

Free-Range Chickens Reared Within an Olive Grove Influenced the Soil Microbial Community and Carbon Sequestration

 , , , ,
, , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Management of Chickens
2.3. Evaluation of Animal Density and Feed Consumption
2.4. Soil Sampling
2.5. Estimation of the Aboveground and Belowground Herbaceous Biomass and Soil Analyses
2.6. Statistical Analysis
3. Results and Discussion
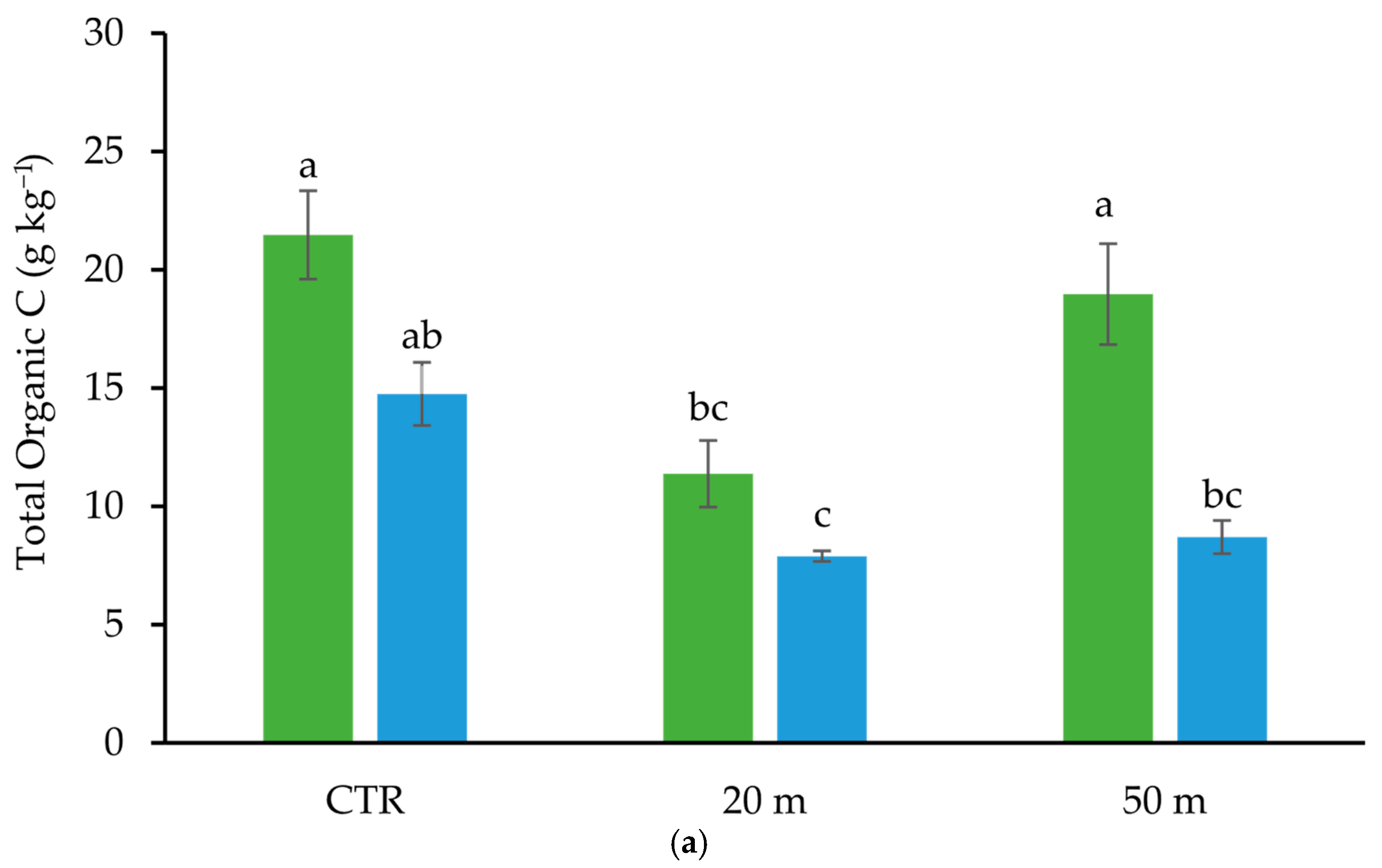
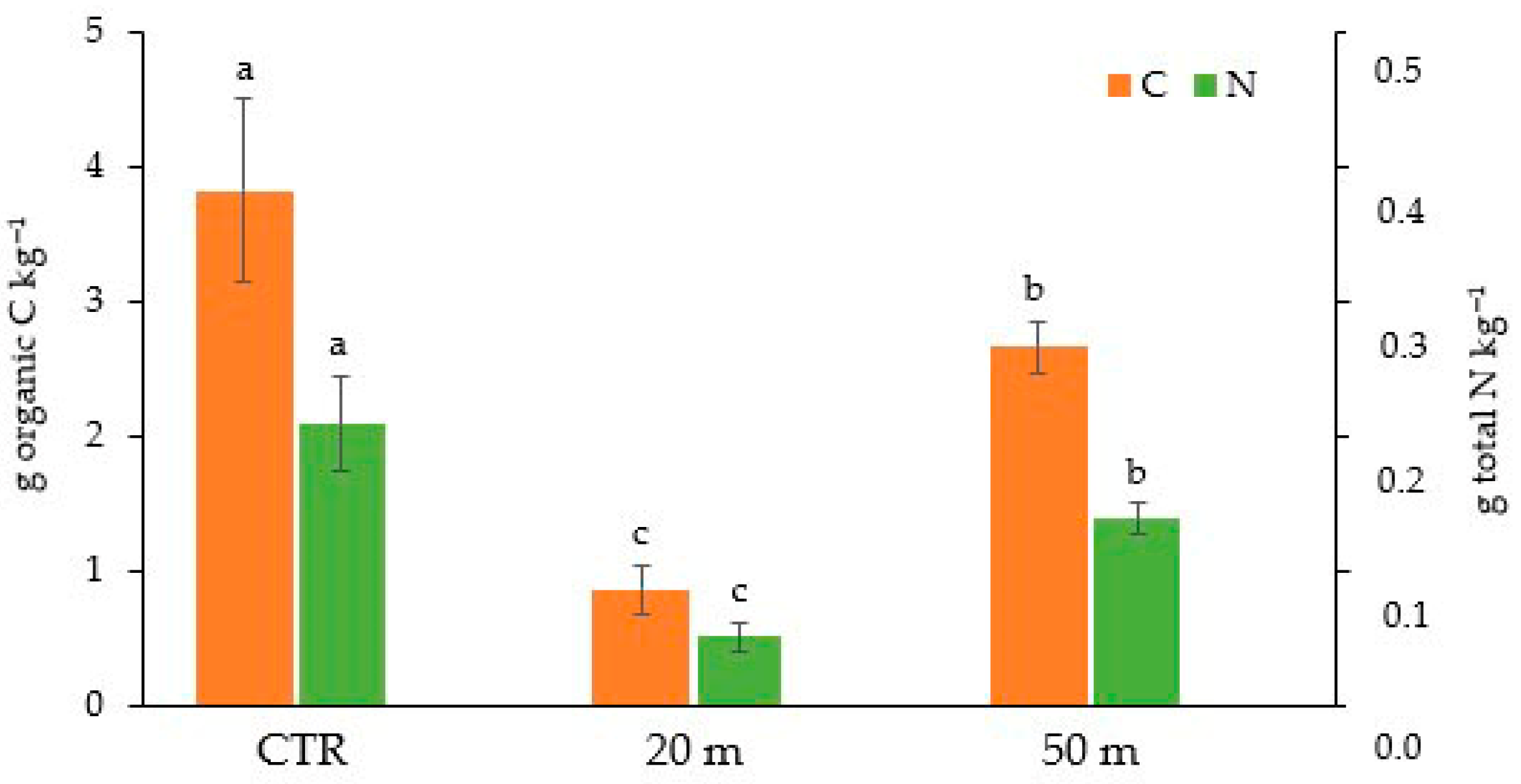
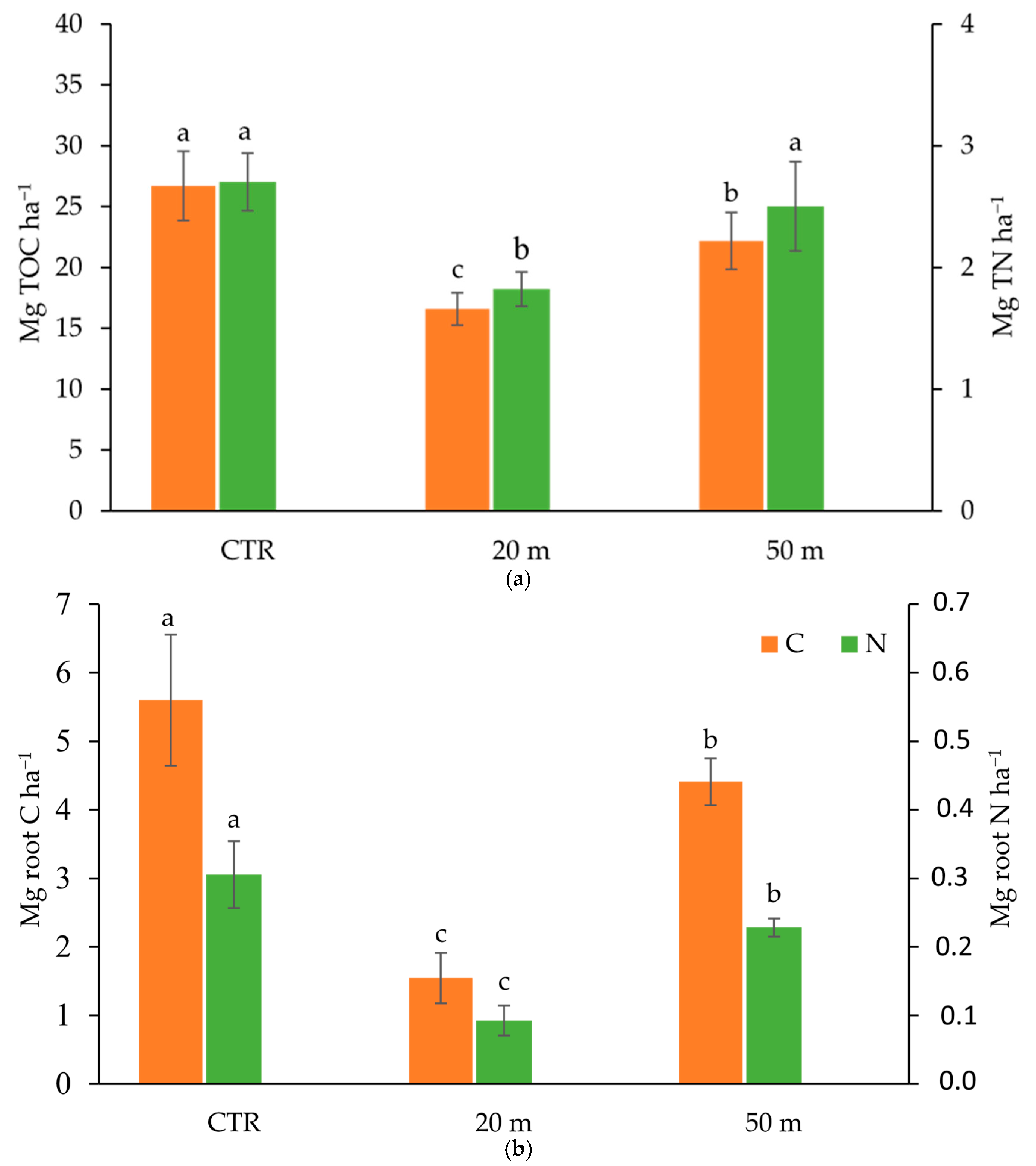
3.1. Grazing Effect on C and N Soil Concentrations and Stocks
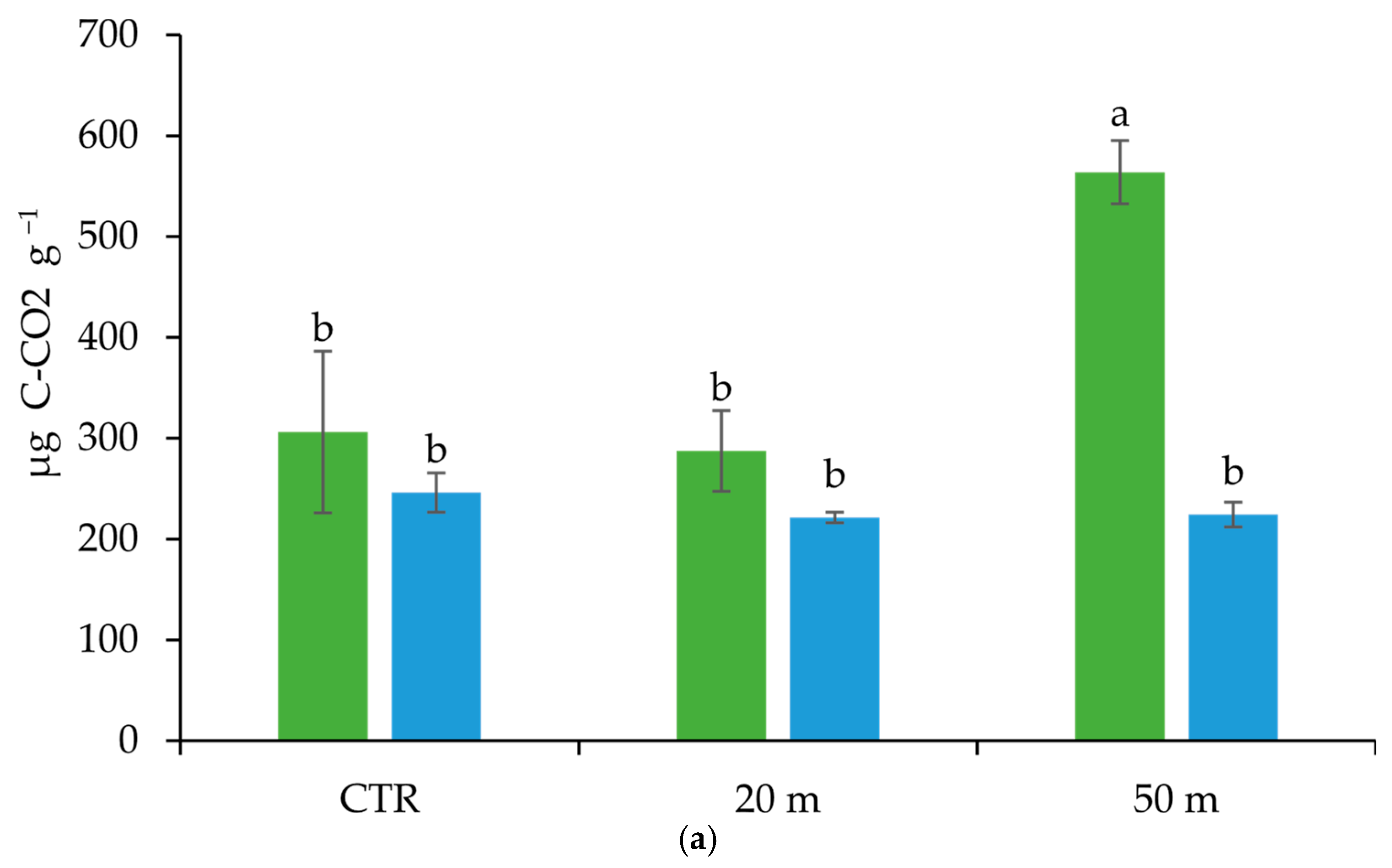
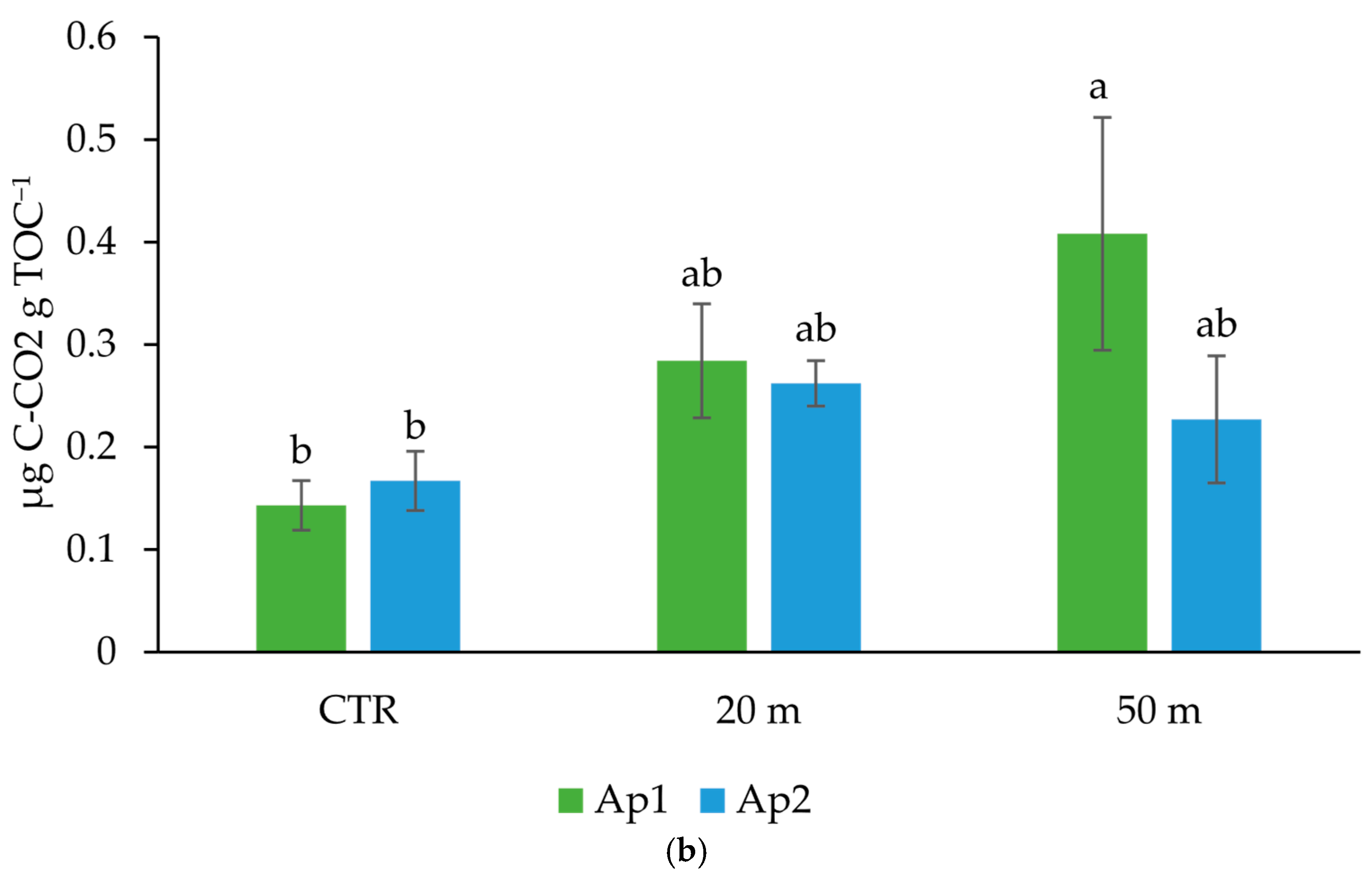
3.2. Grazing Effect on Microbial Community Amount, Activity, and Structure
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gouveia, K.G.; Vaz-Pires, P.; da Costa, P.M. Welfare assessment of broilers through examination of haematomas, foot-pad dermatitis, scratches and breast blisters at processing. Anim. Welf. 2009, 18, 43–48. [Google Scholar] [CrossRef]
- Dal Bosco, A.; Mugnai, C.; Mattioli, S.; Rosati, A.; Ruggeri, S.; Ranucci, D.; Castellini, C. Transfer of bioactive compounds from pasture to meat in organic free-range chickens. Poult. Sci. 2016, 95, 2464–2471. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, S.; Cartoni Mancinelli, A.; Dal Bosco, A.; Ciarelli, C.; Guarino Amato, M.; Angelucci, E.; Chiattelli, D.; Castellini, C. Intake of nutrients (polyunsaturated fatty acids, tocols, and carotenes) and storage efficiency in different slow-growing chicken genotypes reared in extensive systems. PLoS ONE 2022, 17, e0275527. [Google Scholar] [CrossRef] [PubMed]
- Skunca, D.; Tomasevic, I.; Nastasijevic, I.; Tomovic, V.; Djekic, I. Life cycle assessment of the chicken meat chain. J. Clean. Prod. 2018, 184, 440–450. [Google Scholar] [CrossRef]
- Paolotti, L.; Boggia, A.; Castellini, C.; Rocchi, L.; Rosati, A. Combining livestock and tree crops to improve sustainability in agriculture: A case study using the Life Cycle Assessment (LCA) approach. J. Clean. Prod. 2016, 131, 351–363. [Google Scholar] [CrossRef]
- Massaccesi, L.; Cartoni Mancinelli, A.; Mattioli, S.; De Feudis, M.; Castellini, C.; Dal Bosco, A.; Marongiu, L.; Agnelli, A. Geese reared in vineyard: Soil, grass and animals interaction. Animals 2019, 9, 179. [Google Scholar] [CrossRef]
- Chavan, S.B.; Newaj, R.; Rizvi, R.H.; Ajit; Prasad, R.; Alam, B.; Handa, A.K.; Dhyani, S.K.; Amit, J.; Tripathi, D. Reduction of global warming potential vis-à-vis greenhouse gases through traditional agroforestry systems in Rajasthan, India. Environ. Dev. Sustain. 2021, 23, 4573–4593. [Google Scholar] [CrossRef]
- Berton, M.; Huerta, A.; Trocino, A.; Bordignon, F.; Sturaro, E.; Xiccato, G.; Birolo, M. Life Cycle Assessment of broiler chicken production using different genotypes and low-input diets. In Proceedings of the 73rd Annual Meeting of the EEAP, 5–8 September 2022; Porto, Portugal; Wageningen Academic Publishers: Noordwijk, The Netherlands, 2022; p. 502. [Google Scholar]
- Cui, X.; Wang, Y.; Niu, H.; Wu, J.; Wang, S.; Schnug, E.; Rogasik, J.; Fleckenstein, J.; Tang, Y. Effect of long-term grazing on soil organic carbon content in semiarid steppes in Inner Mongolia. Ecol. Res. 2005, 20, 519–527. [Google Scholar] [CrossRef]
- Abdalla, M.; Hastings, A.; Chadwick, D.R.; Jones, D.L.; Evans, C.D.; Jones, M.B.; Rees, R.M.; Smith, P. Critical review of the impacts of grazing intensity on soil organic carbon storage and other soil quality indicators in extensively managed grasslands. Agr. Ecosyst. Environ. 2018, 253, 62–81. [Google Scholar] [CrossRef]
- Poeplau, C. Grassland soil organic carbon stocks along management intensity and warming gradients. Grass Forage Sci. 2021, 76, 186–195. [Google Scholar] [CrossRef]
- Yang, F.; Niu, K.; Collins, C.G.; Yan, X.; Ji, Y.; Ling, N.; Zhou, X.; Du, G.; Guo, H.; Hu, S. Grazing practices affect the soil microbial community composition in a Tibetan alpine meadow. Land Degrad. Dev. 2019, 30, 49–59. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Thorpe, A.S.; Brookshire, E.N.J. Livestock exclusion and belowground ecosystem responses in riparian meadows of eastern Oregon. Ecol. Appl. 2004, 14, 1671–1679. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Leemans, D.K.; Cook, R.; Hobbs, P.J. Seasonality of the soil biota of grazed and ungrazed hill grasslands. Soil Biol. Biochem. 1997, 29, 1285–1294. [Google Scholar] [CrossRef]
- Kohler, F.; Hamelin, J.; Gillet, F.; Gobat, J.M.; Buttler, A. Soil microbial community changes in wooded mountain pastures due to simulated effects of cattle grazing. Plant Soil 2005, 278, 327–340. [Google Scholar] [CrossRef]
- Kardol, P.; Dickie, I.A.; John, M.G.S.; Husheer, S.W.; Bonner, K.I.; Bellingham, P.J.; Wardle, D.A. Soil-mediated effects of invasive ungulates on native tree seedlings. J. Ecol. 2014, 102, 622–631. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Wardle, D.A. Herbivore-mediated linkages between aboveground and belowground communities. Ecology 2003, 84, 2258–2268. [Google Scholar] [CrossRef]
- Eldridge, D.J.; Delgado-Baquerizo, M.; Travers, S.K.; Val, J.; Oliver, I. Do grazing intensity and herbivore type affect soil health? Insights from a semi-arid productivity gradient. J. Appl. Ecol. 2017, 54, 976–985. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Wardle, D.A.; Yeates, G.W. Linking above-ground and below-ground interactions: How plant responses to foliar herbivory influence soil organisms. Soil Biol. Biochem. 1998, 30, 1867–1878. [Google Scholar] [CrossRef]
- Hu, H.; Chen, X.; Hou, F.; Wu, Y.; Cheng, Y. Bacterial and fungal community structures in loess plateau grasslands with different grazing intensities. Front. Microbiol. 2017, 8, 606. [Google Scholar] [CrossRef]
- Xu, H.; You, C.; Tan, B.; Xu, L.; Liu, Y.; Wang, M.; Xu, Z.; Sardans, J.; Peñuelas, J. Effects of livestock grazing on the relationships between soil microbial community and soil carbon in grassland ecosystems. Sci. Total Environ. 2023, 881, 163416. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, M.; Wang, X.; Wang, C. The impact of different grazing intensity and management measures on soil organic carbon density in Zhangye grassland. Sci. Rep. 2024, 14, 17556. [Google Scholar] [CrossRef] [PubMed]
- Napoli, R.; Paolanti, M.; Di Ferdinando, S. Atlante dei Suoli del Lazio; ARSIAL Regione Lazio: Rome, Italy, 2019; ISBN 978-88-904841-2-4. [Google Scholar]
- Akinola, O.S.; Onakomaiya, A.O.; Agunbiade, J.A.; Oso, A.O. Growth performance, apparent nutrient digestibility, intestinal morphology and carcass traits of broiler chickens fed dry, wet and fermented-wet feed. Livest. Sci. 2015, 177, 103–109. [Google Scholar] [CrossRef]
- Gould, I.J.; Quinton, J.N.; Weigelt, A.; De Deyn, G.B.; Bardgett, R.D. Plant diversity and root traits benefit physical properties key to soil function in grasslands. Ecol. Lett. 2016, 19, 1140–1149. [Google Scholar] [CrossRef] [PubMed]
- O’brien, S.L.; Jastrow, J.D.; Grimley, D.A.; Gonzalez-Meler, M.A. Moisture and vegetation controls on decadal-scale accrual of soil organic carbon and total nitrogen in restored grasslands. Glob. Change Biol. 2010, 16, 2573–2588. [Google Scholar] [CrossRef]
- Ma, Z.; Qin, W.; Wang, Z.; Han, C.; Liu, X.; Huang, X. A Meta-Analysis of Soil Organic Carbon Response to Livestock Grazing in Grassland of the Tibetan Plateau. Sustainability 2022, 14, 14065. [Google Scholar] [CrossRef]
- van Reeuwijk, L. Procedures for Soil Analysis, 6th ed.; ISRIC: Wageningen, The Netherlands; FAO: Rome, Italy, 2002. [Google Scholar]
- Anderson, T.H.; Domsch, K.H. The metabolic quotient for CO2 (qCO2) as a specific activity parameter to assess the effects of environmental conditions, such as pH, on the microbial biomass of forest soils. Soil Biol. Biochem. 1993, 25, 393–395. [Google Scholar] [CrossRef]
- Stazi, S.R.; Moscatelli, M.C.; Papp, R.; Crognale, S.; Grego, S.; Martin, M.; Marabottini, R. A multi-biological assay approach to assess microbial diversity in arsenic (as) contaminated soils. Geomicrobiol. J. 2017, 34, 183–192. [Google Scholar] [CrossRef]
- Massaccesi, L.; Benucci, G.M.N.; Gigliotti, G.; Cocco, S.; Corti, G.; Agnelli, A. Rhizosphere effect of three plant species of environment under periglacial conditions (Majella massif, Central Italy). Soil Biol. Biochem. 2015, 89, 184–195. [Google Scholar] [CrossRef]
- Marinari, S.; Marabottini, R.; Falsone, G.; Vianello, G.; Antisari, L.V.; Agnelli, A.; Massaccesi, L.; Cocco, S.; Cardelli, V.; Serrani, D.; et al. Mineral weathering and lessivage affect microbial community and enzyme activity in mountain soils. Appl. Soil Ecol. 2021, 167, 104024. [Google Scholar] [CrossRef]
- McKinley, V.; Peacock, A.D.; White, D.C. Microbial community PLFA and PHB responses to ecosystem restoration in tallgrass prairie soils. Soil Biol. Biochem. 2005, 37, 1946–1958. [Google Scholar] [CrossRef]
- Romaniuk, R.; Costantini, A.; Giuffré, L.; Nannipieri, P. Catabolic response and phospholipid fatty acid profiles as microbial tools to assess soil functioning. Soil Use Manag. 2016, 32, 603–612. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 30 November 2024).
- Dal Bosco, A.; Mugnai, C.; Sirri, F.; Zamparini, C.; Castellini, C. Assessment of a GPS to evaluate activity of organic chickens at pasture. J. Appl. Poult. Res. 2010, 19, 213–218. [Google Scholar] [CrossRef]
- Mohr, D.; Cohnstaedt, L.W.; Topp, W. Wild boar and red deer affect soil nutrients and soil biota in steep oak stands of the Eifel. Soil Biol. Biochem. 2005, 37, 693–700. [Google Scholar] [CrossRef]
- Bilotta, G.S.; Brazier, R.E.; Haygarth, P.M. The impacts of grazing animals on the quality of soils, vegetation, and surface waters in intensively managed grasslands. Adv. Agron. 2007, 94, 237–280. [Google Scholar] [CrossRef]
- Gai, X.; Zhong, Z.; Zhang, X.; Bian, F.; Yang, C. Effects of chicken farming on soil organic carbon fractions and fungal communities in a Lei bamboo (Phyllostachys praecox) forest in subtropical China. For. Ecol. Manag. 2021, 479, 118603. [Google Scholar] [CrossRef]
- Zhong, Y.Q.W.; Yan, W.M.; Shangguan, Z.P. Soil carbon and nitrogen fractions in the soil profile and their response to long-term nitrogen fertilization in a wheat field. Catena 2015, 135, 38–46. [Google Scholar] [CrossRef]
- Urra, J.; Alkorta, I.; Lanzén, A.; Mijangos, I.; Garbisu, C. The application of fresh and composted horse and chicken manure affects soil quality, microbial composition and antibiotic resistance. Appl. Soil Ecol. 2019, 135, 73–84. [Google Scholar] [CrossRef]
- Dawson, L.A.; Grayston, S.J.; Paterson, E. Effects of Grazing on the Roots and Rhizosphere of Grasses. In Grassland Ecophysiology and Grazing Ecology; Lemaire, G., Hodgson, J., de Moraes, A., Nabinger, C., de Faccio Carvalho, P.C., Eds.; CAB International: Wallingford, UK, 2000; pp. 61–84. [Google Scholar]
- Soares, P.R.; Guilherme, R.; Conceição, A.; Galhano, C. Soil macrofauna under laying hens’ grazed fields in two different agroecosystems in Portugal. Open Agric. 2023, 8, 20220172. [Google Scholar] [CrossRef]
- Hayes, M.H.B. Darwin’s “vegetable mould” and some modern concepts of humus structure and soil aggregation. In Earthworm Ecology; Satchell, J.E., Ed.; Chapman and Hall: London, UK; New York, NY, USA, 1983; pp. 19–33. [Google Scholar]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Dikinya, O.; Mufwanzala, N. Chicken manure-enhanced soil fertility and productivity: Effects of application rates. J. Soil Sci. Environ. Manag. 2010, 1, 46–54. Available online: http://www.academicjournals.org/JSSEM (accessed on 20 November 2024).
- Murphy, B.W. Impact of soil organic matter on soil properties—A review with emphasis on Australian soils. Soil. Res. 2015, 53, 605–635. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Fry, E.L.; Eldridge, D.J.; Vries, F.T.; Manning, P.; Hamonts, K.; Kattge, J.; Boenisch, G.; Singh, B.K.; Bardgett, R.D. Plant attributes explain the distribution of soil microbial communities in two contrasting regions of the globe. New Phytol. 2018, 219, 574–587. [Google Scholar] [CrossRef] [PubMed]
- Leff, J.W.; Bardgett, R.D.; Wilkinson, A.; Jackson, B.G.; Pritchard, W.J.; De Long, J.R.; Oakley, S.; Mason, K.E.; Ostle, N.J.; Johnson, D.; et al. Predicting the structure of soil communities from plant community taxonomy, phylogeny, and traits. ISME J. 2018, 12, 1794–1805. [Google Scholar] [CrossRef] [PubMed]
- Bossio, D.A.; Scow, K.M.; Gunapala, N.; Graham, K.J. Determinants of soil microbial communities: Effects of agricultural management, season, and soil type on phospholipid fatty acid profiles. Microbial. Ecol. 1998, 36, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kieft, T.L.; Ringelberg, D.B.; White, D.C. Changes in ester-linked phospholipid fatty acid profiles of subsurface bacteria during starvation and desiccation in a porous medium. Appl. Environ. Microbiol. 1994, 60, 3292–3299. [Google Scholar] [CrossRef]
- Pinkart, H.C.; Ringelberg, D.B.; Piceno, Y.M.; Macnaughton, S.J.; White, D.C. Biochemical approaches to biomass measurements and community structure analysis. Manual Environ. Microbiol. 2002, 2, 101–113. [Google Scholar]
- De Vries, F.T.; Hoffland, E.; Van Eekeren, N.; Brussaard, L.; Bloem, J. Fungal/bacterial ratios in grasslands with contrasting nitrogen management. Soil Biol. Biochem. 2006, 38, 2092–2103. [Google Scholar] [CrossRef]
- Schon, N.L.; Fraser, P.M.; Mackay, A.D.; Dickinson, N. Relationship between earthworm abundance, ecological diversity and soil function in pastures. Soil Res. 2021, 59, 767–777. [Google Scholar] [CrossRef]
- Yao, H.Y.; Jiao, X.D.; Wu, F.Z. Effects of continuous cucumber cropping and alternative rotations under protected cultivation on soil microbial community diversity. Plant Soil 2006, 284, 195–203. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CTR | 20 m | 50 m | |
|---|---|---|---|
| Chicken density (animals m−2) | - | 0.63 (0.33) a | 0.08 (0.05) b |
| Aboveground herbaceous biomass (g d.m. m−2) | 231.0 (33.1) c | 20.0 (1.2) a | 53.0 (4.6) b |
| Belowground plant biomass (g d.m. dm−3) | 2.2 (0.5) a | 0.6 (0.3) b | 1.9 (0.1) ab |
| N content of herbaceous biomass (g d.m. m−2) | 4.7 (0.4) c | 0.4 (0.0) a | 1.1 (0.9) b |
| Chicken excreta (g d.m. m−2 per cycle) | - | 889.6 (4.6) a | 154.8 (2.1) b |
| N excreted with faeces and urine (g d.m. m−2 per cycle) | - | 24.8 (3.0) a | 8.1 (1.0) b |
| CTR | 20 m | 50 m | ||||
|---|---|---|---|---|---|---|
| Ap1 | Ap2 | Ap1 | Ap2 | Ap1 | Ap2 | |
| nmol g−1 | ||||||
| Total EL-FAME | 525.99 (38.6) a | 434.7 ± 56.3 a | 495.2 (26.7) a | 451.0 (32.2) a | 588.45 (75.0) a | 258.1 (40.6) b |
| TB | 404.7 (36.1) a | 400.9 (63.1) a | 403.47 (21.9) a | 362.4 (25.1) a | 441.07 (57.6) a | 282.9 (66.4) a |
| G+ | 99.96 (8.2) a | 90.4 (13.2) a | 94.85 (4.8) a | 85.3 (5.2) a | 100.65 (17.2) a | 60.7 (13.6) a |
| G− | 113.84 (12.6) a | 110.4 (19.6) a | 84.41 (3.1) a | 73.8 (5.1) ab | 111.4 (13.3) a | 63.2 (10.2) b |
| ACT | 15.18 (1.9) a | 12.3 (2.4) a | 16.16 (1.4) a | 15.4 (2.2) a | 22.9 (5.0) a | 13.2 (3.2) a |
| TF | 121.28 (10.0) a | 139.9 (20.2) a | 91.73 (4.8) b | 88.6 (7.3) b | 147.38 (18.0) a | 68.35 (12.8) b |
| SF | 30.14 (9.0) ab | 21.3 (3.1) a | 15.5 (2.1) b | 14.4 (0.8) b | 36.28 (4.8) a | 10.6 (2.2) b |
| AMF | 32.38 (2.4) a | 30.0 (5.1) a | 16.6 (0.7) b | 19.6 (1.6) a | 27.25 (3.7) a | 21.5 (3.5) a |
| SFA | 190.35 (20.4) a | 188.3 (28.1) a | 208.05 (21.2) a | 187.9 (13.7) a | 189.74 (11.7) a | 144.4 (38.0) a |
| MUFA | 173.13 (20.5) ab | 203.3 (28.2) a | 134.38 (2.4) b | 120.2 (9.9) b | 197.25 (27.7) a | 101.9 (17.9) b |
| Iso | 49.79 (4.5) a | 42.1 (7.4) a | 43.59 (2.6) a | 44.8 (3.3) a | 51.99 (9.4) a | 30.1 (6.2) a |
| Anteiso | 50.18 (4.1) a | 48.3 (5.9) a | 51.26 (2.6) a | 40.5 (4.4) a | 48.81 (7.8) a | 30.8 (7.4) a |
| Cy | 32.5 (2.5) a | 25.8 (5.6) ab | 26.27 (2.1) a | 27.8 (2.1) a | 28.97 (3.7) a | 17.5 (3.5) b |
| Pre | 49.2 (3.8) a | 51.9 (7.6) a | 34.3 (1.6) b | 27.1 (1.8) b | 52.81 (6.8) a | 25.9 (4.4) b |
| ratios | ||||||
| G+/G− | 0.79 (0.0) b | 0.8 (0.0) b | 1.13 (0.1) a | 1.2 (0.1) a | 0.96 (0.0) a | 1.0 (0.1) a |
| TF/TB | 0.28 (0.0) b | 0.31 (0.0) a | 0.23 (0.0) c | 0.24 (0.0) b | 0.33 (0.0) a | 0.25 (0.0) ab |
| SFA/MUFA | 1.14 (0.0) b | 1.01 (0.05) b | 1.54 (0.1) a | 1.57 (0.04) a | 1.12 (0.0) b | 1.41 (0.10) a |
| Iso/Anteiso | 0.98 (0.0) a | 0.86 (0.03) a | 0.85 (0.0) b | 1.16 (0.19) a | 1.08 (0.0) a | 0.99 (0.04) a |
| Cy/Pre | 0.62 (0.0) ab | 0.53 (0.05) c | 0.77 (0.1) a | 1.02 (0.01) a | 0.49 (0.0) b | 0.82 (0.03) b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Massaccesi, L.; Marabottini, R.; Poesio, C.; Mattioli, S.; Castellini, C.; Agnelli, A. Free-Range Chickens Reared Within an Olive Grove Influenced the Soil Microbial Community and Carbon Sequestration. Soil Syst. 2025, 9, 69. https://doi.org/10.3390/soilsystems9030069
Massaccesi L, Marabottini R, Poesio C, Mattioli S, Castellini C, Agnelli A. Free-Range Chickens Reared Within an Olive Grove Influenced the Soil Microbial Community and Carbon Sequestration. Soil Systems. 2025; 9(3):69. https://doi.org/10.3390/soilsystems9030069
Chicago/Turabian StyleMassaccesi, Luisa, Rosita Marabottini, Chiara Poesio, Simona Mattioli, Cesare Castellini, and Alberto Agnelli. 2025. "Free-Range Chickens Reared Within an Olive Grove Influenced the Soil Microbial Community and Carbon Sequestration" Soil Systems 9, no. 3: 69. https://doi.org/10.3390/soilsystems9030069
APA StyleMassaccesi, L., Marabottini, R., Poesio, C., Mattioli, S., Castellini, C., & Agnelli, A. (2025). Free-Range Chickens Reared Within an Olive Grove Influenced the Soil Microbial Community and Carbon Sequestration. Soil Systems, 9(3), 69. https://doi.org/10.3390/soilsystems9030069