
Effects of Recombinant Leptin Proteins on the Expression of Key Genes in the HPG Axis and Liver of Tongue Sole In Vitro


 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Quantitative Reverse Transcription Polymerase Chain Reaction (RT-qPCR)
2.3. Statistic Analysis
3. Results
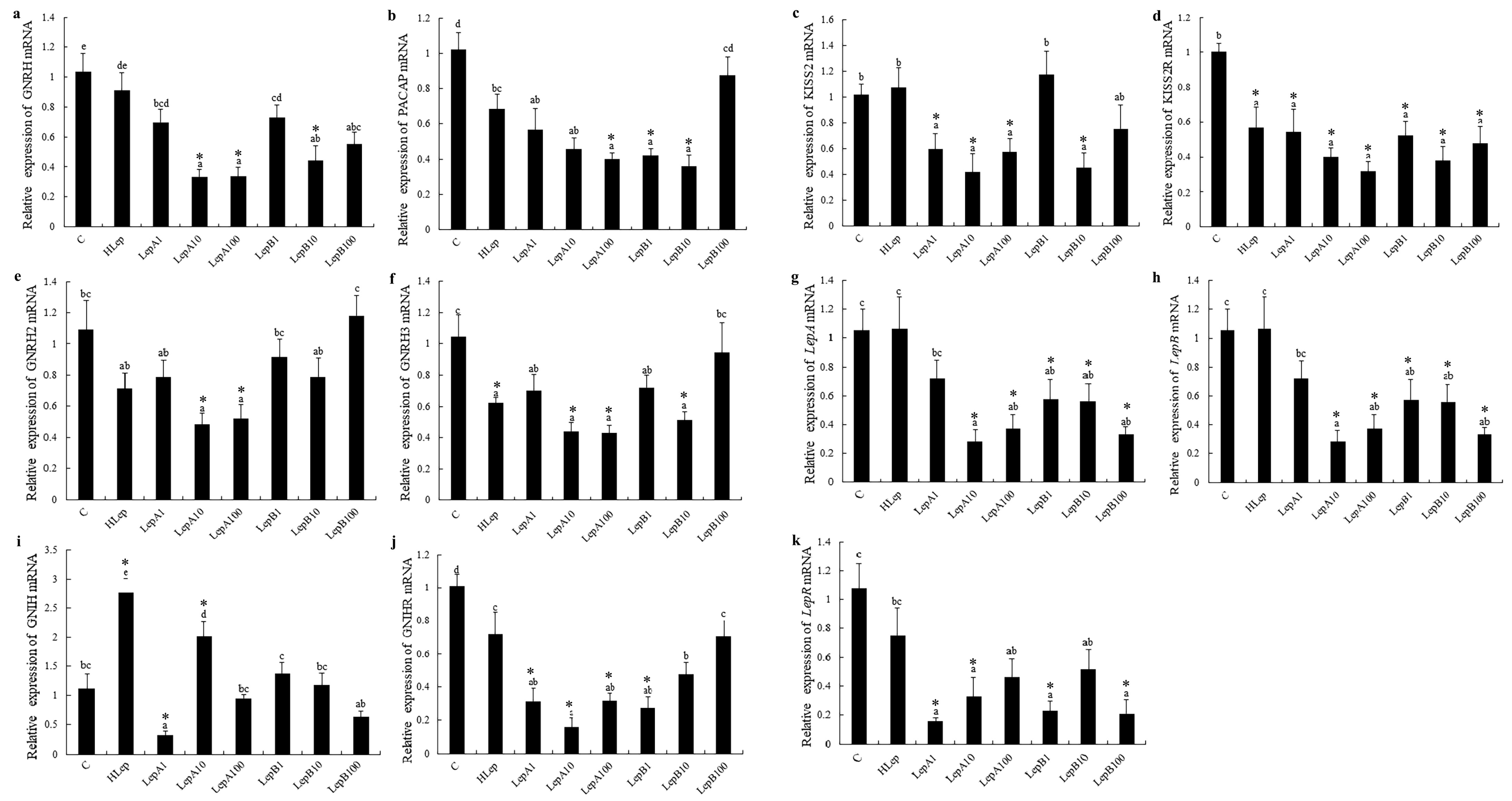
3.1. Hypothalamus
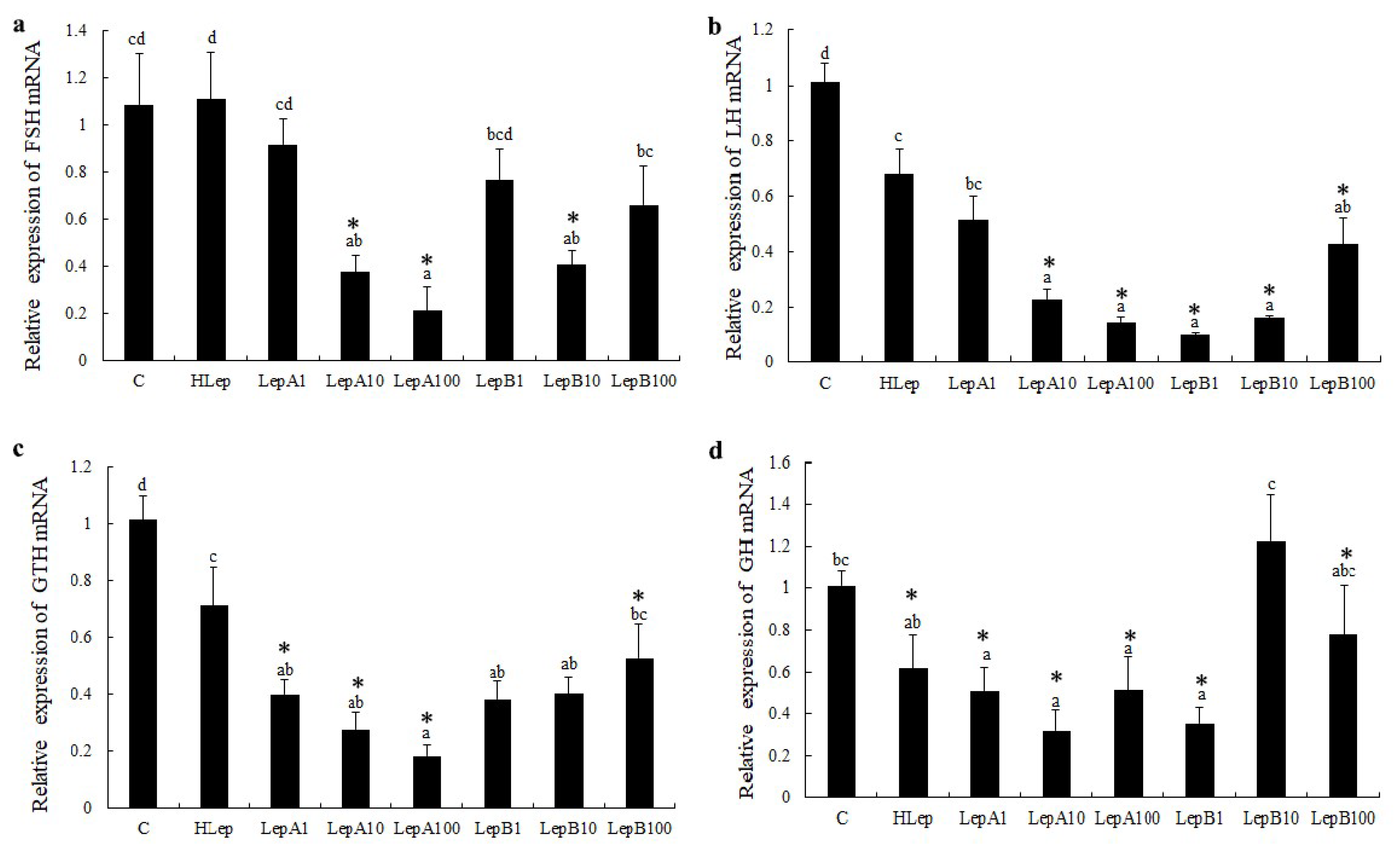
3.2. Pituitary
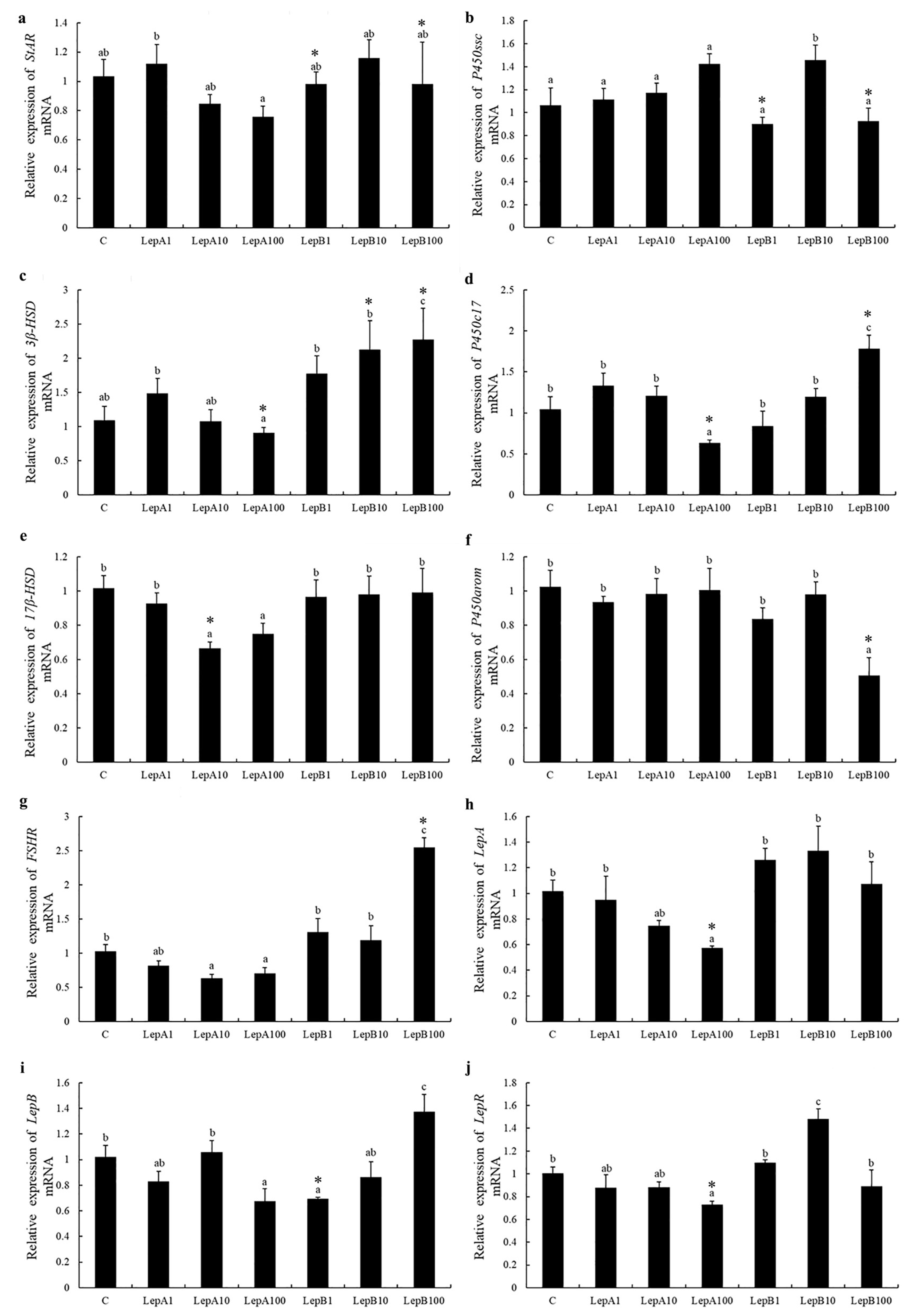
3.3. Ovary
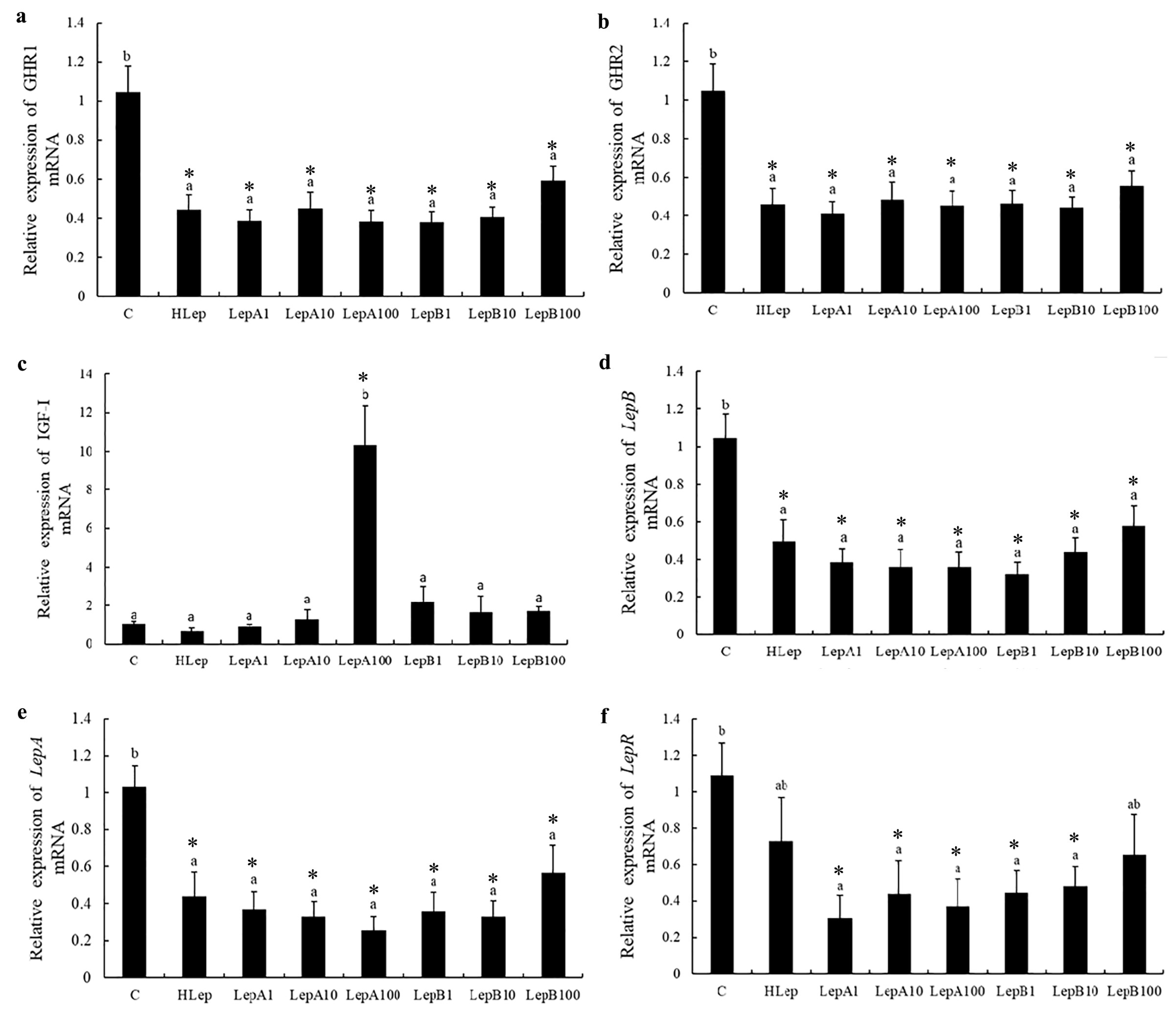
3.4. Liver
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Saladin, R.; De Vos, P.; Guerre-Millo, M.; Leturque, A.; Girard, J.; Staels, B.; Auwerx, J. Transient increase in obese gene expression after food intake or insulin administration. Nature 1995, 377, 527–529. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, T.; Uji, S.; Suzuki, T. Identification of cDNA coding for a homologue to mammalian leptin from pufferfish, Takifugu rubripes. Peptides 2005, 26, 745–750. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Gao, J.; Wu, H.; Cheng, X.; Zhang, Z.; Song, R.; Li, S.; Zhou, J.; Li, C.; Zeng, G. Molecular characterization and expression pattern of leptin in yellow cheek carp (Elopichthys bambusa) and its transcriptional changes in response to fasting and refeeding. Biology 2023, 12, 758. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.H.; Tan, X.Y.; Wu, K.; Zhuo, M.Q.; Song, Y.F.; Chen, Q.L. Regulation and mechanism of leptin on lipid metabolism in ovarian follicle cells from yellow catfish Pelteobagrus fulvidraco. Gen. Comp. Endocrinol. 2015, 222, 116–123. [Google Scholar] [CrossRef]
- Murashita, K.; Uji, S.; Yamamoto, T.; Rønnestad, I.; Kurokawa, T. Production of recombinant leptin and its effects on food intake in rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 150, 377–384. [Google Scholar] [CrossRef]
- Michel, M.; Page-McCaw, P.S.; Chen, W.; Cone, R.D. Leptin signaling regulates glucose homeostasis, but not adipostasis, in the zebrafish. Proc. Natl. Acad. Sci. USA 2016, 113, 3084–3089. [Google Scholar] [CrossRef]
- Li, G.G.; Liang, X.F.; Xie, Q.; Li, G.; Yu, Y.; Lai, K. Gene structure, recombinant expression and functional characterization of grass carp leptin. Gen. Comp. Endocrinol. 2010, 166, 117–127. [Google Scholar] [CrossRef]
- De Pedro, N.; Martínez-Alvarez, R.; Delgado, M.J. Acute and chronic leptin reduces food intake and body weight in goldfish (Carassius auratus). J. Endocrinol. 2006, 188, 513–520. [Google Scholar] [CrossRef]
- Huising, M.O.; Geven, E.J.; Kruiswijk, C.P.; Nabuurs, S.B.; Stolte, E.H.; Spanings, F.A.; Verburg-van Kemenade, B.M.; Flik, G. Increased leptin expression in common Carp (Cyprinus carpio) after food intake but not after fasting or feeding to satiation. Endocrinology 2006, 147, 5786–5797. [Google Scholar] [CrossRef]
- YZhang, B.; Wang, X.; Liu, Y.; Xu, B.; Shi, Y.; Jiang, A.; Cui, Z.; Zhang, R.; Sun, R. Recombinant expression and bioactivity analysis of two leptin genes of Cynoglossus semilaevis. SCXueBao 2019, 43, 2279–2289. [Google Scholar]
- Del Vecchio, G.; Murashita, K.; Verri, T.; Gomes, A.S.; Rønnestad, I. Leptin receptor-deficient (knockout) zebrafish: Effects on nutrient acquisition. Gen. Comp. Endocrinol. 2021, 310, 113832. [Google Scholar] [CrossRef] [PubMed]
- Kamstra, K.; Rizwan, M.Z.; Grattan, D.R.; Horsfield, J.A.; Tups, A. Leptin regulates glucose homeostasis via the canonical Wnt pathway in the zebrafish. Faseb. J. 2022, 36, e22207. [Google Scholar] [CrossRef] [PubMed]
- Trombley, S.; Schmitz, M. Leptin in fish: Possible role in sexual maturation in male Atlantic salmon. Fish. Physiol. Biochem. 2013, 39, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Mariano, G.; Stilo, R.; Terrazzano, G.; Coccia, E.; Vito, P.; Varricchio, E.; Paolucci, M. Effects of recombinant trout leptin in superoxide production and NF-κB/MAPK phosphorylation in blood leukocytes. Peptides 2013, 48, 59–69. [Google Scholar] [CrossRef]
- Zhang, Y.; Chua, S., Jr. Leptin Function and Regulation. Compr. Physiol. 2017, 8, 351–369. [Google Scholar]
- Suter, K.J.; Pohl, C.R.; Wilson, M.E. Circulating concentrations of nocturnal leptin, growth hormone, and insulin-like growth factor-I increase before the onset of puberty in agonadal male monkeys: Potential signals for the initiation of puberty. J. Clin. Endocrinol. Metab. 2000, 85, 808–814. [Google Scholar] [CrossRef]
- Plant, T.M. Leptin, growth hormone, and the onset of primate puberty. J. Clin. Endocrinol. Metab. 2001, 86, 458–460. [Google Scholar] [CrossRef]
- Ghizzoni, L.; Mastorakos, G. Interactions of leptin, GH, and cortisol in normal children. Ann. N. Y. Acad. Sci. 2003, 997, 56–63. [Google Scholar] [CrossRef]
- Zhao, H.; Zeng, C.; Yi, S.; Wan, S.; Chen, B.; Gao, Z. Leptin genes in blunt snout bream: Cloning, phylogeny and expression correlated to gonads development. Int. J. Mol. Sci. 2015, 16, 27609–27624. [Google Scholar] [CrossRef]
- Bagivalu Lakshminarasimha, A.; Puvvada, M.; Hammerschmidt, M.; Michel, M. Leptin modulates oocyte maturation by a central and a direct pathway in zebrafish. J. Endocrinol. 2022, 254, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Weil, C.; Le Bail, P.Y.; Sabin, N.; Le Gac, F. In vitro action of leptin on FSH and LH production in rainbow trout (Onchorynchus mykiss) at different stages of the sexual cycle. Gen. Comp. Endocrinol. 2003, 130, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Ohga, H.; Ito, K.; Kakino, K.; Mon, H.; Kusakabe, T.; Lee, J.M.; Matsuyama, M. Leptin is an important endocrine player that directly activates gonadotropic cells in teleost fish, Chub Mackerel. Cells 2021, 10, 3505. [Google Scholar] [CrossRef] [PubMed]
- Peyon, P.; Zanuy, S.; Carrillo, M. Action of leptin on in vitro luteinizing hormone release in the European sea bass (Dicentrarchus labrax). Biol. Reprod. 2001, 65, 1573–1578. [Google Scholar] [CrossRef] [PubMed]
- Peyon, P.; Vega-Rubín de Celis, S.; Gómez-Requeni, P.; Zanuy, S.; Pérez-Sánchez, J.; Carrillo, M. In vitro effect of leptin on somatolactin release in the European sea bass (Dicentrarchus labrax): Dependence on the reproductive status and interaction with NPY and GnRH. Gen. Comp. Endocrinol. 2003, 132, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Uña, M.; López-Mancheño, Y.; Diéguez, C.; Fernández-Rojo, M.A.; Novelle, M.G. Unraveling the role of leptin in liver function and its relationship with liver diseases. Int. J. Mol. Sci. 2020, 21, 9368. [Google Scholar] [CrossRef] [PubMed]
- Schirman-Hildesheim, T.D.; Gershon, E.; Litichever, N.; Galiani, D.; Ben-Aroya, N.; Dekel, N.; Koch, Y. Local production of the gonadotropic hormones in the rat ovary. Mol. Cell Endocrinol. 2008, 282, 32–38. [Google Scholar] [CrossRef]
- Conchillo, M.; Prieto, J.; Quiroga, J. Insulin-like growth factor I (IGF-I) and liver cirrhosis. Rev. Esp. Enferm. Dig. 2007, 99, 156–164. [Google Scholar]
- Patiño, R.; Yoshizaki, G.; Thomas, P.; Kagawa, H. Gonadotropic control of ovarian follicle maturation: The two-stage concept and its mechanisms. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2001, 129, 427–439. [Google Scholar] [CrossRef]
- Mukherjee, D.; Majumder, S.; Moulick, S.R.; Pal, P.; Mallick, B.; Chakraborty, A.; Gupta, S. Signaling pathways in insulin- and IGF-I mediated oocyte maturation in lower vertebrates. Indian. J. Biochem. Biophys. 2014, 51, 520–526. [Google Scholar]
- Xie, L.; Tang, Q.; Yang, L.; Chen, L. Insulin-like growth factor I promotes oocyte maturation through increasing the expression and phosphorylation of epidermal growth factor receptor in the zebrafish ovary. Mol. Cell Endocrinol. 2016, 419, 198–207. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Ding, Y.; Nowik, N.; Jager, C.; Eeza, M.N.H.; Alia, A.; Baelde, H.J.; Spaink, H.P. Leptin deficiency affects glucose homeostasis and results in adiposity in zebrafish. J. Endocrinol. 2021, 249, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Audira, G.; Sarasamma, S.; Chen, J.R.; Juniardi, S.; Sampurna, B.P.; Liang, S.T.; Lai, Y.H.; Lin, G.M.; Hsieh, M.C.; Hsiao, C.D. Zebrafish mutants carrying leptin a (lepa) gene deficiency display obesity, anxiety, less aggression and fear, and circadian rhythm and color preference dysregulation. Int. J. Mol. Sci. 2018, 19, 4038. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hu, Y.; Cheng, P.; Chen, S. Identification of potential blind-side hypermelanosis-related lncRNA-miRNA-mRNA regulatory network in a flatfish species, Chinese tongue sole (Cynoglossus semilaevis). Front. Genet. 2021, 12, 817117. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhang, Y.; Wang, B.; Liu, X.; Liu, Q.; Song, X.; Shi, B.; Ren, K. Leptin and leptin receptor genes in tongue sole (Cynoglossus semilaevis): Molecular cloning, tissue distribution and differential regulation of these genes by sex steroids. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2018, 224, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Douros, J.D.; Baltzegar, D.A.; Mankiewicz, J.; Taylor, J.; Yamaguchi, Y.; Lerner, D.T.; Seale, A.P.; Grau, E.G.; Breves, J.P.; Borski, R.J. Control of leptin by metabolic state and its regulatory interactions with pituitary growth hormone and hepatic growth hormone receptors and insulin like growth factors in the tilapia (Oreochromis mossambicus). Gen. Comp. Endocrinol. 2017, 240, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Xin, N.; Zhai, Y.; Jiang, L.; Zhai, J.; Zhang, Q.; Qi, J. Reference gene selection for quantitative real-time RT-PCR normalization in the half-smooth tongue sole (Cynoglossus semilaevis) at different developmental stages, in various tissue types and on exposure to chemicals. PLoS ONE 2014, 9, e91715. [Google Scholar] [CrossRef]
- Xu, H.; Cao, L.; Wei, Y.; Zhang, Y.; Liang, M. Effects of different dietary DHA:EPA ratios on gonadal steroidogenesis in the marine teleost, tongue sole (Cynoglossus semilaevis). Br. J. Nutr. 2017, 118, 179–188. [Google Scholar] [CrossRef]
- Wang, B.; Yang, G.; Xu, Y.; Zhang, Y.; Liu, X. In vitro effects of tongue sole LPXRFa and kisspeptin on relative abundance of pituitary hormone mRNA and inhibitory action of LPXRFa on kisspeptin activation in the PKC pathway. Anim. Reprod. Sci. 2019, 203, 1–9. [Google Scholar] [CrossRef]
- Wang, B.; Liu, Q.; Liu, X.; Xu, Y.; Song, X.; Shi, B. Molecular characterization of kiss2 and differential regulation of reproduction-related genes by sex steroids in the hypothalamus of half-smooth tongue sole (Cynoglossus semilaevis). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2017, 213, 46–55. [Google Scholar] [CrossRef]
- Ji, X.S.; Chen, S.L.; Jiang, Y.L.; Xu, T.J.; Yang, J.F.; Tian, Y.S. Growth differences and differential expression analysis of pituitary adenylate cyclase activating polypeptide (PACAP) and growth hormone-releasing hormone (GHRH) between the sexes in half-smooth tongue sole Cynoglossus semilaevis. Gen. Comp. Endocrinol. 2011, 170, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Barash, I.A.; Cheung, C.C.; Weigle, D.S.; Ren, H.; Kabigting, E.B.; Kuijper, J.L.; Clifton, D.K.; Steiner, R.A. Leptin is a metabolic signal to the reproductive system. Endocrinology 1996, 137, 3144–3147. [Google Scholar] [CrossRef] [PubMed]
- Yura, S.; Ogawa, Y.; Sagawa, N.; Masuzaki, H.; Itoh, H.; Ebihara, K.; Aizawa-Abe, M.; Fujii, S.; Nakao, K. Accelerated puberty and late-onset hypothalamic hypogonadism in female transgenic skinny mice overexpressing leptin. J. Clin. Investig. 2000, 105, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Ambati, S.; Duan, J.; Duff, E.; Choi, Y.H.; Hartzell, D.L.; Della-Fera, M.A.; Baile, C.A. Gene expression in arcuate nucleus-median eminence of rats treated with leptin or ciliary neurotrophic factor. Biofactors 2007, 31, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, F.J.; Wuertz, S. Insights into kisspeptin- and leptin-signalling on GnRH mRNA expression in hypothalamic organ cultures of immature pikeperch Sander lucioperca. Int. Aquat. Res. 2016, 8, 191–196. [Google Scholar] [CrossRef][Green Version]
- Ahima, R.S.; Osei, S.Y. Leptin signaling. Physiol. Behav. 2004, 81, 223–241. [Google Scholar] [CrossRef] [PubMed]
- Yaron, Z.; Gur, G.; Melamed, P.; Rosenfeld, H.; Elizur, A.; Levavi-Sivan, B. Regulation of fish gonadotropins. Int. Rev. Cytol. 2003, 225, 131–185. [Google Scholar]
- Hassin, S.; Gothilf, Y.; Blaise, O.; Zohar, Y. Gonadotropin-I and -II subunit gene expression of male striped bass (Morone saxatilis) after gonadotropin-releasing hormone analogue injection: Quantitation using an optimized ribonuclease protection assay. Biol. Reprod. 1999, 58, 1233–1240. [Google Scholar] [CrossRef][Green Version]
- An, K.W.; Nelson, E.R.; Habibi, H.R.; Choi, C.Y. Molecular characterization and expression of three GnRH forms mRNA during gonad sex-change process, and effect of GnRHa on GTH subunits mRNA in the protandrous black porgy (Acanthopagrus schlegeli). Gen. Comp. Endocrinol. 2008, 159, 38–45. [Google Scholar] [CrossRef]
- Chen, H.P.; Cui, X.F.; Wang, Y.R.; Li, Z.Y.; Tian, C.X.; Jiang, D.N.; Zhu, C.H.; Zhang, Y.; Li, S.S.; Li, G.L. Identification, functional characterization, and estrogen regulation on gonadotropin-releasing hormone in the spotted scat, Scatophagus argus. Fish. Physiol. Biochem. 2020, 46, 1743–1757. [Google Scholar] [CrossRef]
- Ren, X.; Huang, Y.; Li, X.; Li, Z.; Yang, H.; He, R.; Zhong, H.; Li, G.; Chen, H. Identification and functional characterization of gonadotropin-releasing hormone in pompano (Trachinotus ovatus). Gen. Comp. Endocrinol. 2022, 316, 113958. [Google Scholar] [CrossRef] [PubMed]
- Zohar, Y.; Muñoz-Cueto, J.A.; Elizur, A.; Kah, O. Neuroendocrinology of reproduction in teleost fish. Gen. Comp. Endocrinol. 2010, 165, 438–455. [Google Scholar] [CrossRef] [PubMed]
- Tilbrook, A.J.; Clarke, I.J. Negative feedback regulation of the secretion and actions of gonadotropin-releasing hormone in males. Biol. Reprod. 2001, 64, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Stocco, D.M. The role of the StAR protein in steroidogenesis: Challenges for the future. J. Endocrinol. 2000, 164, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Nagahama, Y.; Yamashita, M. Regulation of oocyte maturation in fish. Dev. Growth Differ. 2008, 50 (Suppl. S1), S195–S219. [Google Scholar] [CrossRef]
- Senthilkumaran, B.; Sudhakumari, C.C.; Chang, X.T.; Kobayashi, T.; Oba, Y.; Guan, G.; Yoshiura, Y.; Yoshikuni, M.; Nagahama, Y. Ovarian carbonyl reductase-like 20beta-hydroxysteroid dehydrogenase shows distinct surge in messenger RNA expression during natural and gonadotropin-induced meiotic maturation in nile tilapia. Biol. Reprod. 2002, 67, 1080–1086. [Google Scholar] [CrossRef]
- Fouad Mansour, M.; Pelletier, M.; Boulet, M.M.; Mayrand, D.; Brochu, G.; Lebel, S.; Poirier, D.; Fradette, J.; Cianflone, K.; Luu-The, V.; et al. Oxidative activity of 17β-hydroxysteroid dehydrogenase on testosterone in male abdominal adipose tissues and cellular localization of 17β-HSD type 2. Mol. Cell Endocrinol. 2015, 414, 168–176. [Google Scholar] [CrossRef]
- Hojo, Y.; Hattori, T.A.; Enami, T.; Furukawa, A.; Suzuki, K.; Ishii, H.T.; Mukai, H.; Morrison, J.H.; Janssen, W.G.; Kominami, S.; et al. Adult male rat hippocampus synthesizes estradiol from pregnenolone by cytochromes P45017alpha and P450 aromatase localized in neurons. Proc. Natl. Acad. Sci. USA 2004, 101, 865–870. [Google Scholar] [CrossRef]
- Trombley, S.; Rocha, A.; Schmitz, M. Sex steroids stimulate leptin gene expression in Atlantic salmon parr hepatocytes in vitro. Gen. Comp. Endocrinol. 2015, 221, 156–164. [Google Scholar] [CrossRef]
- Kumar, P.A.; Sivakumar, A.V.N.; Pathipati, D.; Chakravarthi, V.P.; Brahmaiah, K.V.; Rao, V.H. Leptin induced in vitro development of ovarian follicles in sheep is related to the expression of P450 aromatase and steroidogenesis. Theriogenology 2019, 136, 1–6. [Google Scholar] [CrossRef]
- Flier, J.S.; Maratos-Flier, E. Lasker lauds leptin. Cell 2010, 143, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Blanco, A.M.; Soengas, J.L. Leptin signalling in teleost fish with emphasis in food intake regulation. Mol. Cell Endocrinol. 2021, 526, 111209. [Google Scholar] [CrossRef] [PubMed]
- Angotzi, A.R.; Stefansson, S.O.; Nilsen, T.O.; Rathore, R.M.; Rønnestad, I. Molecular cloning and genomic characterization of novel leptin-like genes in salmonids provide new insight into the evolution of the Leptin gene family. Gen. Comp. Endocrinol. 2013, 187, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Douros, J.D.; Baltzegar, D.A.; Breves, J.P.; Lerner, D.T.; Seale, A.P.; Grau, E.G.; Borski, R.J. Prolactin is a major inhibitor of hepatic Leptin A synthesis and secretion: Studies utilizing a homologous Leptin A ELISA in the tilapia. Gen. Comp. Endocrinol. 2014, 207, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Won, E.T.; Baltzegar, D.A.; Picha, M.E.; Borski, R.J. Cloning and characterization of leptin in a Perciform fish, the striped bass (Morone saxatilis): Control of feeding and regulation by nutritional state. Gen. Comp. Endocrinol. 2012, 178, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Salmerón, C.; Johansson, M.; Angotzi, A.R.; Rønnestad, I.; Jönsson, E.; Björnsson, B.T.; Gutiérrez, J.; Navarro, I.; Capilla, E. Effects of nutritional status on plasma leptin levels and in vitro regulation of adipocyte leptin expression and secretion in rainbow trout. Gen. Comp. Endocrinol. 2015, 210, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Utz, A.L.; Yamamoto, A.; Hemphill, L.; Miller, K.K. Growth hormone deficiency by growth hormone releasing hormone-arginine testing criteria predicts increased cardiovascular risk markers in normal young overweight and obese women. J. Clin. Endocrinol. Metab. 2008, 93, 2507–2514. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chaves, V.E.; Júnior, F.M.; Bertolini, G.L. The metabolic effects of growth hormone in adipose tissue. Endocrine 2013, 44, 293–302. [Google Scholar] [CrossRef]
- Carotti, S.; Guarino, M.P.L.; Valentini, F.; Porzio, S.; Vespasiani-Gentilucci, U.; Perrone, G.; Zingariello, M.; Gallo, P.; Cicala, M.; Picardi, A.; et al. Impairment of GH/IGF-1 axis in the liver of patients with HCV-related chronic hepatitis. Horm. Metab. Res. 2018, 50, 145–151. [Google Scholar] [CrossRef]
- Elias, C.F.; Purohit, D. Leptin signaling and circuits in puberty and fertility. Cell Mol. Life Sci. 2013, 70, 841–862. [Google Scholar] [CrossRef]
- Landry, D.; Cloutier, F.; Martin, L.J. Implications of leptin in neuroendocrine regulation of male reproduction. Reprod. Biol. 2013, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Frøiland, E.; Jobling, M.; Björnsson, B.T.; Kling, P.; Ravuri, C.S.; Jørgensen, E.H. Seasonal appetite regulation in the anadromous Arctic charr: Evidence for a role of adiposity in the regulation of appetite but not for leptin in signalling adiposity. Gen. Comp. Endocrinol. 2012, 178, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Gorissen, M.; Flik, G. Leptin in teleostean fish, towards the origins of leptin physiology. J. Chem. Neuroanat. 2014, 61–62, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.M.; Johnson, T.M.; Londraville, R.L. Evidence for leptin expression in fishes. J. Exp. Zool. 2000, 286, 718–724. [Google Scholar] [CrossRef]
- Won, E.T.; Douros, J.D.; Hurt, D.A.; Borski, R.J. Leptin stimulates hepatic growth hormone receptor and insulin-like growth factor gene expression in a teleost fish, the hybrid striped bass. Gen. Comp. Endocrinol. 2016, 229, 84–91. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
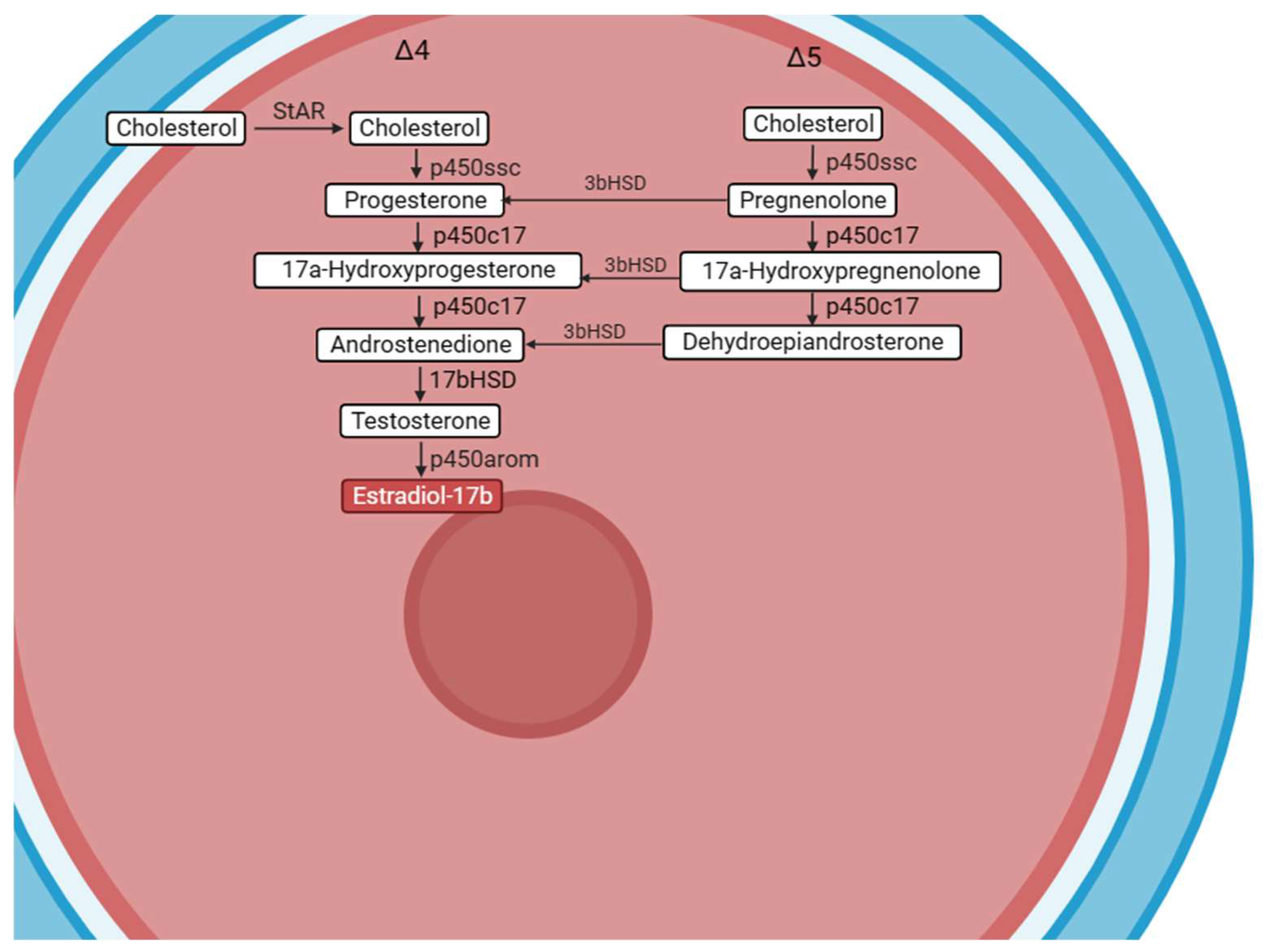{kind=link}
| Tissue Detect | Full Name | Gene Name | Forward Primer Sequence 5′-3′ | Reverse Primer Sequence 5′-3′ | Reference |
|---|---|---|---|---|---|
| Gonad | steroidogenic acute regulatory protein | Csstar | AGGACGGCTGGACCACTGAAAT | ACCTCGTGGGTGACCATCGTGT | [38] |
| Gonad | P450 cholesterol-side-chain-cleavage | csp450ssc | TTCTGTGCTGTATGGCGAAC | CTTTTGACCCAATCCGTCTC | |
| Gonad | 3β-hydroxysteroid dehydrogenase | cs3b-hsd | CACCACTGGGTAAGCACTATC | AGGTTATCGCAAACAGCATT | |
| Gonad | P450 17α-hydroxylase/17,20-lyase | csp450c17 | GCCCACTCGCTCCCTACATACT | GTCTTTCCCATCTCGGGTCAG | |
| Gonad | 17β-hydroxysteroid dehydrogenase | cs17b-hsd | AATGTGCAGGCTCTAACTGCTTC | AGGTTCCTCATGGTGGCGTA | |
| Gonad | P450 aromatase | csp450arom | TGCGATTTCAGCCCGT | TGCGACCCGTGTTCAGA | |
| Gonad | follicle stimulating hormone receptor | Csfshr | AAGATCAAGGGAAAACGCTA | CTCAGATGGTTGGAGGAAAG | [39] |
| Pituitary | growth hormone | Csgh | TTATAGACCAGCGGCGTTTC | ATGCTTGTTGTCGGGGATG | |
| Pituitary | luteinizing hormone | Cslhb | TCCACCTGACACTAACGCTG | GTTTGGTTCCTTTGTTCTGC | |
| Pituitary | follicle stimulating hormone b | Csfshb | TGATGGGTGTCCAGAGGAAG | CAACAAACCGTCCACAGTCC | |
| Four tissues | Leptin A | Cslepa | ATTCTTCAGCAGCACGACGAG | CCTGGATGGTTACAACAGCCTTA | [35] |
| Four tissues | leptin B | Cslepb | TGCGTCTATGATGTGTTGGCT | GTCTGGTCGCTGCTGTTGGTT | |
| Four tissues | leptin receptor | Cslepr | TTGGTTTGACTTCACTCCTGGT | AATACCGCTGTTGTCGCTTCT | |
| Liver | growth hormone receptor 1 | csghr1 | AGATGTTGAAGAAAGGGAAGTTGG | GGTTGATAGGTTGGGGCAGA | |
| Liver | growth hormone receptor 2 | csghr2 | CGTCACTTGAAGATGTGCCCCA | CAAGAGGAGATTTTTTGTTGATGAA | |
| Liver | insulin-like growth factor-Ι | csigf-1 | GTATCTCCTGTAGCCACACCCTCT | GCCTCTCTCTCCACACACAAACT | |
| Hypothalamus | gonadotropin-inhibitory hormone | Csgnih | GGAAATCAGCCTACAGTGACAAAA | GCCTCTCCAAGTCCAAACTCC | [40] |
| Hypothalamus | gonadotropin-inhibitory hormone receptor | Csgnihr | GCTTTTCATGTTGTCCTGGTTG | GGGTTGATGCTTGAGTTGGAG | |
| Hypothalamus | kisspeptin2 | cskiss2 | GGCAACTGCTGTGCAACGA | AAGACAGAAAGCGGGGAGAAC | |
| Hypothalamus | kisspeptin2 receptor | cskiss2r | AGTTGTGATCGTCCTCCTCTTTG | AGTTGGGTTGGTATTTGGGATG | |
| Hypothalamus | gonadotropin-releasing hormone2 | csgnrh2 | GGAATCTGAACTGGAGAACTGCT | TGGCTGCTCACAACTTTATCAC | |
| Hypothalamus | gonadotropin-releasing hormone3 | csgnrh3 | AGGCAGCAGAGTGATCGTG | CACCTGGTAGCCATCCATAAGAC | |
| Four tissues | 18s rRNA | 18S | GGTCTGTGATGCCCTTAGATGTC | AGTGGGGTTCAGCGGGTTAC | |
| Pituitary | gonadotropin | Csgth | TCAGCAGTTTGGAGGTGCTT | GTCCAGTACACTCGGTGGTG | [39] |
| Hypothalamus | gonadotropin-releasing hormone | Csgnrh | TGTGTTTCGGCTGGTTCTGTTG | GAATGCTCCTCCTCTGGGGTC | [40] |
| Gonad | Pituitary adenylate cyclase-activating polypeptide | Cspacap | CCTGCTGACCCAGAATCCTAC | AAAATCAGCAGCGCAACGAG | [41] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, X.; Zhang, Y.; Wang, B.; Cui, A.; Jiang, Y.; Meng, Z.; Li, Y.; Xu, Y. Effects of Recombinant Leptin Proteins on the Expression of Key Genes in the HPG Axis and Liver of Tongue Sole In Vitro. Fishes 2023, 8, 608. https://doi.org/10.3390/fishes8120608
Cai X, Zhang Y, Wang B, Cui A, Jiang Y, Meng Z, Li Y, Xu Y. Effects of Recombinant Leptin Proteins on the Expression of Key Genes in the HPG Axis and Liver of Tongue Sole In Vitro. Fishes. 2023; 8(12):608. https://doi.org/10.3390/fishes8120608
Chicago/Turabian StyleCai, Xin, Yaxing Zhang, Bin Wang, Aijun Cui, Yan Jiang, Zhaojun Meng, Yuting Li, and Yongjiang Xu. 2023. "Effects of Recombinant Leptin Proteins on the Expression of Key Genes in the HPG Axis and Liver of Tongue Sole In Vitro" Fishes 8, no. 12: 608. https://doi.org/10.3390/fishes8120608
APA StyleCai, X., Zhang, Y., Wang, B., Cui, A., Jiang, Y., Meng, Z., Li, Y., & Xu, Y. (2023). Effects of Recombinant Leptin Proteins on the Expression of Key Genes in the HPG Axis and Liver of Tongue Sole In Vitro. Fishes, 8(12), 608. https://doi.org/10.3390/fishes8120608