
Integrated Rice–Fish Culture Alters the Bacterioplankton Community and Its Assembly Processes

 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experiment Design and Sample Collection
2.2. Detection of Nutrients in Water
2.3. 16S rDNA Sequencing
2.4. Statistical Analysis
3. Results
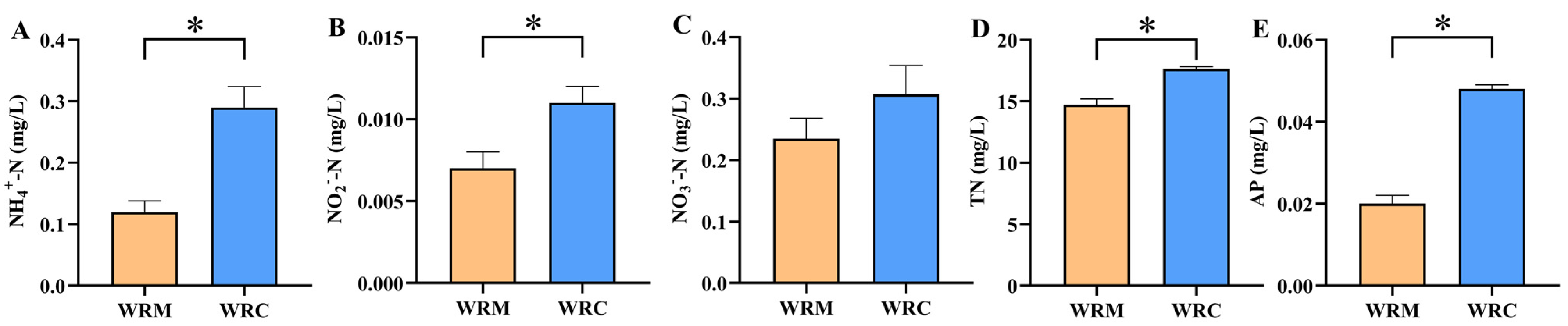
3.1. Water Physicochemical Properties
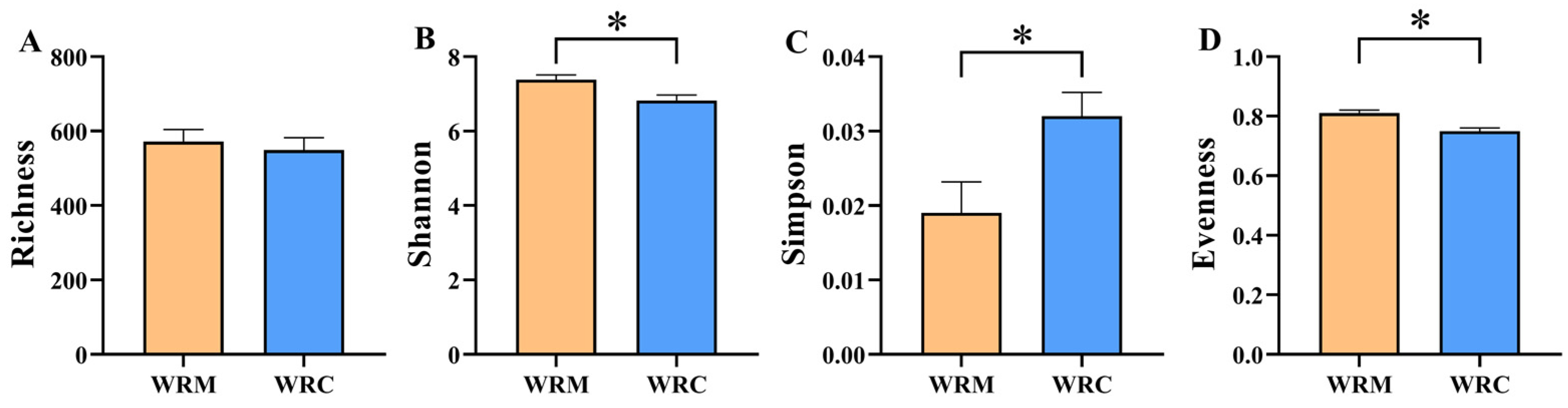
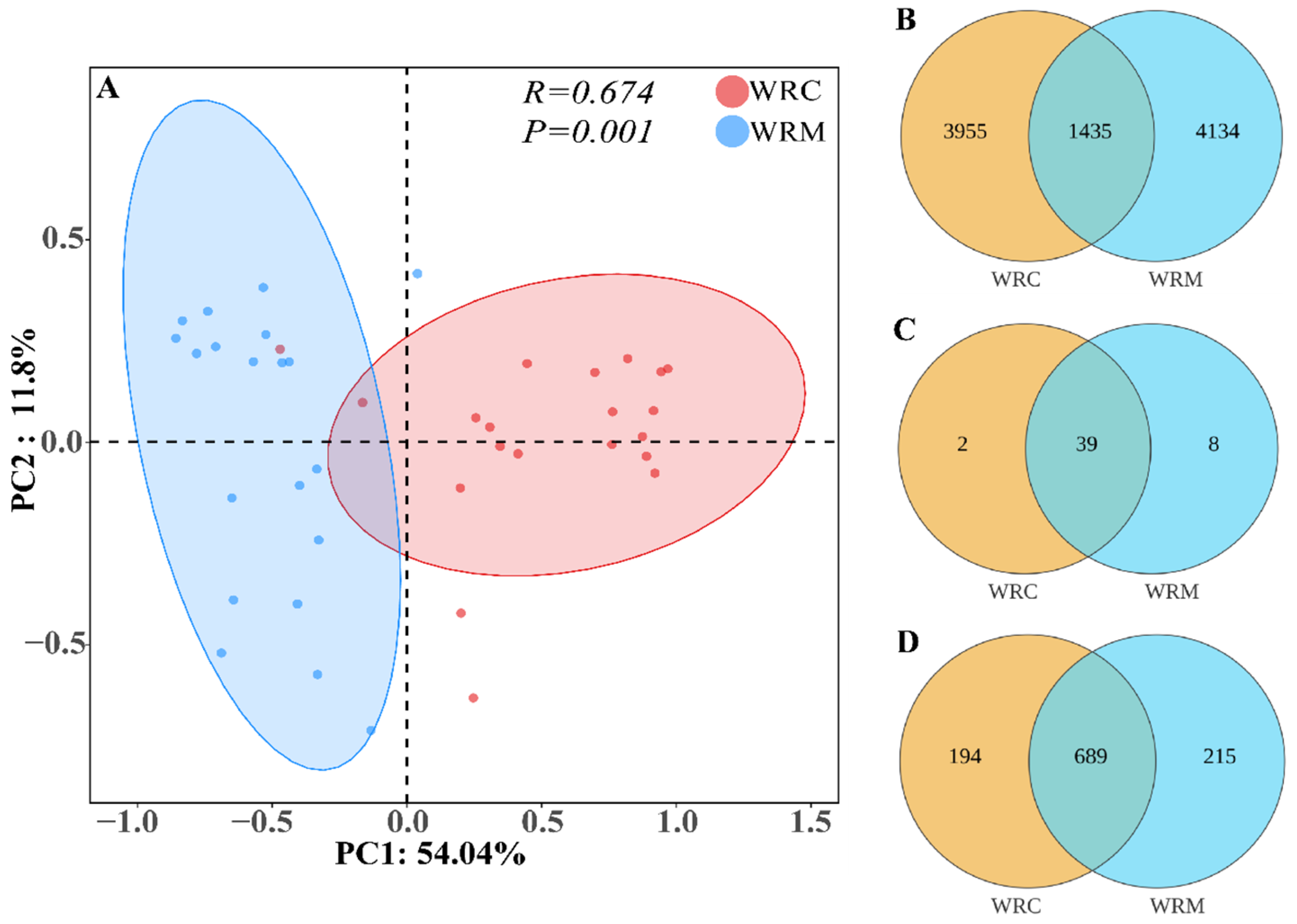
3.2. Alpha and Beta Diversities of Bacteria in Paddy Water
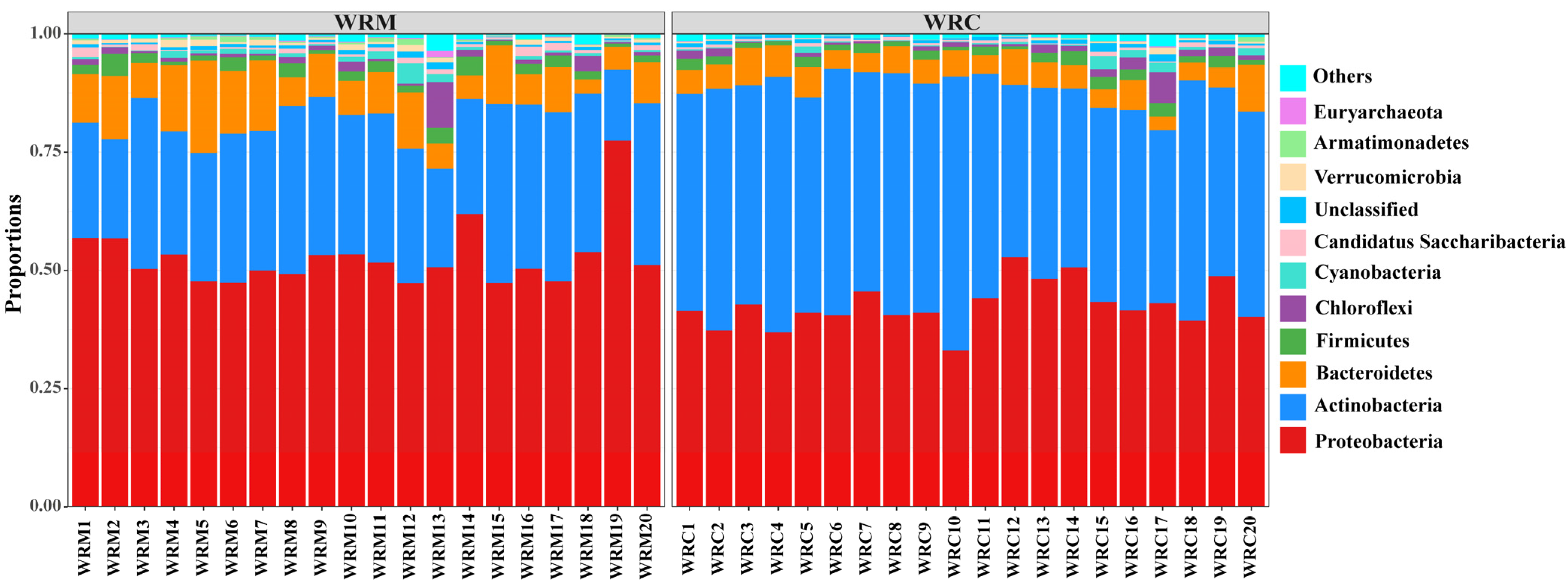
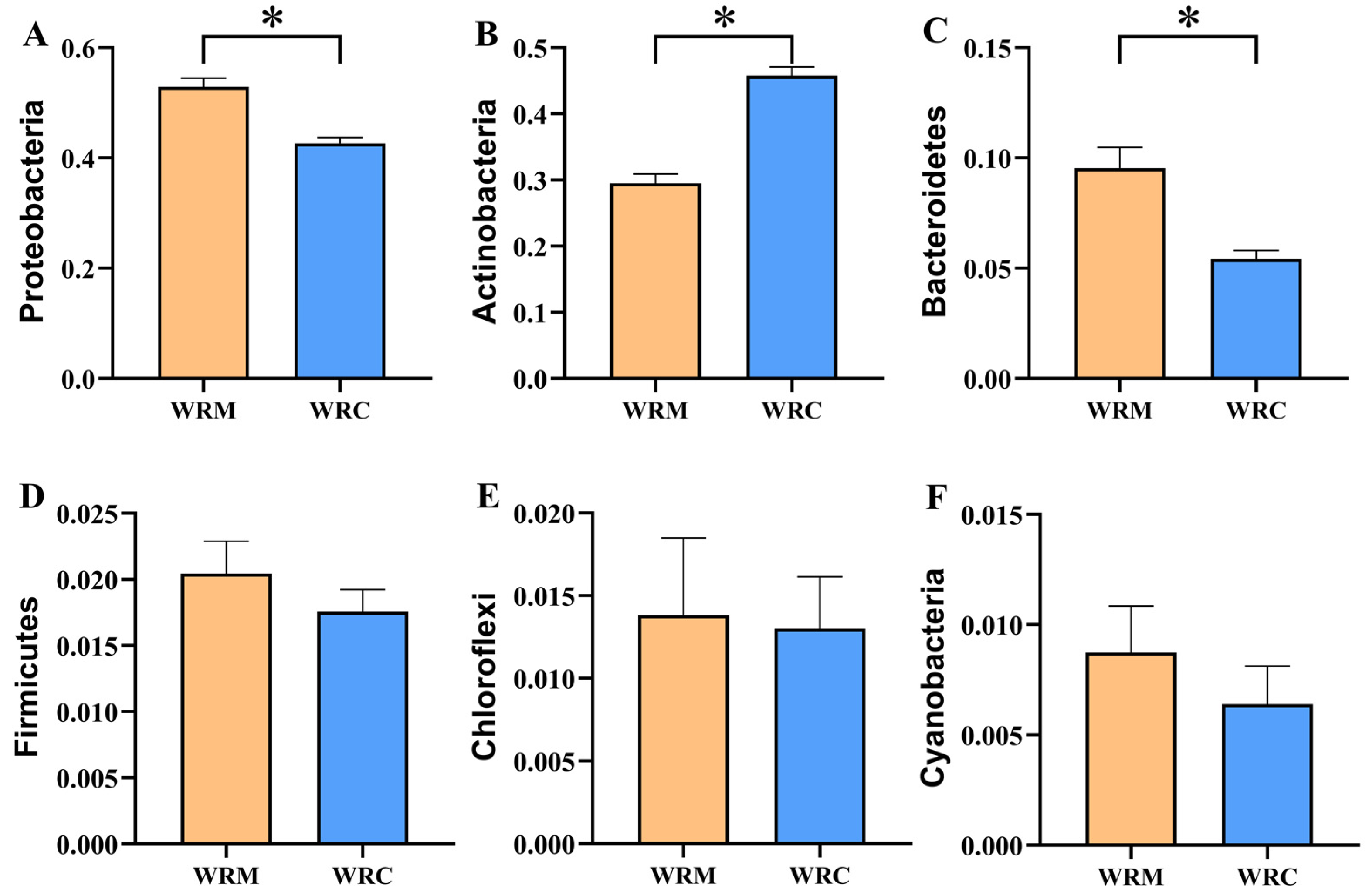
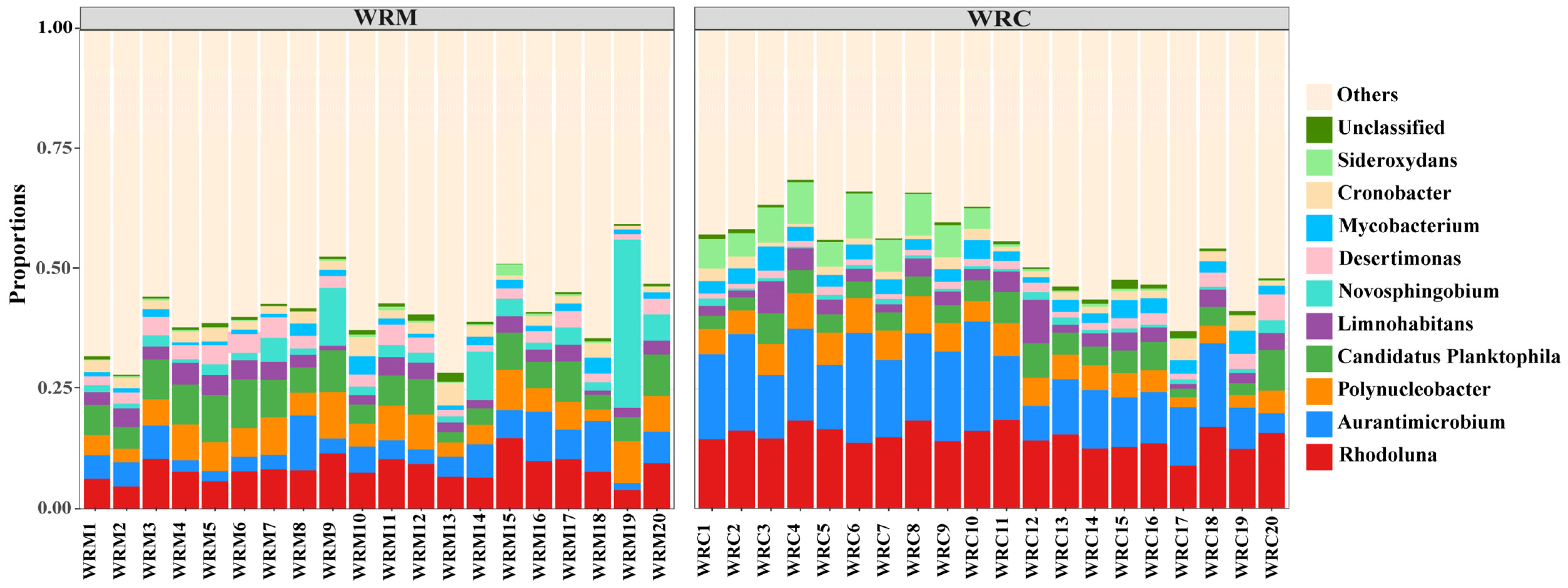
3.3. Bacterial Composition in Paddy Water
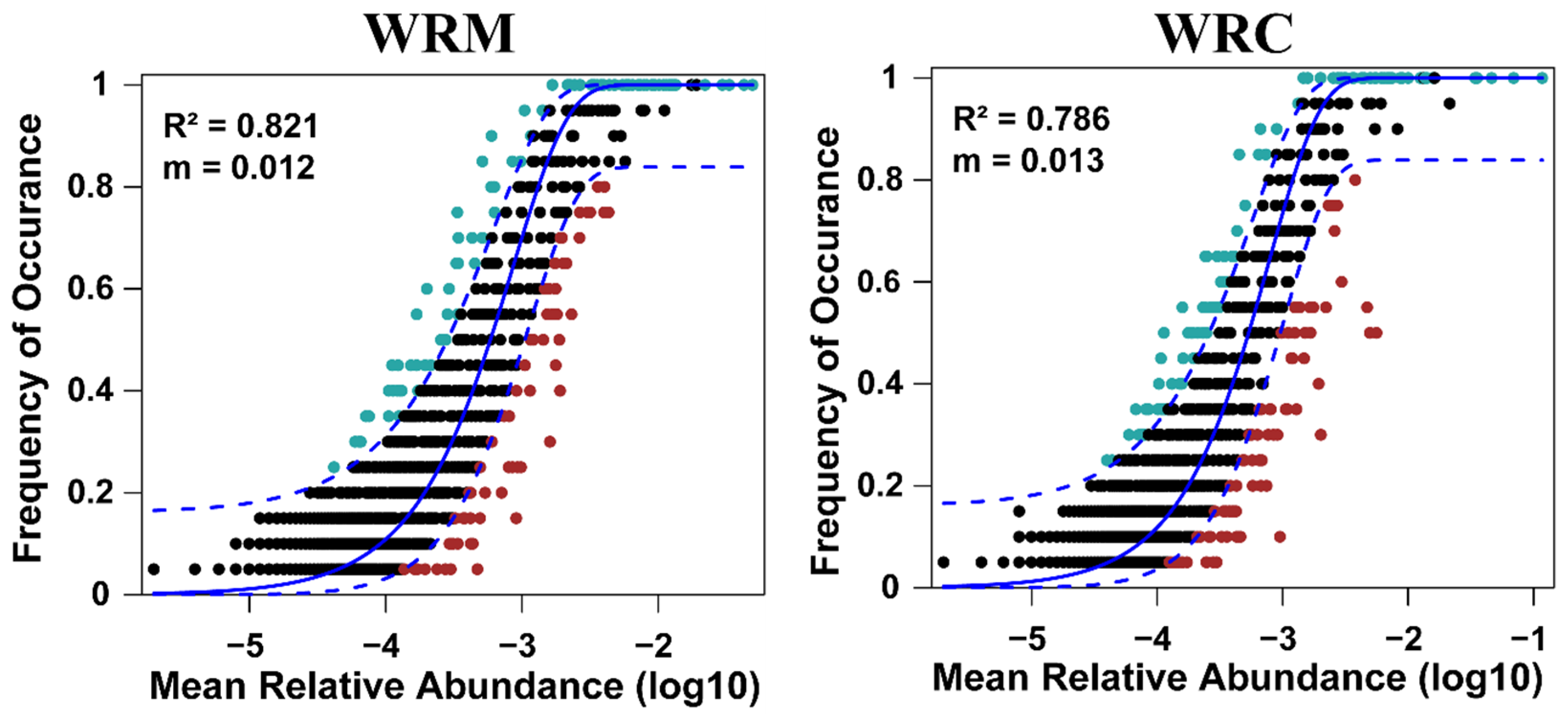
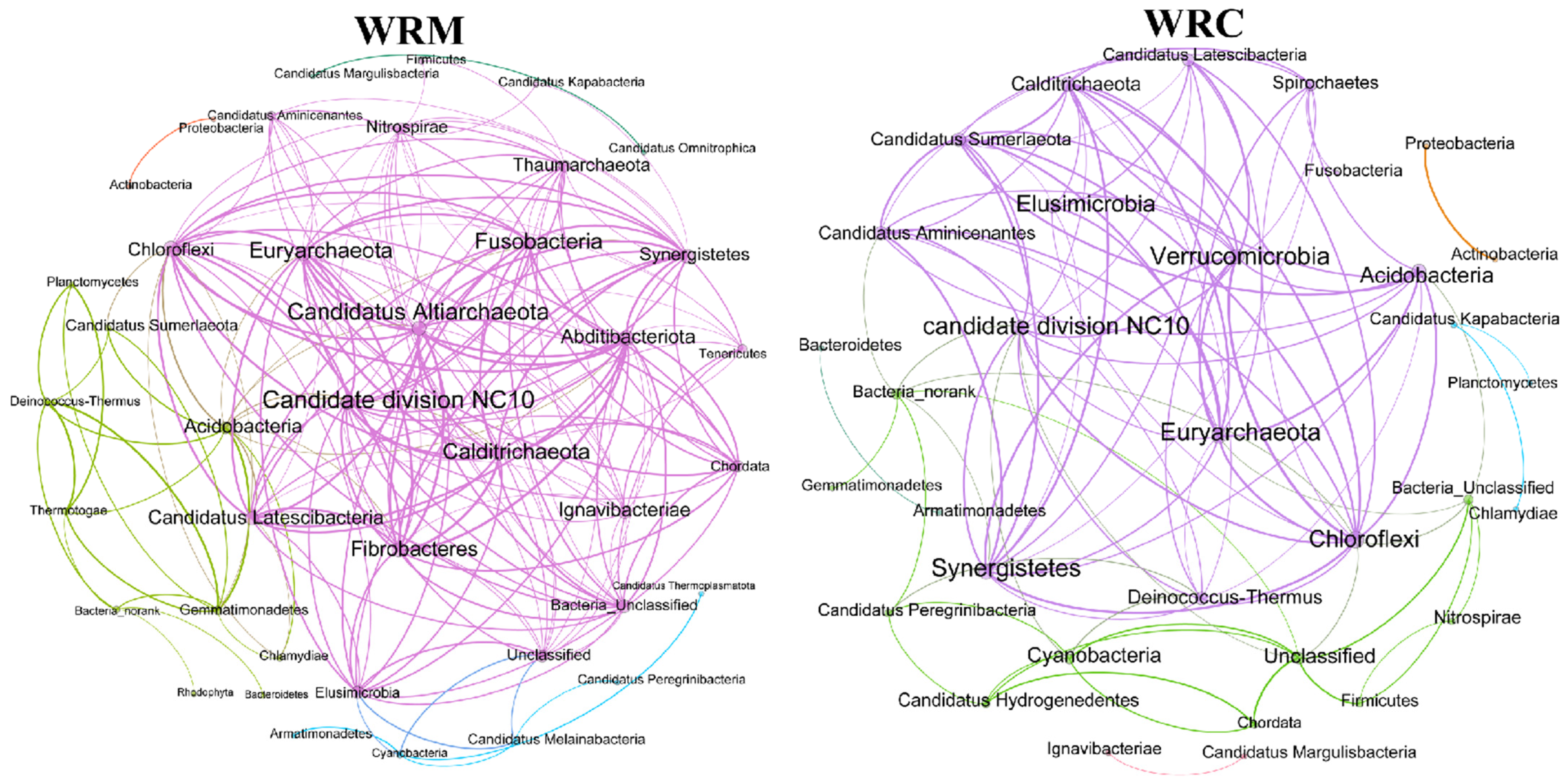
3.4. Assembly Processes and Co-Occurrence Patterns in the Water Bacterial Communities
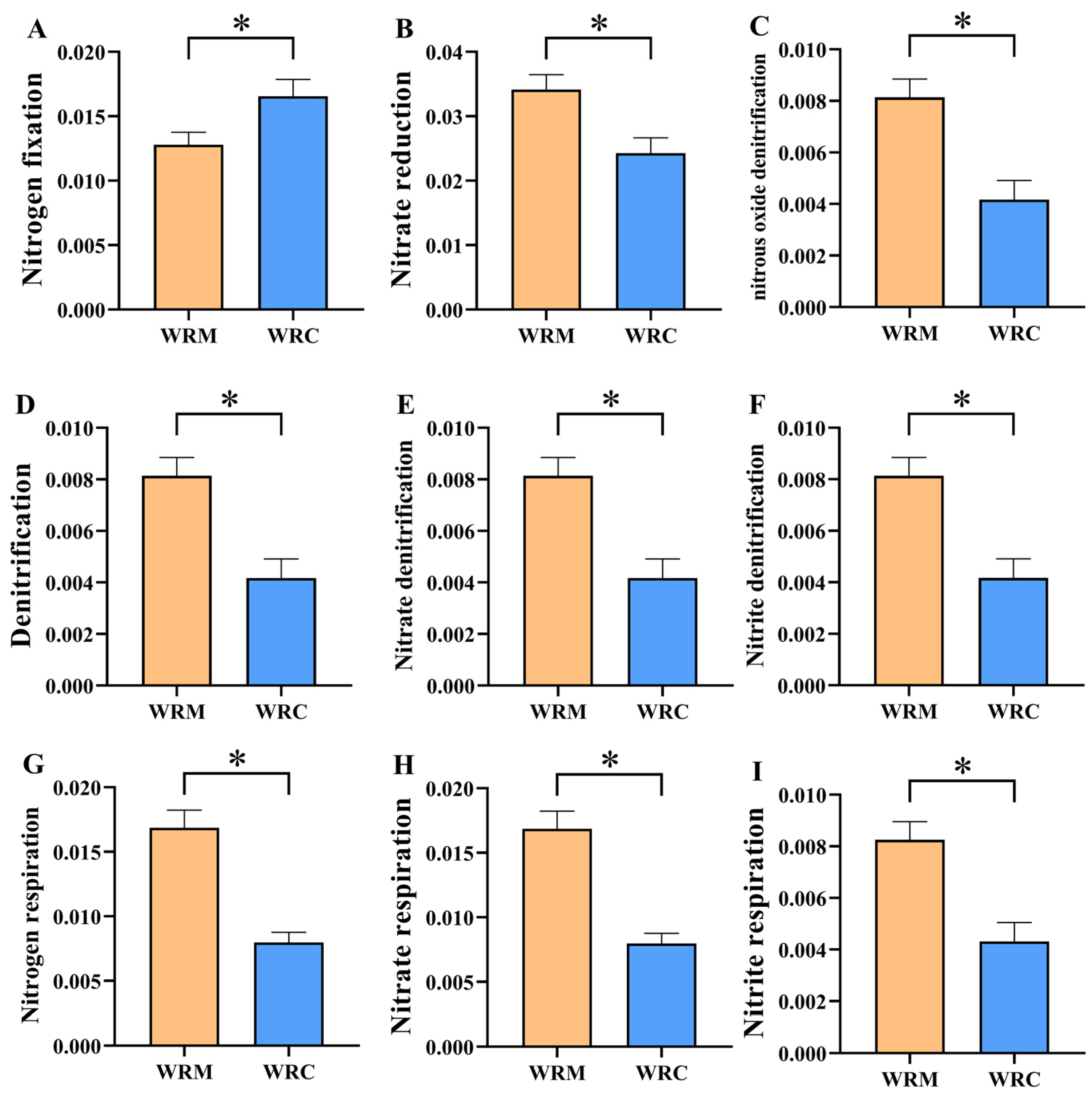
3.5. Function Prediction of Water Bacterial Communities
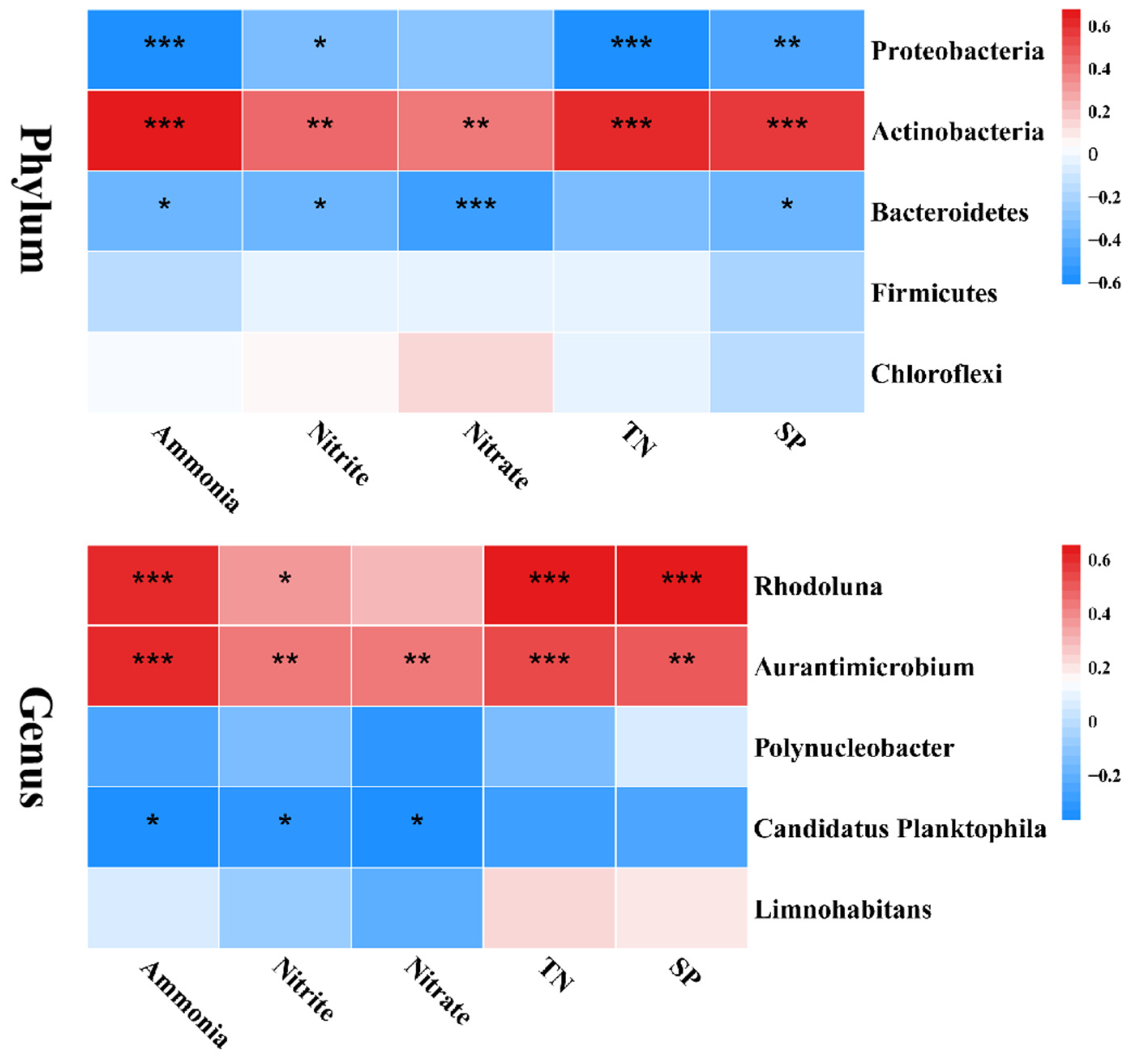
3.6. Relationship between Water Bacterial Community and Physicochemical Properties
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- You, G.; Xu, B.; Su, H.; Zhang, S.; Pan, J.; Hou, X.; Li, J.; Ding, R. Evaluation of Aquaculture Water Quality Based on Improved Fuzzy Comprehensive Evaluation Method. Water 2021, 13, 1019. [Google Scholar] [CrossRef]
- Peter, M.R.; Jacob, K.E. Pond water quality and its relation to fish yield and disease occurrence in small-scale aquaculture in arid areas. Heliyon 2023, 9, e16753. [Google Scholar]
- Yu, J.; Kang, H.; Kong, Q. Research Progress of Aquaculture Tailwater Treatment Technology. Tianjin Agric. Sci. 2023, 29, 83–91. [Google Scholar]
- Liu, C.; Hu, N.; Song, W.; Chen, Q.; Zhu, L. Aquaculture Feeds Can Be Outlaws for Eutrophication When Hidden in Rice Fields? A Case Study in Qianjiang, China. Int. J. Environ. Res. Public Health 2019, 16, 4471. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Li, H.; Guo, X.; Chen, B.; Wang, M. A semi-continuous efficient strategy for removing phosphorus and nitrogen from eel aquaculture wastewater using the self-flocculating microalga Desmodesmus sp. PW1. J. Environ. Manag. 2023, 346, 118970. [Google Scholar] [CrossRef] [PubMed]
- Yi, S. Contingent Valuation of Sustainable Integrated Agriculture-Aquaculture Products: The Case of Rice–Fish Farming Systems in South Korea. Agronomy 2019, 9, 601. [Google Scholar] [CrossRef]
- Yu, X.; Hao, X.; Dang, Z.; Yang, L. Report on the Development of China’s Integrated Rice and Fishery Breeding Industry (2023). Chin. J. Aquac. 2023, 8, 19–26. [Google Scholar]
- Dong, Y.; Jia, R.; Hou, Y.R.; Diao, W.X.; Li, B.; Zhu, J. Effects of stocking density on the growth performance, mitophagy, endocytosis and metabolism of Cherax quadricarinatus in integrated rice-crayfish farming systems. Front. Physiol. 2022, 13, 1040712. [Google Scholar] [CrossRef] [PubMed]
- Xiang, A. Survey and New Inquiry on Fish Breeding in Rice Fields. China Hist. Mater. Sci. Technol. 1995, 16, 62–74. [Google Scholar]
- Lu, S.; Liao, M.; Xie, C.; He, X.; Li, D.; He, L.; Chen, J. Seasonal dynamics of ammonia-oxidizing microorganisms in freshwater aquaculture ponds. Ann. Microbiol. 2014, 65, 651–657. [Google Scholar] [CrossRef]
- Lu, S.; Liu, X.; Liu, C.; Cheng, G.; Zhou, R.; Li, Y. A Review of Ammonia-Oxidizing Archaea and Anaerobic Ammonia-Oxidizing Bacteria in the Aquaculture Pond Environment in China. Front. Microbiol. 2021, 12, 775794. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.J.; Amenyogbe, E.; Li, R.X.; Zhang, J.D.; Xie, R.T.; Wang, Z.L.; Chen, G.; Huang, J.S. The Microflora Structure in the Digestive Tract, Culture Water, and Feed of Hybrid Grouper (Epinephelus fuscoguttatus ♀ × E. polyphekadion ♂) Cultured in an Outdoor Pond Based on a High-Throughput Sequencing Technique. Aquac. Res. 2023, 2023, 9923362. [Google Scholar] [CrossRef]
- Eissa, E.S.H.; Baghdady, E.S.; Gaafar, A.Y.; El-Badawi, A.A.; Bazina, W.K.; Abd Al-Kareem, O.M.; Abd El-Hamed, N.N.B. Assessing the Influence of Dietary Pediococcus acidilactici Probiotic Supplementation in the Feed of European Sea Bass (Dicentrarchus labrax L.) (Linnaeus, 1758) on Farm Water Quality, Growth, Feed Utilization, Survival Rate, Body Composition, Blood Biochemical Parameters, and Intestinal Histology. Aquac. Nutr. 2022, 2022, 5841220. [Google Scholar] [CrossRef]
- Huang, H.H.; Li, C.Y. Adaptability of commercial probiotics to biofloc system: Influences on autochthonal bacterial community, water quality and growth performance of shrimp (Litopenaeus vannamei). Aquaculture 2024, 590, 740992. [Google Scholar] [CrossRef]
- Wuquan, L.; Zexu, L.; Minze, L.; Yuan, X.; Jianing, Z.; Yue, W.; Danqing, H.; Chengbo, S. Effects of sodium humate and probiotics on growth performance enzyme activity and microbial environment of Litopenaeus vannamei in high-density zero-water exchange systems. Front. Mar. Sci. 2022, 9, 989325. [Google Scholar]
- Bentzon-Tilia, M.; Sonnenschein, E.C.; Gram, L. Monitoring and managing microbes in aquaculture—Towards a sustainable industry. Microb. Biotechnol. 2016, 9, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wan, W.; Liu, B.; Xu, W.; Gu, Z. Effects of rice-crayfish integrated system on microbial diversity and community structure in paddy water. J. Huazhong Agric. Univ. 2022, 41, 141–151. [Google Scholar] [CrossRef]
- Arunrat, N.; Sansupa, C.; Kongsurakan, P.; Sereenonchai, S.; Hatano, R. Soil Microbial Diversity and Community Composition in Rice-Fish Co-Culture and Rice Monoculture Farming System. Biology 2022, 11, 1242. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Wang, G.; Gong, W.; Yu, E.; Xia, Y.; Li, Z.; Tian, J.; Xie, J. Study on Organic Carbon, Nitrogen, and Phosphorus Budgets of Zero-Water Exchange Ponds of Grass Carp, Bighead Carp, and Crucian Carp. Prog. Fish. Sci. 2022, 43, 188–198. [Google Scholar] [CrossRef]
- Huang, F.; Tang, Q.; Liang, P.; Xiao, L. Improvement of the ammonium molybdate spectrophotometric method for phosphorus monitoring in fresh water. J. Lake Sci. 2016, 28, 1404–1410. [Google Scholar]
- Smart, M.M.; Rada, R.G.; Donnermeyer, G.N. Determination of total nitrogen in sediments and plants using persulfate digestion. Water Res. 1983, 17, 1207–1211. [Google Scholar] [CrossRef]
- Wu, H.Z.; Cao, A. Preparation and Adding Methods of Nessler’s Reagent Having Effects on Determination of Water Quality Ammonia Nitrogen. Adv. Mater. Res. 2013, 726–731, 1362–1366. [Google Scholar] [CrossRef]
- Lo, H.S.; Lo, K.W.; Yeung, C.F.; Wong, C.Y. Rapid visual and spectrophotometric nitrite detection by cyclometalated ruthenium complex. Anal. Chim. Acta 2017, 990, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Noyes, H.A. Accurate Determination of Soil Nitrates by Phenol Disulfonic Acid Method. J. Ind. Eng. Chem. 2002, 11, 213–218. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, X.; Hang, Y.; Liu, X.; Li, P. Combination Modes and Ecological Effects of Planting-Breeding ecosystem in Rice Field. Chin. Agric. Sci. Bull. 2019, 35, 46–51. [Google Scholar]
- Herlambang, A.; Murwantoko, M.; Istiqomah, I. Dynamic change in bacterial communities in the integrated rice–fish farming system in Sleman, Yogyakarta, Indonesia. Aquac. Res. 2021, 52, 5566–5578. [Google Scholar] [CrossRef]
- Tsuruta, T.; Yamaguchi, M.; Abe, S.-I.; Iguchi, K. Effect of fish in rice-fish culture on the rice yield. Fish. Sci. 2010, 77, 95–106. [Google Scholar] [CrossRef]
- Zhijuan, N.; Feifan, L.; Wenwu, Z.; Gangchun, X.; Bo, L.; Yuyu, W.; Nailiang, S.; Jiawen, H.; Pao, X. Microbial community structure of the rice-carp co-culture systems in Hani Terraces. J. Fish. China 2020, 44, 469–479. [Google Scholar] [CrossRef]
- Li, F.; Gao, J.; Xu, Y.; Nie, Z.; Fang, J.; Zhou, Q.; Xu, G.; Shao, N.; Xu, D.; Xu, P.; et al. Biodiversity and sustainability of the integrated rice-fish system in Hani terraces, Yunnan province, China. Aquac. Rep. 2021, 20, 100763. [Google Scholar] [CrossRef]
- Nayak, P.K.; Nayak, A.K.; Panda, B.B.; Lal, B.; Gautam, P.; Poonam, A.; Shahid, M.; Tripathi, R.; Kumar, U.; Mohapatra, S.D.; et al. Ecological mechanism and diversity in rice based integrated farming system. Ecol. Indic. 2018, 91, 359–375. [Google Scholar] [CrossRef]
- Peng, H.; Zhang, K.; Chen, C.; Xie, J. Effects of Carp (Cyprinus carpio) on Phytoplankton and Environmental Factors in Paddy Fields. J. Tianjin Agric. Univ. 2020, 27, 37–43. [Google Scholar] [CrossRef]
- Wang, S.; Zhao, Z.; Luo, L.; Zhang, R. Effects of Stocking Density on Growth Performance of Chinese Mitten Handed Crab (Erocheir sinensis) and Water Quality in a Rice-Crab Juveniles Co-culture System in Cold Regions. Chin. J. Fish. 2023, 36, 100–108. [Google Scholar]
- Zhao, X.-G.; Luo, H.; Liu, Q.-G.; Zhao, L.-J.; Cai, L.-R.; Dai, L.-L.; Zhang, Z. Influence of the cultured Odontobutis obscurus to the microbial community structure and diversity in rice-fish system. Freshw. Fish. 2017, 47, 8–14. [Google Scholar] [CrossRef]
- Jin, F. The Analysis of Microbial Community on Health Pelodiscus sinensis Breeding Aound Paddy and Penaeus vannamei Pond. Master’s Thesis, East China Normal University, Shanghai, China, 2013. [Google Scholar]
- Goodfellow, M. Ecology of Actinomycetes. Annu. Rev. Microbiol 1983, 37, 189–216. [Google Scholar] [CrossRef] [PubMed]
- Henning, S.M.; Yang, J.; Shao, P.; Lee, R.P.; Huang, J.; Ly, A.; Hsu, M.; Lu, Q.Y.; Thames, G.; Heber, D.; et al. Health benefit of vegetable/fruit juice-based diet: Role of microbiome. Sci. Rep. 2017, 7, 2167. [Google Scholar] [CrossRef]
- Rudi, K.; Berg, F.; Gaustad, E.; Tannes, T.; Vatn, M. Ratios between Alpha-, Beta- and Gamma-proteobacteria in tap water determined by the ProteoQuant assay. Lett. Appl. Microbiol. 2010, 50, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Abbas, A.; Duan, J.; Abdoulaye, A.H.; Fu, Y.; Lin, Y.; Xie, J.; Cheng, J.; Jiang, D. Deciphering Bacterial Community of the Fallow and Paddy Soil Focusing on Possible Biocontrol Agents. Agronomy 2022, 12, 431. [Google Scholar] [CrossRef]
- Zhou, Z.C.; Tran, P.Q.; Kieft, K.; Anantharaman, K. Genome diversification in globally distributed novel marine Proteobacteria is linked to environmental adaptation. ISME J. 2020, 14, 2060–2077. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Jia, R.; Sun, W.; Ding, H.; Li, B.; Zhu, J. Red Claw Crayfish Cherax quadricarinatus Cultivation Influences the Dynamics and Assembly of Benthic Bacterial Communities in Paddy Fields. Environments 2023, 10, 178. [Google Scholar] [CrossRef]
- Zhang, X.; Ma, X.; Wang, N.; Yao, T. New subgroup of Bacteroidetes and diverse microorganisms in Tibetan plateau glacial ice provide a biological record of environmental conditions. FEMS Microbiol. Ecol. 2009, 67, 21–29. [Google Scholar] [CrossRef]
- Wolinska, A.; Kuzniar, A.; Zielenkiewicz, U.; Izak, D.; Szafranek-Nakonieczna, A.; Banach, A.; Blaszczyk, M. Bacteroidetes as a sensitive biological indicator of agricultural soil usage revealed by a culture-independent approach. Appl. Soil Ecol. 2017, 119, 128–137. [Google Scholar] [CrossRef]
- Zhong, L.-Q.; Wang, M.-H.; Zhang, S.-Y.; Jiang, H.-C.; Chen, X.-H.; Zhu, G.-W.; Bian, W.-J. Microbial community structure in a channel catfish pond in Nanjing, China. J. Agro-Environ. Sci. 2020, 39, 1594–1604. [Google Scholar]
- Li, L.; Liang, K.; Li, B.; Xu, H. Comparison of Water Quality Indicatiors and Environmental Organisms in Pond and Rice-fish Culture of Odontobutis obscura. Acta Hydrobiol. Sin. 2024, 48, 625–633. [Google Scholar]
- Peng, H. A Comparative Study of Rice-Fish Ecosystem and Rice-Monoculture Ecosystem. Master’s Thesis, Tianjin Agricultural University, Tianjin, China, 2019. [Google Scholar]
- Burns, A.R.; Stephens, W.Z.; Stagaman, K.; Wong, S.; Rawls, J.F.; Guillemin, K.; Bohannan, B.J. Contribution of neutral processes to the assembly of gut microbial communities in the zebrafish over host development. ISME J. 2016, 10, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Yang, Q.; Li, X.; Chao, X.; Liu, H.; Wei, L.; Ba, S. Deterministic processes dominate the geographic distribution pattern and community assembly of phytoplankton in typical plateau rivers. Biodivers. Sci. 2023, 31, 43–57. [Google Scholar] [CrossRef]
- Huang, X. Effects of Oyster Shell-Loading on Water Variables and Bacterial Community in Shrimop (Litopenaeus vannamei) Culture System. Ph.D. Thesis, Ningbo University, Ningbo, China, 2023. [Google Scholar]
- Wang, R.; Chen, H.; Zhu, Y.; Al-Masqari, Z.A.; Yan, M.; Wang, G.; Dong, P.; Gao, F.; Lu, T.; Zhang, D.; et al. Survival status of Penaeus vannamei is associated with the homeostasis and assembly process of the intestinal bacterial community. Aquaculture 2022, 558, 738398. [Google Scholar] [CrossRef]
- Ruyue, W.; Sai, X.; Yuxiang, Z.; Tao, Z.; Shijian, G. Denitrifying anaerobic methane-oxidizing bacteria in river networks of the Taihu Basin: Community dynamics and assembly process. Front. Microbiol. 2022, 13, 1074316. [Google Scholar]
- Xie, M.; Zhang, J.; Jia, J.; Wang, G.; Qin, Z.; Gao, H.; Li, H. Advances in the Assembly of Soil Microbial Communities in Natural and Farmland Ecosystems. Chin. J. Soil Sci. 2023, 54, 1503–1512. [Google Scholar] [CrossRef]
- Rogueta, A.; Laiglea, G.S.; Theriala, C.; Bressya, A.; Soulignac, F.; Catherine, A.; Lacroix, G.; Jardillier, L.; Bonhomme, C.; Lerch, T.Z.; et al. Neutral community model explains the bacterial community assembly in freshwater lakes. FEMS Microbiol. Ecol. 2015, 91, fiv125. [Google Scholar] [CrossRef]
- Wang, P.; Xu, M.; Yang, W.; Bai, K.; Liu, W.; Zhang, Z.; Ji, B.; Fan, Y.; Zhang, X. Nitrogen addition stimulate random migration of plant community in a semiarid steppe. Glob. Ecol. Conserv. 2021, 26, e01508. [Google Scholar] [CrossRef]
- Wu, J.; Zhu, Z.; Waniek, J.J.; Niu, M.; Wang, Y.; Zhang, Z.; Zhou, M.; Zhang, R. The biogeography and co-occurrence network patterns of bacteria and microeukaryotes in the estuarine and coastal waters. Mar. Environ. Res. 2023, 184, 105873. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Bai, C.; Cai, J.; Dai, J.; Shao, K.; Tang, X.; Gao, G. Co-occurrence Network Reveals the Higher Fragmentation of the Bacterial Community in Kaidu River Than Its Tributaries in Northwestern China. Microbes Environ. 2018, 33, 127–134. [Google Scholar] [CrossRef]
- Ley, R.E.; Harris, J.K.; Wilcox, J.; Spear, J.R.; Miller, S.R.; Bebout, B.M.; Maresca, J.A.; Bryant, D.A.; Sogin, M.L.; Pace, N.R. Unexpected diversity and complexity of the Guerrero Negro hypersaline microbial mat. Appl. Environ. Microbiol. 2006, 72, 3685–3695. [Google Scholar] [CrossRef]
- Richter, D.; Matuła, J. Response of cyanobacteria and algae community from small water bodies to physicochemical parameters. Oceanol. Hydrobiol. Stud. 2012, 41, 18–28. [Google Scholar] [CrossRef]
- Yu, H.; Khan, A.U.; Subramanian, S.; Vesper, D.; Van Aken, B. Microbial Communities in Chesapeake & Ohio Canal National Historical Park and Their Function as Indicators of Water Quality. Geomicrobiol. J. 2019, 36, 673–682. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, G.-Z.; Yang, X.-N.; Wu, F.-P.; Zhao, W.; Zhang, H.-W.; Zhang, X. Microbial Community Structure and Diversity in Cellar Water by 16S rRNA High-throughput Sequencing. Huanjing Kexue 2017, 38, 1704–1716. [Google Scholar] [CrossRef]
- Li, Z.; Han, J.; Bai, H.; Peng, D.; Wang, L.; Bai, L. Effects of novel bioorganic fertilizer application on soil enzymes and bacterial community in multi-site rice paddies in China. AMB Express 2021, 11, 79. [Google Scholar] [CrossRef] [PubMed]
- Bai, Z.; Zheng, L.; Bai, Z.; Jia, A.; Wang, M. Long-term cultivation alter soil bacterial community in a forest-grassland transition zone. Front. Microbiol. 2022, 13, 1001781. [Google Scholar] [CrossRef]
- Zhang, Y.; Hou, Y.; Jia, R.; Li, B.; Zhu, J.; Ge, X. Nitrogen occurrence forms and bacterial community in sediment influenced by Bellamya purificata bioturbation. Front. Mar. Sci. 2022, 9, 1028716. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diao, W.; Yuan, J.; Jia, R.; Hou, Y.; Zhang, L.; Li, B.; Zhu, J. Integrated Rice–Fish Culture Alters the Bacterioplankton Community and Its Assembly Processes. Fishes 2024, 9, 254. https://doi.org/10.3390/fishes9070254
Diao W, Yuan J, Jia R, Hou Y, Zhang L, Li B, Zhu J. Integrated Rice–Fish Culture Alters the Bacterioplankton Community and Its Assembly Processes. Fishes. 2024; 9(7):254. https://doi.org/10.3390/fishes9070254
Chicago/Turabian StyleDiao, Weixu, Julin Yuan, Rui Jia, Yiran Hou, Liqiang Zhang, Bing Li, and Jian Zhu. 2024. "Integrated Rice–Fish Culture Alters the Bacterioplankton Community and Its Assembly Processes" Fishes 9, no. 7: 254. https://doi.org/10.3390/fishes9070254
APA StyleDiao, W., Yuan, J., Jia, R., Hou, Y., Zhang, L., Li, B., & Zhu, J. (2024). Integrated Rice–Fish Culture Alters the Bacterioplankton Community and Its Assembly Processes. Fishes, 9(7), 254. https://doi.org/10.3390/fishes9070254