
Age, Growth, and Utility of Otolith Morphometrics as Predictors of Age in the European Barracuda, Sphyraena sphyraena (Sphyraenidae): Preliminary Data



Abstract
:
1. Introduction
2. Materials and Methods
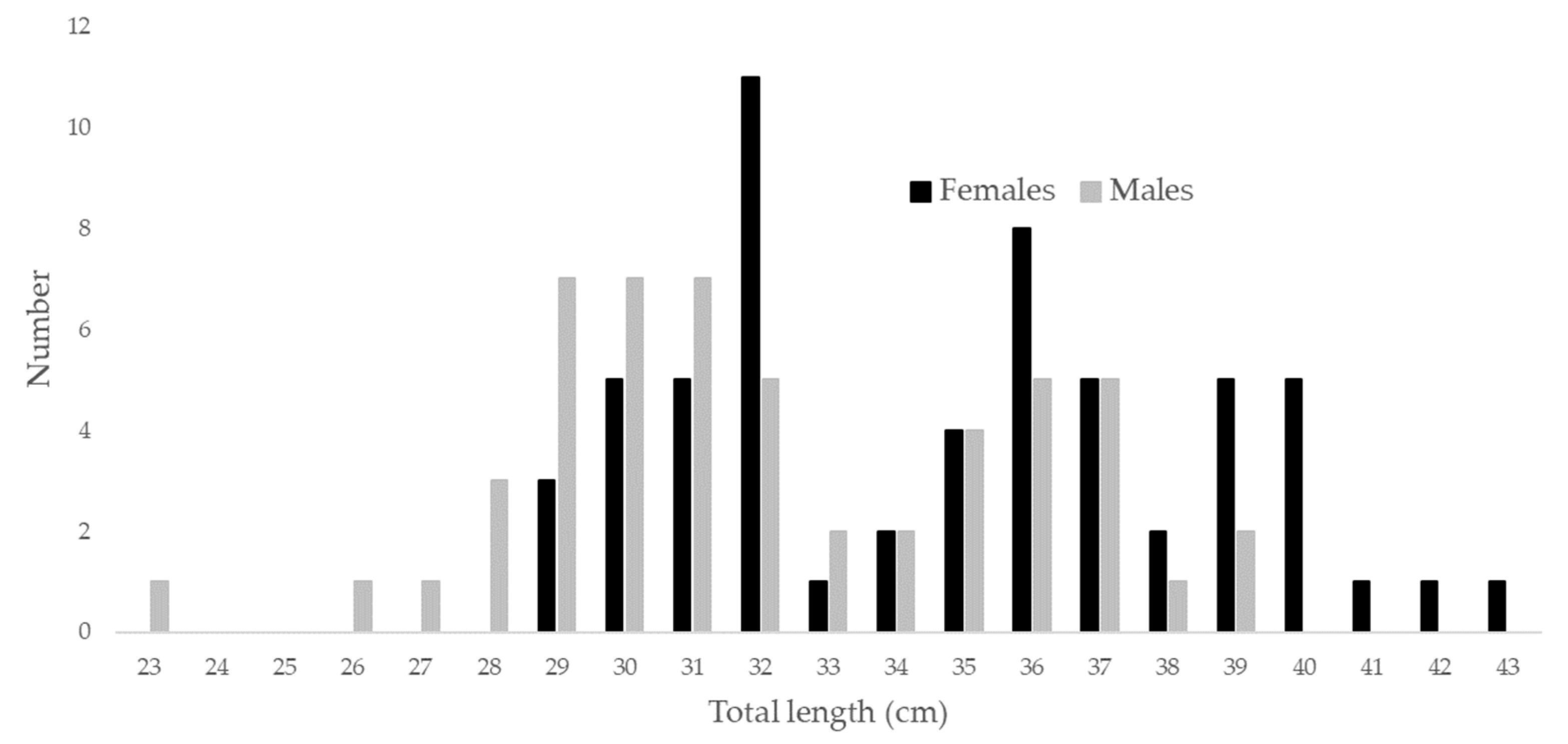
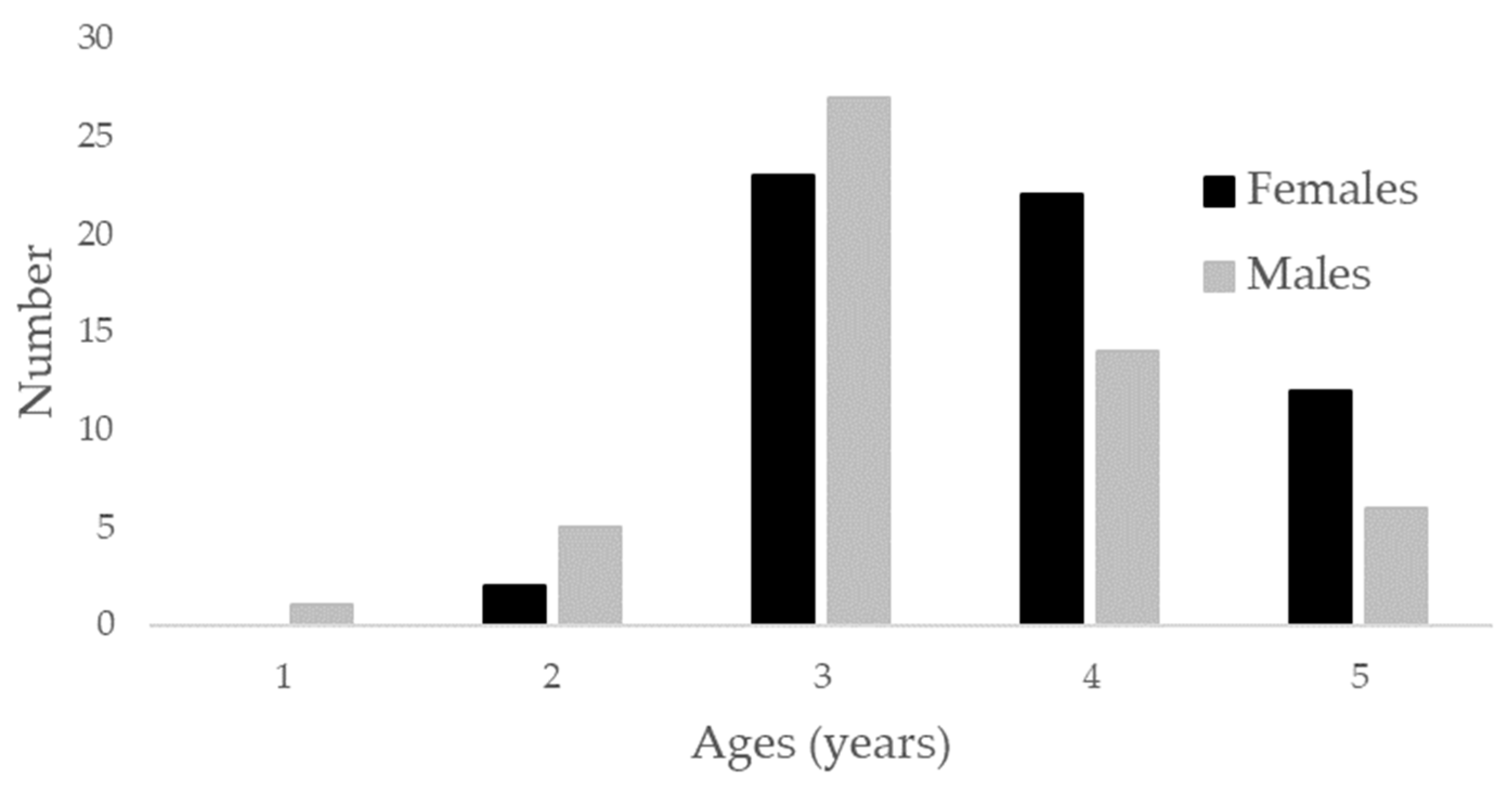
2.1. Sampling
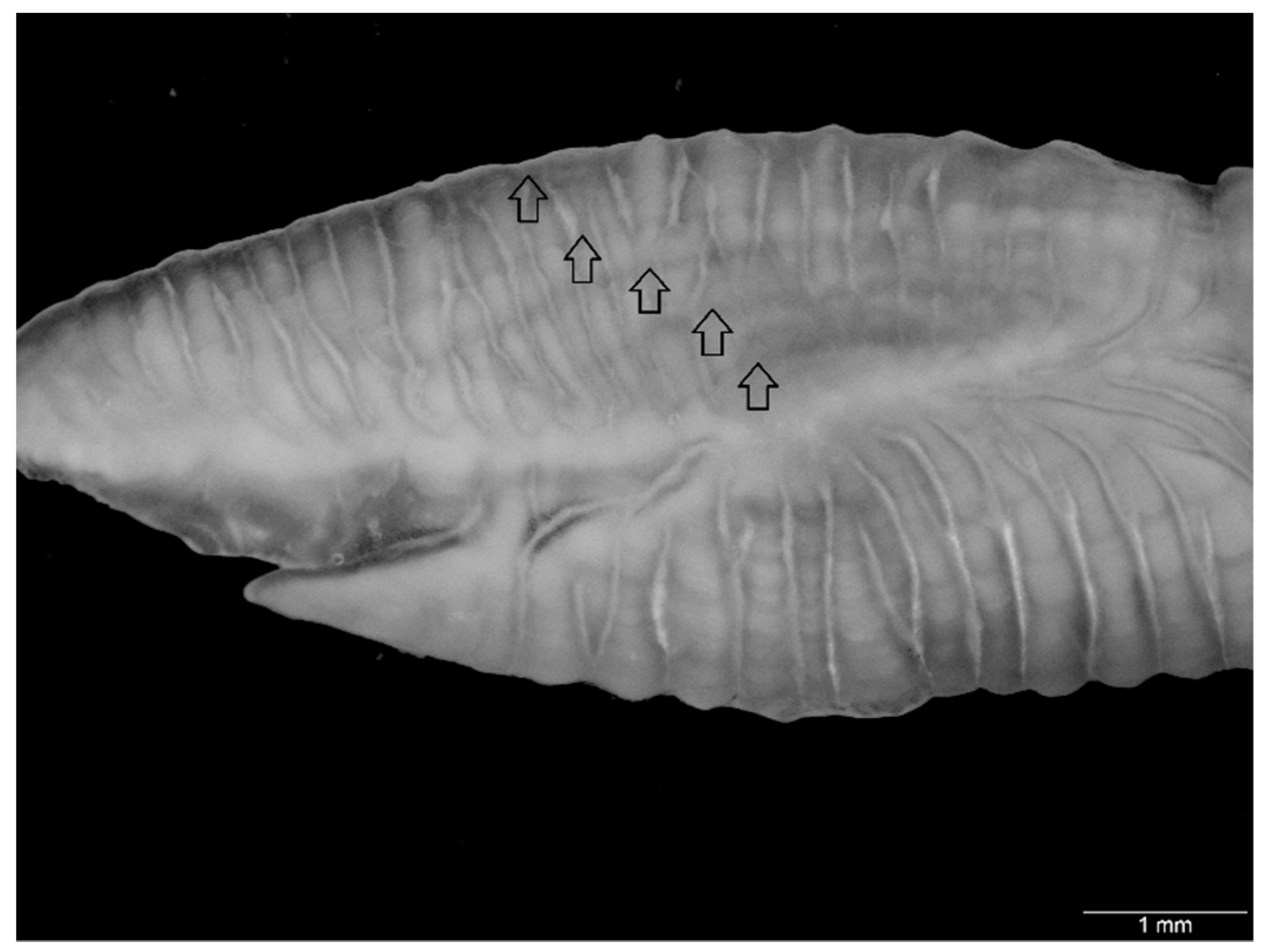
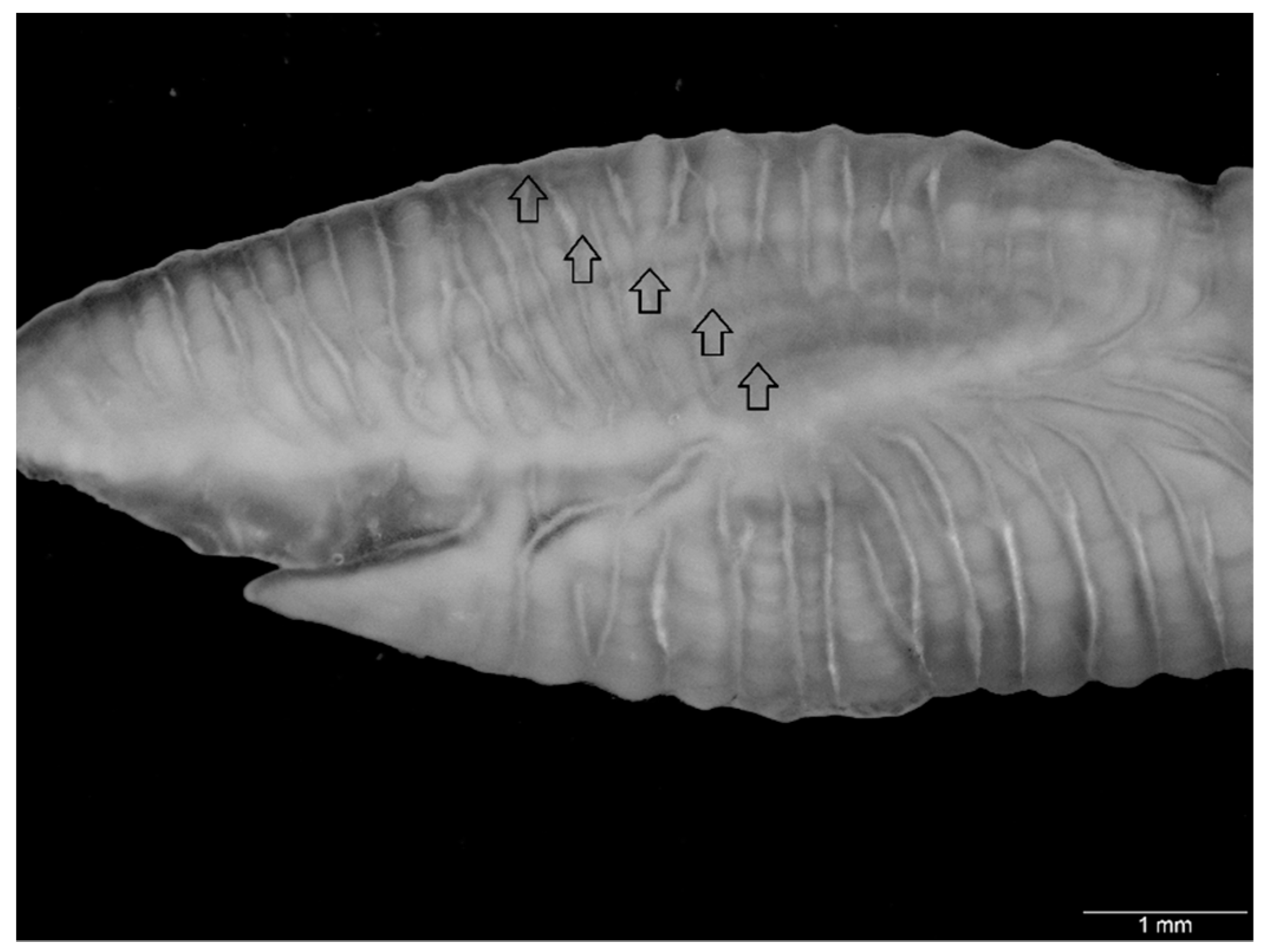
2.2. Otolith Morphometrics
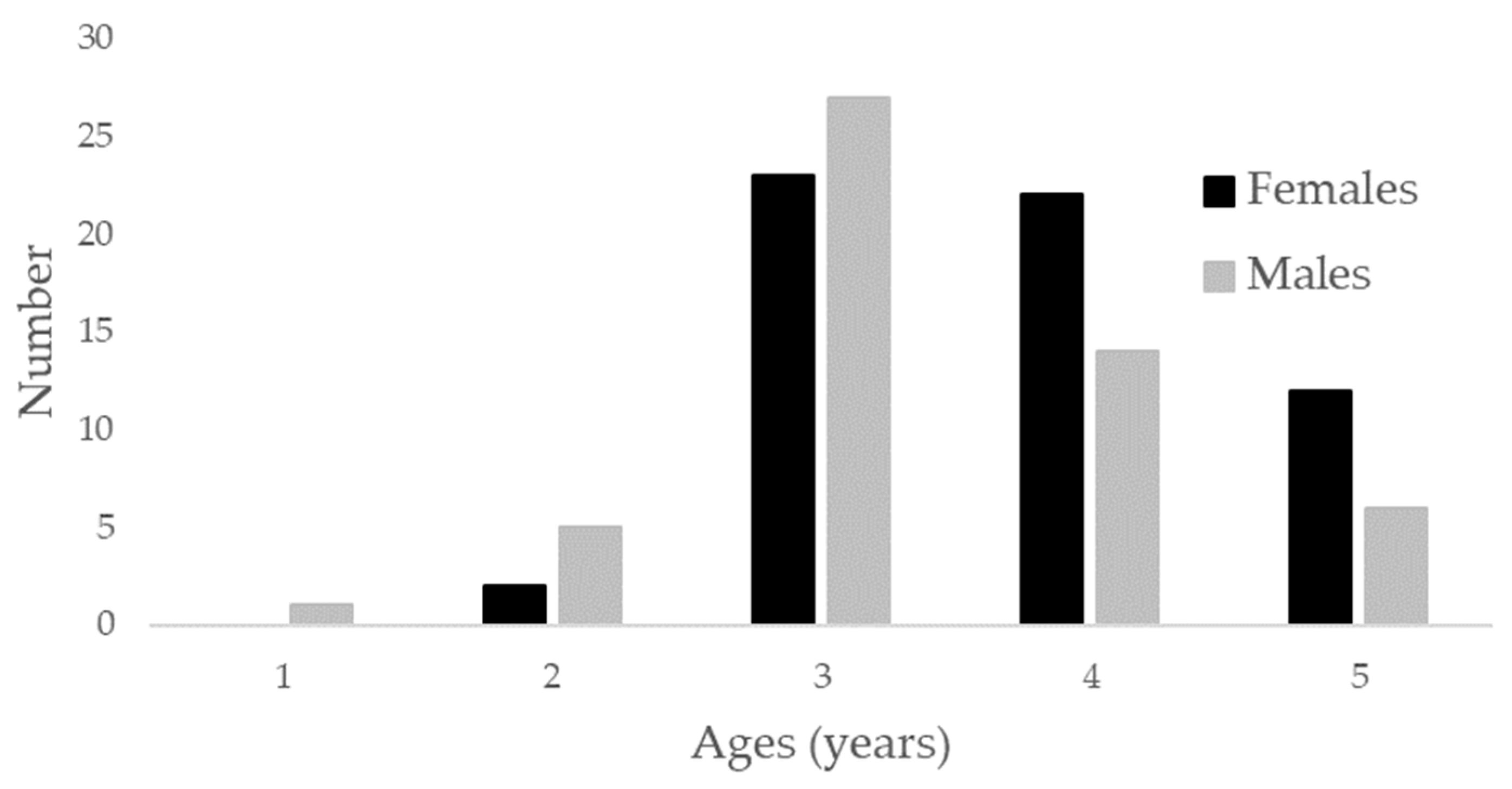
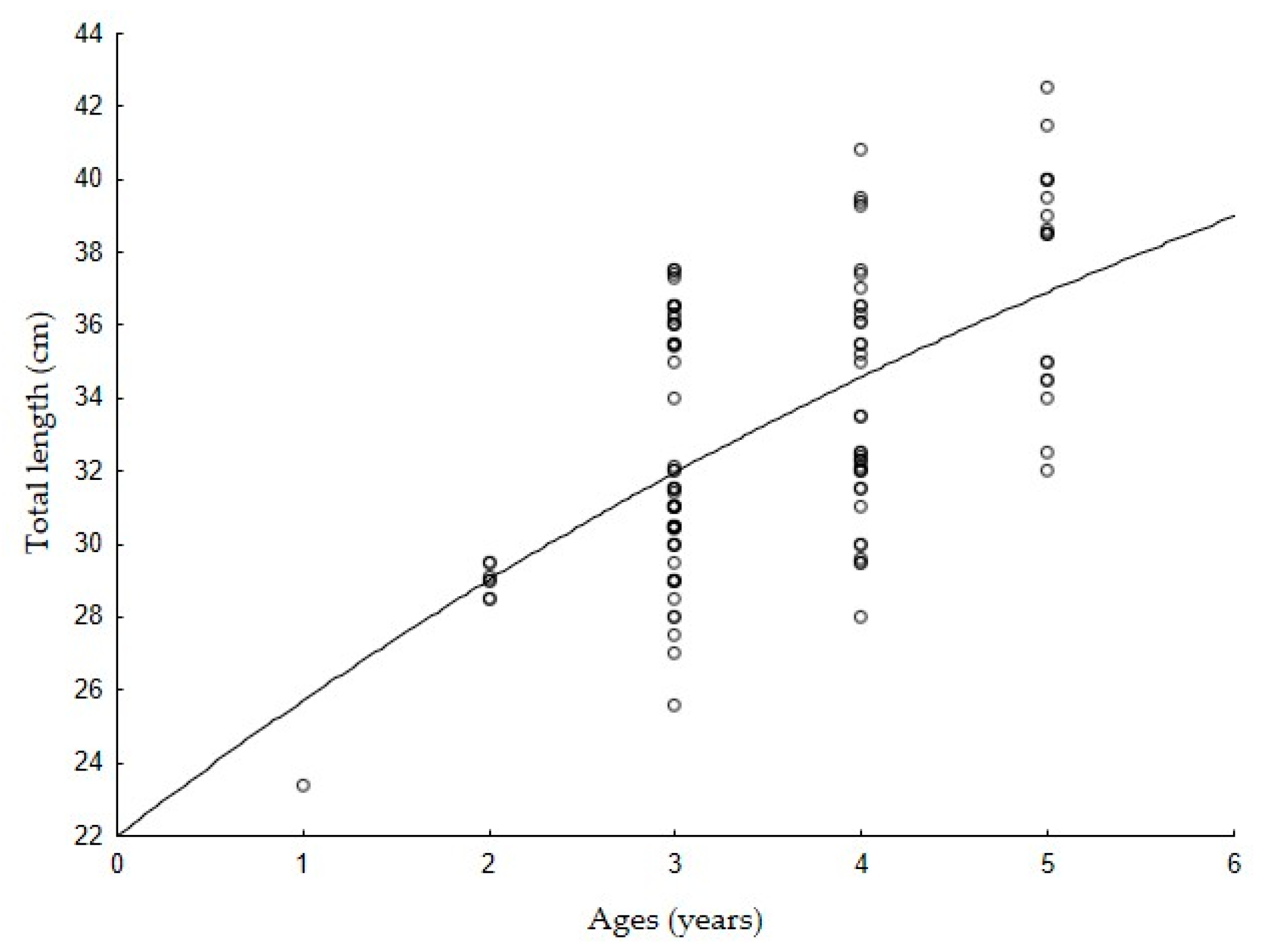
2.3. Age and Growth Estimation
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World, 5th ed.; Wiley: Hoboken, NJ, USA, 2016; pp. 468–472. [Google Scholar]
- ElGanainy, A.; Amin, A.; Ali, A.; Osman, H. Age and growth of two barracuda species Sphyraena chrysotaenia and S. flavicauda (Family: Sphyraenidae) from the Gulf of Suez, Egypt. Egypt. J. Aquat. Res. 2017, 43, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Ballen, G.A. Nomenclature of the Sphyraenidae (Teleostei: Carangaria): A synthesis of fossil- and extant-based classification systems. Zootaxa 2019, 4686, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Wadie, W.F.; Rizkalla, S.I. Fisheries for the genus Sphyraena (Perciformes, Sphyraenidae) in the south-eastern part of the Mediterranean Sea. Pak. J. Mar. Sci. 2001, 10, 21–34. [Google Scholar]
- Villegas-Hernández, H.; Muñoz, M.; Lloret, J. Life-history traits of temperate and thermophilic barracudas (Teleostei: Sphyraenidae) in the context of sea warming in the Mediterranean Sea. J. Fish Biol. 2014, 84, 1940–1957. [Google Scholar] [CrossRef] [PubMed]
- Pastore, A. Sphyraena intermedia sp. nov. (Pisces: Sphyraenidae): A potential new species of barracuda identified from the central Mediterranean Sea. J. Mar. Biol. Assoc. 2009, 89, 1299–1303. [Google Scholar] [CrossRef]
- Bourehail, N.; Morat, F.; Lecomte-Finiger, R.; Kara, M.H. Using otolith shape analysis to distinguish barracudas Sphyraena sphyraena and Sphyraena viridensis from the Algerian coast. Cybium 2015, 39, 271–278. [Google Scholar]
- Yedier, S. Otolith shape analysis and relationships between total length and otolith dimensions of European barracuda, Sphyraena sphyraena in the Mediterranean Sea. Iran. J. Fish. Sci. 2021, 20, 1080–1096. [Google Scholar]
- Froese, R.; Pauly, D. FishBase. World Wide Web Electronic Publication. Version (08/2021). Available online: www.fishbase.org (accessed on 10 January 2022).
- Golani, D.; Öztürk, B.; Başusta, N. The Fishes of Eastern Mediterranean; Turkish Marine Research Foundation: Istanbul, Turkey, 2006; pp. 179–181. [Google Scholar]
- Ben-Tuvia, A. Sphyraenidae. In Fishes of the North-Eastern Atlantic and the Mediterranean; Whitehead, P.J.P., Bauchot, M.L., Hureau, J.C., Nielsen, J., Tortonese, E., Eds.; UNESCO: Paris, France, 1986; pp. 1194–1196. [Google Scholar]
- Kalogirou, S.; Mittermayer, F.; Pihl, L.; Wennhage, H. Feeding ecology of indigenous and non-indigenous fish species within the family Sphyraenidae. J. Fish Biol. 2012, 80, 2528–2548. [Google Scholar] [CrossRef] [PubMed]
- Allam, S.M.; Faltas, S.N.; Ragheb, E. Age and growth of barracudas in the Egyptian Mediterranean waters. Egypt. J. Aquat. Res. 2004, 30, 281–289. [Google Scholar]
- Allam, S.M.; Faltas, S.N.; Ragheb, E. Reproductive biology of Sphyraena species in the Egyptian Mediterranean waters of Alexandria. Egypt. J. Aquat. Res. 2004, 30, 255–270. [Google Scholar]
- Bourehail, N.; Kara, M.H. Age, growth and mortality of the yellowmouth barracuda Sphyraena viridensis (Sphyraenidae) from eastern coasts of Algeria. J. Mar. Biol. Assoc. 2021, 101, 599–608. [Google Scholar] [CrossRef]
- Munro, J.L.; Pauly, D. A simple method for comparing the growth of fishes and invertebrates. Fishbyte 1983, 1, 5–6. [Google Scholar]
- Sparre, P.; Ursin, E.; Venema, S.C. Introduction to Tropical Fish Stock Assessment. Part 1: Manual; FAO Fisheries Technical Paper; FAO: Rome, Italy, 1987; pp. 75–76. [Google Scholar]
- Wootton, R.J. Ecology of Teleost Fishes; Chapman and Hall: England, UK, 2012; pp. 117–158. [Google Scholar]
- Matić-Skoko, S.; Ferri, J.; Škeljo, F.; Bartulović, V.; Glavić, K.; Glamuzina, B. Age, growth and validation of otolith morphometrics as predictors of age in the forkbeard, Phycis phycis (Gadidae). Fish. Res. 2011, 112, 52–58. [Google Scholar] [CrossRef]
- Cardinale, M.; Arrhenius, F. Using otolith weight to estimate the age of haddock (Melanogrammus aeglefinus): A tree model application. J. Appl. Ichthyol. 2004, 20, 470–475. [Google Scholar] [CrossRef]
- Pilling, G.M.; Grandcourt, E.M.; Kirkwood, G.P. The utility of otolith weight as a predictor of age in the emperor Lethrinus mahsena and other tropical fish species. Fish. Res. 2003, 60, 493–506. [Google Scholar] [CrossRef]
- Pino, A.C.; Cubillos, L.A.; Araya, M.; Sepúlveda, A. Otolith weight as an estimator of age in the Patagonian grenadier, Macruronus magellanicus, in central-south Chile. Fish. Res. 2004, 66, 145–156. [Google Scholar] [CrossRef]
- Doering-Arjes, P.; Cardinale, M.; Mosegaard, H. Estimating population age structure using otolith morphometrics: A test with known-age Atlantic cod (Gadus morhua) individuals. Can. J. Fish. Aquat. Sci. 2008, 65, 2341–2350. [Google Scholar] [CrossRef]
- Steward, C.A.; DeMaria, K.D.; Shenker, J.M. Using otolith morphometrics to quickly and inexpensively predict age in the gray angelfish (Pomacanthus arcuatus). Fish. Res. 2009, 99, 123–129. [Google Scholar] [CrossRef]
- Škeljo, F.; Ferri, J.; Brčić, J.; Petrić, M.; Jardas, I. Age, growth and utility of otolith morphometrics as a predictor of age in the wrasse Coris julis (Labridae) from the eastern Adriatic Sea. Sci. Mar. 2012, 76, 587–595. [Google Scholar] [CrossRef] [Green Version]
- Tuset, V.M.; Lombarte, A.; Assis, C.A. Otolith atlas for the western Mediterranean, north and central eastern Atlantic. Sci. Mar. 2008, 72, 7–198. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Length (cm) | Weight (g) | |||
|---|---|---|---|---|
| Range | Mean ± SD | Range | Mean ± SD | |
| Females | 28.5–42.5 | 34.6 ± 3.75 | 91.5–270.0 | 159.9 ± 47.09 |
| Males | 23.4–38.6 | 32.0 ± 3.56 | 47.3–224.2 | 131.7 ± 37.27 |
| All specimens | 23.4–42.5 | 33.4 ± 3.89 | 47.3–270.0 | 146.1 ± 44.95 |
| Otolith Measures | Range | Mean ± SD |
|---|---|---|
| OL (mm) | 7.56–12.96 | 10.20 ± 1.10 |
| OW (mm) | 2.43–4.09 | 3.29 ± 0.34 |
| OM (g) | 0.0157–0.0769 | 0.0390 ± 0.01 |
| Relationships | a | b | R2 |
|---|---|---|---|
| TL vs. OL | 1.683 | 0.256 | 0.865 |
| TL vs. OW | 0.690 | 0.078 | 0.805 |
| TL vs. OM | −0.073 | 0.003 | 0.851 |
| Linear Model | R2 |
|---|---|
| Age = 0.425 OL − 0.761 | 0.277 |
| Age = 1.294 OW − 0.694 | 0.255 |
| Age = 28.912 OM + 2.445 | 0.236 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferri, J.; Brzica, A. Age, Growth, and Utility of Otolith Morphometrics as Predictors of Age in the European Barracuda, Sphyraena sphyraena (Sphyraenidae): Preliminary Data. Fishes 2022, 7, 68. https://doi.org/10.3390/fishes7020068
Ferri J, Brzica A. Age, Growth, and Utility of Otolith Morphometrics as Predictors of Age in the European Barracuda, Sphyraena sphyraena (Sphyraenidae): Preliminary Data. Fishes. 2022; 7(2):68. https://doi.org/10.3390/fishes7020068
Chicago/Turabian StyleFerri, Josipa, and Anđela Brzica. 2022. "Age, Growth, and Utility of Otolith Morphometrics as Predictors of Age in the European Barracuda, Sphyraena sphyraena (Sphyraenidae): Preliminary Data" Fishes 7, no. 2: 68. https://doi.org/10.3390/fishes7020068
APA StyleFerri, J., & Brzica, A. (2022). Age, Growth, and Utility of Otolith Morphometrics as Predictors of Age in the European Barracuda, Sphyraena sphyraena (Sphyraenidae): Preliminary Data. Fishes, 7(2), 68. https://doi.org/10.3390/fishes7020068