

From Microbes to Molecules: Synthetic Biology Approaches for Advanced Materials Design



Abstract

1. Introduction
2. Biomineralization
2.1. Overview
2.2. Bioremediation
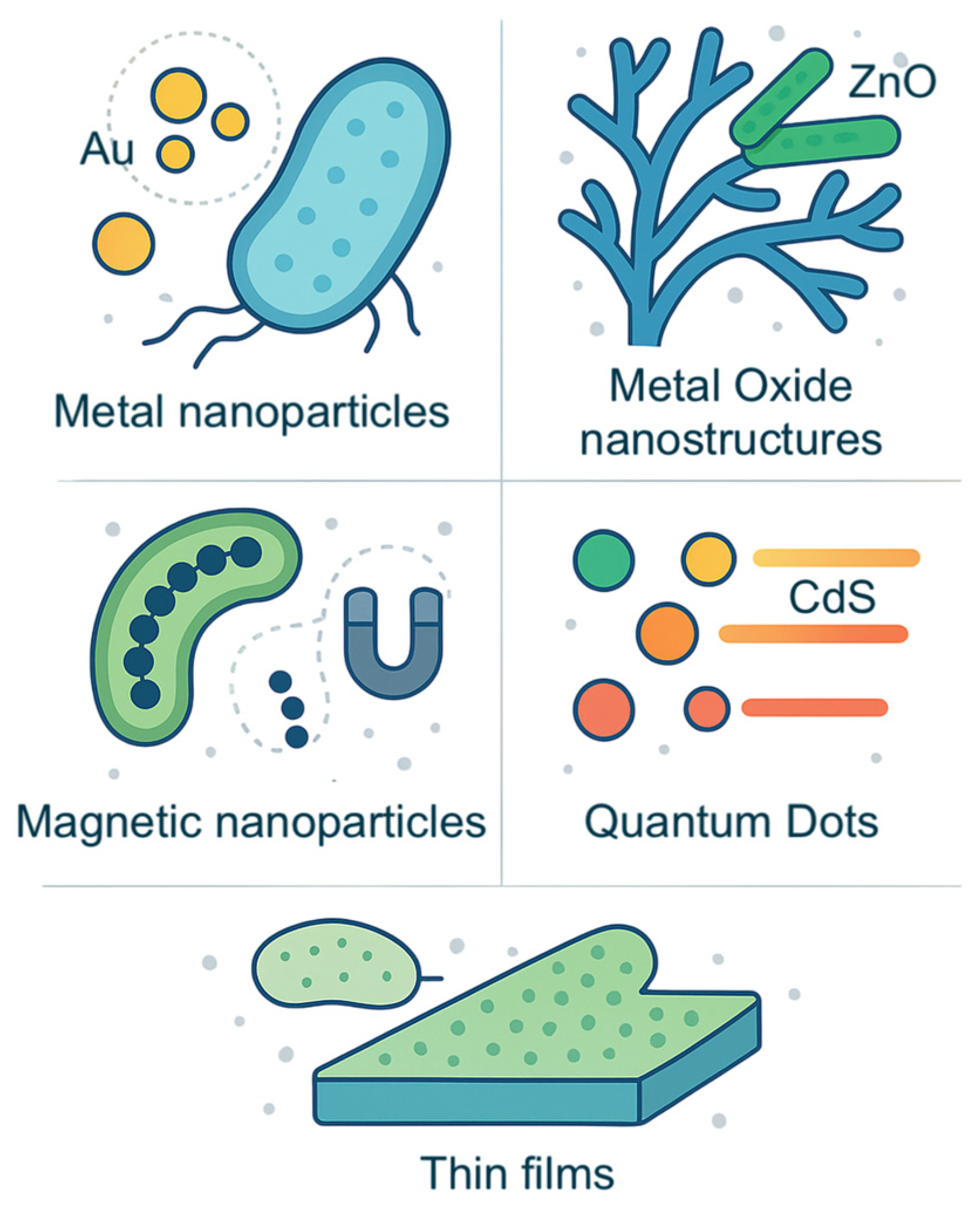
3. Synthesis of Advanced Materials
3.1. Inorganic Nanostructures
3.1.1. Metal Nanoparticles
3.1.2. Metal Oxide Nanostructures
3.1.3. Magnetic Nanoparticles
3.1.4. Semiconductor Nanocrystals
3.2. Thin Films Through Microbial Deposition
3.3. Biopolymers
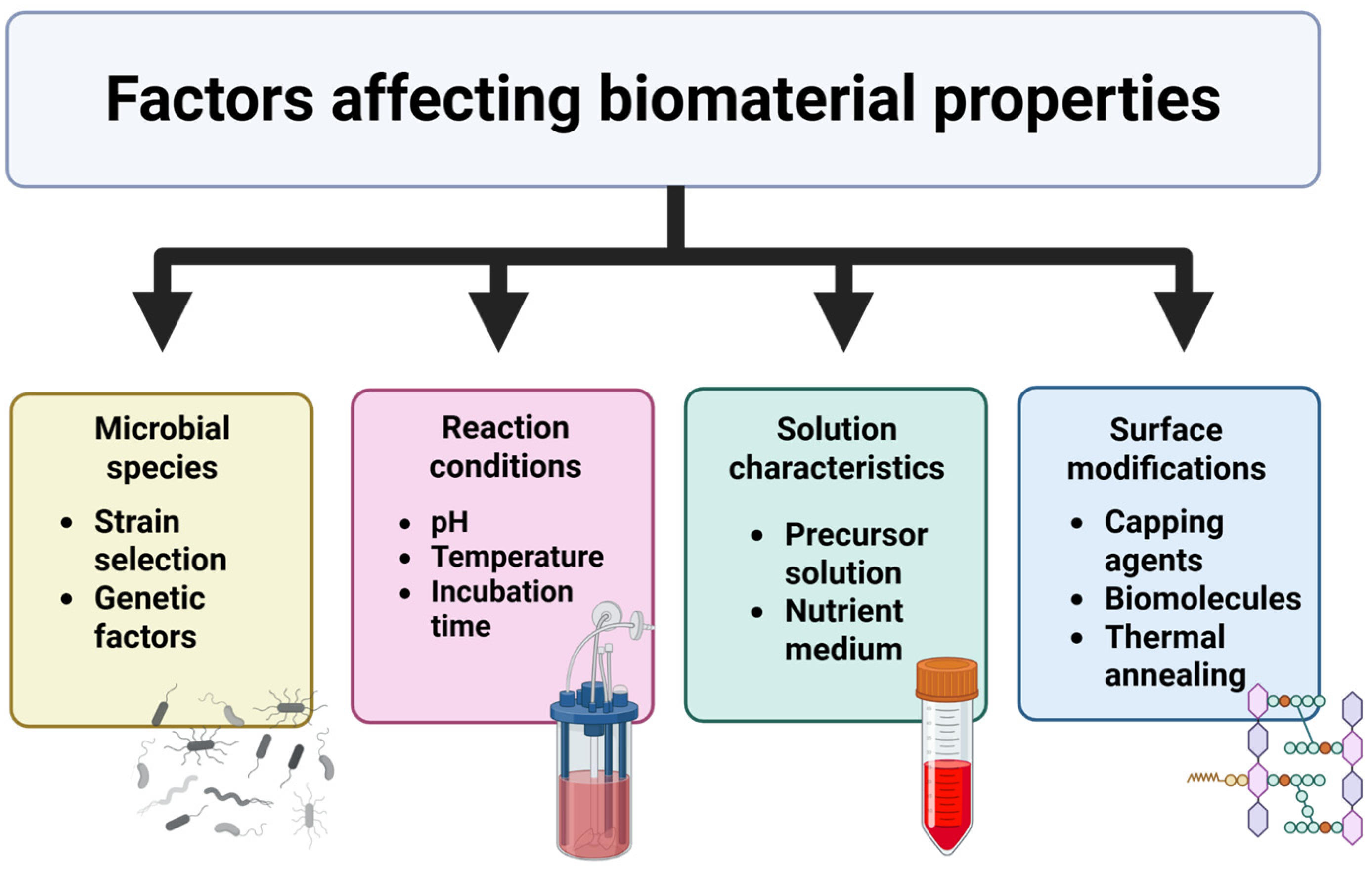
4. Factors Affecting Material Properties
4.1. Microbial Species/Strain Selection
4.2. Precursor and/or Nutrient Characteristics
4.3. Reaction pH, Temperature, and Time
4.4. Biomolecules and Capping Agents
5. Synergistic Approaches
5.1. Protein Engineering and Genome Editing
5.2. Smart and Responsive Living Materials
5.3. Resource Management and Recycling
6. Challenges and Future Directions
6.1. Large-Scale Production
6.2. Polydispersity and Uniformity
6.3. Compatibility and Waste Production
6.4. Ethical Considerations
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Annamalai, J.; Ummalyma, S.B.; Pandey, A.; Bhaskar, T. Recent Trends in Microbial Nanoparticle Synthesis and Potential Application in Environmental Technology: A Comprehensive Review. Environ. Sci. Pollut. Res. Int. 2021, 28, 49362–49382. [Google Scholar] [PubMed]
- Getahun, M.J.; Kassie, B.B.; Alemu, T.S. Recent Advances in Biopolymer Synthesis, Properties, & Commercial Applications: A Review. Process Biochem. 2024, 145, 261–287. [Google Scholar]
- Hulkoti, N.I.; Taranath, T.C. Biosynthesis of Nanoparticles Using Microbes—A Review. Colloids Surf. B Biointerfaces 2014, 121, 474–483. [Google Scholar]
- Raouf Hosseini, M.; Nasiri Sarvi, M. Recent Achievements in the Microbial Synthesis of Semiconductor Metal Sulfide Nanoparticles. Mater. Sci. Semicond. Process. 2015, 40, 293–301. [Google Scholar]
- Hernández-Arriaga, A.M.; Campano, C.; Rivero-Buceta, V.; Prieto, M.A. When Microbial Biotechnology Meets Material Engineering. Microb. Biotechnol. 2022, 15, 149–163. [Google Scholar] [CrossRef]
- Abid, N.; Khan, A.M.; Shujait, S.; Chaudhary, K.; Ikram, M.; Imran, M.; Haider, J.; Khan, M.; Khan, Q.; Maqbool, M. Synthesis of Nanomaterials Using Various Top-down and Bottom-up Approaches, Influencing Factors, Advantages, and Disadvantages: A Review. Adv. Colloid Interface Sci. 2022, 300, 102597. [Google Scholar] [CrossRef]
- Arole, D.V.; Munde, S.V. Fabrication of Nanomaterials by Top-down and Bottom-up Approaches—An Overview. J. Mater. Sci 2014, 1, 89–93. [Google Scholar]
- Jain, A.; Bhatia, P.; Chugh, A. Microbial Synthetic Biology for Human Therapeutics. Syst. Synth. Biol. 2012, 6, 9–22. [Google Scholar]
- Mironov, V.; Trusk, T.; Kasyanov, V.; Little, S.; Swaja, R.; Markwald, R. Biofabrication: A 21st Century Manufacturing Paradigm. Biofabrication 2009, 1, 022001. [Google Scholar]
- Chen, H.-G.; Zhang, Y.-H.P. New Biorefineries and Sustainable Agriculture: Increased Food, Biofuels, and Ecosystem Security. Renew. Sustain. Energy Rev. 2015, 47, 117–132. [Google Scholar] [CrossRef]
- Reid, A.; Buchanan, F.; Julius, M.; Walsh, P.J. A Review on Diatom Biosilicification and Their Adaptive Ability to Uptake Other Metals into Their Frustules for Potential Application in Bone Repair. J. Mater. Chem. B Mater. Biol. Med. 2021, 9, 6728–6737. [Google Scholar] [CrossRef] [PubMed]
- Raman, R.; Bashir, R. Biomimicry, Biofabrication, and Biohybrid Systems: The Emergence and Evolution of Biological Design. Adv. Healthc. Mater. 2017, 6, 1700496. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Chang, J.; Zhu, Y.; Wu, C. 3D Printing of Bioinspired Biomaterials for Tissue Regeneration. Adv. Healthc. Mater. 2020, 9, e2000208. [Google Scholar] [CrossRef]
- Ghosh, S.; Ahmad, R.; Zeyaullah, M.; Khare, S.K. Microbial Nano-Factories: Synthesis and Biomedical Applications. Front. Chem. 2021, 9, 626834. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, G.S.; Brar, S.K.; Kaur, S.; Verma, M. Green Approach for Nanoparticle Biosynthesis by Fungi: Current Trends and Applications. Crit. Rev. Biotechnol. 2012, 32, 49–73. [Google Scholar] [CrossRef]
- Yang, Y.; Waterhouse, G.I.N.; Chen, Y.; Sun-Waterhouse, D.; Li, D. Microbial-Enabled Green Biosynthesis of Nanomaterials: Current Status and Future Prospects. Biotechnol. Adv. 2022, 55, 107914. [Google Scholar] [CrossRef]
- Rodrigues, R.C.; Pereira, H.S.; Senra, R.L.; Ribon, A.d.O.B.; Mendes, T.A.d.O. Understanding the Emerging Potential of Synthetic Biology for Food Science: Achievements, Applications and Safety Considerations. Food Chem. Adv. 2023, 3, 100476. [Google Scholar] [CrossRef]
- Tamerler, C.; Sarikaya, M. Molecular Biomimetics: Nanotechnology and Bionanotechnology Using Genetically Engineered Peptides. Philos. Trans. A Math. Phys. Eng. Sci. 2009, 367, 1705–1726. [Google Scholar] [CrossRef]
- Isar, J.; Jain, D.; Joshi, H.; Dhoot, S.; Rangaswamy, V. MICROBIAL Isoprene Production: An Overview. World J. Microbiol. Biotechnol. 2022, 38, 122. [Google Scholar] [CrossRef]
- Kreyenschulte, D.; Krull, R.; Margaritis, A. Recent Advances in Microbial Biopolymer Production and Purification. Crit. Rev. Biotechnol. 2014, 34, 1–15. [Google Scholar] [CrossRef]
- Bittencourt, D.M.d.C.; Oliveira, P.; Michalczechen-Lacerda, V.A.; Rosinha, G.M.S.; Jones, J.A.; Rech, E.L. Bioengineering of Spider Silks for the Production of Biomedical Materials. Front. Bioeng. Biotechnol. 2022, 10, 958486. [Google Scholar] [CrossRef] [PubMed]
- Dhami, N.K.; Reddy, M.S.; Mukherjee, A. Biomineralization of Calcium Carbonates and Their Engineered Applications: A Review. Front. Microbiol. 2013, 4, 314. [Google Scholar] [CrossRef] [PubMed]
- Benzerara, K.; Miot, J.; Morin, G.; Ona-Nguema, G.; Skouri-Panet, F.; Férard, C. Significance, Mechanisms and Environmental Implications of Microbial Biomineralization. Comptes Rendus Geosci. 2010, 343, 160–167. [Google Scholar] [CrossRef]
- Keykha, H.A.; Zangani, A.; Romiani, H.M.; Asadi, A.; Kawasaki, S.; Radmanesh, N. Characterizing Microbial and CO2-Induced Carbonate Minerals: Implications for Soil Stabilization in Sandy Environments. Minerals 2023, 13, 976. [Google Scholar] [CrossRef]
- Elgendy, I.M.; Elkaliny, N.E.; Saleh, H.M.; Darwish, G.O.; Almostafa, M.M.; Metwally, K.; Yahya, G.; Mahmoud, Y.A.-G. Bacteria-Powered Self-Healing Concrete: Breakthroughs, Challenges, and Future Prospects. J. Ind. Microbiol. Biotechnol. 2024, 52, kuae051. [Google Scholar] [CrossRef]
- Rahman, M.M.; Hora, R.N.; Ahenkorah, I.; Beecham, S.; Karim, M.R.; Iqbal, A. State-of-the-Art Review of Microbial-Induced Calcite Precipitation and Its Sustainability in Engineering Applications. Sustainability 2020, 12, 6281. [Google Scholar] [CrossRef]
- Lazo, D.E.; Dyer, L.G.; Alorro, R.D. Silicate, Phosphate and Carbonate Mineral Dissolution Behaviour in the Presence of Organic Acids: A Review. Miner. Eng. 2017, 100, 115–123. [Google Scholar] [CrossRef]
- Tan, K.H. Degradation of Soil Minerals by Organic Acids. In SSSA Special Publications; Soil Science Society of America: Madison, WI, USA, 2015; pp. 1–27. ISBN 9780891189121. [Google Scholar]
- Moisescu, C.; Ardelean, I.I.; Benning, L.G. The Effect and Role of Environmental Conditions on Magnetosome Synthesis. Front. Microbiol. 2014, 5, 49. [Google Scholar] [CrossRef]
- Perez-Gonzalez, T.; Jimenez-Lopez, C.; Neal, A.L.; Rull-Perez, F.; Rodriguez-Navarro, A.; Fernandez-Vivas, A.; Iañez-Pareja, E. Magnetite Biomineralization Induced by Shewanella Oneidensis. Geochim. Cosmochim. Acta 2010, 74, 967–979. [Google Scholar] [CrossRef]
- Wan, J.; Ji, R.; Liu, J.; Ma, K.; Pan, Y.; Lin, W. Biomineralization in Magnetotactic Bacteria: From Diversity to Molecular Discovery-Based Applications. Cell Rep. 2024, 43, 114995. [Google Scholar] [CrossRef]
- Hoffmann, T.D.; Reeksting, B.J.; Gebhard, S. Bacteria-Induced Mineral Precipitation: A Mechanistic Review. Microbiology 2021, 167, 001049. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Wang, C.-Y.; Ma, Y.-X.; Shen, M.-J.; Li, J.; Jiao, K.; Tay, F.R.; Niu, L.-N. Microbe-Mediated Extracellular and Intracellular Mineralization: Environmental, Industrial, and Biotechnological Applications. Adv. Mater. 2020, 32, e1907833. [Google Scholar] [CrossRef] [PubMed]
- Lai Huat, L.; Swee Pin, Y.; Yao, H.; Huangpu, J.; Shao Feng, S.; Swee Sen, T. Environmental Bioremediation: Microremediation, Mycoremediation, and Phytoremediation. Environ. Claims J. 2024, 36, 186–205. [Google Scholar] [CrossRef]
- Lai, H.; Ding, X.; Cui, M.; Zheng, J.; Chen, Z.; Pei, J.; Zhang, J. Mechanisms and Influencing Factors of Biomineralization Based Heavy Metal Remediation: A Review. Biogeotechnics 2023, 1, 100039. [Google Scholar] [CrossRef]
- Cui, M.-J.; Lai, H.-J.; Wu, S.-F.; Chu, J. Comparison of Soil Improvement Methods Using Crude Soybean Enzyme, Bacterial Enzyme or Bacteria Induced Carbonate Precipitation. Géotechnique 2022, 74, 18–26. [Google Scholar] [CrossRef]
- Qabany, A.A.L.; Soga, K. Effect of Chemical Treatment Used in MICP on Engineering Properties of Cemented Soils. In Bio- and Chemo-Mechanical Processes in Geotechnical Engineering; ICE Publishing: London, UK, 2014; pp. 107–115. ISBN 9780727760531. [Google Scholar]
- Zhou, Y.; Zhao, X.; Jiang, Y.; Ding, C.; Liu, J.; Zhu, C. Synergistic Remediation of Lead Pollution by Biochar Combined with Phosphate Solubilizing Bacteria. Sci. Total Environ. 2023, 861, 160649. [Google Scholar] [CrossRef]
- Yu, X.; Jiang, J. Phosphate Microbial Mineralization Consolidation of Waste Incineration Fly Ash and Removal of Lead Ions. Ecotoxicol. Environ. Saf. 2020, 191, 110224. [Google Scholar] [CrossRef]
- Coelho, E.; Reis, T.A.; Cotrim, M.; Mullan, T.K.; Renshaw, J.; Rizzutto, M.; Corrêa, B. Talaromyces Amestolkiae Uses Organic Phosphate Sources for the Treatment of Uranium-Contaminated Water. Biometals 2022, 35, 335–348. [Google Scholar] [CrossRef]
- Zhang, K.; Xue, Y.; Xu, H.; Yao, Y. Lead Removal by Phosphate Solubilizing Bacteria Isolated from Soil through Biomineralization. Chemosphere 2019, 224, 272–279. [Google Scholar] [CrossRef]
- Zeng, G.; Qiao, S.; Wang, X.; Sheng, M.; Wei, M.; Chen, Q.; Xu, H.; Xu, F. Immobilization of Cadmium by Burkholderia Sp. QY14 through Modified Microbially Induced Phosphate Precipitation. J. Hazard. Mater. 2021, 412, 125156. [Google Scholar] [CrossRef]
- Zeng, G.; Wan, J.; Huang, D.; Hu, L.; Huang, C.; Cheng, M.; Xue, W.; Gong, X.; Wang, R.; Jiang, D. Precipitation, Adsorption and Rhizosphere Effect: The Mechanisms for Phosphate-Induced Pb Immobilization in Soils-A Review. J. Hazard. Mater. 2017, 339, 354–367. [Google Scholar] [CrossRef] [PubMed]
- Yin, T.; Lin, H.; Dong, Y.; Li, B.; He, Y.; Liu, C.; Chen, X. A Novel Constructed Carbonate-Mineralized Functional Bacterial Consortium for High-Efficiency Cadmium Biomineralization. J. Hazard. Mater. 2021, 401, 123269. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, K.B.; Sakthivel, N. Biological Synthesis of Metal Nanoparticles by Microbes. Adv. Colloid Interface Sci. 2010, 156, 1–13. [Google Scholar]
- Vanlalveni, C.; Ralte, V.; Zohmingliana, H.; Das, S.; Anal, J.M.H.; Lallianrawna, S.; Rokhum, S.L. A Review of Microbes Mediated Biosynthesis of Silver Nanoparticles and Their Enhanced Antimicrobial Activities. Heliyon 2024, 10, e32333. [Google Scholar] [CrossRef]
- Sridhar, S. Microbial Genetics; Wisdom Press: New Delhi, India, 2013; ISBN 9789382006213. [Google Scholar]
- Quester, K.; Avalos-Borja, M.; Castro-Longoria, E. Biosynthesis and Microscopic Study of Metallic Nanoparticles. Micron 2013, 54–55, 1–27. [Google Scholar] [CrossRef]
- Siddiqi, K.S.; Husen, A. Fabrication of Metal Nanoparticles from Fungi and Metal Salts: Scope and Application. Nanoscale Res. Lett. 2016, 11, 98. [Google Scholar]
- Korbekandi, H.; Ashari, Z.; Iravani, S.; Abbasi, S. Optimization of Biological Synthesis of Silver Nanoparticles Using Fusarium Oxysporum. Iran. J. Pharm. Res. 2013, 12, 289–298. [Google Scholar]
- Durán, N.; Marcato, P.D.; Alves, O.L.; Souza, G.I.H.D.; Esposito, E. Mechanistic Aspects of Biosynthesis of Silver Nanoparticles by Several Fusarium Oxysporum Strains. J. Nanobiotechnology 2005, 3, 8. [Google Scholar] [CrossRef]
- Tauseef, A.; Hisam, F.; Hussain, T.; Caruso, A.; Hussain, K.; Châtel, A.; Chénais, B. Nanomicrobiology: Emerging Trends in Microbial Synthesis of Nanomaterials and Their Applications. J. Clust. Sci. 2023, 34, 639–664. [Google Scholar]
- Wu, J.-W.; Ng, I.-S. Biofabrication of Gold Nanoparticles by Shewanella Species. Bioresour. Bioprocess. 2017, 4, 50. [Google Scholar] [CrossRef]
- Grasso, G.; Zane, D.; Dragone, R. Microbial Nanotechnology: Challenges and Prospects for Green Biocatalytic Synthesis of Nanoscale Materials for Sensoristic and Biomedical Applications. Nanomaterials 2019, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Konishi, Y.; Tsukiyama, T.; Tachimi, T.; Saitoh, N.; Nomura, T.; Nagamine, S. Microbial Deposition of Gold Nanoparticles by the Metal-Reducing Bacterium Shewanella Algae. Electrochim. Acta 2007, 53, 186–192. [Google Scholar] [CrossRef]
- Kitching, M.; Ramani, M.; Marsili, E. Fungal Biosynthesis of Gold Nanoparticles: Mechanism and Scale up: Fungal Biosynthesis of AuNPs. Microb. Biotechnol. 2015, 8, 904–917. [Google Scholar] [CrossRef]
- Hammad, S.E.; El-Rouby, M.N.; Abdel-Aziz, M.M.; El-Sayyad, G.S.; Elshikh, H.H. Endophytic Fungi–assisted Biomass Synthesis of Gold, and Zinc Oxide Nanoparticles for Increasing Antibacterial, and Anticancer Activities. Biomass Convers. Biorefin. 2025, 15, 2285–2302. [Google Scholar] [CrossRef]
- Ahmad, A.; Mukherjee, P.; Senapati, S.; Mandal, D.; Khan, M.I.; Kumar, R.; Sastry, M. Extracellular Biosynthesis of Silver Nanoparticles Using the Fungus Fusarium Oxysporum. Colloids Surf. B Biointerfaces 2003, 28, 313–318. [Google Scholar] [CrossRef]
- Gowramma, B.; Keerthi, U.; Rafi, M.; Muralidhara Rao, D. Biogenic Silver Nanoparticles Production and Characterization from Native Stain of Corynebacterium Species and Its Antimicrobial Activity. 3 Biotech 2015, 5, 195–201. [Google Scholar] [CrossRef]
- Rilda, Y.; Rinaldi, R.; Syukri, S.; Armaini, A.; Refinel, R.; Agustien, A.; Pardi, H. Biosynthesis of Zinc Oxide (ZnO) Using the Biomass of Aspergillus Niger to Impart Cotton Fabric with Antimicrobial Properties. ChemistrySelect 2022, 7, e202103824. [Google Scholar] [CrossRef]
- Sharma, J.L.; Dhayal, V.; Sharma, R.K. White-Rot Fungus Mediated Green Synthesis of Zinc Oxide Nanoparticles and Their Impregnation on Cellulose to Develop Environmental Friendly Antimicrobial Fibers. 3 Biotech 2021, 11, 269. [Google Scholar] [CrossRef]
- Hajiali, S.; Daneshjou, S.; Daneshjoo, S.; Khajeh, K. Biosynthesis Optimization of Antibacterial-Magnetic Iron Oxide Nanoparticles from Bacillus Megaterium. Biol. Trace Elem. Res. 2025, 203, 467–484. [Google Scholar] [CrossRef]
- Vasanth, V.; Murugesh, K.A.; Susikaran, S. Synthesis of Titanium Dioxide Nanoparticles Using Spirulina Platensis Algae Extract. Pharma Innov. J. 2022, 11, 266–269. [Google Scholar]
- Saha, M.; Sarkar, S.; Sarkar, B.; Sharma, B.K.; Bhattacharjee, S.; Tribedi, P. Microbial Siderophores and Their Potential Applications: A Review. Environ. Sci. Pollut. Res. Int. 2016, 23, 3984–3999. [Google Scholar] [CrossRef] [PubMed]
- De Windt, W.; Boon, N.; Van den Bulcke, J.; Rubberecht, L.; Prata, F.; Mast, J.; Hennebel, T.; Verstraete, W. Biological Control of the Size and Reactivity of Catalytic Pd(0) Produced by Shewanella Oneidensis. Antonie Van Leeuwenhoek 2006, 90, 377–389. [Google Scholar] [CrossRef] [PubMed]
- ELtarahony, M.; Zaki, S.; Kheiralla, Z.; Abd-El-Haleem, D. Biogenic Synthesis of Iron Oxide Nanoparticles via Optimization of Nitrate Reductase Enzyme Using Statistical Experimental Design. J. Adv. Biotechnol 2016, 5, 667–684. [Google Scholar]
- Elblbesy, M.A.; Madbouly, A.; Hamdan, T. Bio-Synthesis of Magnetite Nanoparticles by Bacteria. Am. J. Nano Res. Appl. 2014, 2, 98. [Google Scholar]
- Hossain, S.; Bahreini, B.; Pasteur, E. Magnetotactic Bacteria and Magnetosomes–an Overview. Mater. Sci. Eng. 2024, 8, 83–100. [Google Scholar] [CrossRef]
- Noguchi, Y.; Fujiwara, T.; Yoshimatsu, K.; Fukumori, Y. Iron Reductase for Magnetite Synthesis in the Magnetotactic Bacterium Magnetospirillum Magnetotacticum. J. Bacteriol. 1999, 181, 2142–2147. [Google Scholar] [CrossRef]
- Fdez-Gubieda, M.L.; Muela, A.; Alonso, J.; García-Prieto, A.; Olivi, L.; Fernández-Pacheco, R.; Barandiarán, J.M. Magnetite Biomineralization in Magnetospirillum Gryphiswaldense: Time-Resolved Magnetic and Structural Studies. ACS Nano 2013, 7, 3297–3305. [Google Scholar] [CrossRef]
- Amor, M.; Ceballos, A.; Wan, J.; Simon, C.P.; Aron, A.T.; Chang, C.J.; Hellman, F.; Komeili, A. Magnetotactic Bacteria Accumulate a Large Pool of Iron Distinct from Their Magnetite Crystals. Appl. Environ. Microbiol. 2020, 86, e01278-20. [Google Scholar] [CrossRef]
- Khan, A.A.; Khan, S.; Khan, S.; Rentschler, S.; Laufer, S.; Deigner, H.-P. Biosynthesis of Iron Oxide Magnetic Nanoparticles Using Clinically Isolated Pseudomonas Aeruginosa. Sci. Rep. 2021, 11, 20503. [Google Scholar] [CrossRef]
- Liße, D.; Monzel, C.; Vicario, C.; Manzi, J.; Maurin, I.; Coppey, M.; Piehler, J.; Dahan, M. Engineered Ferritin for Magnetogenetic Manipulation of Proteins and Organelles inside Living Cells. Adv. Mater. 2017, 29, 1700189. [Google Scholar] [CrossRef]
- Zhou, Y.; Gao, Y.; Xie, Q.; Wang, J.; Yue, Z.; Wei, L.; Yang, Y.; Li, L.; Chen, T. Reduction and Transformation of Nanomagnetite and Nanomaghemite by a Sulfate-Reducing Bacterium. Geochim. Cosmochim. Acta 2019, 256, 66–81. [Google Scholar] [CrossRef]
- Pan, J.; Qian, H.; Sun, Y.; Miao, Y.; Zhang, J.; Li, Y. Microbially Synthesized Nanomaterials: Advances and Applications in Biomedicine. Precis. Med. Eng. 2025, 2, 100019. [Google Scholar] [CrossRef]
- Bao, H.; Lu, Z.; Cui, X.; Qiao, Y.; Guo, J.; Anderson, J.M.; Li, C.M. Extracellular Microbial Synthesis of Biocompatible CdTe Quantum Dots. Acta Biomater. 2010, 6, 3534–3541. [Google Scholar] [CrossRef]
- Mi, C.; Wang, Y.; Zhang, J.; Huang, H.; Xu, L.; Wang, S.; Fang, X.; Fang, J.; Mao, C.; Xu, S. Biosynthesis and Characterization of CdS Quantum Dots in Genetically Engineered Escherichia Coli. J. Biotechnol. 2011, 153, 125–132. [Google Scholar] [CrossRef]
- Tusher, M.M.H. Microbial Synthesis of Cadmium Selenide Quantum Dots (CdSe QDs), Influencing Factors and Applications. Opt. Quantum Electron. 2023, 55, 332. [Google Scholar] [CrossRef]
- Alghuthaymi, M.A.; Almoammar, H.; Rai, M.; Said-Galiev, E.; Abd-Elsalam, K.A. Myconanoparticles: Synthesis and Their Role in Phytopathogens Management. Biotechnol. Biotechnol. Equip. 2015, 29, 221–236. [Google Scholar] [CrossRef]
- Su, Z.; Li, X.; Xi, Y.; Xie, T.; Liu, Y.; Liu, B.; Liu, H.; Xu, W.; Zhang, C. Microbe-Mediated Transformation of Metal Sulfides: Mechanisms and Environmental Significance. Sci. Total Environ. 2022, 825, 153767. [Google Scholar] [CrossRef]
- Chen, Y.-L.; Tuan, H.-Y.; Tien, C.-W.; Lo, W.-H.; Liang, H.-C.; Hu, Y.-C. Augmented Biosynthesis of Cadmium Sulfide Nanoparticles by Genetically Engineered Escherichia Coli. Biotechnol. Prog. 2009, 25, 1260–1266. [Google Scholar] [CrossRef]
- Mirzadeh, S.; Darezereshki, E.; Bakhtiari, F.; Fazaelipoor, M.H.; Hosseini, M.R. Characterization of Zinc Sulfide (ZnS) Nanoparticles Biosynthesized by Fusarium Oxysporum. Mater. Sci. Semicond. Process. 2013, 16, 374–378. [Google Scholar] [CrossRef]
- Senapati, S.; Syed, A.; Khan, S.; Pasricha, R.; Khan, M.; Kumar, R.; Ahmad, A. Extracellular Biosynthesis of Metal Sulfide Nanoparticles Using the Fungus Fusarium Oxysporum. Curr. Nanosci. 2014, 10, 588–595. [Google Scholar] [CrossRef]
- Oliva-Arancibia, B.; Órdenes-Aenishanslins, N.; Bruna, N.; Ibarra, P.S.; Zacconi, F.C.; Pérez-Donoso, J.M.; Poblete-Castro, I. Co-Synthesis of Medium-Chain-Length Polyhydroxyalkanoates and CdS Quantum Dots Nanoparticles in Pseudomonas Putida KT2440. J. Biotechnol. 2017, 264, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Hardman, R. A Toxicologic Review of Quantum Dots: Toxicity Depends on Physicochemical and Environmental Factors. Environ. Health Perspect. 2006, 114, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Sobhanan, J.; Jones, P.; Kohara, R.; Sugino, S.; Vacha, M.; Subrahmanyam, C.; Takano, Y.; Lacy, F.; Biju, V. Toxicity of Nanomaterials due to Photochemical Degradation and the Release of Heavy Metal Ions. Nanoscale 2020, 12, 22049–22058. [Google Scholar] [CrossRef]
- Calvo-Olvera, A.; De Donato-Capote, M.; Pool, H.; Rojas-Avelizapa, N.G. In Vitro Toxicity Assessment of Fungal-Synthesized Cadmium Sulfide Quantum Dots Using Bacteria and Seed Germination Models. J. Environ. Sci. Health A Tox. Hazard. Subst. Environ. Eng. 2021, 56, 713–722. [Google Scholar] [CrossRef]
- Feng, J.; Chen, B.; Sun, W.; Wang, Y. Microbial Induced Calcium Carbonate Precipitation Study Using Bacillus Subtilis with Application to Self-Healing Concrete Preparation and Characterization. Constr. Build. Mater. 2021, 280, 122460. [Google Scholar] [CrossRef]
- Ahmad, S.S.E.; Elmahdy, M.A.R.; ELShami, A.A.; Yousry, E.-S.M. Bacterial Sustainable Concrete for Repair and Rehabilitation of Structural Cracks. J. Sustain. Cem.-Based Mater. 2024, 20, e03188. [Google Scholar] [CrossRef]
- Fan, Q.; Fan, L.; Quach, W.-M.; Zhang, R.; Duan, J.; Sand, W. Application of Microbial Mineralization Technology for Marine Concrete Crack Repair: A Review. J. Build. Eng. 2023, 69, 106299. [Google Scholar] [CrossRef]
- Renninger, N.; Knopp, R.; Nitsche, H.; Clark, D.S.; Keasling, J.D. Uranyl Precipitation by Pseudomonas Aeruginosa via Controlled Polyphosphate Metabolism. Appl. Environ. Microbiol. 2004, 70, 7404–7412. [Google Scholar] [CrossRef]
- Liang, X.; Hillier, S.; Pendlowski, H.; Gray, N.; Ceci, A.; Gadd, G.M. Uranium Phosphate Biomineralization by Fungi. Environ. Microbiol. 2015, 17, 2064–2075. [Google Scholar] [CrossRef]
- Manobala, T.; Shukla, S.K.; Rao, T.S.; Kumar, M.D. Uranium Sequestration by Biofilm-Forming Bacteria Isolated from Marine Sediment Collected from Southern Coastal Region of India. Int. Biodeterior. Biodegrad. 2019, 145, 104809. [Google Scholar] [CrossRef]
- Toster, J.; Harnagea, C.; Iyer, K.; Rosei, F.; Raston, C. Controlling Anatase Coating of Diatom Frustules by Varying the Binding Layer. CrystEngComm 2012, 14, 3446–3450. [Google Scholar] [CrossRef]
- Lahiri, D.; Nag, M.; Dutta, B.; Dey, A.; Sarkar, T.; Pati, S.; Edinur, H.A.; Abdul Kari, Z.; Mohd Noor, N.H.; Ray, R.R. Bacterial Cellulose: Production, Characterization, and Application as Antimicrobial Agent. Int. J. Mol. Sci. 2021, 22, 12984. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.U.; Ullah, M.W.; Khan, S.; Shah, N.; Park, J.K. Strategies for Cost-Effective and Enhanced Production of Bacterial Cellulose. Int. J. Biol. Macromol. 2017, 102, 1166–1173. [Google Scholar] [CrossRef]
- Kadier, A.; Ilyas, R.A.; Huzaifah, M.R.M.; Harihastuti, N.; Sapuan, S.M.; Harussani, M.M.; Azlin, M.N.M.; Yuliasni, R.; Ibrahim, R.; Atikah, M.S.N.; et al. Use of Industrial Wastes as Sustainable Nutrient Sources for Bacterial Cellulose (BC) Production: Mechanism, Advances, and Future Perspectives. Polymers 2021, 13, 3365. [Google Scholar] [CrossRef]
- Song, J.; Winkeljann, B.; Lieleg, O. Biopolymer-based Coatings: Promising Strategies to Improve the Biocompatibility and Functionality of Materials Used in Biomedical Engineering. Adv. Mater. Interfaces 2020, 7, 2000850. [Google Scholar] [CrossRef]
- Rehm, B.H.; Moradali, M.F. (Eds.) Biopolymers for Biomedical and Biotechnological Applications; Wiley: Hoboken, NJ, USA, 2021; ISBN 9783527818310. [Google Scholar]
- Rosalam, S.; England, R. Review of Xanthan Gum Production from Unmodified Starches by Xanthomonas Comprestris sp. Enzym. Microb. Technol. 2006, 39, 197–207. [Google Scholar] [CrossRef]
- Ronďošová, S.; Legerská, B.; Chmelová, D.; Ondrejovič, M.; Miertus, S. Optimization of Growth Conditions to Enhance PHA Production by Cupriavidus Necator. Fermentation 2022, 8, 451. [Google Scholar] [CrossRef]
- Zytner, P.; Kumar, D.; Elsayed, A.; Mohanty, A.; Ramarao, B.V.; Misra, M. A Review on Polyhydroxyalkanoate (PHA) Production through the Use of Lignocellulosic Biomass. RSC Sustain. 2023, 1, 2120–2134. [Google Scholar] [CrossRef]
- Policastro, G.; Panico, A.; Fabbricino, M. Improving Biological Production of poly(3-Hydroxybutyrate-Co-3-Hydroxyvalerate) (PHBV) Co-Polymer: A Critical Review. Rev. Environ. Sci. Biotechnol. 2021, 20, 479–513. [Google Scholar] [CrossRef]
- Tian, X.; Chen, H.; Liu, H.; Chen, J. Recent Advances in Lactic Acid Production by Lactic Acid Bacteria. Appl. Biochem. Biotechnol. 2021, 193, 4151–4171. [Google Scholar] [CrossRef]
- Dessie, W.; Xin, F.; Zhang, W.; Jiang, Y.; Wu, H.; Ma, J.; Jiang, M. Opportunities, Challenges, and Future Perspectives of Succinic Acid Production by Actinobacillus Succinogenes. Appl. Microbiol. Biotechnol. 2018, 102, 9893–9910. [Google Scholar] [PubMed]
- Pateraki, C.; Patsalou, M.; Vlysidis, A.; Kopsahelis, N.; Webb, C.; Koutinas, A.A.; Koutinas, M. Actinobacillus Succinogenes: Advances on Succinic Acid Production and Prospects for Development of Integrated Biorefineries. Biochem. Eng. J. 2016, 112, 285–303. [Google Scholar] [CrossRef]
- Eiteman, M.A.; Ramalingam, S. Microbial Production of Lactic Acid. Biotechnol. Lett. 2015, 37, 955–972. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Microbial Production of Lactic Acid: The Latest Development. Crit. Rev. Biotechnol. 2016, 36, 967–977. [Google Scholar] [CrossRef]
- Amara, N.I.; Chukwuemeka, E.S.; Obiajulu, N.O.; Chukwuma, O.J. Yeast-Driven Valorization of Agro-Industrial Wastewater: An Overview. Environ. Monit. Assess. 2023, 195, 1252. [Google Scholar] [CrossRef]
- Saravanan, M.; Gopinath, V.; Chaurasia, M.K.; Syed, A.; Ameen, F.; Purushothaman, N. Green Synthesis of Anisotropic Zinc Oxide Nanoparticles with Antibacterial and Cytofriendly Properties. Microb. Pathog. 2018, 115, 57–63. [Google Scholar] [CrossRef]
- Jacob, P.J.; Masarudin, M.J.; Hussein, M.Z.; Rahim, R.A. Optimization of Process Parameters Influencing the Sustainable Construction of Iron Oxide Nanoparticles by a Novel Tropical Wetlands Streptomyces spp. J. Clean. Prod. 2019, 232, 193–202. [Google Scholar] [CrossRef]
- Rajput, V.D.; Minkina, T.; Kimber, R.L.; Singh, V.K.; Shende, S.; Behal, A.; Sushkova, S.; Mandzhieva, S.; Lloyd, J.R. Insights into the Biosynthesis of Nanoparticles by the Genus Shewanella. Appl. Environ. Microbiol. 2021, 87, e01390-21. [Google Scholar] [CrossRef]
- Khort, A.; Brookman-Amissah, M.; Hedberg, J.; Chang, T.; Mei, N.; Lundberg, A.; Sturve, J.; Blomberg, E.; Odnevall, I. Influence of Natural Organic Matter on the Transformation of Metal and Metal Oxide Nanoparticles and Their Ecotoxic Potency In Vitro. NanoImpact 2022, 25, 100386. [Google Scholar] [CrossRef]
- Mohammadkazemi, F.; Azin, M.; Ashori, A. Production of Bacterial Cellulose Using Different Carbon Sources and Culture Media. Carbohydr. Polym. 2015, 117, 518–523. [Google Scholar] [CrossRef]
- Hu, H.; Catchmark, J.M.; Demirci, A. Effects of Pullulan Additive and Co-Culture of Aureobasidium Pullulans on Bacterial Cellulose Produced by Komagataeibacter Hansenii. Bioprocess Biosyst. Eng. 2022, 45, 573–587. [Google Scholar]
- Chavan, S.; Baig, M. Relationship of Biomass and Xanthan Gum Production by Xanthomonas Campestris: Optimization of Parameters. Br. Biotechnol. J. 2016, 11, 1–8. [Google Scholar] [CrossRef]
- Trindade, R.A.; Munhoz, A.P.; Burkert, C.A.V. Impact of a Carbon Source and Stress Conditions on Some Properties of Xanthan Gum Produced by Xanthomonas Campestris Pv. Mangiferaeindicae. Biocatal. Agric. Biotechnol. 2018, 15, 167–172. [Google Scholar] [CrossRef]
- Zulfiqar, Z.; Khan, R.R.M.; Summer, M.; Saeed, Z.; Pervaiz, M.; Rasheed, S.; Shehzad, B.; Kabir, F.; Ishaq, S. Plant-Mediated Green Synthesis of Silver Nanoparticles: Synthesis, Characterization, Biological Applications, and Toxicological Considerations: A Review. Biocatal. Agric. Biotechnol. 2024, 57, 103121. [Google Scholar] [CrossRef]
- Ibrahim, S.; Ahmad, Z.; Manzoor, M.Z.; Mujahid, M.; Faheem, Z.; Adnan, A. Optimization for Biogenic Microbial Synthesis of Silver Nanoparticles through Response Surface Methodology, Characterization, Their Antimicrobial, Antioxidant, and Catalytic Potential. Sci. Rep. 2021, 11, 770. [Google Scholar]
- Nejadmansouri, M.; Shad, E.; Razmjooei, M.; Safdarianghomsheh, R.; Delvigne, F.; Khalesi, M. Production of Xanthan Gum Using Immobilized Xanthomonas Campestris Cells: Effects of Support Type. Biochem. Eng. J. 2020, 157, 107554. [Google Scholar] [CrossRef]
- Li, P.; Li, T.; Zeng, Y.; Li, X.; Jiang, X.; Wang, Y.; Xie, T.; Zhang, Y. Biosynthesis of Xanthan Gum by Xanthomonas Campestris LRELP-1 Using Kitchen Waste as the Sole Substrate. Carbohydr. Polym. 2016, 151, 684–691. [Google Scholar] [CrossRef]
- Baker, S.; Mohan Kumar, K.; Santosh, P.; Rakshith, D.; Satish, S. Extracellular Synthesis of Silver Nanoparticles by Novel Pseudomonas Veronii AS41G Inhabiting Annona Squamosa L. and Their Bactericidal Activity. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2015, 136 Pt C, 1434–1440. [Google Scholar] [CrossRef]
- Saravana Kumar, P.; Balachandran, C.; Duraipandiyan, V.; Ramasamy, D.; Ignacimuthu, S.; Al-Dhabi, N.A. Extracellular Biosynthesis of Silver Nanoparticle Using Streptomyces Sp. 09 PBT 005 and Its Antibacterial and Cytotoxic Properties. Appl. Nanosci. 2015, 5, 169–180. [Google Scholar] [CrossRef]
- Guo, E.; Chen, G.; Yu, D.; Qiu, Y.; Li, S.; Yu, Y. Optimization of Dry Anaerobic Co-Fermentation of Sludge and Corn Straw with Magnetite (Fe3O4). J. Environ. Chem. Eng. 2022, 10, 108618. [Google Scholar]
- Tyagi, S.; Tyagi, P.K.; Gola, D.; Chauhan, N.; Bharti, R.K. Extracellular Synthesis of Silver Nanoparticles Using Entomopathogenic Fungus: Characterization and Antibacterial Potential. SN Appl. Sci. 2019, 1, 1545. [Google Scholar] [CrossRef]
- Balashanmugam, P.; Balakumaran, M.D.; Murugan, R.; Dhanapal, K.; Kalaichelvan, P.T. Phytogenic Synthesis of Silver Nanoparticles, Optimization and Evaluation of in Vitro Antifungal Activity against Human and Plant Pathogens. Microbiol. Res. 2016, 192, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Krishna Leela, J.; Sharma, G. Studies on Xanthan Production from Xanthomonas Campestris. Bioprocess Biosyst. Eng. 2000, 23, 687–689. [Google Scholar]
- Chakraborty, I.; Feliu, N.; Roy, S.; Dawson, K.; Parak, W.J. Protein-Mediated Shape Control of Silver Nanoparticles. Bioconjug. Chem. 2018, 29, 1261–1265. [Google Scholar]
- Chakraborty, I.; Parak, W.J. Protein-induced Shape Control of Noble Metal Nanoparticles. Adv. Mater. Interfaces 2019, 6, 1801407. [Google Scholar]
- Murphy, C.J.; Chang, H.-H.; Falagan-Lotsch, P.; Gole, M.T.; Hofmann, D.M.; Hoang, K.N.L.; McClain, S.M.; Meyer, S.M.; Turner, J.G.; Unnikrishnan, M.; et al. Virus-Sized Gold Nanorods: Plasmonic Particles for Biology. Acc. Chem. Res. 2019, 52, 2124–2135. [Google Scholar]
- Duman, E.; Şahin Kehribar, E.; Ahan, R.E.; Yuca, E.; Şeker, U.Ö.Ş. Biomineralization of Calcium Phosphate Crystals Controlled by Protein-Protein Interactions. ACS Biomater. Sci. Eng. 2019, 5, 4750–4763. [Google Scholar]
- Mohammadi, P.; Gandier, J.-A.; Nonappa; Wagermaier, W.; Miserez, A.; Penttilä, M. Bioinspired Functionally Graded Composite Assembled Using Cellulose Nanocrystals and Genetically Engineered Proteins with Controlled Biomineralization. Adv. Mater. 2021, 33, e2102658. [Google Scholar]
- Nie, Z.; Zhang, Y.; Tang, R.; Wang, X. Biomimetic Mineralization: An Emerging Organism Engineering Strategy for Biomedical Applications. J. Inorg. Biochem. 2022, 232, 111815. [Google Scholar]
- Dickerson, M.B.; Sandhage, K.H.; Naik, R.R. Protein- and Peptide-Directed Syntheses of Inorganic Materials. Chem. Rev. 2008, 108, 4935–4978. [Google Scholar]
- Tamerler, C.; Kacar, T.; Sahin, D.; Fong, H.; Sarikaya, M. Genetically Engineered Polypeptides for Inorganics: A Utility in Biological Materials Science and Engineering. Mater. Sci. Eng. C Mater. Biol. Appl. 2007, 27, 558–564. [Google Scholar]
- Qin, K.; Zheng, Z.; Wang, J.; Pan, H.; Tang, R. Biomineralization Strategy: From Material Manufacturing to Biological Regulation. Giant 2024, 19, 100317. [Google Scholar] [CrossRef]
- Miserez, A.; Yu, J.; Mohammadi, P. Protein-Based Biological Materials: Molecular Design and Artificial Production. Chem. Rev. 2023, 123, 2049–2111. [Google Scholar]
- Schulz, A.; Wang, H.; Rijn, P.; Böker, A. Synthetic Inorganic Materials by Mimicking Biomineralization Processes Using Native and Non-Native Protein Functions. J. Mater. Chem. 2011, 21, 18903–18918. [Google Scholar]
- Wang, Y.; Guo, J.; Zhou, L.; Ye, C.; Omenetto, F.G.; Kaplan, D.L.; Ling, S. Design, Fabrication, and Function of Silk-Based Nanomaterials. Adv. Funct. Mater. 2018, 28, 1805305. [Google Scholar]
- Chandraker, S.K.; Kumar, R. Biogenic Biocompatible Silver Nanoparticles: A Promising Antibacterial Agent. Biotechnol. Genet. Eng. Rev. 2024, 40, 3113–3147. [Google Scholar] [CrossRef]
- Russo, P.; De Simone, N.; Capozzi, V.; Mohedano, M.L.; Ruiz-Masó, J.Á.; Del Solar, G.; López, P.; Spano, G. Selection of Riboflavin Overproducing Strains of Lactic Acid Bacteria and Riboflavin Direct Quantification by Fluorescence. Methods Mol. Biol. 2021, 2280, 3–14. [Google Scholar]
- Kant, G.; Pandey, A.; Kumari, S.; Bux, F.; Srivastava, S. Enhancing Bio-Isoprene Production in Escherichia Coli through a Combinatorial Optimization Approach. Process Biochem. 2024, 144, 210–219. [Google Scholar]
- De Carluccio, G.; Fusco, V.; di Bernardo, D. Engineering a Synthetic Gene Circuit for High-Performance Inducible Expression in Mammalian Systems. Nat. Commun. 2024, 15, 3311. [Google Scholar]
- Wang, Y.; Liu, Y.; Li, J.; Chen, Y.; Liu, S.; Zhong, C. Engineered Living Materials (ELMs) Design: From Function Allocation to Dynamic Behavior Modulation. Curr. Opin. Chem. Biol. 2022, 70, 102188. [Google Scholar]
- Sadeghianmaryan, A.; Ahmadian, N.; Wheatley, S.; Alizadeh Sardroud, H.; Nasrollah, S.A.S.; Naseri, E.; Ahmadi, A. Advancements in 3D-Printable Polysaccharides, Proteins, and Synthetic Polymers for Wound Dressing and Skin Scaffolding—A Review. Int. J. Biol. Macromol. 2024, 266, 131207. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, D.G.; Curtin, C.; O’Brien, F. Articulation Inspired by Nature: A Review of Biomimetic and Biologically Active 3D Printed Scaffolds for Cartilage Tissue Engineering. Biomater. Sci. 2022, 10, 2462–2483. [Google Scholar] [CrossRef] [PubMed]
- Heveran, C.M.; Williams, S.L.; Qiu, J.; Artier, J.; Hubler, M.H.; Cook, S.M.; Cameron, J.C.; Srubar, W.V., III. Biomineralization and Successive Regeneration of Engineered Living Building Materials. Matter 2020, 2, 481–494. [Google Scholar] [CrossRef]
- Bi, S.; He, C.; Liu, R.; Zhao, X.; Liu, J.; Gu, J.; Liu, W.; Yan, B. On-Demand Dissociable Antibacterial Self-Healing Hydrogel with Rapid Arginine-Related Biosynthesis Capacity to Promote Full-Thickness Bacteria-Infected Wound Healing. Ind. Eng. Chem. Res. 2023, 62, 7492–7503. [Google Scholar] [CrossRef]
- Yang, M.; Yang, F.; Chen, W.; Liu, S.; Qiu, L.; Chen, J. Bacteria-Mediated Cancer Therapies: Opportunities and Challenges. Biomater. Sci. 2021, 9, 5732–5744. [Google Scholar] [CrossRef]
- Fan, J.-X.; Niu, M.-T.; Qin, Y.-T.; Sun, Y.-X.; Zhang, X.-Z. Progress of Engineered Bacteria for Tumor Therapy. Adv. Drug Deliv. Rev. 2022, 185, 114296. [Google Scholar] [CrossRef]
- Lin, D.; Feng, X.; Mai, B.; Li, X.; Wang, F.; Liu, J.; Liu, X.; Zhang, K.; Wang, X. Bacterial-Based Cancer Therapy: An Emerging Toolbox for Targeted Drug/gene Delivery. Biomaterials 2021, 277, 121124. [Google Scholar] [CrossRef]
- Jiang, W.; Wu, Z.; Gao, Z.; Wan, M.; Zhou, M.; Mao, C.; Shen, J. Artificial Cells: Past, Present and Future. ACS Nano 2022, 16, 15705–15733. [Google Scholar] [CrossRef]
- Choi, S. Electrogenic Bacteria Promise New Opportunities for Powering, Sensing, and Synthesizing. Small 2022, 18, e2107902. [Google Scholar] [CrossRef]
- Shin, H.J. Genetically Engineered Microbial Biosensors for in Situ Monitoring of Environmental Pollution. Appl. Microbiol. Biotechnol. 2011, 89, 867–877. [Google Scholar] [CrossRef]
- Girotti, S.; Ferri, E.N.; Fumo, M.G.; Maiolini, E. Monitoring of Environmental Pollutants by Bioluminescent Bacteria. Anal. Chim. Acta 2008, 608, 2–29. [Google Scholar] [CrossRef] [PubMed]
- Belkin, S. Microbial Whole-Cell Sensing Systems of Environmental Pollutants. Curr. Opin. Microbiol. 2003, 6, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Belkin, S. Genetically Engineered Microorganisms for Pollution Monitoring. In NATO Science Series; Springer: Dordrecht, The Netherlands, 2007; pp. 147–160. ISBN 9781402047268. [Google Scholar]
- Parakh, P.D.; Nanda, S.; Kozinski, J.A. Eco-Friendly Transformation of Waste Biomass to Biofuels. Curr. Biochem. Eng. 2020, 6, 120–134. [Google Scholar] [CrossRef]
- Zeng, J.; Zeng, H.; Wang, Z. Review on Technology of Making Biofuel from Food Waste. Int. J. Energy Res. 2022, 46, 10301–10319. [Google Scholar] [CrossRef]
- Zhang, F.-L.; Zhang, L.; Zeng, D.-W.; Liao, S.; Fan, Y.; Champreda, V.; Runguphan, W.; Zhao, X.-Q. Engineering Yeast Cell Factories to Produce Biodegradable Plastics and Their Monomers: Current Status and Prospects. Biotechnol. Adv. 2023, 68, 108222. [Google Scholar] [CrossRef]
- Kitamoto, H.K.; Shinozaki, Y.; Cao, X.-H.; Morita, T.; Konishi, M.; Tago, K.; Kajiwara, H.; Koitabashi, M.; Yoshida, S.; Watanabe, T.; et al. Phyllosphere Yeasts Rapidly Break down Biodegradable Plastics. AMB Express 2011, 1, 44. [Google Scholar] [CrossRef]
- Naidu, S.; Singh, I.K.; Singh, A. Microbial Synthesis of Magnetic Nanoparticles for Plant Science and Agriculture. Plant Nano Biol. 2023, 4, 100036. [Google Scholar] [CrossRef]
- Sadh, P.K.; Chawla, P.; Kumar, S.; Das, A.; Kumar, R.; Bains, A.; Sridhar, K.; Duhan, J.S.; Sharma, M. Recovery of Agricultural Waste Biomass: A Path for Circular Bioeconomy. Sci. Total Environ. 2023, 870, 161904. [Google Scholar] [CrossRef]
- Chen, C.-C.; Dai, L.; Ma, L.; Guo, R. Enzymatic Degradation of Plant Biomass and Synthetic Polymers. Nat. Rev. Chem. 2020, 4, 114–126. [Google Scholar] [CrossRef]
- Banerjee, A.; Chatterjee, K. Enzymatic Degradation of Polymers: A Brief Review. Mater. Sci. Technol. 2014, 30, 567–573. [Google Scholar] [CrossRef]
- de Jesús Montaño López, J.; Duran, L.A.; Avalos, J. Physiological Limitations and Opportunities in Microbial Metabolic Engineering. Nat. Rev. Microbiol. 2021, 20, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Varjani, S.J.; Parameswaran, B.; Kumar, S.; Khare, S.K. (Eds.) Biosynthetic Technology and Environmental Challenges, 1st ed.; Energy, Environment, and Sustainability; Springer: Singapore, 2017; ISBN 9789811074332. [Google Scholar]
- Ahmed, I.A. Ethical Issues of Microbial Products for Industrialization. In Interdisciplinary Biotechnological Advances; Springer Nature: Singapore, 2023; pp. 393–411. ISBN 9789819917365. [Google Scholar]
- Meena, S.S.; Mohanty, A. Ethical, Patent, and Regulatory Issues in Microbial Engineering. In Engineering of Microbial Biosynthetic Pathways; Springer: Singapore, 2020; pp. 133–142. ISBN 9789811526039. [Google Scholar]
- Casadevall, A. The Future of Biological Warfare: The Future of Biological Warfare. Microb. Biotechnol. 2012, 5, 584–587. [Google Scholar] [CrossRef] [PubMed]
- Granato, E.T.; Meiller-Legrand, T.A.; Foster, K.R. The Evolution and Ecology of Bacterial Warfare. Curr. Biol. 2019, 29, R521–R537. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material | Microbe | Category | Reference |
|---|---|---|---|
| Alginate | Azotobacter sp. Pseudomonas sp. | Biopolymer | [5] |
| Bacterial cellulose | Komagataeibacter xylinus | Biopolymer | [95,96,97] |
| CdS CdS | Escherichia coli | QD | [81] |
| Pseudomonas putida | QD | [75,84] | |
| CdTe | Escherichia coli | QD | [76] |
| Fe3O4 | Magnetospirillum strain AMB-1 | NP | [67] |
| Magnetospirillum gryphiswaldense | [70] | ||
| Magnetospirillum magnetotacticum | [69] | ||
| Shewanella oneidensis | [30] | ||
| Gold | Shewanella oneidensis | NP | [52,53,54,55] |
| Polyhydroxyalkanoates | Cupriavidus necator | Biopolymer | [101] |
| Polylactic acid | Lactobacillus sp. | Biopolymer | [104] |
| Silver | Pseudomonas aeruginosa | NP | [49,52] |
| Fusarium oxysporum | NP | [51,58] | |
| Shewanella oneidensis | NP | [54,56] | |
| Succinic acid | Actinobacillus succinogenes | Monomer | [104,105,106,107,108] |
| TiO2 | Bacillariophyceae (diatoms) | Thin films | [94] |
| Spirulina platensis | NP | [54,63] | |
| Uranium phosphate | Pseudomonas sp. | Thin films | [93] |
| Xanthan gum | Xanthomonas campestris | Biopolymer | [100] |
| ZnO | Aspergillus niger | NP | [60,61] |
| ZnS | Fusarium oxysporum | QD | [82,83] |
| Factor | Traditional Synthesis | Microbial Synthesis |
|---|---|---|
| Energy Requirements | High temperatures and pressures | Ambient or mild conditions |
| Environmental Impact | High waste generation and toxic byproducts | Generally low waste and greener processes |
| Precision/Control | Moderate control over structure/composition | Polydispersity and uniformity remain ongoing challenges; high tunability via genetic/metabolic engineering |
| Raw Material Source | Petroleum-based and non-renewable | Renewable feedstocks (e.g., sugars and agricultural waste) |
| Scalability | Established large-scale industrial processes | Still challenging; limited by bioreactor design and yield |
| Cost | Higher due to being energy and purification intensive | Potentially lower, but requires optimization |
| Speed of Production | Fast, depending on the method | Slower and dependent on microbial growth and metabolic rates |
| Material Diversity | Broad, well-studied, and chemically intensive | Growing area of research, influenced by microbe strain |
| Biocompatibility | Often poor; requires surface modification | Often high, especially for medical/biological applications |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramachandran, R.; Macabenta, F.; Bettencourt, G.; Feng, S. From Microbes to Molecules: Synthetic Biology Approaches for Advanced Materials Design. BioChem 2025, 5, 12. https://doi.org/10.3390/biochem5020012
Ramachandran R, Macabenta F, Bettencourt G, Feng S. From Microbes to Molecules: Synthetic Biology Approaches for Advanced Materials Design. BioChem. 2025; 5(2):12. https://doi.org/10.3390/biochem5020012
Chicago/Turabian StyleRamachandran, Roshini, Frank Macabenta, Grace Bettencourt, and Shulammite Feng. 2025. "From Microbes to Molecules: Synthetic Biology Approaches for Advanced Materials Design" BioChem 5, no. 2: 12. https://doi.org/10.3390/biochem5020012
APA StyleRamachandran, R., Macabenta, F., Bettencourt, G., & Feng, S. (2025). From Microbes to Molecules: Synthetic Biology Approaches for Advanced Materials Design. BioChem, 5(2), 12. https://doi.org/10.3390/biochem5020012