 117, 289–320. [Google Scholar] [CrossRef] [PubMed]
- Falcón, J.; Torriglia, A.; Attia, D.; Viénot, F.; Gronfier, C.; Behar-Cohen, F.; Martinsons, C.; Hicks, D. Exposure to Artificial Light at Night and the Consequences for Flora, Fauna, and Ecosystems. Front. Neurosci. 2020, 14, 602796. [Google Scholar] [CrossRef]
- Chaney, W.R. Does Night Lighting Harm Trees. Purdue Univ. For. Nat. Resour. FAQ 2002, 17, 1–4. [Google Scholar]
- Škvareninová, J.; Tuhárska, M.; Škvarenina, J.; Babálová, D.; Hana, S.; Mária, T.; Jaroslav, Š.; Jana, Š. Effects of Light Pollution on Tree Phenology in the Urban Environment. Morav. Geogr. Rep. 2017, 25, 282–290. [Google Scholar] [CrossRef]
- Singhal, R.K.; Kumar, M.; Bose, B. Eco-Physiological Responses of Artificial Night Light Pollution in Plants. Russ. J. Plant Physiol. 2019, 66, 190–202. [Google Scholar] [CrossRef]
- Sysoeva, M.I.; Markovskaya, E.F.; Shibaeva, T.G. Plants under Continuous Light: A Review. Plant Stress 2010, 4, 5–17. [Google Scholar]
- Luo, H.; Merope, T.; Zhang, Y.; Zhang, W. Combining Gas Exchange and Chlorophyll a Fluorescence Measurements to Analyze the Photosynthetic Activity of Drip-Irrigated Cotton under Different Soil Water Deficits. J. Integr. Agric. 2016, 15, 1256–1266. [Google Scholar] [CrossRef]
- Strasser, R.J.; Stirbet, A.D. Heterogeneity of Photosystem II Probed by the Numerically Simulated Chlorophyll a Fluorescence Rise (O–J–I–P). Math. Comput. Simul. 1998, 48, 3–9. [Google Scholar] [CrossRef]
- Ranjbar, F.A.; Dehghani, B.R. Impact of Salinity Stress on Photochemical Efficiency of Photosystem Ii, Chlorophyll Content and Nutrient Elements of Nitere Bush (Nitraria schoberi L.) Plants. J. Rangel. Sci. 2016, 6, 1–9. [Google Scholar]
- Schreiber, U.; Bilger, W.; Neubauer, C. Chlorophyll Fluorescence as a Nonintrusive Indicator for Rapid Assessment of In Vivo Photosynthesis. In Ecophysiology of Photosynthesis; Springer: Berlin/Heidelberg, Germany, 1995. [Google Scholar]
- Schansker, G.; Srivastava, A.; Strasser, R.J. Characterization of the 820-Nm Transmission Signal Paralleling the Chlorophyll a Fluorescence Rise (OJIP) in Pea Leaves. Funct. Plant Biol. 2003, 30, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Goltsev, V.; Zaharieva, I.; Chernev, P.; Strasser, R.J. Delayed Fluorescence in Photosynthesis. Photosynth. Res. 2009, 101, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Liu, W.; Wang, X.; Yang, L.; Han, S.; Chen, S.; Strasser, R.J.; Valverde, B.E.; Qiang, S. Comparative Phytotoxicity of Usnic Acid, Salicylic Acid, Cinnamic Acid and Benzoic Acid on Photosynthetic Apparatus of Chlamydomonas Reinhardtii. Plant Physiol. Biochem. 2018, 128, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Goltsev, V.; Zaharieva, I.; Lambrev, P.; Yordanov, I.; Strasser, R. Simultaneous Analysis of Prompt and Delayed Chlorophyll a Fluorescence in Leaves during the Induction Period of Dark to Light Adaptation. J. Theor. Biol. 2003, 225, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.J.; Stirbet, A.D. Estimation of the Energetic Connectivity of PS II Centres in Plants Using the Fluorescence Rise O–J–I–P: Fitting of Experimental Data to Three Different PS II Models. Math. Comput. Simul. 2001, 56, 451–462. [Google Scholar] [CrossRef]
- Solovchenko, A.; Solovchenko, O.; Khozin-Goldberg, I.; Didi-Cohen, S.; Pal, D.; Cohen, Z.; Boussiba, S. Probing the Effects of High-Light Stress on Pigment and Lipid Metabolism in Nitrogen-Starving Microalgae by Measuring Chlorophyll Fluorescence Transients: Studies with a Δ5 Desaturase Mutant of Parietochloris incisa (Chlorophyta, Trebouxiophyceae). Algal Res. 2013, 2, 175–182. [Google Scholar] [CrossRef]
- Balarinová, K.; Barták, M.; Hazdrová, J.; Hájek, J.; Jílková, J. Changes in Photosynthesis, Pigment Composition and Glutathione Contents in Two Antarctic Lichens during a Light Stress and Recovery. Photosynthetica 2014, 52, 538–547. [Google Scholar] [CrossRef]
- Kono, M.; Noguchi, K.; Terashima, I. Roles of the Cyclic Electron Flow around PSI (CEF-PSI) and O2-Dependent Alternative Pathways in Regulation of the Photosynthetic Electron Flow in Short-Term Fluctuating Light in Arabidopsis Thaliana. Plant Cell Physiol. 2014, 55, 990–1004. [Google Scholar] [CrossRef]
- Hamdani, S.; Khan, N.; Perveen, S.; Qu, M.; Jiang, J.; Zhu, X.-G. Changes in the Photosynthesis Properties and Photoprotection Capacity in Rice (Oryza sativa) Grown under Red, Blue, or White Light. Photosynth. Res. 2019, 139, 107–121. [Google Scholar] [CrossRef]
- Stirbet, A. On the Relation between the Kautsky Effect (Chlorophyll a Fluorescence Induction) and Photosystem II: Basics and Applications of the OJIP Fluorescence Transient. J. Photochem. Photobiol. B Biol. 2011, 104, 236–257. [Google Scholar] [CrossRef]
- Yao, X.C.; Tu, H.Q.; Wang, X.L.; Wang, J. The Effect of Supplemental LED Night Lighting on the Growth and Physiology of the Para Rubber Tree. J. Rubber Res. 2021, 24, 321–326. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Bosa, K.; Kościelniak, J.; Hossain, Z. Chlorophyll a Fluorescence—A Useful Tool for the Early Detection of Temperature Stress in Spring Barley (Hordeum vulgare L.). OMICS J. Integr. Biol. 2011, 15, 925–934. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Qiang, S.; Goltsev, V. Simultaneous in Vivo Recording of Prompt and Delayed Fluorescence and 820-Nm Reflection Changes during Drying and after Rehydration of the Resurrection Plant Haberlea Rhodopensis. Biochim. Biophys. Acta Bioenerg. 2010, 1797, 1313–1326. [Google Scholar] [CrossRef] [PubMed]
- Tóth, S.Z.; Schansker, G.; Strasser, R.J. A Non-Invasive Assay of the Plastoquinone Pool Redox State Based on the OJIP-Transient. Photosynth. Res. 2007, 93, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.M.; Vyas, D.K.; Sher, A.A.; Grenis, K. Light Pollution Affects Invasive and Native Plant Traits Important to Plant Competition and Herbivorous Insects. Biol. Invasions 2022, 24, 599–602. [Google Scholar] [CrossRef]
- Kwak, M.J.; Je, S.M.; Cheng, H.C.; Seo, S.M.; Park, J.H.; Baek, S.G.; Khaine, I.; Lee, T.; Jang, J.; Li, Y.; et al. Night Light-Adaptation Strategies for Photosynthetic Apparatus in Yellow-Poplar (Liriodendron tulipifera L.) Exposed to Artificial Night Lighting. Forests 2018, 9, 74. [Google Scholar] [CrossRef]
- Mendes, M.M.; Pinheiro, A.C.R.; Pires, F.R.; Fernandes, A.A.; de Menezes, L.F.; Pereira, I.A.P.; Dos Santos, V.F.; de Almeida Leite, L.; Cassol, D.; Falqueto, A.R. Photosynthesis and Leaf Traits of Tree Species Influenced by Green Manure Associated with Soil Treatments. Commun. Soil Sci. Plant Anal. 2022, 53, 2064–2081. [Google Scholar] [CrossRef]
- Roach, T.; Krieger-Liszkay, A. Regulation of Photosynthetic Electron Transport and Photoinhibition. Curr. Protein Pept. Sci. 2014, 15, 351–362. [Google Scholar] [CrossRef]
- Tian, Y.; Sacharz, J.; Ware, M.A.; Zhang, H.; Ruban, A.V. Effects of Periodic Photoinhibitory Light Exposure on Physiology and Productivity of Arabidopsis Plants Grown under Low Light. J. Exp. Bot. 2017, 68, 4249–4262. [Google Scholar] [CrossRef]
- Zivcak, M.; Brestic, M.; Kunderlikova, K.; Sytar, O.; Allakhverdiev, S.I. Repetitive Light Pulse-Induced Photoinhibition of Photosystem I Severely Affects CO2 Assimilation and Photoprotection in Wheat Leaves. Photosynth. Res. 2015, 126, 449–463. [Google Scholar] [CrossRef] [PubMed]
- Kalaji, H.M.; Carpentier, R.; Allakhverdiev, S.I.; Bosa, K. Fluorescence Parameters as Early Indicators of Light Stress in Barley. J. Photochem. Photobiol. B Biol. 2012, 112, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Krüger, G.H.J.; Tsimilli-Michael, M.; Strasser, R.J. Light Stress Provokes Plastic and Elastic Modifications in Structure and Function of Photosystem II in Camellia Leaves. Physiol. Plant. 1997, 101, 265–277. [Google Scholar] [CrossRef]
- Heber, U.; Soni, V.; Strasser, R.J. Photoprotection of Reaction Centers: Thermal Dissipation of Absorbed Light Energy vs. Charge Separation in Lichens. Physiol. Plant. 2011, 142, 65–78. [Google Scholar] [CrossRef]
- Kumar, D.; Singh, H.; Raj, S.; Soni, V. Chlorophyll a Fluorescence Kinetics of Mung Bean (Vigna radiata L.) Grown under Artificial Continuous Light. Biochem. Biophys. Rep. 2020, 24, 100813. [Google Scholar] [CrossRef]
- Ceusters, N.; Valcke, R.; Frans, M.; Claes, J.E.; den Ende, W.; Ceusters, J. Performance Index and PSII Connectivity under Drought and Contrasting Light Regimes in the CAM Orchid Phalaenopsis. Front. Plant Sci. 2019, 10, 1012. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the Chlorophyll a Fluorescence Transient. In Chlorophyll a Fluorescence; Springer: Berlin/Heidelberg, Germany, 2004; pp. 321–362. [Google Scholar]
- Kalaji, H.M.; Rastogi, A.; Živčák, M.; Brestic, M.; Daszkowska-Golec, A.; Sitko, K.; Alsharafa, K.Y.; Lotfi, R.; Stypiński, P.; Samborska, I.A.; et al. Prompt Chlorophyll Fluorescence as a Tool for Crop Phenotyping: An Example of Barley Landraces Exposed to Various Abiotic Stress Factors. Photosynthetica 2018, 56, 953–961. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a Fluorescence as a Tool to Monitor Physiological Status of Plants under Abiotic Stress Conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef]
- Faseela, P.; Sinisha, A.K.; Brestič, M.; Puthur, J.T. Chlorophyll a Fluorescence Parameters as Indicators of a Particular Abiotic Stress in Rice. Photosynthetica 2019, 57, 108–115. [Google Scholar]
- Hermans, C.; Smeyers, M.; Rodriguez, R.M.; Eyletters, M.; Strasser, R.J.; Delhaye, J.-P. Quality Assessment of Urban Trees: A Comparative Study of Physiological Characterisation, Airborne Imaging and on Site Fluorescence Monitoring by the OJIP-Test. J. Plant Physiol. 2003, 160, 81–90. [Google Scholar] [CrossRef]
- Roháček, K. Chlorophyll Fluorescence Parameters: The Definitions, Photosynthetic Meaning, and Mutual Relationships. Photosynthetica 2002, 40, 13–29. [Google Scholar] [CrossRef]
- Schöttler, M.A.; Tóth, S.Z. Photosynthetic Complex Stoichiometry Dynamics in Higher Plants: Environmental Acclimation and Photosynthetic Flux Control. Front. Plant Sci. 2014, 5, 188. [Google Scholar] [PubMed]
- Ruban, A.V. Plants in Light. Commun. Integr. Biol. 2009, 2, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Fails, B.S.; Lewis, A.J.; Barden, J.A. Light Acclimatization Potential of Ficus Benjamina1. J. Am. Soc. Hortic. Sci. 1982, 107, 762–766. [Google Scholar] [CrossRef]
- Fails, B.S.; Lewis, A.J.; Barden, J.A. Anatomy and Morphology of Sun-and Shade-Grown Ficus Benjamina1. J. Am. Soc. Hortic. Sci. 1982, 107, 754–757. [Google Scholar] [CrossRef]
- Fjell, I. Anatomy of the Xeromorphic Leaves of Allamanda Neriifolia, Thevetia Peruviana and Vinca Minor (Apocynaceae). Nord. J. Bot. 1983, 3, 383–392. [Google Scholar] [CrossRef]
- DeLucia, E.H.; Day, T.A.; Vogelman, T.C. Ultraviolet-B and Visible Light Penetration into Needles of Two Species of Subalpine Conifers during Foliar Development. Plant Cell Environ. 1992, 15, 921–929. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, D.; Singh, H.; Bhatt, U.; Soni, V. Analyzing the Effects of Urban Photopollution on Photosynthetic Efficiency of Certain Trees through Chlorophyll Fluorescence OJIP Transient. Stresses 2022, 2, 437-449. https://doi.org/10.3390/stresses2040030
Kumar D, Singh H, Bhatt U, Soni V. Analyzing the Effects of Urban Photopollution on Photosynthetic Efficiency of Certain Trees through Chlorophyll Fluorescence OJIP Transient. Stresses. 2022; 2(4):437-449. https://doi.org/10.3390/stresses2040030
Chicago/Turabian StyleKumar, Deepak, Hanwant Singh, Upma Bhatt, and Vineet Soni. 2022. "Analyzing the Effects of Urban Photopollution on Photosynthetic Efficiency of Certain Trees through Chlorophyll Fluorescence OJIP Transient" Stresses 2, no. 4: 437-449. https://doi.org/10.3390/stresses2040030
APA StyleKumar, D., Singh, H., Bhatt, U., & Soni, V. (2022). Analyzing the Effects of Urban Photopollution on Photosynthetic Efficiency of Certain Trees through Chlorophyll Fluorescence OJIP Transient. Stresses, 2(4), 437-449. https://doi.org/10.3390/stresses2040030