

Evidence for the Growth Superiority and Delayed Ovarian Development in Tetraploid Loach Misgurnus anguillicaudatus

Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Fish Materials and Sampling
2.3. Gonadal-Somatic Index and Gonadal Histology
2.4. RNA Isolation and Sequencing
2.5. De Novo Assembly and Annotation
2.6. Identification of Differentially Expressed Genes
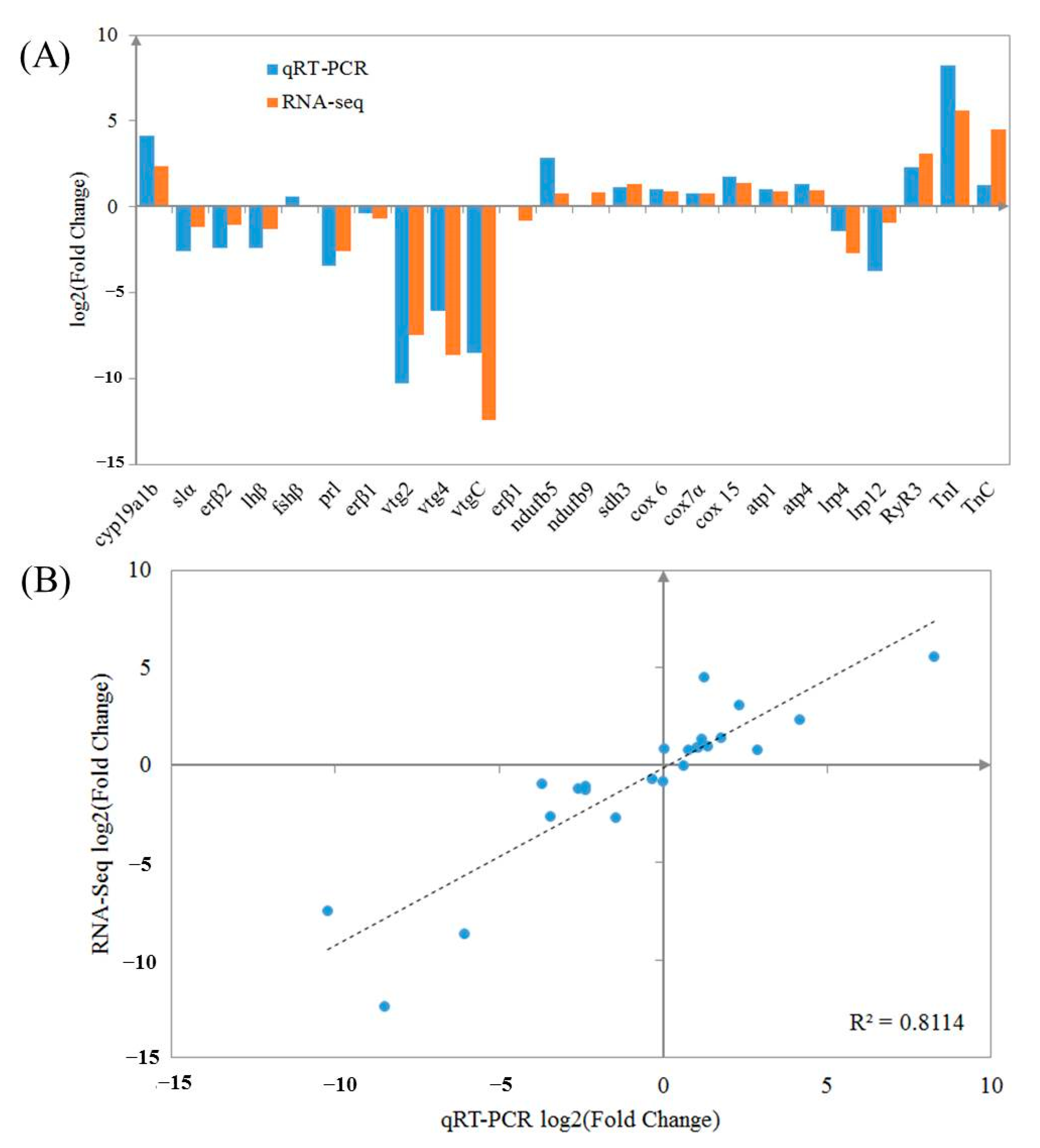
2.7. Validation of RNA-Seq Data via qRT-PCR Assay
2.8. Statistical Analysis
3. Results
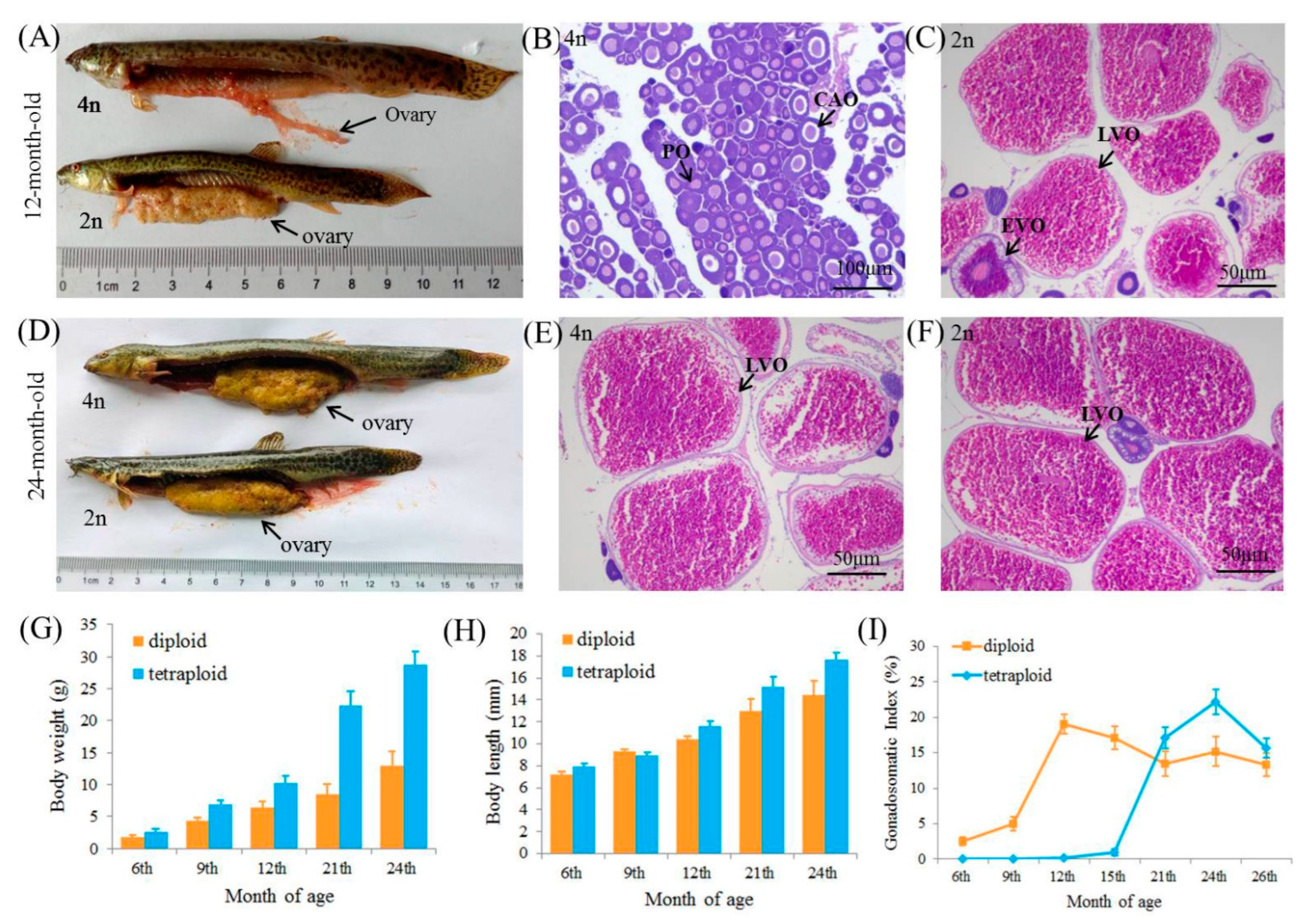
3.1. Growth and Ovarian Development of Diploids and Tetraploids
3.2. Assembly and Annotation of Transcripts
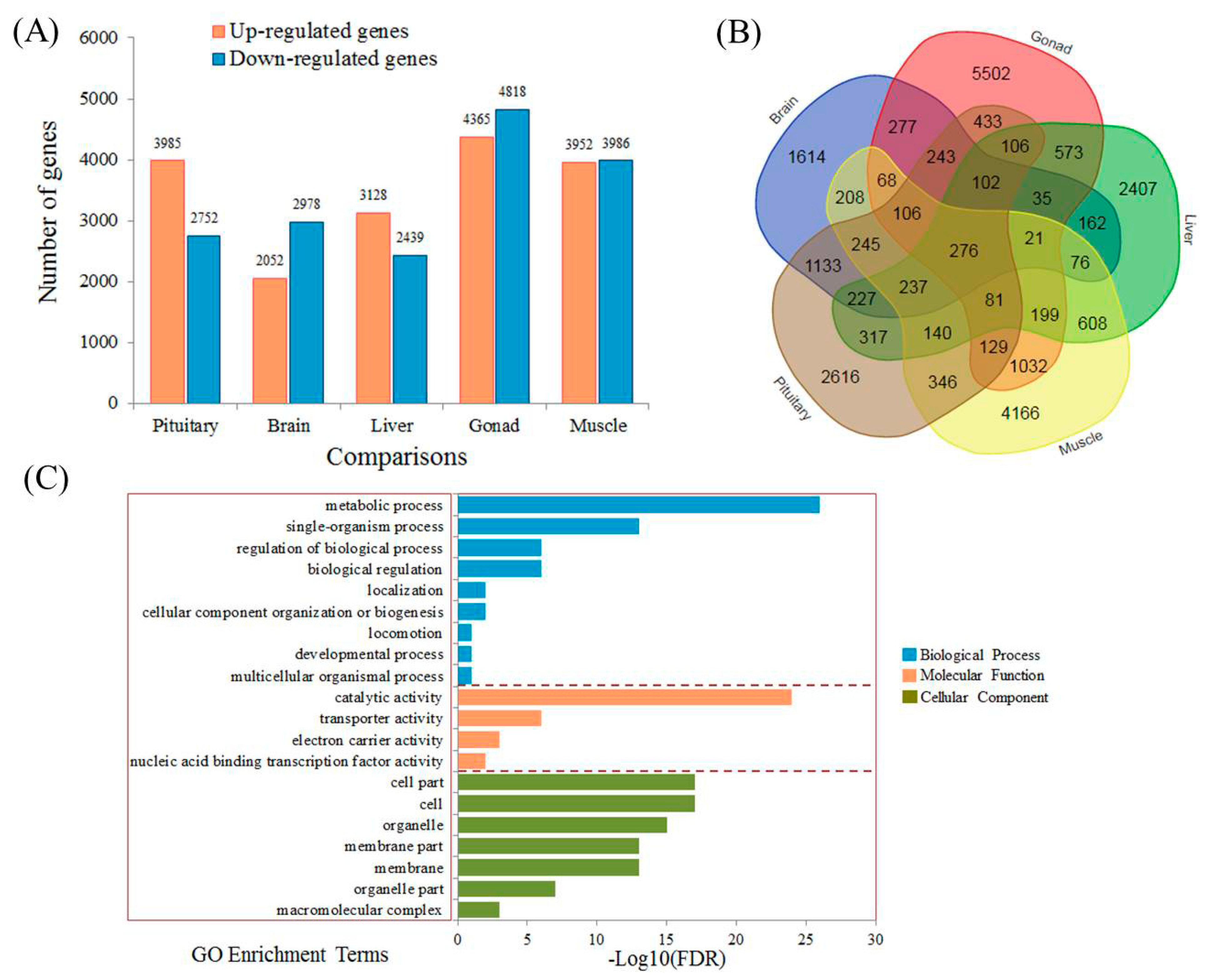
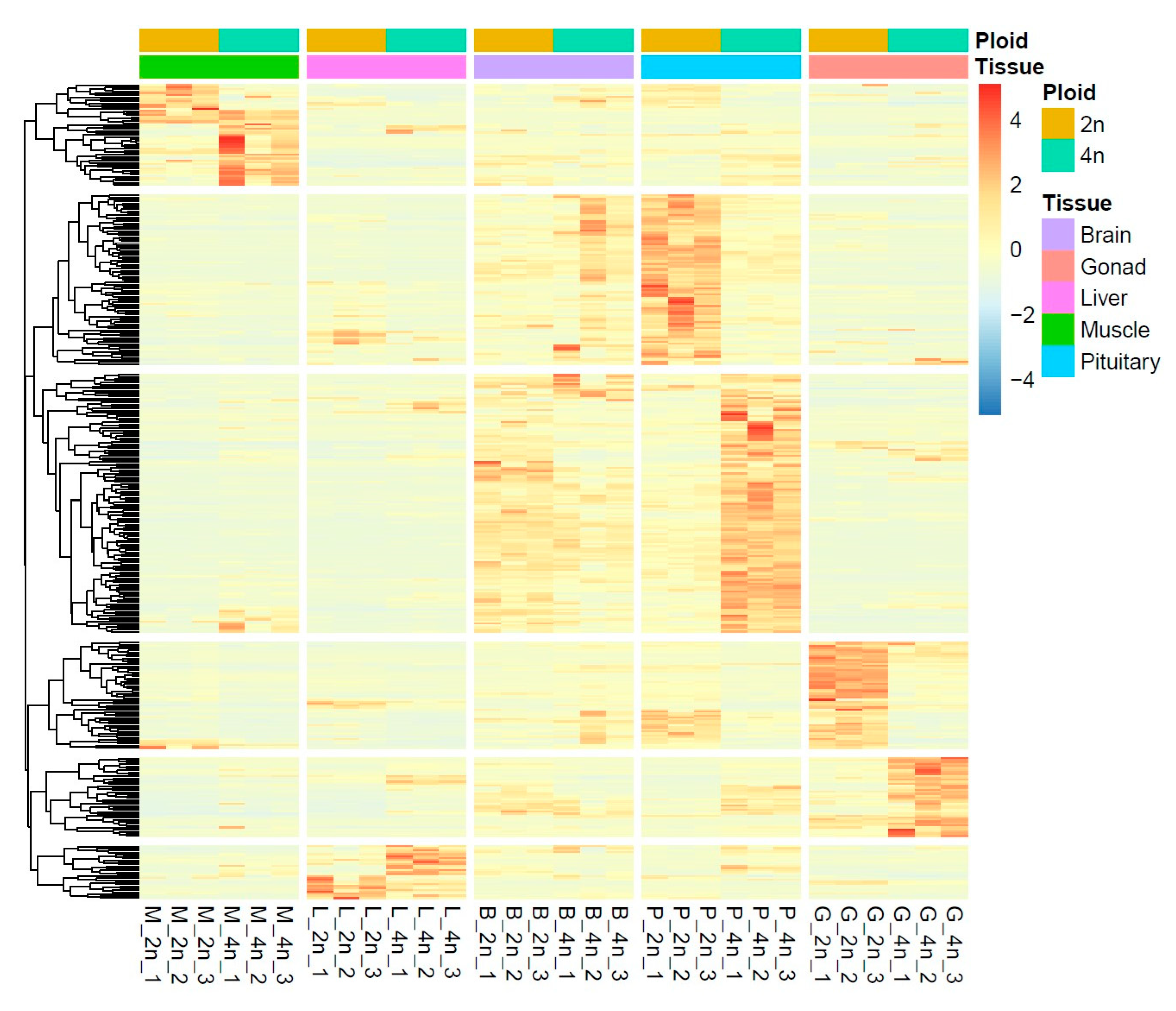
3.3. Identification of Differentially Expressed Genes
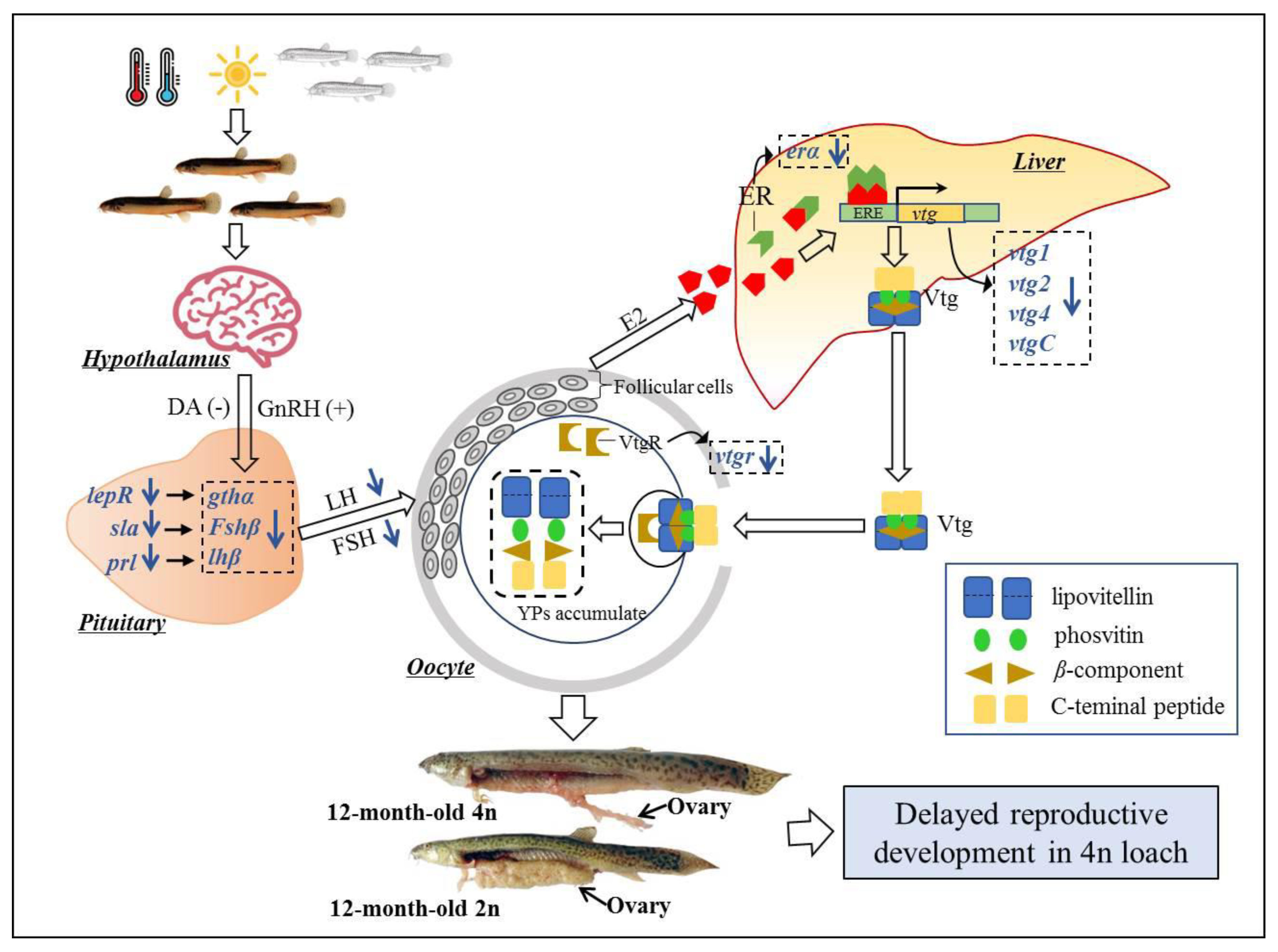
3.4. Candidate Genes Mediating Delayed Ovarian Development of Tetraploids
3.5. Genetic Regulation Patterns of Growth Superiority in Tetraploids
3.6. Correlation of Ovarian Development and Growth in Tetraploids and Diploids
3.7. qRT-PCR Validation of DEGs
4. Discussion
4.1. Up-Regulation of Some Key GH/IGF-Related Genes in Tetraploids
4.2. Genetic Difference of Ovarian Development between Diploids and Tetraploids
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Le Comber, S.C.; Smith, C. Polyploidy in fishes: Patterns and processes. Biol. J. Linn. Soc. 2004, 82, 431–442. [Google Scholar] [CrossRef]
- Benfey, T.J. The physiology of triploid salmonids in relation to aquaculture. Can. Rep. Fish. Aqua. Sci. 1991, 1789, 73–80. [Google Scholar]
- Benfey, T.J. The physiology and behavior of triploid fishes. Rev. Fish. Sci. 1999, 7, 39–67. [Google Scholar] [CrossRef]
- Leggatt, A.L.; Iwama, G.K. Occurrence of polyploidy in the fishes. Rev. Fish Biol. Fisher. 2003, 13, 237–246. [Google Scholar] [CrossRef]
- Malison, J.A.; Procarione, L.S.; Held, J.A.; Kayes, T.B.; Amundson, C.H. The influence of triploidy and heat and hydrostatic pressure shocks on the growth and reproductive development of juvenile yellow perch (Perca flavescens). Aquaculture 1993, 116, 121–133. [Google Scholar] [CrossRef]
- Felip, A.; Piferrer, F.; Zanuy, S.; Carrillo, M. Comparative growth performance of diploid and triploid european sea bass over the first four spawning seasons. J. Fish Biol. 2001, 58, 76–88. [Google Scholar] [CrossRef]
- Yamazaki, F.; Goodier, J. Cytogenetic effects of hydrostatic pressure treatment to suppress the first cleavage of salmon embryos. Aquaculture 1993, 110, 51–59. [Google Scholar] [CrossRef]
- Rab, P.; Machordom, A.; Perdices, A.; Guegan, J.F. Karyotypes of three “small” Barbus species (Cyprinidae) from Republic of Guinea (Western Africa) with a review on karyology of african small Barbus. Caryologia 1995, 48, 299–307. [Google Scholar] [CrossRef]
- Golubtsov, A.S.; Krysanov, E.Y. Karyological study of some cyprinid species from ethiopia-the ploidy differences between large and small Barbus of Africa. J. Fish Biol. 1993, 42, 445–455. [Google Scholar] [CrossRef]
- Sezaki, K.; Kobayasi, H. Comparison of erythrocytic size between diploid and tetraploid in spinous loach, Cobitis Biwae. Nippon Suisan Gakk. 1978, 44, 851–854. [Google Scholar] [CrossRef]
- Kitagawa, T.; Watanabe, M.; Kitagawa, E.; Yoshioka, M.; Kashiwagi, M.; Okazaki, T. Phylogeography and the maternal origin of the tetraploid form of the Japanese spined loach, Cobitis biwae, revealed by mitochondrial DNA analysis. Ichthyol. Res. 2003, 50, 318–325. [Google Scholar] [CrossRef]
- Qin, C.G.; Huang, K.X.; Xu, H.B. Protective effect of polysaccharide from the loach on the in vitro and in vivo peroxidative damage of hepatocyte. J. Nutr. Biochem. 2002, 13, 592–597. [Google Scholar] [CrossRef]
- Yi, S.; Zhou, X.; Zhang, M.; Luo, S. Full-length transcriptome of Misgurnus anguillicaudatus provides insights into evolution of genus Misgurnus. Sci. Rep. 2018, 8, 11699. [Google Scholar] [CrossRef]
- Zhang, Q.; Arai, K. Distribution and reproductive capacity of natural triploid individuals and occurrence of unreduced eggs as a cause of polyploidization in the loach, Migurnus anguillicaudatus. Ichthyol. Res. 1999, 46, 153–161. [Google Scholar] [CrossRef]
- Morishima, K.; Horie, S.; Yamaha, E.; Arai, K. A cryptic clonal line of the loach Misgurnus Anguillicaudatus (Teleostei: Cobitidae) evidenced by induced gynogenesis, interspecific hybridization, microsatellite genotyping and multilocus DNA fingerprinting. Zool. Sci. 2002, 19, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Arai, K. Genetics of the loach, Misgurnus anguillicaudatus, recent progress and perspective. Folia Biol. 2003, 51, 107–117. [Google Scholar]
- Li, Y.; Yu, Z.; Zhang, M.; Qian, C.; Abe, S.; Arai, K. The origin of natural tetraploid loach Misgurnus anguillicaudatus (Teleostei: Cobitidae) inferred from meiotic chromosome configurations. Genetica 2011, 139, 805–811. [Google Scholar] [CrossRef]
- Yu, X.J.; Zhou, T.; Li, Y.C.; Li, K.; Zhou, M. Chromsomes of Chinese Freshwater Fishes; Science Press: Beijing, China, 1989. (In Chinese) [Google Scholar]
- Yin, J.; Zhao, Z.S.; Chen, X.Q.; Li, Y.Q.; Zhu, L.Y. Karyotype comparison of diploid and tetraploid loach, Misgurnus anguillicanudatus. Acta Hydrob. Sin. 2005, 29, 469–472. (In Chinese) [Google Scholar]
- Li, Y.; Yin, J.; Wang, J.; Yuan, X.; Wei, J.; Sun, X.; Arai, K. A study on the distribution of polyploid loach in China. Nippon Suisan Gakk. 2008, 74, 177–182. [Google Scholar] [CrossRef]
- Li, K.; Li, Y.; Zhou, D. A comparative study of the karyotypes in two species of mud loaches. Zool. Res. 1983, 4, 75–81. [Google Scholar]
- Arai, K.; Fujimoto, T. Genomic constitution and atypical reproduction in polyploid and unisexual lineages of the Misgurnus loach, a teleost fish. Cytogenet. Genome Res. 2013, 140, 226–240. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Soonjin, V.Y.; Li, R.; Zhou, X. Comparison of age and growth performance of diploid and tetraploid loach Misgurnus anguillicaudatus, in the Yangtze River basin, China. Environ. Biol. Fish. 2017, 100 (Suppl. 1), 815–828. [Google Scholar] [CrossRef]
- Zhou, X.; Abbas, K.; Li, M.; Fang, L.; Li, S.; Wang, W. Comparative studies on survival and growth performance among diploid, triploid and tetraploid dojo loach Misgurnus anguillicaudatus. Aquac. Int. 2010, 18, 349–359. [Google Scholar] [CrossRef]
- Li, Y.J.; Zhang, M.Z.; Qian, C.; Gao, M.; Arai, K. Fertility and ploidy of gametes of diploid, triploid and tetraploid loaches, Misgurnus anguillicaudatus, in China. J. Appl. Ichthyol. 2012, 28, 900–905. [Google Scholar] [CrossRef]
- Reinecke, M. Influences of the environment on the endocrine and paracrine fish growth hormone-insulin-like growth factor-I system. J. Fish Biol. 2010, 76, 1233–1254. [Google Scholar] [CrossRef]
- Fu, B.; Wang, X.; Feng, X.; Yu, X.; Tong, J. Comparative transcriptomic analyses of two bighead carp (Hypophthalmichthys nobilis) groups with different growth rates. Comp. Biochem. Phys. D 2016, 20, 111–117. [Google Scholar] [CrossRef]
- Reinecke, M.; Björnsson, B.T.; Dickhoff, W.W.; McCormick, S.D.; Navarro, I.; Power, D.M.; Gutiérrez, J. Growth hormone and insulin-like growth factors in fish: Where we are and where to go. Gen. Comp. Endocr. 2005, 142, 20–24. [Google Scholar] [CrossRef]
- Escalante-Rojas, M.; Peña, E.; Hernández, C.; Llera-Herrera, R.; Garcia-Gasca, A. De novo transcriptome assembly for the rose spotted snapper Lutjanus guttatus and expression analysis of growth/atrophy-related genes. Aquac. Res. 2018, 49, 1709–1722. [Google Scholar] [CrossRef]
- Kopchick, J.J.; Andry, J.M. Growth Hormone (GH), GH Receptor, and Signal Transduction. Mol. Genet. Metab. 2000, 71, 293–314. [Google Scholar] [CrossRef]
- Butler, A.A.; LeRoith, D. Minireview: Tissue-specific versus generalized gene targeting of the igf1 and igf1r genes and their roles in insulin-like growth factor physiology. Endocrinology 2001, 142, 1685–1688. [Google Scholar] [CrossRef]
- Zhong, H.; Zhou, Y.; Liu, S.; Tao, M.; Long, Y.; Liu, Z.; Zhang, C.; Duan, W.; Hu, J.; Song, C.; et al. Elevated expressions of GH/IGF axis genes in triploid crucian carp. Gen. Comp. Endocrinol. 2012, 178, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.Y.; Li, M.Y.; Abbas, K.; Gao, Z.X.; Wang, W.M. Comparison of ploidy level screening methods in chinese dojo loach (Misgurnus anguillicaudatus). J. Appl. Ichthyol. 2008, 24, 664–669. [Google Scholar] [CrossRef]
- Domagała, J.; Kirczuk, L.; Pilecka-Rapacz, M. Annual development cycle of gonads of Eurasian ruffe (Gymnocephalus cernuus L.) females from lower Odra River sections differing in the influence of cooling water. J. Fresh. Ecol. 2013, 28, 423–437. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644. [Google Scholar] [CrossRef] [PubMed]
- Pertea, G.; Huang, X.Q.; Liang, F.; Antonescu, V.; Sultana, R.; Karamycheva, S.; Lee, Y.; White, J.; Cheung, F.; Parvizi, B.; et al. TIGR Gene Indices clustering tools (TGICL): A software system for fast clustering of large est datasets. Bioinformatics 2003, 19, 651–652. [Google Scholar] [CrossRef]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. Busco: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Med. 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-seq Data with or without a reference genome. BMC Bioinform. 2011, 12, 93–99. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 392, 316–322. [Google Scholar] [CrossRef] [PubMed]
- De-Santis, C.; Jerry, D.R. Candidate growth genes in finfish-where should we be looking? Aquaculture 2007, 272, 22–38. [Google Scholar] [CrossRef]
- Sun, Y.; Huang, Y.; Hu, G.; Zhang, X.; Ruan, Z.; Zhao, X.; Guo, C.Y.; Tang, Z.J.; Li, X.F.; You, X.X.; et al. Comparative transcriptomic study of muscle provides new insights into the growth superiority of a novel grouper hybrid. PLoS ONE 2016, 11, e0168802. [Google Scholar] [CrossRef]
- Rescan, P.Y.; Montfort, J.; Ralliere, C.; Le Cam, A.; Esquerre, D.; Hugot, K. Dynamic gene expression in fish muscle during recovery growth induced by a fasting-refeeding schedule. BMC Genom. 2007, 8, 438. [Google Scholar] [CrossRef]
- Duan, C. The insulin-like growth factor system and its biological actions in fish. Integr. Comp. Biol. 1997, 37, 491. [Google Scholar] [CrossRef]
- Duan, C. Nutritional and developmental regulation of insulin-like growth factors in fish. J. Nutr. 1998, 128, 306S. [Google Scholar] [CrossRef]
- Reindl, K.M.; Kittilson, J.D.; Bergan, H.E.; Sheridan, M.A. Growth hormone-stimulated insulin-like growth factor-1 expression in rainbow trout (Oncorhynchus mykiss) hepatocytes is mediated by ERK, PI3K-AKT, and JAK-STAT. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R236–R243. [Google Scholar] [CrossRef]
- Pierce, A.L.; Dickey, J.T.; Larsen, D.A.; Fukada, H.; Swanson, P.; Dickhoff, W.W. A quantitative real-time RT-PCR assay for salmon IGF-I mRNA, and its application in the study of GH regulation of IGF-I gene expression in primary culture of salmon hepatocytes. Gen. Comp. Endocrinol. 2004, 135, 401–411. [Google Scholar] [CrossRef]
- Glass, D.J. Skeletal muscle hypertrophy and atrophy signaling pathways. Int.J. Biochem. Cell Biol. 2005, 37, 1974–1984. [Google Scholar] [CrossRef]
- Fuentes, E.N.; Björnsson, B.T.; Valdés, J.A.; Einarsdottir, I.E.; Lorca, B.; Alvarez, M.; Molina, A. IGF-I/PI3K/Akt and IGF-I/MAPK/ERK pathways in vivo in skeletal muscle are regulated by nutrition and contribute to somatic growth in the fine flounder. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R1532–R1542. [Google Scholar] [CrossRef] [PubMed]
- Cleveland, B.M.; Weber, G.M. Effects of insulin-like growth factor-I, insulin, and leucine on protein turnover and ubiquitin ligase expression in rainbow trout primary myocytes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R341. [Google Scholar] [CrossRef] [PubMed]
- Niu, D.; Wang, L.; Bai, Z.; Xie, S.; Zhao, H.; Li, J. Identification and expression characterization of the myostatin (MSTN) gene and association analysis with growth traits in the razor clam Sinonovacula constricta. Gene 2015, 555, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Tostivint, H.; Lihrmann, I.; Vaudry, H. New insight into the molecular evolution of the somatostatin family. Mol. Cell Endocrinol. 2008, 286, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Volkoff, H.; Eykelbosh, A.J.; Peter, R.E. Role of leptin in the control of feeding of goldfish Carassius auratus: Interactions with cholecystokinin, neuropeptide Y and orexin A, and modulation by fasting. Brain Res. 2003, 972, 90–109. [Google Scholar] [CrossRef]
- Londraville, R.L.; Duvall, C.S. Murine leptin injections increase intracellular fatty acid-binding protein in green sunfish (Lepomis cyanellus). Gen. Comp. Endocrinol. 2002, 129, 56–62. [Google Scholar] [CrossRef]
- Trombley, S.; Schmitz, M. Leptin in fish: Possible role in sexual maturation in male Atlantic salmon. Fish Physiol. Biochem. 2013, 39, 103–106. [Google Scholar] [CrossRef]
- Chen, J.; Cao, M.; Zhang, A.; Shi, M.; Tao, B.; Li, Y.; Wang, Y.; Zhu, Z.; Trudeau, V.L.; Hu, W. Growth hormone overexpression disrupts reproductive status through actions on leptin. Front. Endocrinol. 2018, 27, 131. [Google Scholar] [CrossRef]
- Levavi-Sivan, B.; Bogerd, J.; Mañanós, E.L.; Gómez, A.; Lareyre, J.J. Perspectives on fish gonadotropins and their receptors. Gen. Comp. Endocrinol. 2010, 165, 412–437. [Google Scholar] [CrossRef]
- Ma, X.; Liu, X.; Zhang, Y.; Zhu, P.; Ye, W.; Lin, H. Two growth hormone receptors in Nile tilapia (Oreochromis niloticus): Molecular characterization, tissue distribution and expression profiles in the gonad during the reproductive cycle. Comp. Biochem. Physiol. B 2007, 147, 325–339. [Google Scholar] [CrossRef]
- Luo, W.; Liu, C.; Cao, X.; Huang, S.; Wang, W.; Wang, Y. Transcriptome profile analysis of ovarian tissues from diploid and tetraploid loaches Misgurnus anguillicaudatus. Int. J. Mol. Sci. 2015, 16, 16017–16033. [Google Scholar] [CrossRef] [PubMed]
- Raven, P.A.; Sakhrani, D.; Beckman, B.; Neregard, L.; Sundström, L.F.; Björnsson, B.T.; Devlin, R.H. Growth and endocrine effects of recombinant bovine growth hormone treatment in non-transgenic and growth hormone transgenic coho salmon. Gen. Comp. Endocrinol. 2012, 177, 143–152. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Unigene ID | Log2FC | Annotation | Gene Name | FDR |
|---|---|---|---|---|
| TRINITY_DN27311_c2_g2 | 2.2012 | insulin-like growth factor I | igf1 | 3.50 × 10−10 |
| TRINITY_DN30348_c3_g1 | 1.7627 | insulin-like growth factor II | Igf2 | 2.53 × 10−10 |
| TRINITY_DN26313_c4_g1 | 2.0406 | growth hormone-releasing hormone receptor | GHRHR | 5.10 × 10−8 |
| TRINITY_DN30415_c0_g2 | 1.1232 | growth hormone regulated TBC protein 1 | GRTP1 | 3.94 × 10−16 |
| TRINITY_DN38250_c0_g3 | 1.4433 | insulin-like growth factor-binding protein 7 | Igfbp7 | 1.07 × 10−28 |
| TRINITY_DN24955_c0_g1 | 1.3327 | insulin-like growth factor-binding protein 10 | Igfbp10 | 2.28 × 10−12 |
| TRINITY_DN25644_c0_g1 | 2.7081 | elongation factor 1-α | ef1α | 8.63 × 10−4 |
| TRINITY_DN40659_c0_g1 | 2.2222 | transforming growth factor beta 1 | TGF-β1 | 9.20 × 10−15 |
| TRINITY_DN31880_c1_g1 | 2.6882 | transforming growth factor beta-3 | TGF-β3 | 3.45 × 10−109 |
| TRINITY_DN48440_c2_g3 | −1.2520 | luteinizing hormone-β | lhβ | 1.10 × 10−234 |
| TRINITY_DN20712_c3_g3 | −2.9352 | follicle-stimulating hormone-β | fshβ | 0 |
| TRINITY_DN43724_c0_g4 | −1.0389 | brain aromatase | cyp19a1b | 4.04 × 10−19 |
| TRINITY_DN33263_c0_g1 | −2.2631 | prolactin | prl | 0 |
| TRINITY_DN40445_c1_g4 | −1.8500 | somatolactin alpha | slα | 0 |
| TRINITY_DN23324_c0_g2 | −4.4445 | leptin | leptin | 8.32 × 10−36 |
| TRINITY_DN28846_c0_g1 | 3.2100 | gonadal aromatase | cyp19a1a | 3.64 × 10−43 |
| TRINITY_DN48025_c3_g3 | 1.2916 | estrogen receptor α | erα | 3.90 × 10−5 |
| Unigene ID | Log2FC | Annotation | Gene Name | FDR |
|---|---|---|---|---|
| TRINITY_DN23324_c0_g2 | −4.4445 | leptin A2 | leptin | 8.32 × 10−36 |
| TRINITY_DN46583_c1_g2 | −2.3537 | Leptin receptor | lepR | 6.96 × 10−16 |
| TRINITY_DN44455_c0_g2 | −1.3513 | Proopiomelanocortin | POMC | 0 |
| TRINITY_DN26070_c2_g3 | −1.3450 | Cocaine- and amphetamine-regulated transcript protein | CART | 3.24 × 10−9 |
| TRINITY_DN34546_c0_g4 | 1.2470 | agouti-related peptide | AgRP | 0.17 × 10−3 |
| TRINITY_DN32690_c1_g1 | 3.5570 | Neuropeptide Y | NPY | 2.75 × 10−36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, X.; Gao, Z.; Luo, S.; Su, J.; Yi, S. Evidence for the Growth Superiority and Delayed Ovarian Development in Tetraploid Loach Misgurnus anguillicaudatus. Fishes 2022, 7, 322. https://doi.org/10.3390/fishes7060322
Zhou X, Gao Z, Luo S, Su J, Yi S. Evidence for the Growth Superiority and Delayed Ovarian Development in Tetraploid Loach Misgurnus anguillicaudatus. Fishes. 2022; 7(6):322. https://doi.org/10.3390/fishes7060322
Chicago/Turabian StyleZhou, Xiaoyun, Zexia Gao, Shuangshuang Luo, Junxiao Su, and Shaokui Yi. 2022. "Evidence for the Growth Superiority and Delayed Ovarian Development in Tetraploid Loach Misgurnus anguillicaudatus" Fishes 7, no. 6: 322. https://doi.org/10.3390/fishes7060322
APA StyleZhou, X., Gao, Z., Luo, S., Su, J., & Yi, S. (2022). Evidence for the Growth Superiority and Delayed Ovarian Development in Tetraploid Loach Misgurnus anguillicaudatus. Fishes, 7(6), 322. https://doi.org/10.3390/fishes7060322