
Fractional Lévy Stable Motion from a Segmentation Perspective


Abstract
1. Introduction
2. Fractional Lévy Stable Motion
3. Segment Analysis of Diffusive Trajectories
3.1. Identification of Motion Switches
3.2. Types of Confined Motion
- •
- Brownian motion subordinated by special random processes;
- •
- coupled Langevin equations;
- •
- stochastic resetting.
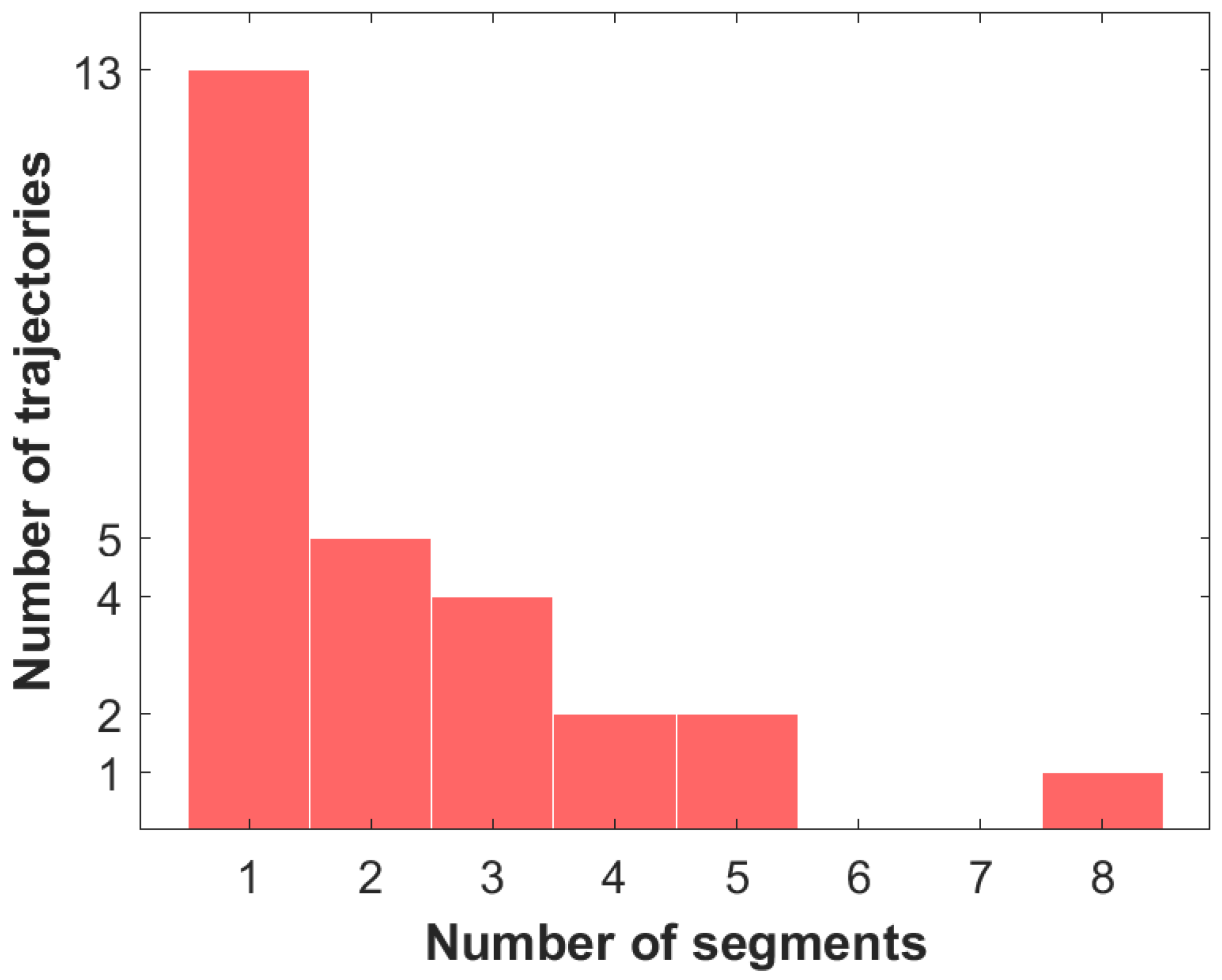
4. Transient Motion Classification Results for the Golding–Cox Data
5. Simulation Study
- •
- : a data sampling is free diffusion;
- •
- : a data sampling is subdiffusion;
- •
- : a data sampling is superdiffusion.
6. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saxton, M.J.; Jacobson, K. Single-particle tracking: Applications to membrane dynamics. Annu. Rev. Biophys. Biomol. Struct. 1997, 26, 373–399. [Google Scholar] [CrossRef]
- El Beheiry, M.; Dahan, M. High-speed localization microscopy and single-particle tracking. In Encyclopedia of Cell Biology; Bradshaw, R.A., Stahl, P.D., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 2, pp. 121–128. [Google Scholar]
- Manzo, C.; Garcia-Parajo, M.F. A review of progress in single particle tracking: From methods to biophysical insights. Rep. Prog. Phys. 2015, 78, 124601. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Welsher, K. Information-efficient, off-center sampling results in improved precision in 3D single-particle tracking microscopy. Entropy 2021, 23, 498. [Google Scholar] [CrossRef]
- Saxton, M.J. Anomalous diffusion due to obstacles: A Monte Carlo study. Biophys. J. 1994, 66, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Saxton, M.J. Single-particle tracking: The distribution of diffusion coefficients. Biophys. J. 1997, 72, 1744–1753. [Google Scholar] [CrossRef] [PubMed]
- Kowalek, P.; Loch-Olszewska, H.; Szwabiński, J. Classification of diffusion modes in single-particle tracking data: Feature-based versus deep-learning approach. Phys. Rev. E 2019, 100, 032410. [Google Scholar] [CrossRef]
- Parry, B.R.; Surovtsev, I.V.; Cabeen, M.T.; O’Hern, C.S.; Dufresne, E.R.; Jacobs-Wagner, C. The bacterial cytoplasm has glass-like properties and is fluidized by metabolic activity. Cell 2014, 156, 183–194. [Google Scholar] [CrossRef]
- Zhou, H.-X.; Rivas, G.; Minton, A.P. Macromolecular crowding and confinement: Biochemical, biophysical, and potential physiological consequences. Annu. Rev. Biophys. 2008, 37, 375–397. [Google Scholar] [CrossRef]
- Weber, S.C.; Spakowitz, A.J.; Theriot, J.A. Bacterial chromosomal loci move subdiffusively through a viscoelastic cytoplasm. Phys. Rev. Lett. 2010, 104, 238102. [Google Scholar] [CrossRef]
- Muñoz-Gil, G.; Garcia-March, M.A.; Manzo, C.; Martín-Guerrero, J.D.; Lewenstein, M. Single trajectory characterization via machine learning. New J. Phys. 2020, 22, 013010. [Google Scholar] [CrossRef]
- Han, D.; Korabel, N.; Chen, R.; Johnston, M.; Gavrilova, A.; Allan, V.J.; Fedotov, S.; Waigh, T.A. Deciphering anomalous heterogeneous intracellular transport with neural networks. eLife 2020, 9, e52224. [Google Scholar] [CrossRef] [PubMed]
- Stoev, S.; Taqqu, M.S. Simulation methods for linear fractional stable motion and FARIMA using the fast Fourier transform. Fractals 2004, 12, 95–121. [Google Scholar] [CrossRef]
- Weron, A.; Burnecki, K.; Mercik, S.; Weron, K. Complete description of all self-similar models driven by Lévy stable noise. Phys. Rev. E 2005, 71, 016113. [Google Scholar] [CrossRef] [PubMed]
- Golding, I.; Cox, E.C. RNA dynamics in live Escherichia coli cells. Proc. Natl. Acad. Sci. USA 2004, 101, 11310. [Google Scholar] [CrossRef] [PubMed]
- Golding, I.; Paulsson, J.; Zawilski, S.M.; Cox, E.C. Real-time kinetics of gene activity in individual bacteria. Cell 2005, 123, 1025. [Google Scholar] [CrossRef] [PubMed]
- Golding, I.; Cox, E.C. Physical nature of bacterial cytoplasm. Phys. Rev. Lett. 2006, 96, 098102. [Google Scholar] [CrossRef] [PubMed]
- Sikora, G.; Burnecki, K.; Wylomanska, A. Mean-squared-displacement statistical test for fractional Brownian motion. Phys. Rev. E 2017, 95, 032110. [Google Scholar] [CrossRef] [PubMed]
- Burnecki, K. Identification, Validation and Prediction of Fractional Dynamical Systems; Oficyna Wydawnicza Politechniki Wroclawskiej: Wroclaw, Poland, 2012. [Google Scholar]
- Balcerek, M.; Burnecki, K. Testing of fractional Brownian motion in a noisy environment. Chaos Solitons Fractals 2020, 140, 110097. [Google Scholar] [CrossRef]
- Klafter, J.; Lim, S.C.; Metzler, R. Fractional Dynamics. Recent Advances; World Scientific: Singapore, 2012. [Google Scholar]
- Burnecki, K. FARIMA processes with application to biophysical data. J. Stat. Mech. 2012, P05015. [Google Scholar] [CrossRef]
- Janczura, J.; Burnecki, K.; Muszkieta, M.; Stanislavsky, A.; Weron, A. Classification of random trajectories based on the fractional Lévy stable motion. Chaos Solitons Fractals 2022, 154, 111606. [Google Scholar] [CrossRef]
- Vega, A.R.; Freeman, S.A.; Grinstein, S.; Jaqaman, K. Multistep track segmentation and motion classification for transient mobility analysis. Biophys. J. 2018, 114, 1018. [Google Scholar] [CrossRef] [PubMed]
- Mandelbrot, B.B.; Van Ness, J.W. Fractional Brownian motions, fractional noises and applications. SIAM Rev. 1968, 10, 422–437. [Google Scholar] [CrossRef]
- Lutz, E. Fractional Langevin equation. Phys. Rev. E 2001, 64, 051106. [Google Scholar] [CrossRef] [PubMed]
- Watkins, N.W.; Credgington, D.; Sanchez, R.; Rosenberg, S.J.; Chapman, S.C. Kinetic equation of linear fractional stable motion and applications to modeling the scaling of intermittent bursts. Phys. Rev. E 2009, 79, 041124. [Google Scholar] [CrossRef] [PubMed]
- Samorodnitsky, G.; Taqqu, M.S. Stable Non-Gaussian Random Processes; Chapman & Hall: New York, NY, USA, 1994. [Google Scholar]
- Burnecki, K.; Sikora, G. Identification and validation of stable ARFIMA processes with application to UMTS data. Chaos Solitons Fractals 2017, 102, 456–466. [Google Scholar] [CrossRef]
- Dybiec, B.; Gudowska-Nowak, E.; Chechkin, A. To hit or to pass it over—Remarkable transient behavior of first arrivals and passages for Lévy flights in finite domains. J. Phys. A Math Theor. 2016, 49, 504001. [Google Scholar] [CrossRef]
- Song, W.; Chen, J.; Wang, Z.; Kudreyko, A.; Qi, D.; Zio, E. Remaining useful life prediction of lithium-ion battery based on adaptive fractional Lévy stable motion with capacity regeneration and random fluctuation phenomenon. Fractal Fract. 2023, 7, 827. [Google Scholar] [CrossRef]
- Teuerle, M.; Wyłomańska, A.; Sikora, G. Modeling anomalous diffusion by a subordinated fractional Lévy-stable process. J. Stat. Mech. 2013, 2013, P05016. [Google Scholar] [CrossRef]
- Beck, C.; Cohen, E.G. Superstatistics. Physica A 2003, 322, 267–275. [Google Scholar] [CrossRef]
- Beck, C. Generalized statistical mechanics for superstatistical systems. Phil. Trans. R. Soc. A 2011, 369, 453–465. [Google Scholar] [CrossRef]
- Ślęzak, J.; Burnecki, K.; Metzler, R. Random coefficient autoregressive processes describe Brownian yet non-Gaussian diffusion in heterogeneous systems. New J. Phys. 2019, 21, 073056. [Google Scholar] [CrossRef]
- Sadoon, A.; Wang, Y. Anomalous, non-Gaussian, viscoelastic, and age-dependent dynamics of histonelike nucleoid-structuring proteins in live Escherichia coli. Phys. Rev. E 2018, 98, 042411. [Google Scholar] [CrossRef]
- Huet, S.; Karatekin, E.; Tran, V.S.; Fanget, I.; Cribier, S.; Henry, J.P. Analysis of transient behavior in complex trajectories: Application to secretory vesicle dynamics. Biophys. J. 2006, 91, 3542–3559. [Google Scholar] [CrossRef]
- Das, R.; Cairo, C.W.; Coombs, D. A hidden Markov model for single particle tracks quantifies dynamic interactions between LFA-1 and the actin cytoskeleton. PLoS Comput. Biol. 2009, 5, e1000556. [Google Scholar] [CrossRef] [PubMed]
- Stanislavsky, A.A.; Weron, A. Confined random motion with Laplace and Linnik statistics. J. Phys. A Math. Theor. 2021, 54, 055009. [Google Scholar] [CrossRef]
- Stanislavsky, A.A.; Weron, A. Confined modes of single-particle trajectories induced by stochastic resetting. Phys. Rev. E 2023, 108, 044130. [Google Scholar] [CrossRef]
- Uhlenbeck, G.E.; Ornstein, L.S. On the theory of Brownian motion. Phys. Rev. 1930, 36, 823–841. [Google Scholar] [CrossRef]
- Kotz, S.; Kozubowski, T.; Podgórski, K. The Laplace Distribution and Generalizations: A Revisit with Applications to Communications, Economics, Engineering, and Finance; Birkhäuser: Boston, MA, USA, 2001. [Google Scholar]
- Kundu, D. Discriminating between normal and Laplace distributions. In Advances in Ranking and Selection, Multiple Comparisons, and Reliability. Statistics for Industry and Technology; Balakrishnan, N., Nagaraja, H.N., Kannan, N., Eds.; Birkhäuser: Boston, MA, USA, 2005; pp. 65–79. [Google Scholar]
- Anderson, D.N.; Arnold, B.C. Linnik distributions and processes. J. Appl. Probab. 1993, 30, 330–340. [Google Scholar] [CrossRef]
- Lampo, T.J.; Stylianidou, S.; Backlund, M.P.; Wiggins, P.A.; Spakowitz, A.J. Cytoplasmic RNA-protein particles exhibit non-Gaussian subdiffusive behavior. Biophys. J. 2017, 112, 532–542. [Google Scholar] [CrossRef]
- Neyman, J.; Pearson, E.S. On the problem of the most efficient tests of statistical hypotheses. Philos. Trans. R. Soc. Ser. A 1933, 231, 289–337. [Google Scholar]
- Neyman, J. Outline of a theory of statistical estimation based on the classical theory of probability. Philos. Trans. R. Soc. Ser. A 1937, 236, 333–380. [Google Scholar]
- Briane, M.V.; Kervrann, C.; Vimond, M. A statistical analysis of particle trajectories in living cells. Phys. Rev. E 2018, 97, 062121. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trajectory | x Coordinate | y Coordinate | ||||
|---|---|---|---|---|---|---|
| No. | ||||||
| 1 | 1.88 | 0.00 | 0.54 | 1.68 | −0.13 | 0.47 |
| 2 | 1.77 | −0.18 | 0.39 | 1.81 | −0.10 | 0.45 |
| 3 | 1.89 | −0.25 | 0.28 | 1.70 | −0.14 | 0.45 |
| 4 | 1.63 | −0.05 | 0.57 | 1.81 | −0.16 | 0.40 |
| 5 | 2.00 | −0.11 | 0.39 | 1.68 | −0.30 | 0.30 |
| 6 | 1.86 | −0.16 | 0.38 | 1.90 | −0.08 | 0.45 |
| 7 | 1.73 | −0.27 | 0.31 | 1.79 | −0.06 | 0.50 |
| 8 | 1.85 | −0.30 | 0.24 | 1.89 | −0.11 | 0.42 |
| 9 | 1.81 | −0.00 | 0.55 | 1.86 | −0.15 | 0.39 |
| 10 | 1.74 | −0.23 | 0.34 | 1.71 | −0.31 | 0.28 |
| 11 | 1.82 | −0.06 | 0.49 | 1.95 | −0.21 | 0.30 |
| 12 | 1.98 | −0.18 | 0.33 | 1.96 | −0.32 | 0.19 |
| 13 | 1.70 | −0.08 | 0.51 | 1.82 | −0.13 | 0.42 |
| 14 | 1.95 | −0.09 | 0.42 | 1.84 | −0.17 | 0.37 |
| 15 | 1.89 | −0.08 | 0.45 | 1.63 | 0.04 | 0.66 |
| 16 | 0.90 | −0.34 | 0.78 | 1.81 | 0.07 | 0.63 |
| 17 | 1.89 | −0.18 | 0.35 | 1.95 | −0.17 | 0.35 |
| 18 | 1.83 | −0.22 | 0.33 | 1.97 | −0.26 | 0.25 |
| 19 | 0.80 | −0.48 | 0.77 | 2.00 | −0.20 | 0.30 |
| 20 | 1.88 | −0.37 | 0.17 | 2.00 | −0.09 | 0.41 |
| 21 | 2.00 | −0.23 | 0.27 | 1.86 | −0.23 | 0.31 |
| 22 | 1.73 | −0.12 | 0.45 | 1.79 | −0.06 | 0.49 |
| 23 | 1.80 | −0.17 | 0.38 | 1.72 | −0.15 | 0.43 |
| 24 | 1.84 | −0.31 | 0.23 | 1.82 | −0.35 | 0.20 |
| 25 | 1.62 | −0.19 | 0.43 | 1.61 | −0.14 | 0.48 |
| 26 | 1.86 | −0.17 | 0.37 | 1.87 | −0.12 | 0.42 |
| 27 | 1.81 | −0.33 | 0.22 | 1.71 | −0.21 | 0.37 |
| Trajectory | Length | Length of | Segm. of | Confined | Confined | Confined |
|---|---|---|---|---|---|---|
| No. | of traj. | Long Parts | This Part | segm. 1 | segm. 2 | segm. 3 |
| 1 | 384 | 118 | ‘f’ | – | – | – |
| 2 | 490 | 338 | ‘fcfci’ | xy:lin,norm | xy: normal | – |
| 3 | 488 | 416 | ‘c’ | xy: normal | – | – |
| 4 | 1600 | 969 | ‘cfifcfcf’ | xy:lap,norm | xy:norm,lin | xy: normal |
| 5 | 399 | 67 | ‘c’ | xy: normal | – | – |
| 6 | 483 | 172 | ‘f’ | – | – | – |
| 7 | 475 | 329 | ‘c’ | xy: normal | – | – |
| 8 | 479 | 340 | ‘fcfc’ | xy:norm,lap | xy: normal | – |
| 9 | 479 | 141 | ‘fc’ | xy:lap,norm | – | – |
| 10 | 447 | 100 | ‘if’ | – | – | – |
| 11 | 419 | 52 | ‘c’ | xy: normal | – | – |
| 12 | 486 | 326 | ‘ifc’ | xy: normal | – | – |
| 13 | 482 | 288 | ‘c’ | xy: normal | – | – |
| 14 | 488 | 386 | ‘c’ | xy:lap,norm | – | – |
| 15 | 140 | 87 | ‘f’ | – | – | – |
| 16 | 461 | 70 | ‘fc’ | xy:lin,norm | – | – |
| 17 | 477 | 147 | ‘fcf’ | xy: normal | – | – |
| 18 | 368 | 108 | ‘cic’ | xy: normal | xy:lin,norm | – |
| 19 | 375 | 66 | ‘c’ | xy:lin,lap | – | – |
| 20 | 330 | 40 | ‘f’ | – | – | – |
| 21 | 412 | 74 | ‘c’ | xy: normal | – | – |
| 22 | 491 | 491 | ‘cfcfc’ | xy: normal | xy:norm,lin | xy: normal |
| 23 | 1628 | 417 | ‘cf’ | xy: normal | – | – |
| 24 | 1074 | 87 | ‘c’ | xy: normal | – | – |
| 25 | 680 | 273 | ‘fcic’ | xy:lap,norm | xy: normal | – |
| 26 | 921 | 200 | ‘cf’ | xy: normal | – | – |
| 27 | 1144 | 431 | ‘cfc’ | xy: normal | xy: normal | – |
| Cases | Subdiffusion | Free Diffusion | Superdiffusion |
|---|---|---|---|
| (a) | 57.8% | 42.2% | 0 |
| (b) | 52.4% | 47.6% | 0 |
| (c) | 44.4% | 55.6% | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stanislavsky, A.A.; Weron, A. Fractional Lévy Stable Motion from a Segmentation Perspective. Fractal Fract. 2024, 8, 336. https://doi.org/10.3390/fractalfract8060336
Stanislavsky AA, Weron A. Fractional Lévy Stable Motion from a Segmentation Perspective. Fractal and Fractional. 2024; 8(6):336. https://doi.org/10.3390/fractalfract8060336
Chicago/Turabian StyleStanislavsky, Aleksander A., and Aleksander Weron. 2024. "Fractional Lévy Stable Motion from a Segmentation Perspective" Fractal and Fractional 8, no. 6: 336. https://doi.org/10.3390/fractalfract8060336
APA StyleStanislavsky, A. A., & Weron, A. (2024). Fractional Lévy Stable Motion from a Segmentation Perspective. Fractal and Fractional, 8(6), 336. https://doi.org/10.3390/fractalfract8060336