1. Introduction
Describing the evolution of the population using the logistic growth model has an extensive research history. Such a model has been incorporated in numerous fields of biological applications, such as modeling yeast cultures [
1], describing the human population [
2], investigating a population of microorganisms [
3], modeling the growth of fish school populations [
4] and many other applications, presented in [
5,
6].
Considering the logistic model with constant environmental coefficients, the expression of the population behavior is well known in the literature—see, for example, the works of Banks [
5] and Broun [
6]. However, it is unrealistic to ignore the variation of the biotic environment which arises from the evolution of the ecosystem. Accordingly, Mayer and Ausubel [
7] investigated the behavior of the human population utilizing the logistic model with a constant intrinsic growth rate, gradually increasing the carrying capacity. Their analysis provided the numerical solution for the population and shows very good agreement with experimental data of England and Japan populations. Similarly, Al-Moqbali et al. [
8] investigated the stability along with obtaining the numerical solution of the prey–predator model with assuming a variable carrying capacity and fixed growth rate.
Assuming time-varying coefficients, the exact solution of the logistic model is no longer available, except in very special cases, so the numerical techniques must be adopted. However, although such a method provides expressions of the population behavior, it shows limited information for the evolution of the population in view of the influence of the environmental coefficients. In order to have a better understanding, an analytical approximate expression of the population evolution may be obtained via applying a multi-time scaling technique. Such a technique is well established in the literature for various fields—see, for example, [
9,
10,
11,
12]. Thus, Shepherd et al. [
13] successfully applied the multi-time scaling technique to obtain the analytical expression of the logistic model, assuming a slow variation of the carrying capacity and a stationary growth rate. Their calculations were compared well with the exact solution. For the general case that the carrying capacity and growth rate both vary with time, Grozdanovski et al. [
14] and Shepherd et al. [
15] developed the methodology of applying the multi-time scaling method and obtained the analytical approximation for the population. They validated their result through comparing with the numerical solution under which was shown a very good accuracy with the analytical one. Following the calculation of [
14], Idlango et al. [
16,
17,
18] investigated the logistic model with harvesting, displaying the Allee effect and saturating harvesting. Their analysis shows the analytical expression for the population, in explicit and implicit forms, in the situation of subcritical and supercritical harvesting. These findings were compared with the numerical solution and showed favorable accuracy. Recently, Alsharidi et al. [
19] investigated the anaerobic digestion model and obtained the expression by applying a parallel technique of those in [
16,
17,
18].
Therefore, we aim in this paper to investigate the slow logistic model with asymmetric variation between the carrying capacity and the growth rate. This will allow the possibility of a clear separation in evolution of the environmental coefficients. We consider the multi-time scaling technique together with the perturbation approach to construct the analytical approximate expression of the population behavior. In addition, since the exact solution for our assumption is not available in the literature, we consider the numerical simulation using a fourth-order Runge–Kutta technique; we compare with the analytical approximation and find that the accuracy between the two methods is very good indeed.
2. Analytical Analysis
The non-autonomous single species population model may be written in the form,
where
and
represent the population density, growth rate and carrying capacity, respectively; all of them depend on the variable
T, while
is the initial population when
. Note that an explicit solution of (
1) may be written as
but due to the integral terms in (
2), a full explicit solution to the population is only available for very special choices of
and
functions. Otherwise, a numerical technique must be employed.
Before constructing our analysis, we first non-dimensionalize the non-autonomous logistic model (
1) by assuming
and
varying on the time scales
and
, respectively. So, the functions
and
may be expressed in dimensionless form as follows
where
r and
k are the dimensionless growth rate and carrying capacity, respectively, while
and
are the characteristic constants. Note that assuming constant environments, i.e.,
R and/or
K are constant, which means that
(and hence
) and/or
(and hence
).
Regarding the dimensionless population
, we introduce the dimensionless time by
and the dimensionless population by
. Thus, in a view of the above scalings, the dimensionless logistic model (
1) can be expressed as
where
is the dimensionless initial population corresponding to
.
At this stage, and are assumed to be varying on hierarchical time scales. So, there are two situations that are identified, which are and . In what follows, we consider analyzing the situation where , and we exhibit the solution of the population evolution when in the Discussion section.
Note that, from (
4), when the time scales
and both of them are large relative to
, the variation of
is smaller than the small
. Accordingly, the hierarchical relationship in variation between the coefficients might be established based on one small parameter as follows:
where
.
Applying (
5) into (
4) yields to the general dimensionless population model as
3. Multi-Time Scaling
Since the model (
6) involves three time scales, which are the ordinary time variable
t and the two slow time variables,
and
, we adopt the multi-time scaling method by presenting the generalized ordinary time variable,
, and the two slow time variables,
and
, as
where the subscripts
k and
r refer to the time scale for the carrying capacity and growth rate, respectively. Note that
and
are expected to be positive, continuous and differentiable functions on
and
, with
and
, respectively.
With respect to
, the derivative of those in (
7) are given by
and we point out, from (
8), that the relationship between
and
t as well as
and
are governed by the functions
and
. Accordingly, we assume
and
to ensure that such relationships are monotonic. In a view of the above, we express the population solution
as a function
of these variables. With the application of the chain rule, the differential Equation (
6) is translated to multi-timing scaled logistic equation as
where
and
represent the partial derivatives of
and
, respectively. Therefore, the multi-time scaling is successfully applied to the differential Equation (
6) and such an equation becomes a partial differential equation for the unknown
. Since the parameter
, which governs the order of the coefficients variation, appears explicitly in (
9) rather than implicitly in (
6), the perturbation method based on a small
is employed to explicitly obtain the analytical approximate expression for the population of (
6) that is valid for all
.
4. Perturbation Analysis
In what follows, we apply the perturbation approach based on a small
to the partial differential Equation (
9). This yields to express
in power of
as
then substituting (
10) into (
9) and equating the coefficients of like power of
give the leading and higher orders terms as
Solving (
11) for
gives
where
and
are given by
and
is an arbitrary function of
and
to be determined later.
In a similar manner, solving (
12), on applying (
14), we obtain the particular solution for
as
where the primes in
and
represent the derivative with respect to
.
It is important to note that as
,
, given in (
14), reaches the carrying capacity limit
, and in (
16),
reaches
. However, the rate of convergence of
is exponential, while that of
is not. This asymmetry arises due to the presence of a quadratic polynomial function of
in (
16). Consequently, we argue that the rate of convergence of
must be aligned to reach its limit at the same time as that of
. To achieve this, we apply parallel justifications to those of [
16] and separately set the coefficients of
and
in (
16) to zero. This yields
Thus, the first of (
17) leads to concluding that
, while in the second, we only have to select
to satisfy the equation, and hence
is constant. In particular and for simplicity, we may choose
and, accordingly, the first of (
15) becomes
.
Considering the above observations and results, (
16) is reduced to
Solving the second correction term, given in (
13), we obtain the solution of
as follows
where the primes in
and
denote the derivative with respect to
, while in
, they represent the derivative with respect to
. Applying analogous observations to that in (
16), we conclude that
, and hence
, where
c is an arbitrary constant to be found using the initial condition, given in (
6). Furthermore,
, so
is constant, and for simplicity we choose
. Consequently, we conclude form the second of (
15) that
and accordingly, the first and the second of (
8) become
defining the variables
and
completely in terms of the growth rate function.
With respect to the observation above, (
19) is reduced to
To recapitulate, applying (
14), (
18) and (
21) into the expansion (
10) yields, on considering the results arising from the observations,
where the prime denotes to the derivative with respect to
. Since our expansion, given in (
10), is built with
and
, we assume that
c in (
22) follows a similar manner and take the form
. Thus, with
c expansion, (
22) yields, on expansion of the like power of
,
where the function
is given by
and the constants
and
are given by substituting the initial condition (
6) into (
23) on equating the like power of
and then are solved. Thus, these take the form as
5. Results and Discussion
In this paper, the analytical approximate expression for the population of the slow logistic model with the non-parallel variation of the coefficients is readily and explicitly obtained, and the expansion (
23) represents the result. It should be noted that the perturbation expansion (
10) is limited to the first three terms of the series, and this is due to the observation that the three-term solution provides a remarkable accuracy without swell of the expression. In the limit of
along with considering a constant coefficients, (
23) reduces to an exact solution—see, for example, [
5]. In addition, when
, the population in (
23) reaches
varying close to the carrying capacity,
, where
is given in the second of (
20). Note that for the long term, the slope of the carrying capacity trajectory becomes zero, i.e.,
(and hence
. Thus, the population
will eventually reach
.
The relationship between the variables
and
is described in (
20), and when the growth rate is assumed constant, for example,
, then
and
, where
. Such a relationship is realistic in describing the population through a variation of a biotic environment.
Since there is no exact solution for our assumption provided in the literature, we consider obtaining a numerical solution of our system, utilizing the fourth-order Runge–Kutta technique. Then, such a result is compared with the analytical approximation to validate the accuracy. To do that, we first consider that the carrying capacity,
is a periodic function, while the growth rate,
, is an exponential decay function. These are given by
Accordingly,
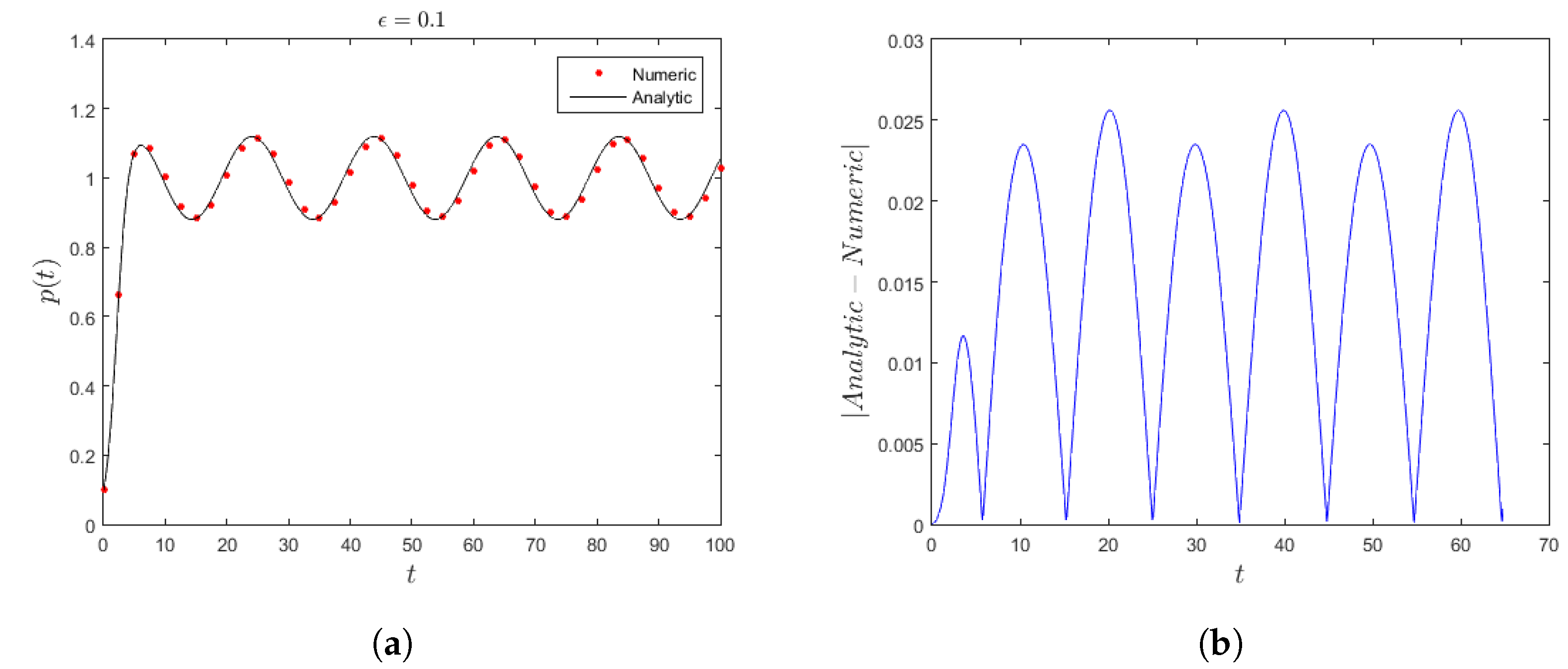
Figure 1a, generated from the analytical approximation (
23) (solid curve) along with the numerical solution (dotted curve), displays the population evolution from
for slowly varying
and
coefficients, given in (
27). It can clearly be seen that the comparison between the two methods shows very good agreement for
. In the same context,
Figure 1b, obtained by measuring the absolute difference between the approximate and numerical solution as
, displays that the maximum discrepancy is less than
, which is a favorable accuracy.
Note that we assume, earlier in this paper, that
. So, it is convenient to consider a solution for the population evolution when
. Consequently, applying analogous line of the above calculations, we reach the population expression as follows:
where
and
are given by
respectively.
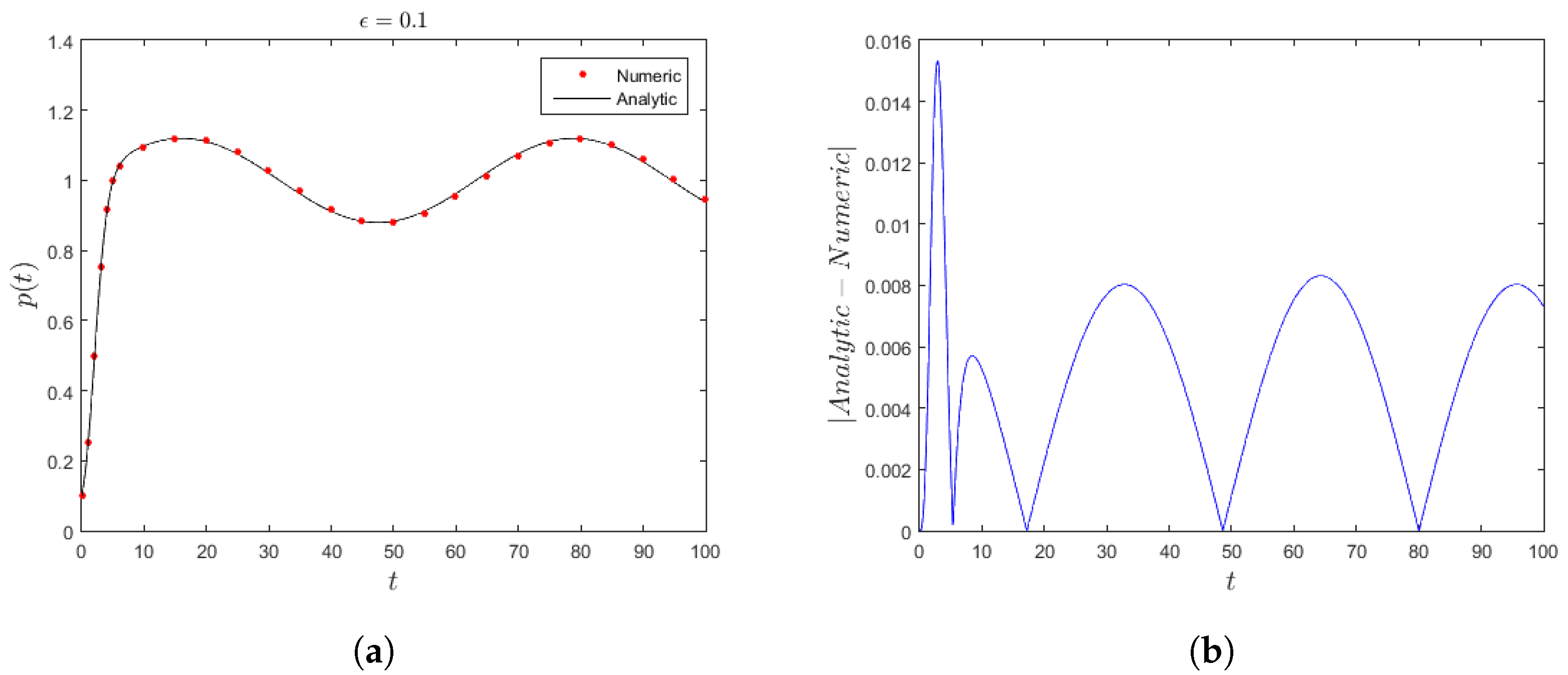
Applying (
27) in (
28) together with considering the fourth-order Runge–Kutta technique for the numerical result generates
Figure 2a, which displays the population evolution from
to the equilibrium state where the coefficients very slowly. The accuracy between the two methods is very good for
. In addition,
Figure 2a shows that the variation of the carrying capacity is much slower than that in
Figure 1a for the same period of time. This is due to changing the order of the coefficients’ variation. Moreover, for
, the absolute difference between the approximate solution and the numerical simulation is less than
as shown in
Figure 2b.


{kind=link}
{kind=link}