
Synergistic Effects of Climate Change and Alien Fish Invasions in Freshwater Ecosystems: A Review




Abstract
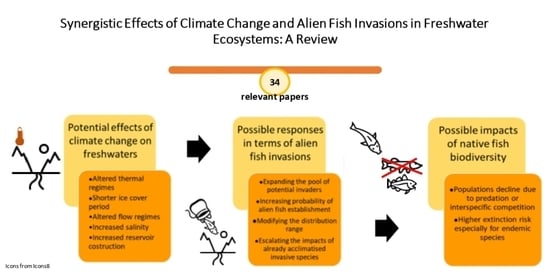
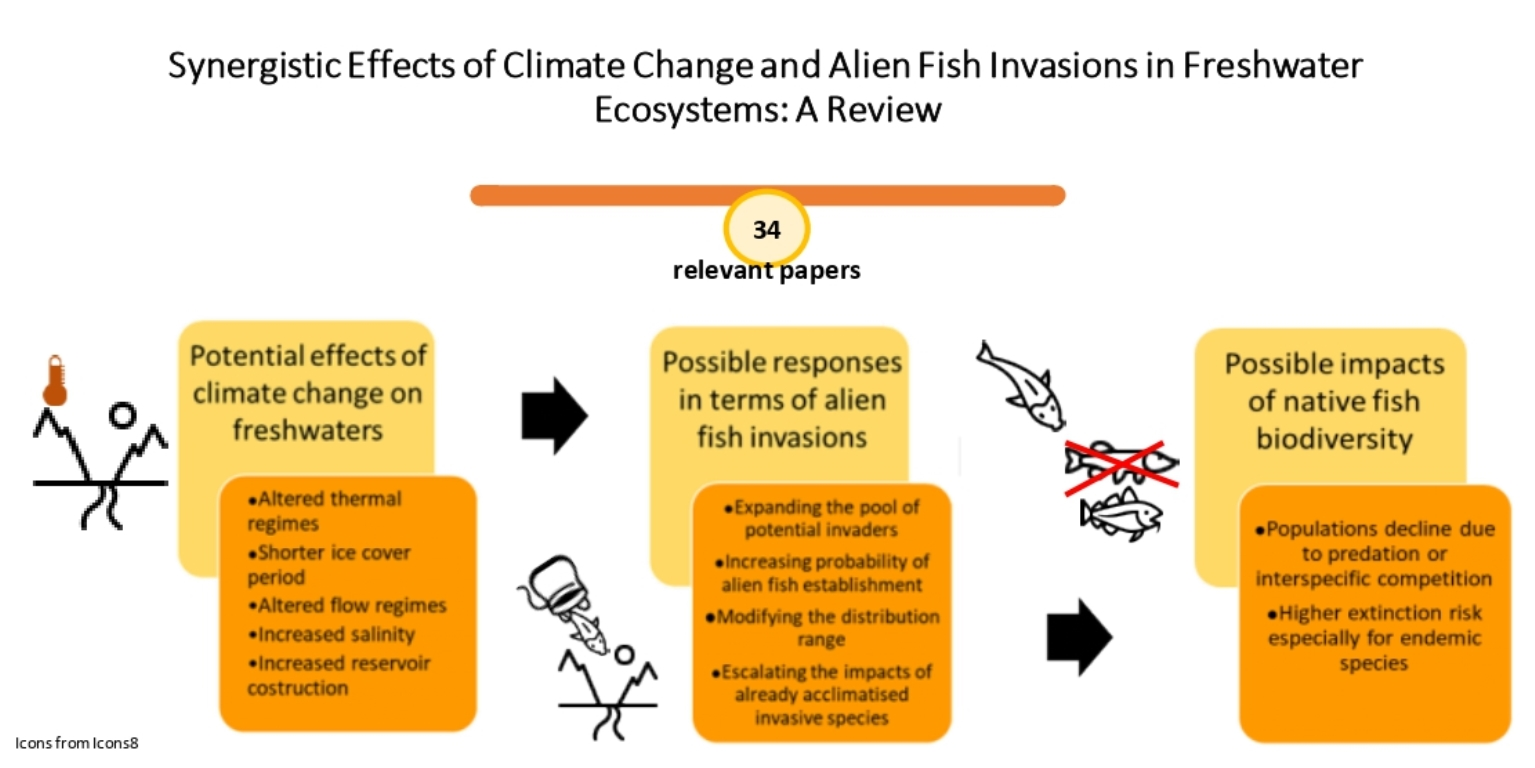
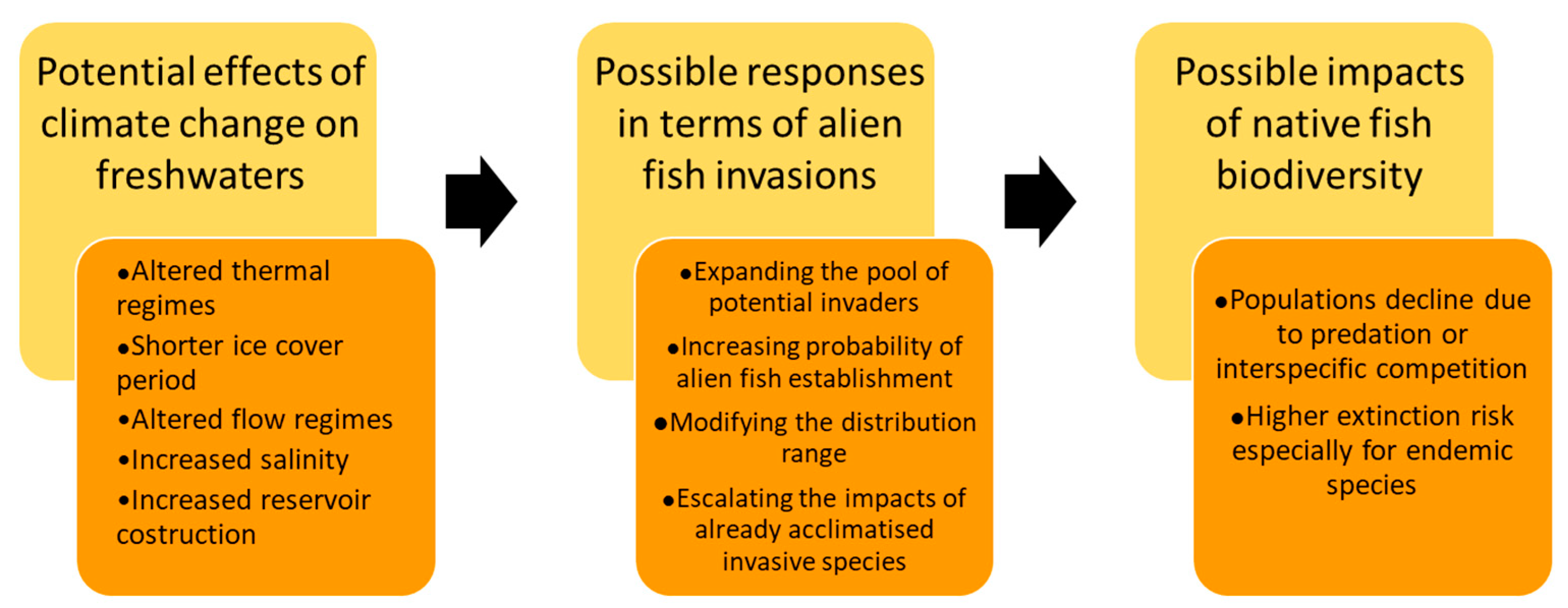
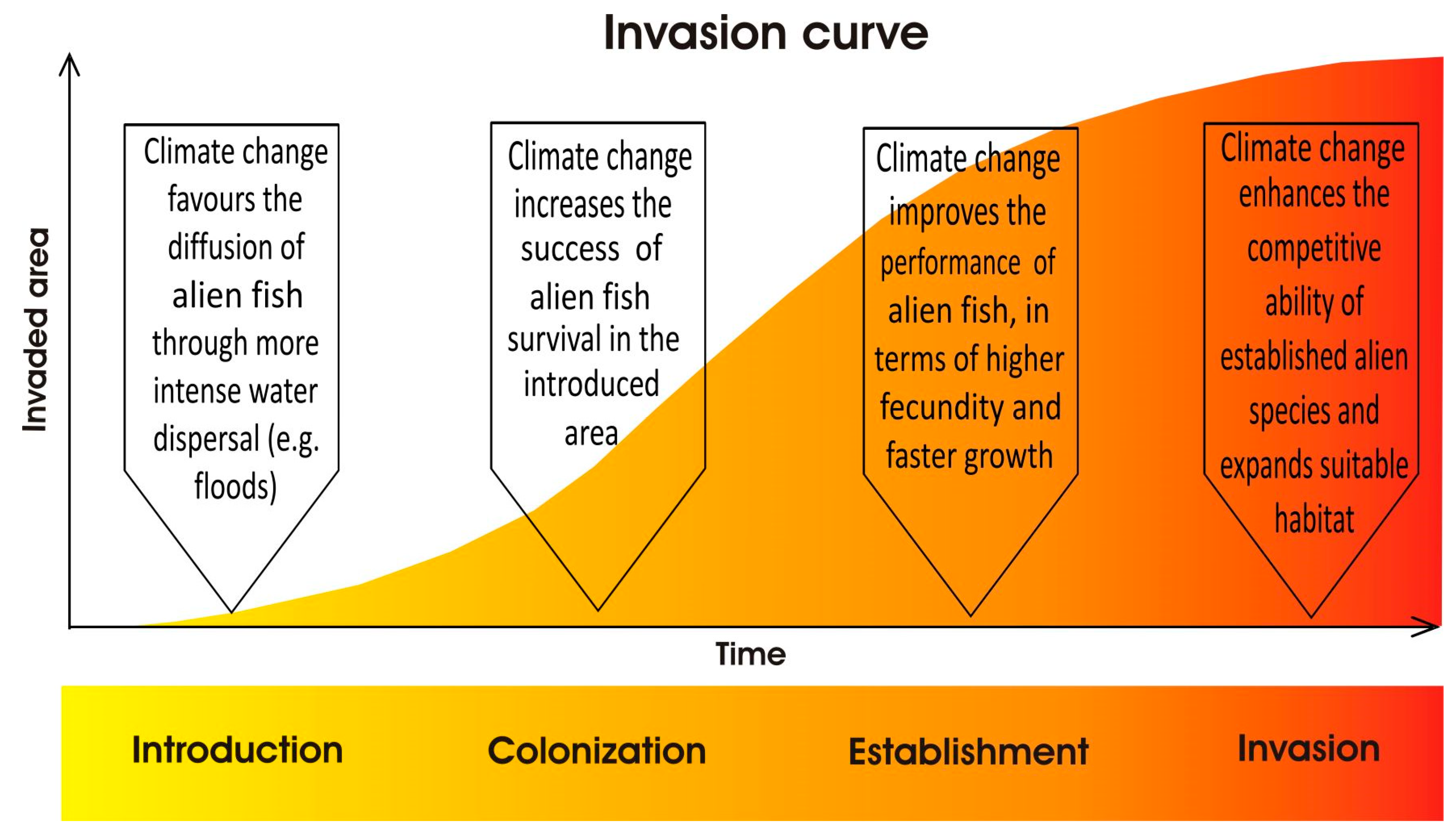
1. Introduction
2. Materials and Methods
3. Results
3.1. Altered Ecological Connectivity in Freshwaters
3.1.1. Impacts of Altered Ecological Connectivity in Inland Waters
3.1.2. Implications for Climate Changes, Alien Fish Invasions, and Native Biodiversity
3.1.3. Possible Mitigation Measures and Management Implications
3.1.4. Knowledge Gaps and Future Directions
3.2. Altered Species Distribution Due to Changes in Thermal Regime
3.2.1. Ecological Impacts of Altered Thermal Regime in Freshwaters
3.2.2. Implications for Climate Changes, Alien Fish Invasions, and Native Biodiversity
3.2.3. Possible Mitigation Measures and Management Implications
3.2.4. Knowledge Gaps and Future Research Directions
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cantonati, M.; Poikane, S.; Pringle, C.M.; Stevens, L.E.; Turak, E.; Heino, J.; Richardson, J.S.; Bolpagni, R.; Borrini, A.; Cid, N.; et al. Characteristics, main impacts, and stewardship of natural and artificial freshwater environments: Consequences for biodiversity conservation. Water 2020, 12, 260. [Google Scholar] [CrossRef]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef] [PubMed]
- Strayer, D.L.; Dudgeon, D. Freshwater biodiversity conservation: Recent progress and future challenges. J. N. Am. Benthol. Soc. 2010, 29, 344–358. [Google Scholar] [CrossRef]
- Geist, J. Integrative freshwater ecology and biodiversity conservation. Ecol. Indic. 2011, 11, 1507–1516. [Google Scholar] [CrossRef]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.-H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Dudgeon, D. Multiple threats imperil freshwater biodiversity in the Anthropocene. Curr. Biol. 2019, 29, R960–R967. [Google Scholar] [CrossRef] [PubMed]
- WWF (World Wildlife Fund). Living Planet Report 2022—Building a Nature-Positive Society; WWF: Gland, Switzerland, 2022. [Google Scholar]
- Smith, K.G.; Barrios, V.; Darwall, W.R.T.; Numa, C. The status and distribution of freshwater biodiversity in the Eastern Mediterranean; IUCN: Cambridge, UK; Malaga, Spain; Gland, Switzerland, 2014; p. 132. [Google Scholar]
- GEO BON and FWBON. Inland Waters in the Post-2020 Global Biodiversity Framework. 2022. Available online: https://geobon.org/science-briefs/ (accessed on 18 August 2023).
- Poff, N.L.; Olden, J.D.; Strayer, D.L. Climate change and freshwater fauna extinction risk. In Saving a Million Species: Extinction Risk from Climate Change; Hannah, L., Ed.; Island Press/Center for Resource Economics: Washington, DC, USA, 2012; pp. 309–336. [Google Scholar] [CrossRef]
- Darwall, W.R.; Freyhof, J. Lost fishes, who is counting? The extent of the threat to freshwater fish biodiversity. In Conservation of Freshwater Fishes, 1st ed.; Closs, G.P., Krkosek, M., Olden, J.D., Eds.; Cambridge University Press: Cambridge, UK, 2016; pp. 1–36. [Google Scholar]
- Fricke, R.; Eschmeyer, W.N.; Van der Laan, R. (Eds.) Eschmeyer’s Catalog of Fishes: Genera, Species, References. Version of 11 Jan 2023. Available online: https://www.calacademy.org/scientists/projects/eschmeyers-catalog-of-fishes (accessed on 5 February 2023).
- Abell, R.; Thieme, M.L.; Revenga, C.; Bryer, M.; Kottelat, M.; Bogutskaya, N.; Coad, B.; Mandrak, N.; Balderas, S.C.; Bussing, W.; et al. Freshwater ecoregions of the world: A new map of biogeographic units for freshwater biodiversity conservation. BioScience 2008, 58, 403–414. [Google Scholar] [CrossRef]
- Geiger, M.F.; Herder, F.; Monaghan, M.T.; Almada, V.; Barbieri, R.; Bariche, M.; Berrebi, P.; Bohlen, J.; Casal-Lopez, M.; Delmastro, G.B.; et al. Spatial heterogeneity in the Mediterranean Biodiversity Hotspot affects barcoding accuracy of its freshwater fishes. Mol. Ecol. Resour. 2014, 14, 1210–1221. [Google Scholar] [CrossRef]
- Ramsar Convention on Wetlands. Global Wetland Outlook: State of the World’s Wetlands and their Services to People; Ramsar Convention Secretariat: Gland, Switzerland, 2018; p. 84. [Google Scholar]
- Jordaan, M.S.; Chakona, A.; Van der Colff, D. Protected areas and endemic freshwater fishes of the Cape Fold Ecoregion: Missing the boat for fish conservation? Front. Environ. Sci. 2020, 8, 502042. [Google Scholar] [CrossRef]
- Doria, C.R.D.C.; Agudelo, E.; Akama, A.; Barros, B.; Bonfim, M.; Carneiro, L.; Briglia-Ferreira, S.R.; Carvalho, L.N.; Bonilla-Castillo, C.A.; Charvet, P.; et al. The Silent Threat of Non-native Fish in the Amazon: ANNF Database and Review. Front. Ecol. Evol. 2021, 9, 646702. [Google Scholar] [CrossRef]
- Collen, B.; Whitton, F.; Dyer, E.E.; Baillie, J.E.; Cumberlidge, N.; Darwall, W.R.; Pollock, C.; Richman, N.I.; Soulsby, A.-M.; Böhm, M. Global patterns of freshwater species diversity, threat and endemism. Glob. Ecol. Biogeogr. 2014, 23, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Woodward, G.; Perkins, D.M.; Brown, L.E. Climate change and freshwater ecosystems: Impacts across multiple levels of organization. Phil. Trans. R. Soc. B 2010, 365, 2093–2106. [Google Scholar] [CrossRef]
- Morrongiello, J.R.; Beatty, S.J.; Bennett, J.C.; Crook, D.A.; Ikedife, D.N.; Kennard, M.J.; Kerezsy, A.; Lintermans, M.; McNeil, D.G.; Pusey, B.J.; et al. Climate change and its implications for Australia’s freshwater fish. Mar. Freshw. Res. 2011, 62, 1082–1098. [Google Scholar] [CrossRef]
- Jeppesen, E.; Meerhoff, M.; Holmgren, K.; González-Bergonzoni, I.; Teixeira-de Mello, F.; Declerck, S.A.; De Meester, L.; Søndergaard, M.; Lauridsen, T.L.; Bjerring, R.; et al. Impacts of climate warming on lake fish community structure and potential effects on ecosystem function. Hydrobiologia 2010, 646, 73–90. [Google Scholar] [CrossRef]
- Sharma, S.; Richardson, D.C.; Woolway, R.I.; Imrit, M.A.; Bouffard, D.; Blagrave, K.; Daly, J.; Filazzola, A.; Granin, N.; Korhonen, J.; et al. Loss of ice cover, shifting phenology, and more extreme events in Northern Hemisphere lakes. J. Geophys. Res. Biogeosci. 2021, 126, e2021JG006348. [Google Scholar] [CrossRef]
- Patil, R.; Wei, Y.; Pullar, D.; Shulmeister, J. Sensitivity of streamflow patterns to river regulation and climate change and its implications for ecological and environmental management. J. Environ. Manag. 2022, 319, 115680. [Google Scholar] [CrossRef] [PubMed]
- Dhungel, S.; Tarboton, D.G.; Jin, J.; Hawkins, C.P. Potential Effects of Climate Change on Ecologically Relevant Streamflow Regimes. River Res. Appl. 2016, 32, 1827–1840. [Google Scholar] [CrossRef]
- Castillo, A.M.; Sharpe, D.M.; Ghalambor, C.K.; De León, L. Exploring the effects of salinization on trophic diversity in freshwater ecosystems: A quantitative review. Hydrobiologia 2018, 807, 1–17. [Google Scholar] [CrossRef]
- Fenoglio, S.; Bo, T.; Cucco, M.; Mercalli, L.; Malacarne, G. Effects of global climate change on freshwater biota: A review with special emphasis on the Italian situation. Ital. J. Zool. 2010, 77, 374–383. [Google Scholar] [CrossRef]
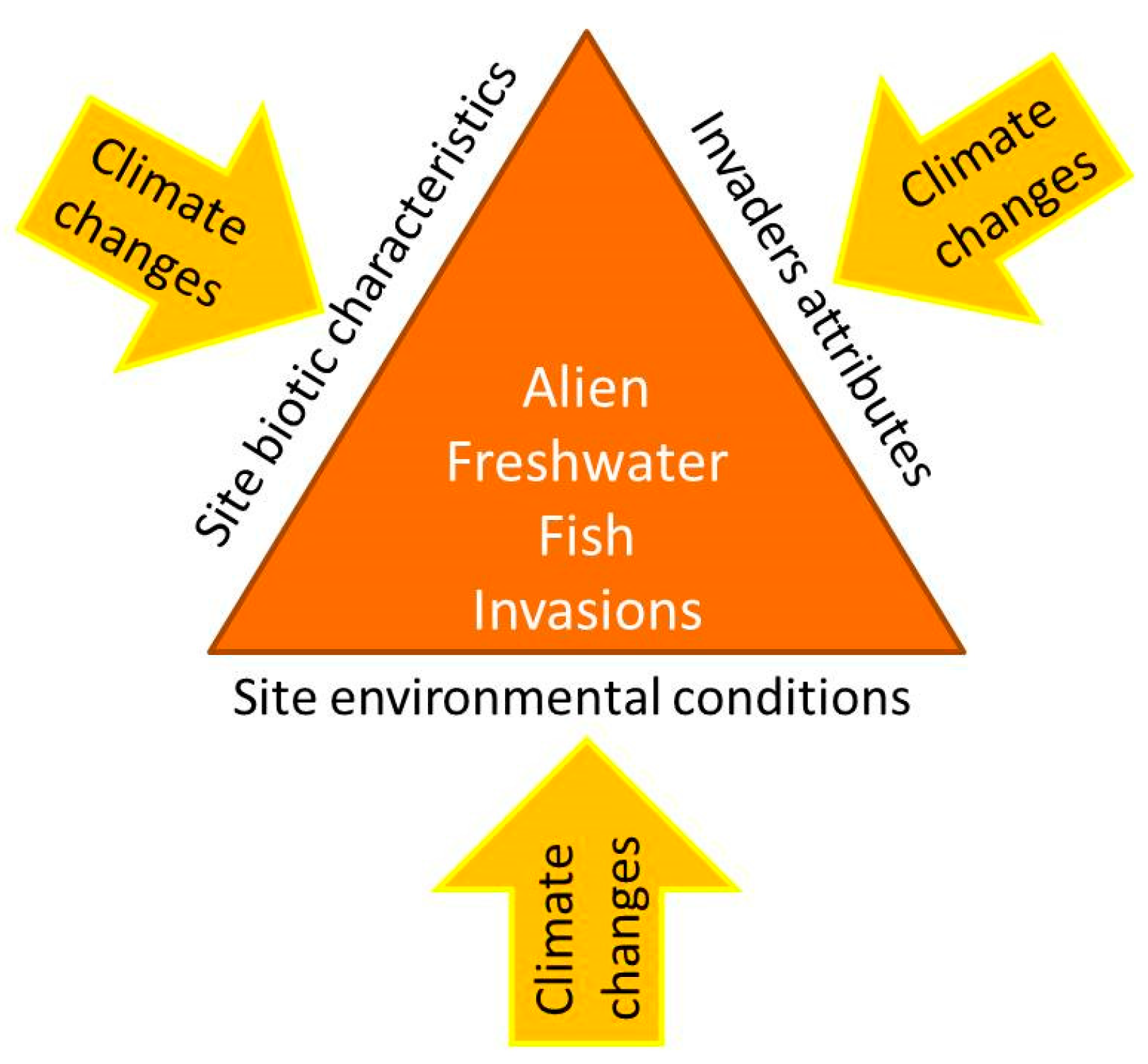
- Perkins, L.B.; Leger, E.A.; Nowak, R.S. Invasion triangle: An organizational framework for species invasion. Ecol. Evol. 2011, 1, 610–625. [Google Scholar] [CrossRef]
- Hulme, P.E. Climate change and biological invasions: Evidence, expectations, and response options. Biol. Rev. 2017, 92, 1297–1313. [Google Scholar] [CrossRef] [PubMed]
- Döll, P.; Bunn, S.E. Cross-chapter box on the impact of climate change on freshwater ecosystems due to altered river flow regimes. In Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V.R., Dokken, D.J., Mach, K.J., Mastrandrea, M.D., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. 143–146. [Google Scholar]
- Liew, J.H.; Tan, H.H.; Yeo, D.C. Dammed rivers: Impoundments facilitate fish invasions. Freshw. Biol. 2016, 61, 1421–1429. [Google Scholar] [CrossRef]
- Clavero, M.; Hermoso, V. Reservoirs promote the taxonomic homogenization of fish communities within river basins. Biodivers. Conserv. 2011, 20, 41–57. [Google Scholar] [CrossRef]
- Zenetos, A.; Çinar, M.E.; Pancucci-Papadopoulou, M.A.; Harmelin, J.G.; Furnari, G.; Andaloro, F.; Bellou, N.; Streftaris, N.; Zibrowius, H. Annotated list of marine alien species in the Mediterranean with records of the worst invasive species. Mediterr. Mar. Sci. 2005, 6, 63–118. [Google Scholar] [CrossRef]
- Lockwood, J.L.; Cassey, P.; Blackburn, T. The role of propagule pressure in explaining species invasions. Trends Ecol. Evol. 2005, 20, 223–228. [Google Scholar] [CrossRef]
- Di Santo, V.; Jordan, H.L.; Cooper, B.; Currie, R.J.; Beitinger, T.L.; Bennett, W.A. Thermal tolerance of the invasive red-bellied pacu and the risk of establishment in the United States. J. Therm. Biol. 2018, 74, 110–115. [Google Scholar] [CrossRef]
- Ficke, A.D.; Myrick, C.A.; Hansen, L.J. Potential impacts of global climate change on freshwater fisheries. Rev. Fish Biol. Fish. 2007, 17, 581–613. [Google Scholar] [CrossRef]
- Walther, G.R.; Roques, A.; Hulme, P.E.; Sykes, M.T.; Pyšek, P.; Kühn, I.; Zobel, M.; Bacher, S.; Botta-Dukát, Z.; Bugmann, H.; et al. Alien species in a warmer world: Risks and opportunities. Trends Ecol. Evol. 2009, 24, 686–693. [Google Scholar] [CrossRef]
- Nunn, A.D.; Cowx, I.G.; Frear, P.A.; Harvey, J.P. Is water temperature an adequate predictor of recruitment success in cyprinid fish populations in lowland rivers? Freshw. Biol. 2003, 48, 579–588. [Google Scholar] [CrossRef]
- Johnson, A.C.; Acreman, M.C.; Dunbar, M.J.; Feist, S.W.; Giacomello, A.M.; Gozlan, R.E.; Hinsley, S.A.; Ibbotson, A.T.; Jarvie, H.P.; Jones, J.I.; et al. The British river of the future: How climate change and human activity might affect two contrasting river ecosystems in England. Sci. Total Environ. 2009, 407, 4787–4798. [Google Scholar] [CrossRef]
- Invasive Species Council. Inquiry into Ecosystem Decline in Victoria by the Environment and Planning Committee of the Victorian Parliament. Submission by the Invasive Species Council November 2020. Available online: https://www.parliament.vic.gov.au/images/stories/committees/SCEP/Ecosystem_Decline/submissions/S943_-_Invasive_Species_Council_Redacted.pdf (accessed on 1 February 2023).
- Sharma, S.; Vander Zanden, M.J.; Magnuson, J.J.; Lyons, J. Comparing climate change and species invasions as drivers of coldwater fish population extirpations. PLoS ONE 2011, 6, e22906. [Google Scholar] [CrossRef] [PubMed]
- Muhlfeld, C.C.; Kovach, R.P.; Jones, L.A.; Al-Chokhachy, R.; Boyer, M.C.; Leary, R.F.; Lowe, W.H.; Luikart, G.; Allendorf, F.W. Invasive hybridization in a threatened species is accelerated by climate change. Nat. Clim. Chang. 2014, 4, 620–624. [Google Scholar] [CrossRef]
- Gomez-Maldonado, S.; Camacho-Cervantes, M. Effect of a temperature gradient on the behaviour of an endangered Mexican topminnow and an invasive freshwater fish. Sci. Rep. 2022, 12, 20584. [Google Scholar] [CrossRef] [PubMed]
- DeWeber, J.T.; Baer, J.; Rösch, R.; Brinker, A. Turning summer into winter: Nutrient dynamics, temperature, density dependence and invasive species drive bioenergetic processes and growth of a keystone coldwater fish. Oikos 2022, 2022, e09316. [Google Scholar] [CrossRef]
- Avlijaš, S.; Mandrak, N.E.; Ricciardi, A. Effects of substrate and elevated temperature on the growth and feeding efficiency of an invasive cyprinid fish, Tench (Tinca tinca). Biol. Invasions 2022, 24, 2383–2397. [Google Scholar] [CrossRef]
- To, D.A.L.; Gomez, G.M.L.; Ramos, D.R.Y.; Palillo, C.D.V.; Go, N.K.C.; Gilles, A.S.; Briones, J.C.A.; Pavia, R.T.B. Invasiveness Risk Assessment of Non-native Freshwater Fish Species (Order: Siluriformes) for Lake Taal, Philippines. Philipp. J. Sci. 2022, 151, 833–841. [Google Scholar] [CrossRef]
- Cerrilla, C.; Afrika, J.; Impson, D.; Jordaan, M.S.; Kotze, N.; Paxton, B.R.; Reed, C.; Schumann, M.; van der Walt, J.A.; Shelton, J.M. Rapid population decline in one of the last recruiting populations of the endangered Clanwilliam sandfish (Labeo seeberi): The roles of climate change and non-native fish. Aquat. Conserv. Mar. Freshw. Ecosyst. 2022, 32, 781–796. [Google Scholar] [CrossRef]
- Souza, A.T.; Argillier, C.; Blabolil, P.; Děd, V.; Jarić, I.; Monteoliva, A.P.; Reynaud, N.; Ribeiro, F.; Ritterbusch, D.; Sala, P.; et al. Empirical evidence on the effects of climate on the viability of common carp (Cyprinus carpio) populations in European lakes. Biol. Invasions 2022, 24, 1213–1227. [Google Scholar] [CrossRef]
- Tytar, V.; Nekrasova, O.; Pupins, M.; Čeirāns, A.; Skute, A. Modelling the range expansion of pumpkinseed Lepomis gibbosus across Europe, with special focus on Latvia and Ukraine. North West J. Zool. 2021, 18, 143–150. [Google Scholar]
- Radtke, G.; Wolnicki, J.; Kapusta, A.; Przybylski, M.; Kaczkowski, Z. Critical thermal maxima of three small-bodied fish species (Cypriniformes) of different origin and protection status. Eur. Zool. J. 2022, 89, 1351–1361. [Google Scholar] [CrossRef]
- Mumladze, L.; Kuljanishvili, T.; Japoshvili, B.; Epitashvili, G.; Kalous, L.; Vilizzi, L.; Piria, M. Risk of invasiveness of non-native fishes in the South Caucasus biodiversity and geopolitical hotspot. NeoBiota 2022, 76, 109–133. [Google Scholar] [CrossRef]
- Tarkan, A.S.; Emiroğlu, Ö.; Aksu, S.A.D.İ.; Başkurt, S.; Aksu, İ.; Vilizzi, L.; Yoğurtçuoğlu, B. Coupling molecular and risk analysis to investigate the origin, distribution and potential impact of non-native species: An application to ruffe Gymnocephalus cernua in Turkey. Eur. Zool. J. 2022, 89, 109–121. [Google Scholar] [CrossRef]
- Jourdan, J.; Riesch, R.; Cunze, S. Off to new shores: Climate niche expansion in invasive mosquitofish (Gambusia spp.). Ecol. Evol. 2021, 11, 18369–18400. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, J.S.; Fraker, M.E.; Hood, J.M.; Frank, K.T.; DuFour, M.R.; Gorman, A.M.; Ludsin, S.A. Functional traits reveal the dominant drivers of long-term community change across a North American Great Lake. Glob. Chang. Biol. 2021, 27, 6232–6251. [Google Scholar] [CrossRef]
- Lin, L.; Deng, W.; Huang, X.; Liu, Y.; Huang, L.; Kang, B. How fish traits and functional diversity respond to environmental changes and species invasion in the largest river in Southeastern China. PeerJ 2021, 9, e11824. [Google Scholar] [CrossRef]
- Pirroni, S.; de Pennafort Dezen, L.; Santi, F.; Riesch, R. Comparative gut content analysis of invasive mosquitofish from Italy and Spain. Ecol. Evol. 2021, 11, 4379–4398. [Google Scholar] [CrossRef]
- Costa, A.C.; Balibrea, A.; Raposeiro, P.M.; Santos, S.; Souto, M.; Gonçalves, V. Non-indigenous and invasive freshwater species on the Atlantic Islands of the Azores Archipelago. Front. Ecol. Evol. 2021, 9, 631214. [Google Scholar] [CrossRef]
- Carosi, A.; Ghetti, L.; Lorenzoni, M. The role of climate changes in the spread of freshwater fishes: Implications for alien cool and warm-water species in a Mediterranean basin. Water 2021, 13, 347. [Google Scholar] [CrossRef]
- Bounas, A.; Catsadorakis, G.; Koutseri, I.; Nikolaou, H.; Nicolas, D.; Malakou, M.; Crivelli, A.J. Temporal trends and determinants of fish biomass in two contrasting natural lake systems: Insights from a spring long-term monitoring scheme. Knowl. Manag. Aquat. Ecosyst. 2021, 422, 28. [Google Scholar] [CrossRef]
- Rubenson, E.S.; Olden, J.D. An invader in salmonid rearing habitat: Current and future distributions of smallmouth bass (Micropterus dolomieu) in the Columbia River Basin. Can. J. Fish. Aquat. Sci. 2020, 77, 314–325. [Google Scholar] [CrossRef]
- Zięba, G.; Vilizzi, L.; Copp, G.H. How likely is Lepomis gibbosus to become invasive in Poland under conditions of climate warming? Acta Ichthyol. Piscat. 2020, 50, 35–51. [Google Scholar] [CrossRef]
- Liu, C.; Comte, L.; Xian, W.; Chen, Y.; Olden, J.D. Current and projected future risks of freshwater fish invasions in China. Ecography 2019, 42, 2074–2083. [Google Scholar] [CrossRef]
- Jia, Y.; Kennard, M.J.; Liu, Y.; Sui, X.; Chen, Y.; Li, K.; Wang, G.; Chen, Y. Understanding invasion success of Pseudorasbora parva in the Qinghai-Tibetan Plateau: Insights from life-history and environmental filters. Sci. Total Environ. 2019, 694, 133739. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Liu, Y.; Chen, K.; Sun, H.; Chen, Y. Climate, habitat and human disturbance driving the variation of life-history traits of the invasive goldfish Carassius auratus (Linnaeus, 1758) in a Tibetan Plateau river. Aquat. Invasions 2019, 14, 724–737. [Google Scholar] [CrossRef]
- Radinger, J.; Alcaraz-Hernández, J.D.; García-Berthou, E. Environmental filtering governs the spatial distribution of alien fishes in a large, human-impacted Mediterranean river. Divers. Distrib. 2019, 25, 701–714. [Google Scholar] [CrossRef]
- Shelton, J.M.; Weyl, O.L.; Esler, K.J.; Paxton, B.R.; Impson, N.D.; Dallas, H.F. Temperature mediates the impact of non-native rainbow trout on native freshwater fishes in South Africa’s Cape Fold Ecoregion. Biol. Invasions 2018, 20, 2927–2944. [Google Scholar] [CrossRef]
- Rubenson, E.S.; Olden, J.D. Dynamism in the upstream invasion edge of a freshwater fish exposes range boundary constraints. Oecologia 2017, 184, 453–467. [Google Scholar] [CrossRef]
- Rolls, R.J.; Hayden, B.; Kahilainen, K.K. Conceptualising the interactive effects of climate change and biological invasions on subarctic freshwater fish. Ecol. Evol. 2017, 7, 4109–4128. [Google Scholar] [CrossRef]
- Tarkan, A.S.; Sarı, H.M.; İlhan, A.; Kurtul, I.; Vilizzi, L. Risk screening of non-native and translocated freshwater fish species in a Mediterranean-type shallow lake: Lake Marmara (West Anatolia). Zool. Middle East 2017, 63, 48–57. [Google Scholar] [CrossRef]
- Copp, G.H.; Britton, J.R.; Guo, Z.; Ronni Edmonds-Brown, V.; Pegg, J.; Vilizzi, L.; Davison, P.I. Trophic consequences of non-native pumpkinseed Lepomis gibbosus for native pond fishes. Biol. Invasions 2017, 19, 25–41. [Google Scholar] [CrossRef]
- Crook, D.A.; Lowe, W.H.; Allendorf, F.W.; Erős, T.; Finn, D.S.; Gillanders, B.M.; Hadwen, W.L.; Harrod, C.; Hermoso, V.; Jennings, S.; et al. Human effects on ecological connectivity in aquatic ecosystems: Integrating scientific approaches to support management and mitigation. Sci. Total Environ. 2015, 534, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Rabitsch, W.; Milasowszky, N.; Nehring, S.; Wiesner, C.; Wolter, C.; Essl, F. The times are changing: Temporal shifts in patterns of fish invasions in central European fresh waters. J. Fish Biol. 2013, 82, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Kornis, M.S.; Mercado-Silva, N.; Vander Zanden, M.J. Twenty years of invasion: A review of round goby Neogobius melanostomus biology, spread and ecological implications. J. Fish Biol. 2012, 80, 235–285. [Google Scholar] [CrossRef] [PubMed]
- Britton, J.R.; Cucherousset, J.; Davies, G.D.; Godard, M.J.; Copp, G.H. Non-native fishes and climate change: Predicting species responses to warming temperatures in a temperate region. Freshw. Biol. 2010, 55, 1130–1141. [Google Scholar] [CrossRef]
- Benejam, L.; Alcaraz, C.; Sasal, P.; Simon-Levert, G.; García-Berthou, E. Life history and parasites of the invasive mosquitofish (Gambusia holbrooki) along a latitudinal gradient. Biol. Invasions 2009, 11, 2265–2277. [Google Scholar] [CrossRef]
- Rahel, F.J.; Olden, J.D. Assessing the effects of climate change on aquatic invasive species. Conserv. Biol. 2008, 22, 521–533. [Google Scholar] [CrossRef]
- Franchi, E.; Carosi, A.; Ghetti, L.; Giannetto, D.; Pedicillo, G.; Pompei, L.; Lorenzoni, M. Changes in the fish community of the upper Tiber River after construction of a hydro-dam. J. Limnol. 2014, 73, 876. [Google Scholar] [CrossRef]
- Drouineau, H.; Carter, C.; Rambonilaza, M.; Beaufaron, G.; Bouleau, G.; Gassiat, A.; Lambert, P.; le Floch, S.; Tétard, S.; De Oliveira, E. River continuity restoration and diadromous fishes: Much more than an ecological issue. Environ. Manag. 2018, 61, 671–686. [Google Scholar] [CrossRef]
- Merg, M.L.; Dézerald, O.; Kreutzenberger, K.; Demski, S.; Reyjol, Y.; Usseglio-Polatera, P.; Belliard, J.C. Modeling diadromous fish loss from historical data: Identification of anthropogenic drivers and testing of mitigation scenarios. PLoS ONE 2020, 15, e0236575. [Google Scholar] [CrossRef]
- Carosi, A.; Ghetti, L.; La Porta, G.; Lorenzoni, M. Ecological effects of the European barbel Barbus barbus (L., 1758) (Cyprinidae) invasion on native barbel populations in the Tiber River basin (Italy). Eur. Zool. J. 2017, 84, 420–435. [Google Scholar] [CrossRef]
- Carosi, A.; Ghetti, L.; Forconi, A.; Lorenzoni, M. Fish community of the river Tiber basin (Umbria-Italy): Temporal changes and possible threats to native biodiversity. Knowl. Manag. Aquat. Ecosyst. 2015, 416, 22. [Google Scholar] [CrossRef]
- Isaak, D.J.; Rieman, B.E. Stream isotherm shifts from climate change and implications for distributions of ectothermic organisms. Glob. Chang. Biol. 2013, 19, 742–751. [Google Scholar] [CrossRef] [PubMed]
- Marr, S.M.; Olden, J.D.; Leprieur, F.; Arismendi, I.; Ćaleta, M.; Morgan, D.L.; Nocita, A.; Šanda, R.; Tarkan, A.S.; García-Berthou, E. A global assessment of freshwater fish introductions in mediterranean-climate regions. Hydrobiologia 2013, 719, 317–329. [Google Scholar] [CrossRef]
- Sauz-Sánchez, J.D.J.; Rodiles-Hernández, R.; Andrade-Velázquez, M.; Mendoza-Carranza, M. Modelling the potential distribution of two tropical freshwater fish species under climate change scenarios. Aquat. Conserv. Mar. Freshw. Ecosys. 2021, 31, 2737–2751. [Google Scholar] [CrossRef]
- Nickus, U.; Bishop, K.; Erlandsson, M.; Evans, C.D.; Forsius, M.; Laudon, H.; Livingstone, D.M.; Monteith, D.; Thies, H. Direct impacts of climate change on freshwater ecosystems. In Climate Change Impacts on Freshwater Ecosystems, 1st ed.; Kernan, M., Battarbee, R., Moss, B., Eds.; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2010; pp. 38–64. [Google Scholar] [CrossRef]
- Jeppesen, E.; Kronvang, B.; Jørgensen, T.B.; Larsen, S.E.; Andersen, H.E.; Søndergaard, M.; Liboriussen, L.; Bjerring, R.; Johansson, L.; Trolle, D.; et al. Recent climate-induced changes in freshwaters in Denmark. In Climatic Change and Global Warming of Inland Waters: Impacts and Mitigation for Ecosystems and Societies, 1st ed.; Goldman, C.R., Kumagai, M., Robarts, R.D., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 155–171. [Google Scholar]
- Filipe, A.F.; Lawrence, J.E.; Bonada, N. Vulnerability of stream biota to climate change in mediterranean climate regions: A synthesis of ecological responses and conservation challenges. Hydrobiologia 2013, 719, 331–351. [Google Scholar] [CrossRef]
- Carosi, A.; Padula, R.; Ghetti, L.; Lorenzoni, M. Endemic freshwater fish range shifts related to global climate changes: A long-term study provides some observational evidence for the Mediterranean area. Water 2019, 11, 2349. [Google Scholar] [CrossRef]
- Mulholland, P.J.; Best, G.R.; Coutant, C.C.; Hornberger, G.M.; Meyer, J.L.; Robinson, P.J.; Stenberg, J.R.; Turner, R.E.; Vera-Herrera, F.; Wetzel, R.G. Effects of climate change on freshwater ecosystems of the south-eastern United States and the Gulf Coast of Mexico. Hydrol. Process. 1997, 11, 949–970. [Google Scholar] [CrossRef]
- Danladi Bello, A.A.; Hashim, N.B.; Mohd Haniffah, M.R. Predicting impact of climate change on water temperature and dissolved oxygen in tropical rivers. Climate 2017, 5, 58. [Google Scholar] [CrossRef]
- Copp, G.H.; Russell, I.C.; Peeler, E.J.; Gherardi, F.; Tricarico, E.; MacLeod, A.; Cowx, I.G.; Nunn, A.D.; Occhipinti Ambrogi, A.; Savini, D.; et al. European Non-native Species in Aquaculture Risk Analysis Scheme–a summary of assessment protocols and decision support tools for use of alien species in aquaculture. Fish. Manag. Ecol. 2016, 23, 1–11. [Google Scholar] [CrossRef]
- Buisson, L.; Thuiller, W.; Lek, S.; Lim, P.U.Y.; Grenouillet, G. Climate change hastens the turnover of stream fish assemblages. Glob. Chang. Biol. 2008, 14, 2232–2248. [Google Scholar] [CrossRef]
- Fobert, E.; Fox, M.G.; Ridgway, M.; Copp, G.H. Heated competition: How climate change will affect non-native pumpkinseed Lepomis gibbosus and native perch Perca fluviatilis interactions in the UK. J. Fish Biol. 2011, 79, 1592–1607. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Climatic Area | Geographic Area | Reference | Target Species | Methodological Approach |
|---|---|---|---|---|
| Tropical wet/semiarid | Central Mexico | Gomez-Maldonado and Camacho-Cervantes (2022) [42] | Skiffia bilineata (Bean, 1887) Poecilia reticulata Peters, 1859 | Behavioral observations in aquarium |
| Temperate | Germany | DeWeber et al. (2022) [43] | Coregonus wartmanni (Bloch, 1784) | Bioenergetics and growth modeling |
| Continental | North America | Avlijaš et al. (2022) [44] | Tinca tinca (L., 1758) | Growth and functional response experiments |
| Tropical | Philippines | To et al. (2022) [45] | Clarias batrachus (L., 1758) | Aquatic Species Invasiveness Screening kit (AS-ISK) |
| Mediterranean | South Africa | Cerrilla et al. (2022) [46] | Labeo seeberi Gilchrist & Thompson, 1911 | Observational data analysis |
| Continental, Temperate oceanic, Mediterranean | Europe | Souza et al. (2022) [47] | Cyprinus carpio L., 1758 | Observational data analysis |
| Temperate | Ukraine, Latvia | Tytar et al. (2022) [48] | Lepomis gibbosus (L., 1758) | Ecological niches models |
| Temperate | Poland | Radtke et al. (2022) [49] | Eupallasella percnurus (Pallas, 1814), Leucaspius delineates (Heckel, 1843), Pseudorasbora parva (Temminck & Schlegel, 1846) | Critical Thermal Method in aquarium |
| Continental mesophilic | South Caucasus | Mumladze et al. (2022) [50] | 32 alien species | AS-ISK |
| Temperate, Mediterranean | Turkey | Tarkan et al. (2022) [51] | Gymnocephalus cernua (L., 1758) | AS-ISK, mtDNA barcoding |
| - | Global | Jourdan et al. (2021) [52] | Gambusia spp. | Species distribution models |
| Humid continental | North America | Sinclair et al. (2021) [53] | Lake Eire fish community | Functional trait compositional changes |
| Humid subtropical, Oceanic | Southeastern China | Lin et al. (2021) [54] | Min River fish community | Functional diversity changes |
| Mediterranean | Italy, Spain | Pirroni et al. (2021) [55] | Gambusia holbrooki Girard, 1859 | Feeding ecology |
| Humid subtropical, Mediterranean | Azores Archipelago | Costa et al. (2021) [56] | 132 alien freshwater species | Literature review |
| Mediterranean | Italy | Carosi et al. (2021) [57] | Barbus barbus(L., 1758), Gobio gobio (L., 1758), Padogobius bonelli (Bonaparte, 1846), P. parva | Observational data analysis |
| Temperate, Mediterranean | Southeastern Europe | Bounas et al. (2021) [58] | Prespa Lakes fish community | Structural change models, random forest |
| Continental, Temperate oceanic | North America | Rubenson and Olden (2020) [59] | Micropterus dolomieu Lacepède, 1802 | Species distribution models, environmental DNA |
| Temperate | UK, France, Poland | Zięba et al. (2020) [60] | L. gibbosus | AS-ISK |
| Tropical, Subarctic, Alpine, Temperate, Arid | China | Liu et al. (2019) [61] | Freshwater fish | Species distribution model |
| Alpine | Tibetan Plateau | Jia et al. (2019) [62] | P. parva | Linear mixed-effects models, random forest |
| Alpine | Tibetan Plateau | Jia et al. (2019) [63] | Carassius auratus (L., 1758) | Multiple regression, hierarchical partitioning models |
| Alpine, Semiarid, Mediterranean | Iberian peninsula | Radinger et al. (2019) [64] | Freshwater fish | Boosted regression tree, species distribution model |
| Mediterranean | South Africa | Shelton et al. (2018) [65] | Oncorhynchus mykiss (Walbaum, 1792) | Distance-based Linear Models |
| Continental, Temperate oceanic | North America | Rubenson and Olden (2017) [66] | M. dolomieu | Telemetry analysis |
| Subpolar | Subarctic regions | Rolls et al. (2017) [67] | Freshwater fish | Literature review |
| Mediterranean | West Anatolia | Tarkan et al. (2017) [68] | 35 alien species in Marmara Lake | AS-ISK |
| Temperate | UK | Copp et al. (2017) [69] | L.gibbosus | Stable isotope analysis |
| - | Global | Crook et al. (2015) [70] | Freshwater animals | Literature review |
| Temperate, Continental | Germany, Austria | Rabitsch et al. (2013) [71] | 15 alien species | Species spatial distribution |
| Temperate, Continental | North America, Eurasia, Baltic Sea | Kornis et al. (2012) [72] | Neogobius melanostomus (Pallas, 1814) | Literature review |
| Temperate | UK | Britton et al. (2010) [73] | C. carpio, Silurus glanis L., 1758 | Climate models, spatial distribution models |
| Mediterranean | France, Spain | Benejam et al. (2009) [74] | G. holbrooki | Observational data analysis |
| - | Global | Rahel and Olden (2008) [75] | Alien species | Literature review |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carosi, A.; Lorenzoni, F.; Lorenzoni, M. Synergistic Effects of Climate Change and Alien Fish Invasions in Freshwater Ecosystems: A Review. Fishes 2023, 8, 486. https://doi.org/10.3390/fishes8100486
Carosi A, Lorenzoni F, Lorenzoni M. Synergistic Effects of Climate Change and Alien Fish Invasions in Freshwater Ecosystems: A Review. Fishes. 2023; 8(10):486. https://doi.org/10.3390/fishes8100486
Chicago/Turabian StyleCarosi, Antonella, Francesca Lorenzoni, and Massimo Lorenzoni. 2023. "Synergistic Effects of Climate Change and Alien Fish Invasions in Freshwater Ecosystems: A Review" Fishes 8, no. 10: 486. https://doi.org/10.3390/fishes8100486
APA StyleCarosi, A., Lorenzoni, F., & Lorenzoni, M. (2023). Synergistic Effects of Climate Change and Alien Fish Invasions in Freshwater Ecosystems: A Review. Fishes, 8(10), 486. https://doi.org/10.3390/fishes8100486