



Hypoxia Activates HIF-1α and Affects Gene Expression and Transcriptional Regulation of PHD in Tegillarca granosa

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Hypoxia Stress Experiment
2.1.1. Experimental Samples Preparation
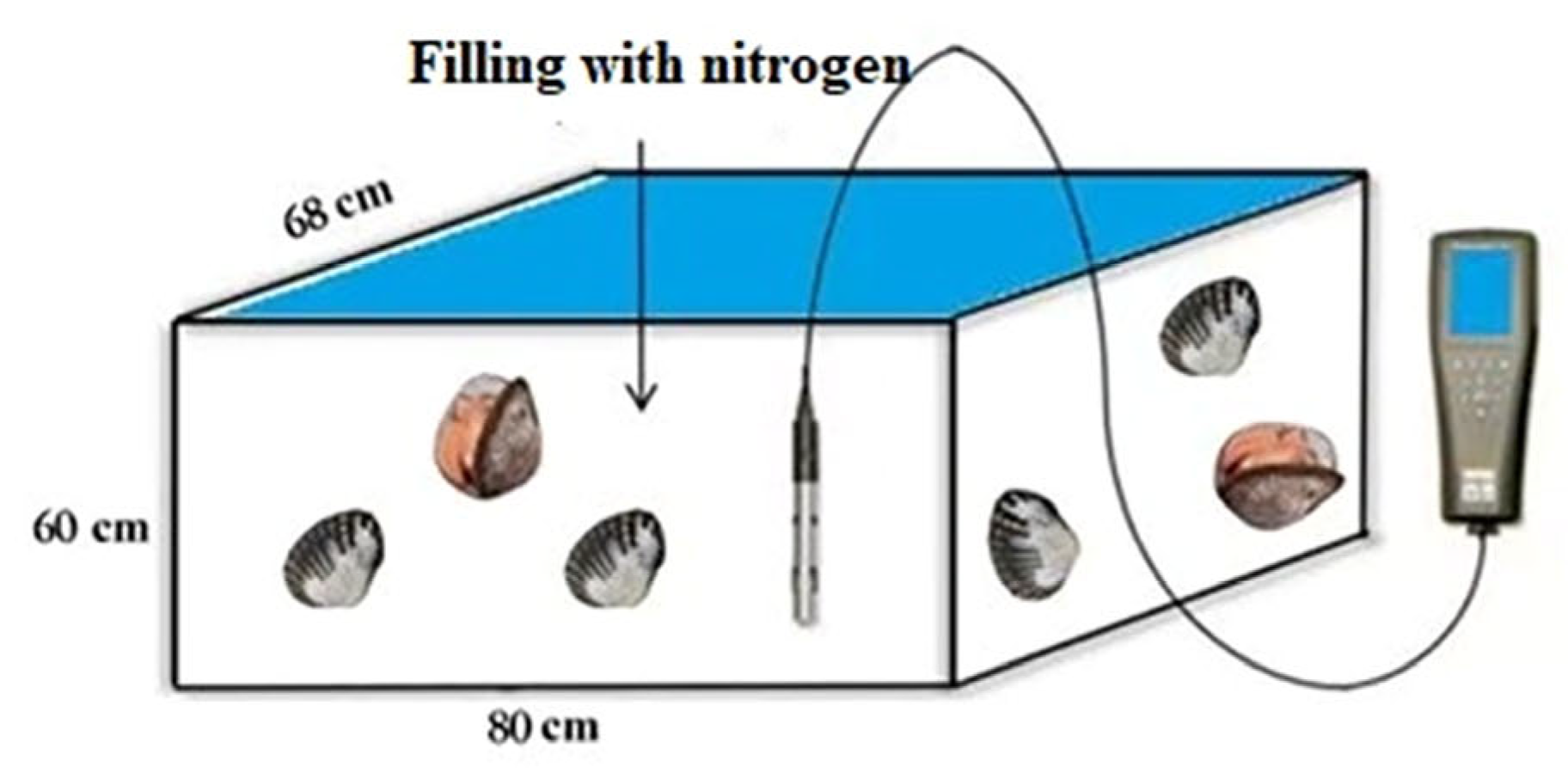
2.1.2. Hypoxia Experiment and Tissue Collection
2.2. PCR Amplification of Tg-HIF-1α ORF and Sequence Analysis of Tg-HIF-1α
2.3. Quantitative Real-Time PCR
2.4. Amplification of Tg-PHD and Tg-Hb Promoter and Construction of Recombinant Plasmid
2.5. HEK293T Cell Culture, Transient Transfection, and Transcriptional Regulation of Tg-PHD and Tg- Hb by Tg-HIF-1α under Hypoxia
2.6. Statistical Analysis
3. Results
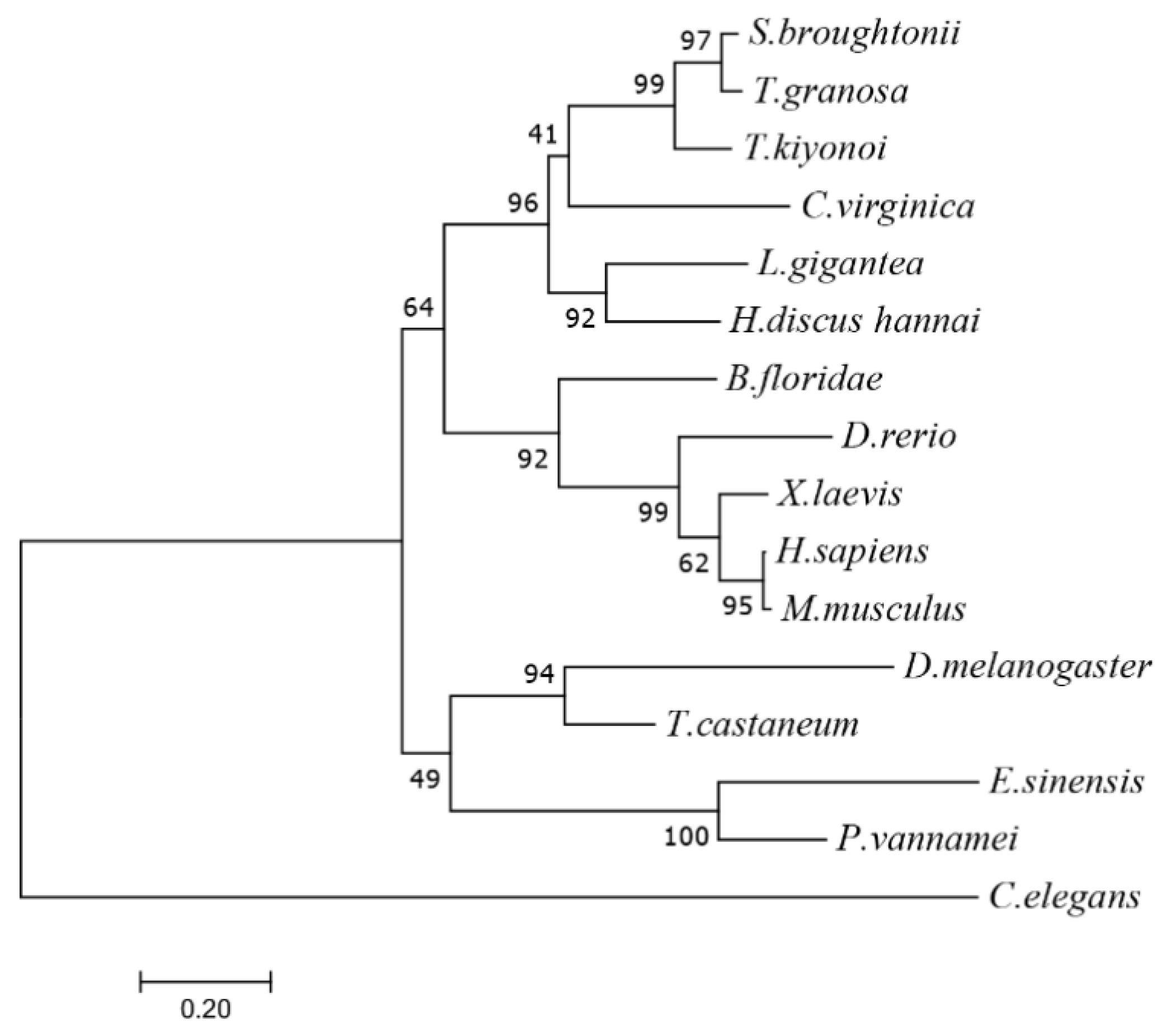
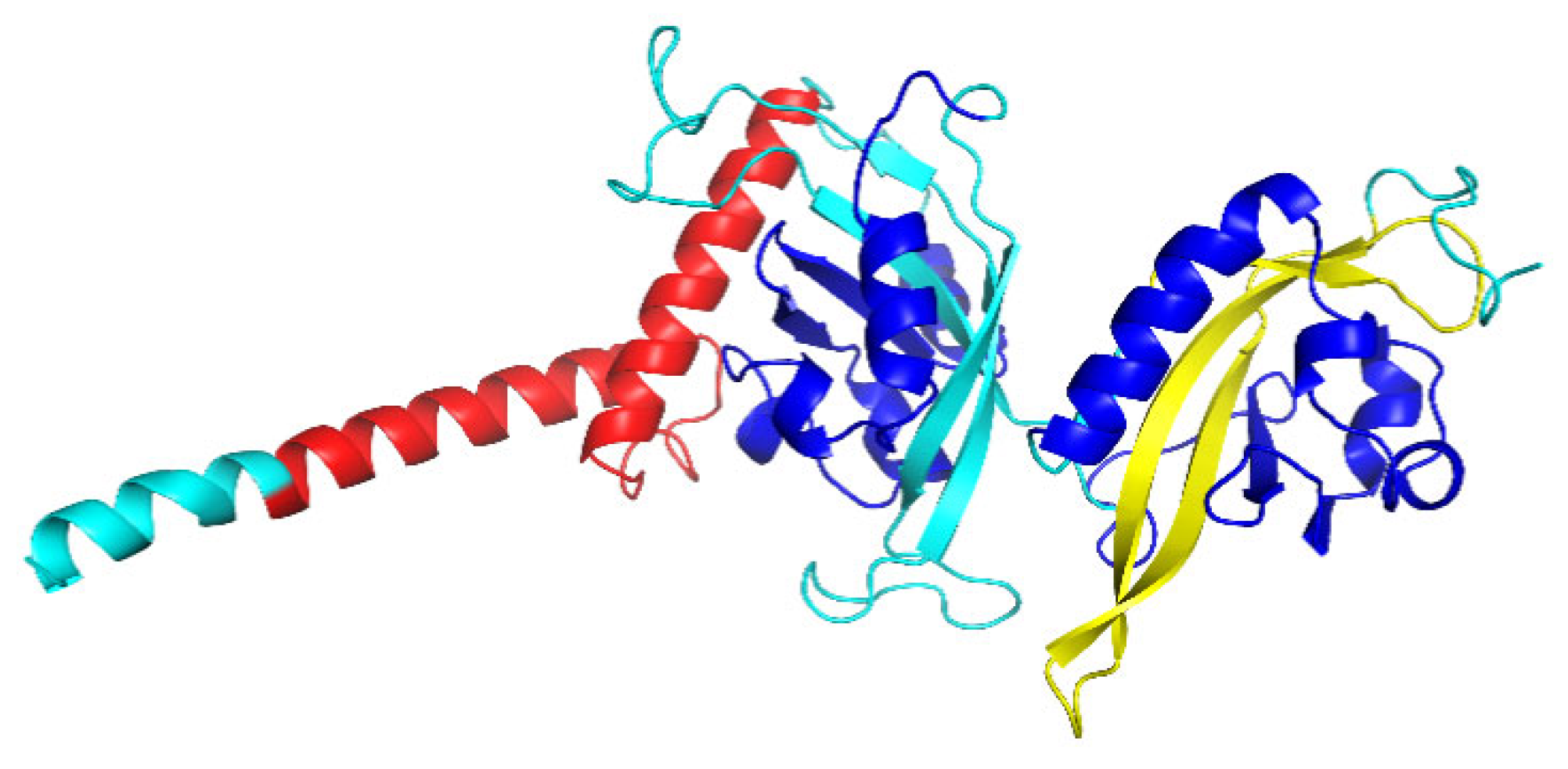
3.1. The Characterization of Tg-HIF-1α
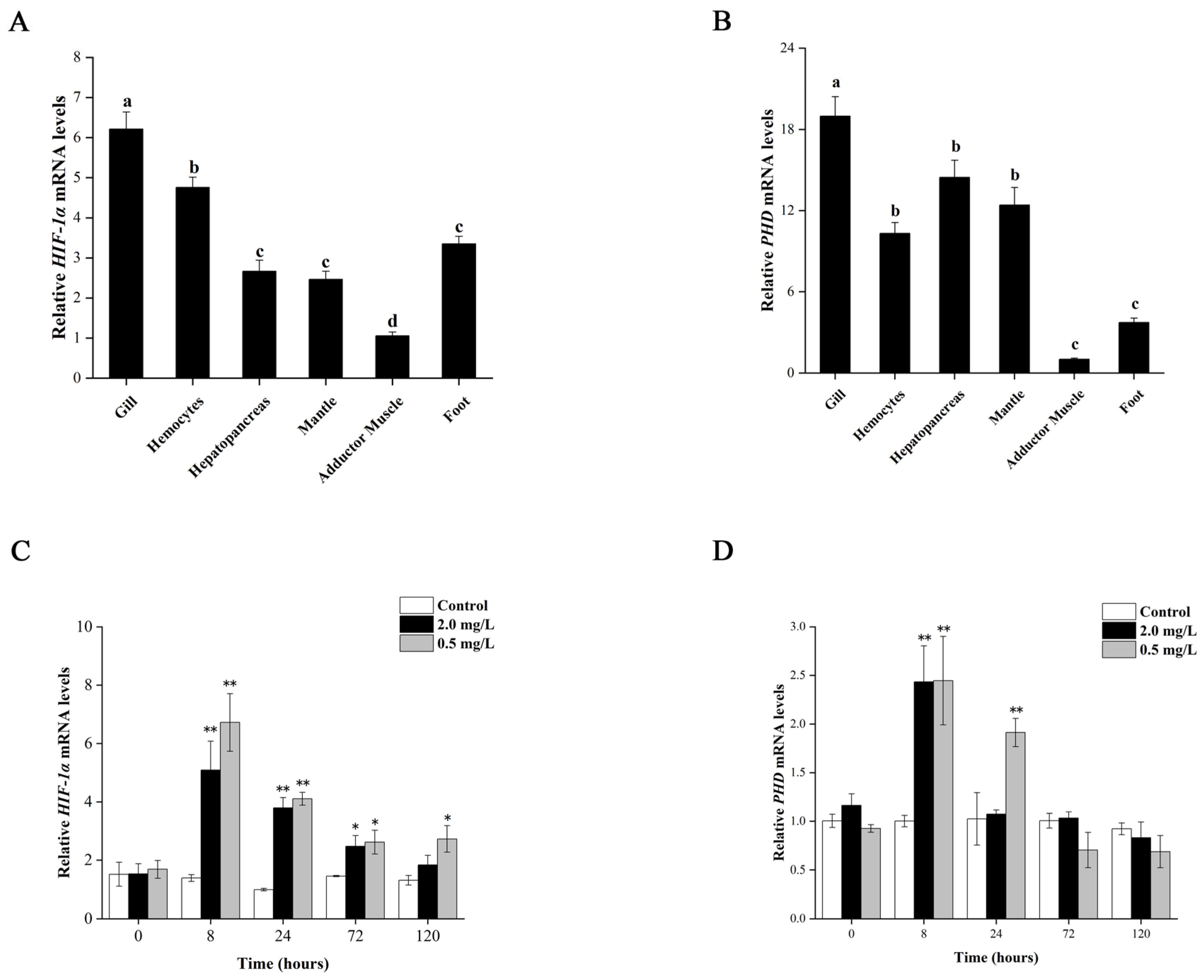
3.2. Differential Expression of Tg-HIF-1α and Tg-PHD in Different Tissues
3.3. Expression of Tg-HIF-1α and Tg-PHD under Different DO Concentrations
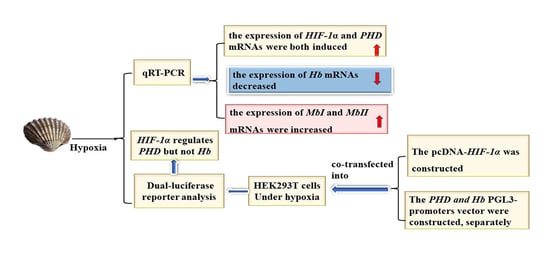
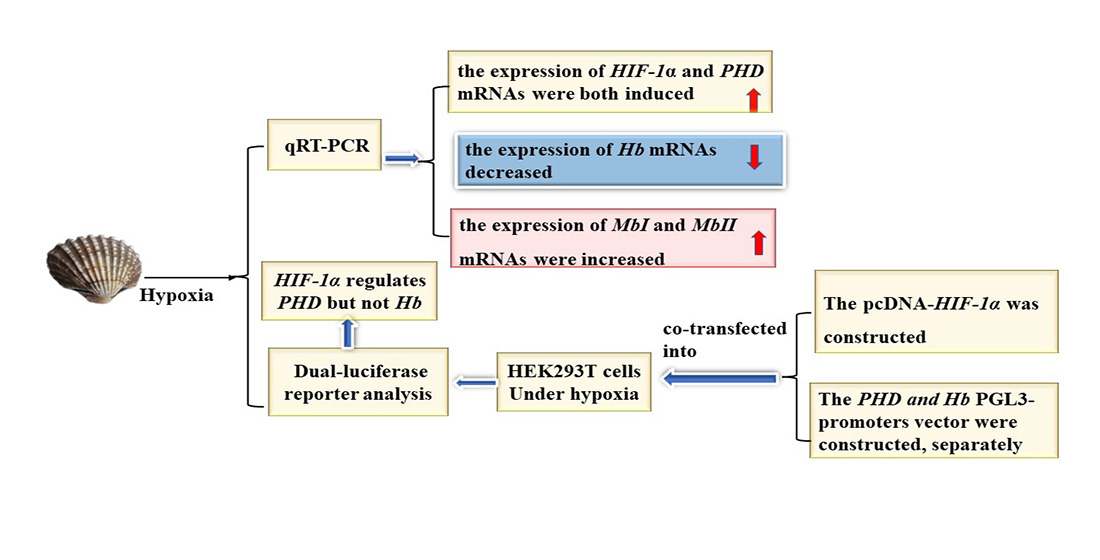
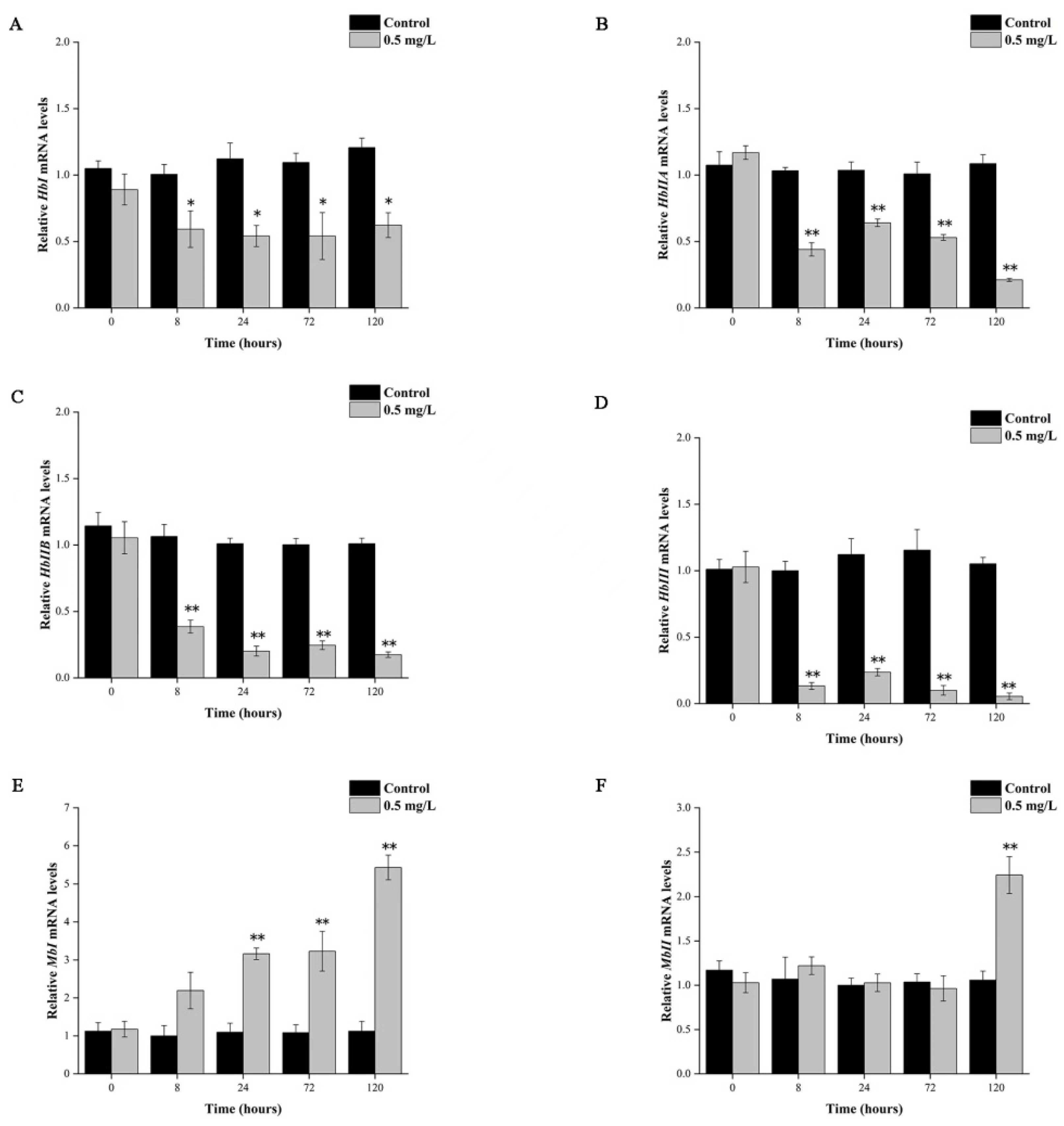
3.4. Hypoxia Affected mRNA Expression of Tg-Hb and Tg-Mb
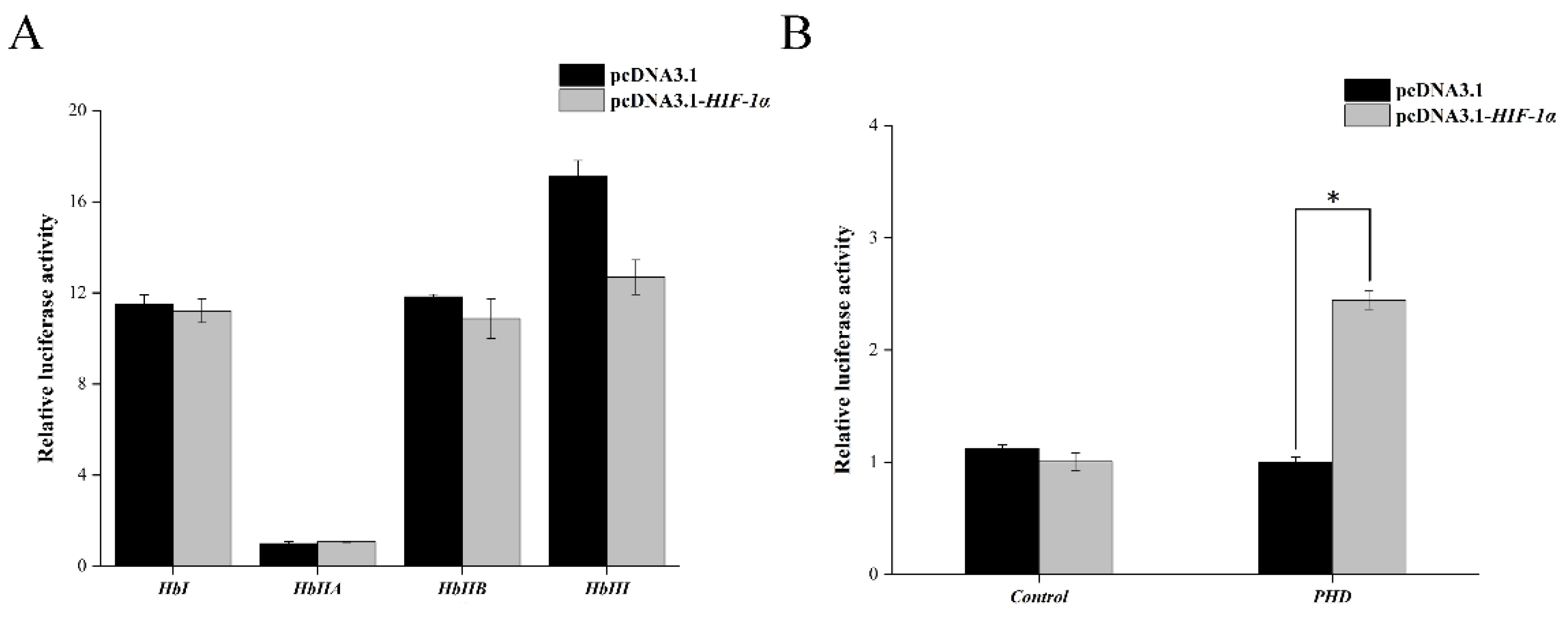
3.5. Transcription Activity Assay of Tg-HIF-1α on Tg-PHD and Tg-Hb
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Appendix A





References
- Chen, C.C.; Gong, G.C.; Shiah, F.K. Hypoxia in the East China Sea: One of the largest coastal low-oxygen areas in the world. Mar. Environ. Res. 2007, 64, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Brouwer, M. Hypoxia-inducible factor, gsHIF, of the grass shrimp Palaemonetes Pugio: Molecular characterization and response to hypoxia. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 147, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Velislava, T.; Vadeboncoeur, C.; Ting, J.; Perr, S.F. Effects of hypoxia-induced gill remodeling on the innervation and distribution of ionocytes in the gill of goldfish, Carassius auratus. J. Comp. Neurol. 2014, 522, 118–130. [Google Scholar]
- Ledford, H.; Callaway, E. Biologists who decoded how cells sense oxygen win medicine Nobel. Nature 2019, 574, 161–162. [Google Scholar] [CrossRef] [PubMed]
- Kewley, R.J.; Whitelaw, M.L.; Chapman, S.A. The mammalian basic helix-loop-helix/PAS’ family of transcriptional regulators. Int. J. Biochem. Cell Biol. 2004, 36, 189–204. [Google Scholar] [CrossRef] [PubMed]
- Kaelin, W.G.; Ratcliffe, P.J. Oxygen sensing by metazoans: The central role of the HIF hydroxylase pathway. Mol. Cell. 2008, 30, 393–402. [Google Scholar] [CrossRef]
- Epstein, A.C.; Gleadle, J.M.; McNeill, L.A.; Hewitson, K.S. C. elegans EGL-9 and mammalian homologs define a family of dioxygenases that regulate HIF by prolyl hydroxylation. Cell 2001, 107, 43–54. [Google Scholar] [CrossRef]
- Appelhoff, R.J.; Tian, Y.M.; Raval, R.R. Differential function of the prolyl hydroxylases PHD1, PHD2, and PHD3 in the regulation of hypoxia-inducible factor. J. Biol. Chem. 2004, 279, 38458–38465. [Google Scholar] [CrossRef]
- Yu, F.; White, S.B.; Zhao, Q.; Lee, F.S. HIF-1alpha binding to VHL is regulated by stimulus-sensitive proline hydroxylation. Proc. Natl. Acad. Sci. USA. 2001, 98, 9630–9635. [Google Scholar] [CrossRef]
- Hon, W.C.; Wilson, M.I.; Harlos, K.; Claridge, T.W.; Schofield, C.J.; Pugh, C.W.; Maxwell, P.H.; Ratcliffe, P.J.; Stuart, D.I.; Jones, E.Y. Structural basis for the recognition of hydroxyproline in HIF-1 alpha by pVHL. Nature 2002, 417, 975–978. [Google Scholar] [CrossRef]
- Semenza, G.L. The genomics and genetics of oxygen homeostasis. Annu. Rev. Genom. Hum. Genet. 2020, 21, 183–204. [Google Scholar] [CrossRef] [PubMed]
- Pugh, C.W.; O’Rourke, J.F.; Nagao, M.; Gleadle, J.M.; Ratcliffe, P.J. Activation of hypoxia-inducible factor-1; Definition of regulatory domains within the alpha subunit. J. Bio. Chem. 1997, 272, 11205–11214. [Google Scholar] [CrossRef]
- Yu, A.; Frid, M.G.; Shimoda, L.A.; Wiener, C.M.; Stenmark, K.; Semenza, G.L. Temporal, spatial, and oxygen-regulated expression of hypoxia-inducible Factor-1 in the lung. Am. J. Physiol. 1998, 275, L818–L826. [Google Scholar] [CrossRef] [PubMed]
- Kallio, P.J.; Okamoto, K.; O’Brien, S.; Carrero, P.; Makino, Y.; Tanaka, H.; Poellinger, L. Signal transduction in hypoxic cells: Inducible nuclear translocation and recruitment of the CBP/p300 coactivator by the hypoxia-inducible factor-1 alpha. EMBO J. 1998, 17, 6573–6586. [Google Scholar] [CrossRef]
- Wenger, R.H.; Camenisch, G.; Stiehl, D.P.; Katschinski, D.M. HIF prolyl-4-hydroxylase interacting proteins: Consequences for drug targeting. Curr. Pharm. Des. 2009, 15, 3886–3894. [Google Scholar] [CrossRef]
- Rytkönen, K.T.; Williams, T.A.; Renshaw, G.M.; Primmer, C.R.; Nikinmaa, M. Molecular evolution of the metazoan PHD-HIF oxygen-sensing system. Mol. Biol. Evol. 2011, 28, 1913–1926. [Google Scholar] [CrossRef] [PubMed]
- Roesner, A.; Mitz, S.A.; Hankeln, T.; Burmester, T. Globins and hypoxia adaptation in the goldfish, Carassius auratus. FEBS J. 2008, 275, 3633–3643. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Xiao, S.S.; Zhang, R.; Liu, L.L.; Zhu, H. Physiological changes and transcriptional modulation of HIF-αs in Siberian sturgeon in response to hypoxia. Aquaculture 2021, 545, 737219. [Google Scholar] [CrossRef]
- Xiao, W.H. The hypoxia signaling pathway and hypoxic adaptation in fishes. Sci. China Life Sci. 2015, 58, 148–155. [Google Scholar] [CrossRef]
- Dunwoodie, S.L. The role of hypoxia in development of the mammalian embryo. Dev. Cell. 2009, 17, 755–773. [Google Scholar] [CrossRef]
- Gary, J.S.; Wu, R.S.; Ying, Y. Effects of hypoxia and organic enrichment on the coastal marine environment. Mar. Ecol. Prog. Ser. 2002, 238, 249–279. [Google Scholar] [CrossRef]
- Wang, T.; Meng, J.; Li, L.; Zhang, G.F. Characterization of CgHIFα-Like, a novel bHLH-PAS transcription factor family member, and its role under hypoxia stress in the Pacific oyster Crassostrea gigas. PLoS ONE. 2016, 11, e0166057. [Google Scholar] [CrossRef]
- Shen, G.Y.; Huang, L.F.; Guo, F. Marine Ecology, 3rd ed.; Science Press China: Beijing, China, 2010; pp. 100–125. [Google Scholar]
- Lappin, T.R.; Lee, F.S. Update on mutations in the HIF: EPO pathway and their role in erythrocytosis. Blood Rev. 2019, 37, 100590. [Google Scholar] [CrossRef] [PubMed]
- Gorr, T.A.; Cahn, J.D.; Yamagata, H.F. Hypoxia-induced synthesis of hemoglobin in the crustacean Daphnia magna is hypoxia-inducible factor-dependent. J. Biol. Chem. 2004, 279, 36038–36047. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.B.; Wang, J.; Li, C.; Li, P.; Wang, S.; Lin, Z. A preliminary study on the antibacterial mechanism of Tegillarca granosa hemoglobin by derived peptides and peroxidase activity. Fish Shellfish Immunol. 2016, 51, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Zha, S.J.; Peng, Z.L.; Lin, Z.H.; Bao, Y.B. Hypoxia-mediated immunotoxicity in the blood clam Tegillarca granosa. Mar. Environ. Res. 2022, 177, 105632. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.B.; Wang, Q.; Lin, Z.H. Hemoglobin of the blood clam Tegillarca granosa (Tg-HbI) is involved in the immune response against bacterial infection. Fish Shellfish Immunol. 2011, 31, 517–523. [Google Scholar] [CrossRef]
- Bao, Y.B.; Zeng, Q.F.; Wang, J.; Zhang, Z.L.; Zhang, Y.; Wang, S.F.; Wong, N.K.; Yuan, W.B.; Huang, Y.Y.; Zhang, W.F.; et al. Genomic insights into the origin and evolution of molluscan red-bloodedness in the blood clam Tegillarca granosa. Mol. Biol. Evol. 2021, 38, 2351–2365. [Google Scholar] [CrossRef]
- Riggs, A.F.; Gorr, T.A. A globin in every cell. Proc. Natl. Acad. Sci USA. 2006, 103, 2469–2470. [Google Scholar] [CrossRef]
- Zha, S.J.; Rong, J.H.; Guan, X.F.; Tang, Y.; Han, Y.; Liu, G.X. Immunotoxicity of four nanoparticles to a marine bivalve species, Tegillarca granosa. J. Hazard. Mater. 2019, 377, 237–248. [Google Scholar] [CrossRef]
- Zhang, W. Influence of hypoxia stress on physiological metabolism of Ruditapes philippinarum. Chin. J. Ecol. 2014, 33, 2448–2453. [Google Scholar]
- Mu, Y.N.; Li, W.R.; He, L.H.; Chen, J.; Chen, X.H. Transcriptome analysis reveals new insights into immune response to hypoxia challenge of large yellow croaker (larimichthys crocea). Fish. Shellfish Immun. 2020, 98, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.W.; Wu, B.; Liu, Z.L.; Zhou, L.Q.; Sun, X.J.; Zhao, Q.; Yang, A.G. Structural characteristics of HIF-1α from Scapharca broughtonii and expression analysis under hypoxia. J. Fish. Sci. China. 2019, 26, 646–656. [Google Scholar]
- Wu, L. Effects of hypoxic preconditioning on the physiological and biochemical characteristics of Scapharca broughtonii under hypoxia stress. Prog. Fish. Sci. 2022, 43, 1–10. [Google Scholar]
- Piontkivska, H.; Chung, J.S.; Ivanina, A.V.; Sokolov, E.P.; Techa, S.; Sokolova, I.M. Molecular characterization and mRNA expression of two key enzymes of hypoxia-sensing pathways in eastern oysters Crassostrea virginica (Gmelin): Hypoxia-inducible factor α (HIF-α) and HIF-prolyl hydroxylase (PHD). Comp. Biochem. Physiol. Part D Genom. Proteom. 2011, 6, 103–114. [Google Scholar] [CrossRef]
- Kawabe, S.; Yokoyama, Y. Role of hypoxia-inducible factor alpha in response to hypoxia and heat shock in the Pacific oyster Crassostrea gigas. Mar. Biotechnol. 2012, 14, 106–119. [Google Scholar] [CrossRef]
- Soñanez, O.J.G.; Peregrino, A.B.; Gómez, J.S.; López, Z.A.; Forman, H.J.; Plascencia, Y.G. Molecular characterization of hypoxia inducible factor-1 (HIF-1) from the white shrimp Litopenaeus vannamei and tissue-specific expression under hypoxia. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2009, 150, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Hardy, K.M.; Follett, C.R.; Burnett, L.E.; Lema, S.C. Gene transcripts encoding hypoxia-inducible factor (HIF) exhibit tissue and muscle fiber type-dependent responses to hypoxia and hypercapnic hypoxia in the Atlantic blue crab, Callinectes sapidus. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2012, 163, 137–146. [Google Scholar] [CrossRef]
- Soitamo, A.J.; Rabergh, C.M.; Gassmann, M.; Sistonen, L.; Nikinmaa, M. Characterization of a hypoxia-inducible factor (HIF-1a) from rainbow trout accumulation of protein occurs at normal venous oxygen tension. J. Biol. Chem. 2001, 276, 19699–19705. [Google Scholar] [CrossRef]
- Cai, X.H.; Huang, Y.T.; Zhang, X.; Wang, S.; Zou, Z.H.; Wang, G.D.; Wang, Y.L.; Zhang, Z.P. Cloning, characterization, hypoxia, and heat shock response of hypoxia inducible factor-1 (HIF-1) from the small abalone Haliotis diversicolor. Gene 2014, 534, 256–264. [Google Scholar] [CrossRef]
- Wang, T. The Molecular Mechanism of Hypoxia Signal Pathway in the Pacific Oyster (Crassostrea gigas). Ph.D. Thesis, University of Chinese Academy of Science, Beijing, China, 2017; pp. 1–135, (In Chinese with an English abstract). [Google Scholar]
- Berra, E.; Ginouvés, A.; Pouysségur, J. The hypoxia-inducible factor hydroxylases bring fresh air into hypoxia signaling. EMBO Rep. 2006, 7, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Rosner, A.; Hankeln, T.; Burmester, T. Hypoxia induces a complex response of globin expression in zebrafish (Danio rerio). Exp. Biol. 2006, 209, 2129–2137. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.S.; Qi, Z.T.; Tian, J.Y.; Qiu, M.; Zhao, W.H.; Wang, A.M.; Huang, J.T.; Guo, X.J. Cloning of hemoglobin-α1 from half-smooth tongue sole (Cynoglossus semilaevis) and its expression under short-term hypoxia. Zool. Res. 2011, 32, 641–646. [Google Scholar]
- Spriggs, K.A.; Bushell, M.; Willis, A.E. Translational regulation of gene expression during conditions of cell stress. Mol. Cell 2010, 40, 228–237. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′-3′) | Application | Length (bp) |
|---|---|---|---|
| qPCR-Tg-HIF-1α-F | ATTCCAGCCCAGACCTT | qRT-PCR | 106 |
| qPCR-Tg-HIF-1α-R | GGGAGCCAAATGAGTAAGA | ||
| qPCR-Tg-PHD-F | GACGTGCCAGAAGGAAT | qRT-PCR | 134 |
| qPCR-Tg-PHD-R | CGATAATAAGAGGCTTTACACT | ||
| qPCR-Tg-HbI-F | TACTTCAAGCGTTTGGGAGA | qRT-PCR | 116 |
| qPCR-Tg-HbI-R | GCTGGGCTGTCGAGATGA | ||
| qPCR-Tg-HbIIA-F | GACATACGCACTCAACAACTT | qRT-PCR | 114 |
| qPCR-Tg-HbIIA-R | AATGCTTCGGCACTGAT | ||
| qPCR-Tg-HbIIB-F | GGACACGCCATCACCTT | qRT-PCR | 172 |
| qPCR-Tg-HbIIB-R | TTGCCTTCAATACTGCTCTAA | ||
| qPCR-Tg-HbIII-F | AAATGGCGTAGCTCTAATG | qRT-PCR | 195 |
| qPCR-Tg-HbIII-R | TTTCGTGCAACATCTTCTA | ||
| qPCR-Tg-MbI-F | AGCATGGCGTTGAACTA | qRT-PCR | 141 |
| qPCR-Tg-MbI-R | CATACATAAATGTGGTGGC | ||
| qPCR-Tg-MbII-F | GTTGGGCATATCTTGAATC | qRT-PCR | 150 |
| qPCR-Tg-MbII-R | GACCACGCATACTACGG | ||
| qPCR-Tg-18S rRNA-F | CTTTCAAATGTCTGCCCTATCAACT | qRT-PCR | 148 |
| qPCR-Tg-18S rRNA-R | TCCCGTATTGTTATTTTTCGTCACT | qRT-PCR | |
| PGL3-Tg-HbI-F | CGAGCTCTTACCCTTCCCGTCTTC | HbI | 1745 |
| PGL3-Tg-HbI-R | CCCTCGAGTTAGCCCAACCTCACAA | ||
| PGL3-Tg-HbIIA-F | CGAGCTCAGGTCAGATGTACCAGCAC | HbIIA | 2071 |
| PGL3-Tg-HbIIA-R | CCCTCGAGTTCCGCAACAGAAAGAG | ||
| PGL3-Tg-HbIIB-F | CGAGCTCATTTGTGGCAGCGTTCA | HbIIB | 1593 |
| PGL3-Tg-HbIIB-R | CCCTCGAGAAAGGGCGTCTGTAGCA | ||
| PGL3-Tg-HbIII-F | CGAGCTCCTTTAAGGCTTGTCACTC | HbIII | 1655 |
| PGL3-Tg-HbIII-R | CCCTCGAGCAACCTCTGGGTCTGTG | ||
| PGL3-Tg-PHD-F | CGAGCTCTTGCCTTTGTCTTTCTC | PHD | 1844 |
| PGL3-Tg-PHD-R | CCCTCGAGTTCTTCAGGACCTATGC | ||
| pcDNA3.1-Tg-HIF-1α-F | CGGGATCCAAATGGCCGGTAAAAGAAG | HIF-1α | 2142 |
| pcDNA3.1-Tg-HIF-1α-R | CCCTCGAGCTTTTTTCCTCAATAATGCTG |
| HLH | PAS-A | PAS-B | PAC | ORF | |
|---|---|---|---|---|---|
| S. broughtoniiHIF-1α | 98.15 | 88.06 | 94.03 | 95.45 | 87.96 |
| T. kiyonoi HIF-1α | 87.04 | 79.10 | 86.57 | 84.09 | 69.49 |
| C. virginica HIF-1α | 60.71 | 58.21 | 62.96 | 77.27 | 47.55 |
| H. discusi HIF-1α | 58.93 | 53.73 | 73.13 | 75.00 | 43.69 |
| D. rerio HIF-1α | 62.96 | 47.76 | 55.22 | 59.09 | 29.77 |
| M. musculus HIF-1α | 55.36 | 49.25 | 62.69 | 61.36 | 28.43 |
| H. sapiensHIF-1α | 55.36 | 49.25 | 62.69 | 61.36 | 28.40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, Z.; Liu, X.; Jin, M.; Zhan, Y.; Zhang, X.; Bao, Y.; Liu, M. Hypoxia Activates HIF-1α and Affects Gene Expression and Transcriptional Regulation of PHD in Tegillarca granosa. Fishes 2023, 8, 359. https://doi.org/10.3390/fishes8070359
Peng Z, Liu X, Jin M, Zhan Y, Zhang X, Bao Y, Liu M. Hypoxia Activates HIF-1α and Affects Gene Expression and Transcriptional Regulation of PHD in Tegillarca granosa. Fishes. 2023; 8(7):359. https://doi.org/10.3390/fishes8070359
Chicago/Turabian StylePeng, Zhilan, Xiaomin Liu, Ming Jin, Yu Zhan, Xiaolin Zhang, Yongbo Bao, and Minhai Liu. 2023. "Hypoxia Activates HIF-1α and Affects Gene Expression and Transcriptional Regulation of PHD in Tegillarca granosa" Fishes 8, no. 7: 359. https://doi.org/10.3390/fishes8070359
APA StylePeng, Z., Liu, X., Jin, M., Zhan, Y., Zhang, X., Bao, Y., & Liu, M. (2023). Hypoxia Activates HIF-1α and Affects Gene Expression and Transcriptional Regulation of PHD in Tegillarca granosa. Fishes, 8(7), 359. https://doi.org/10.3390/fishes8070359