Use of Environmental DNA to Detect Grass Carp Spawning Events

 , and
, and
Abstract
1. Introduction
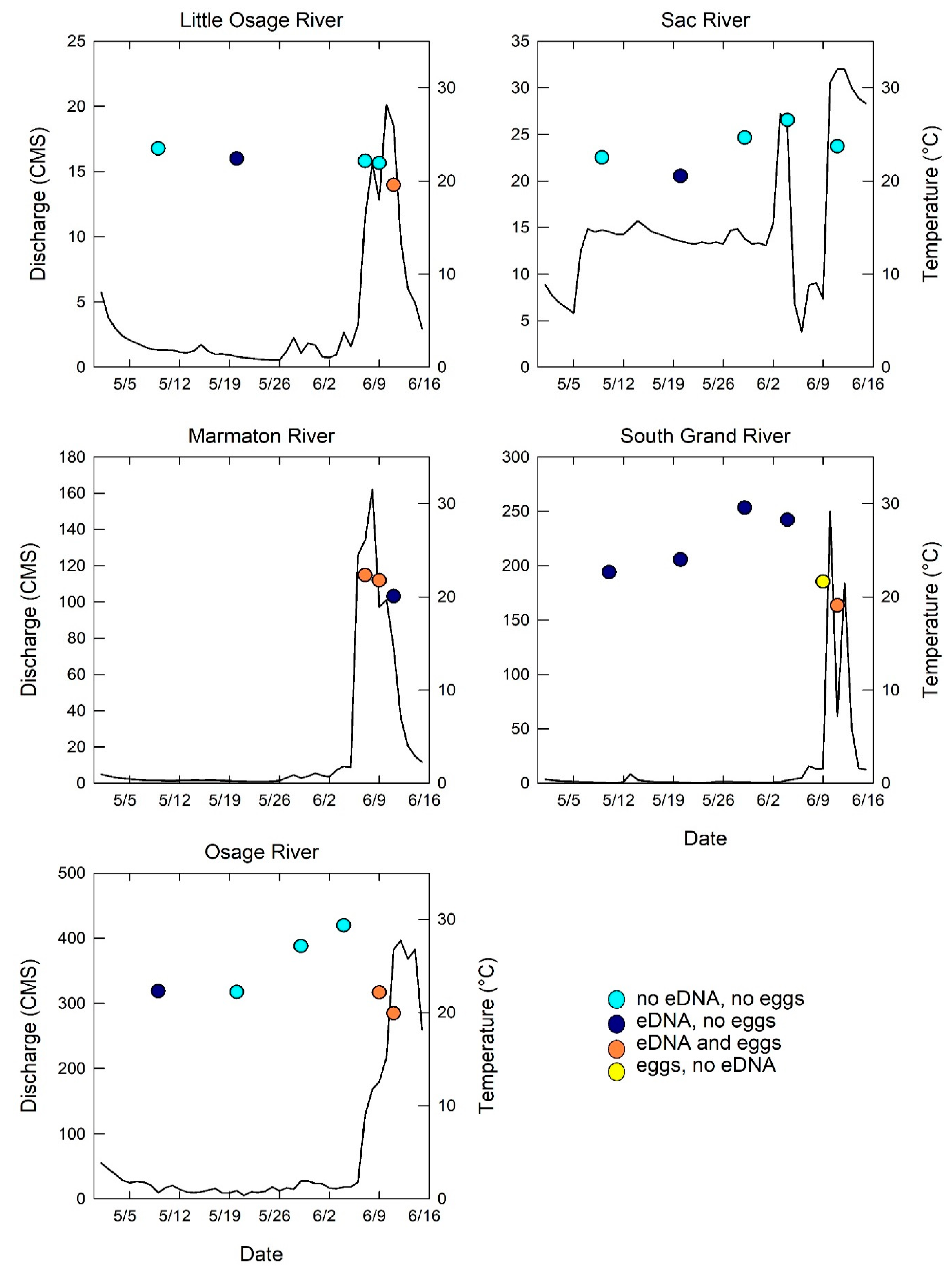
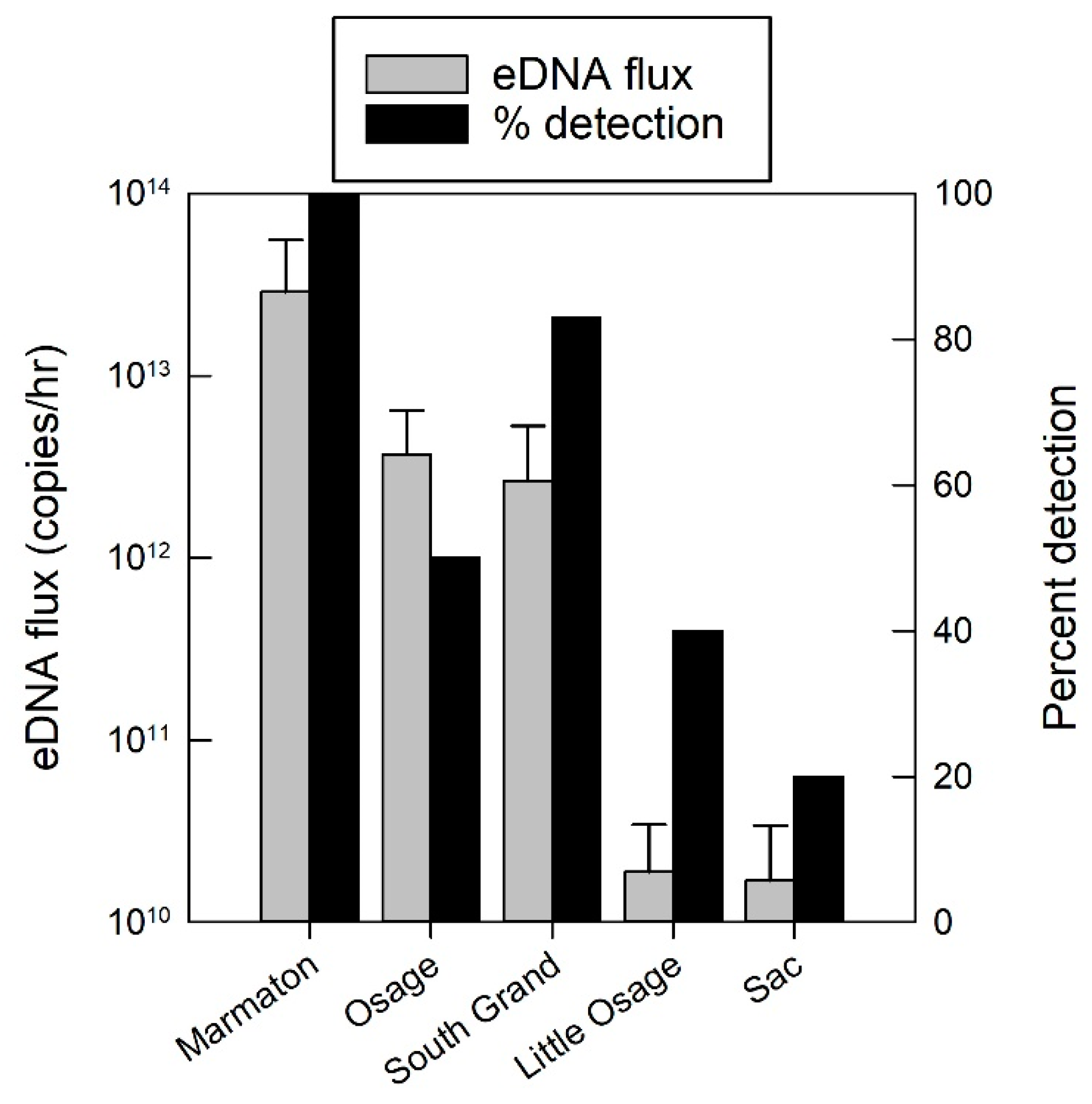
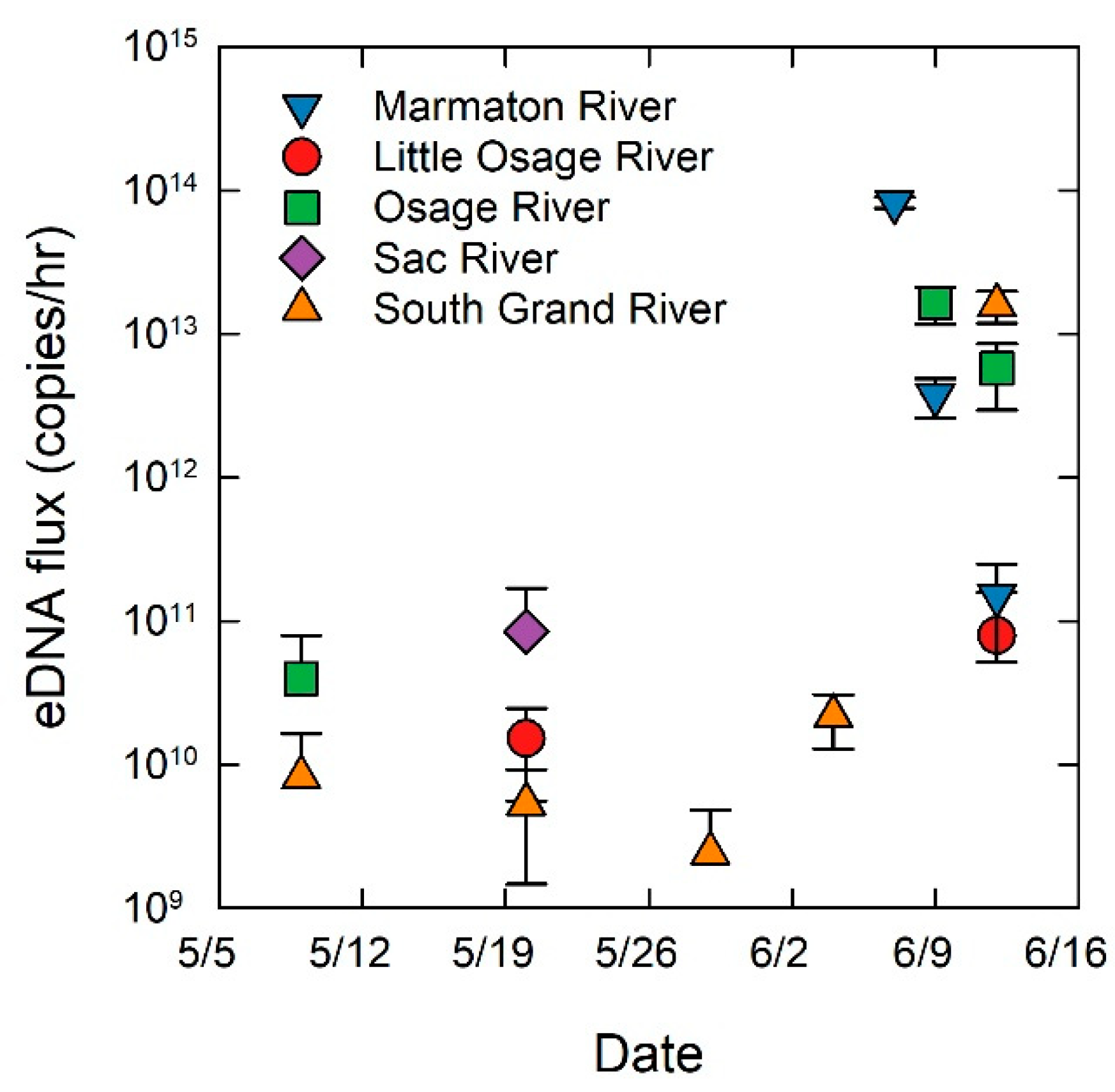
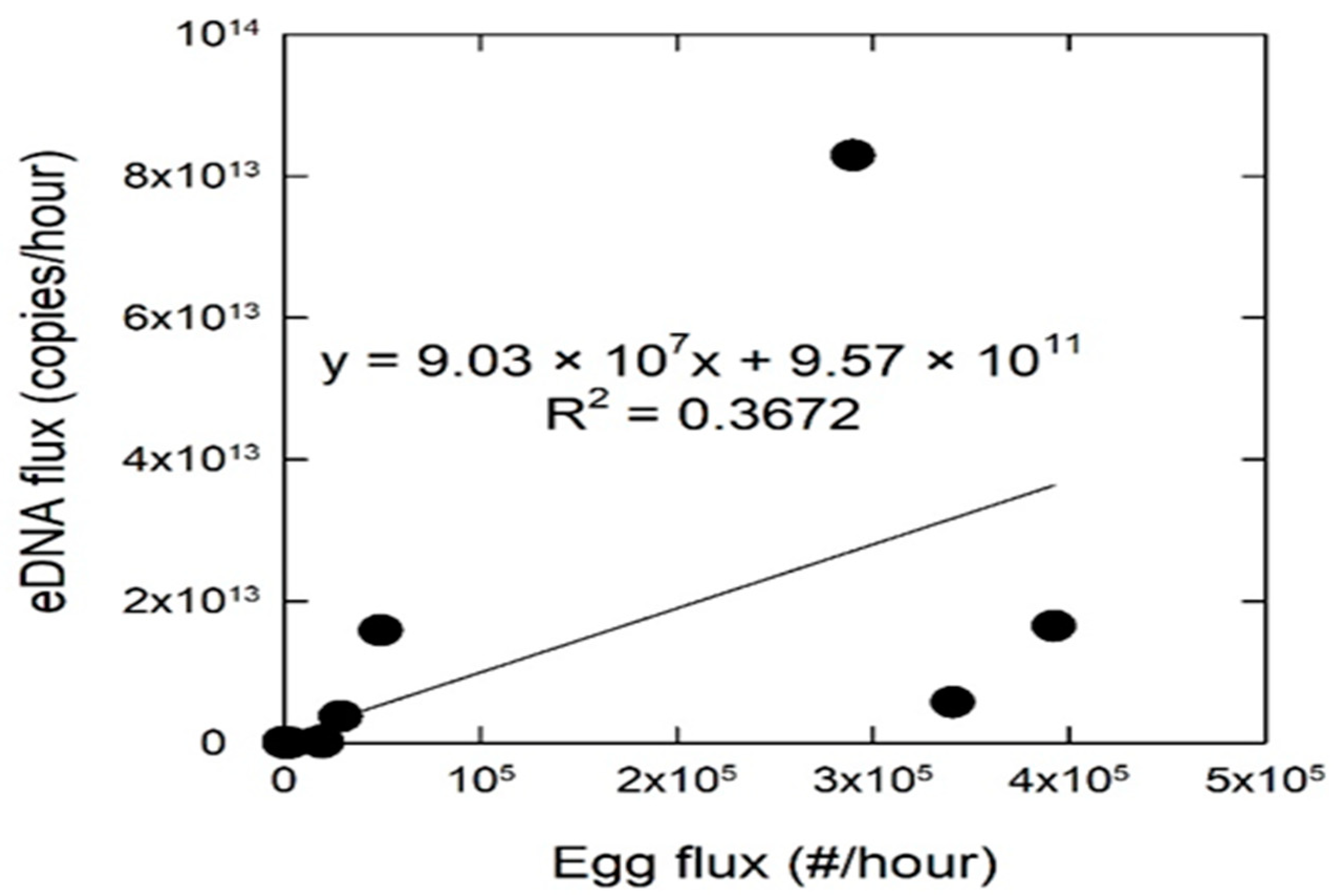
2. Results
3. Discussion
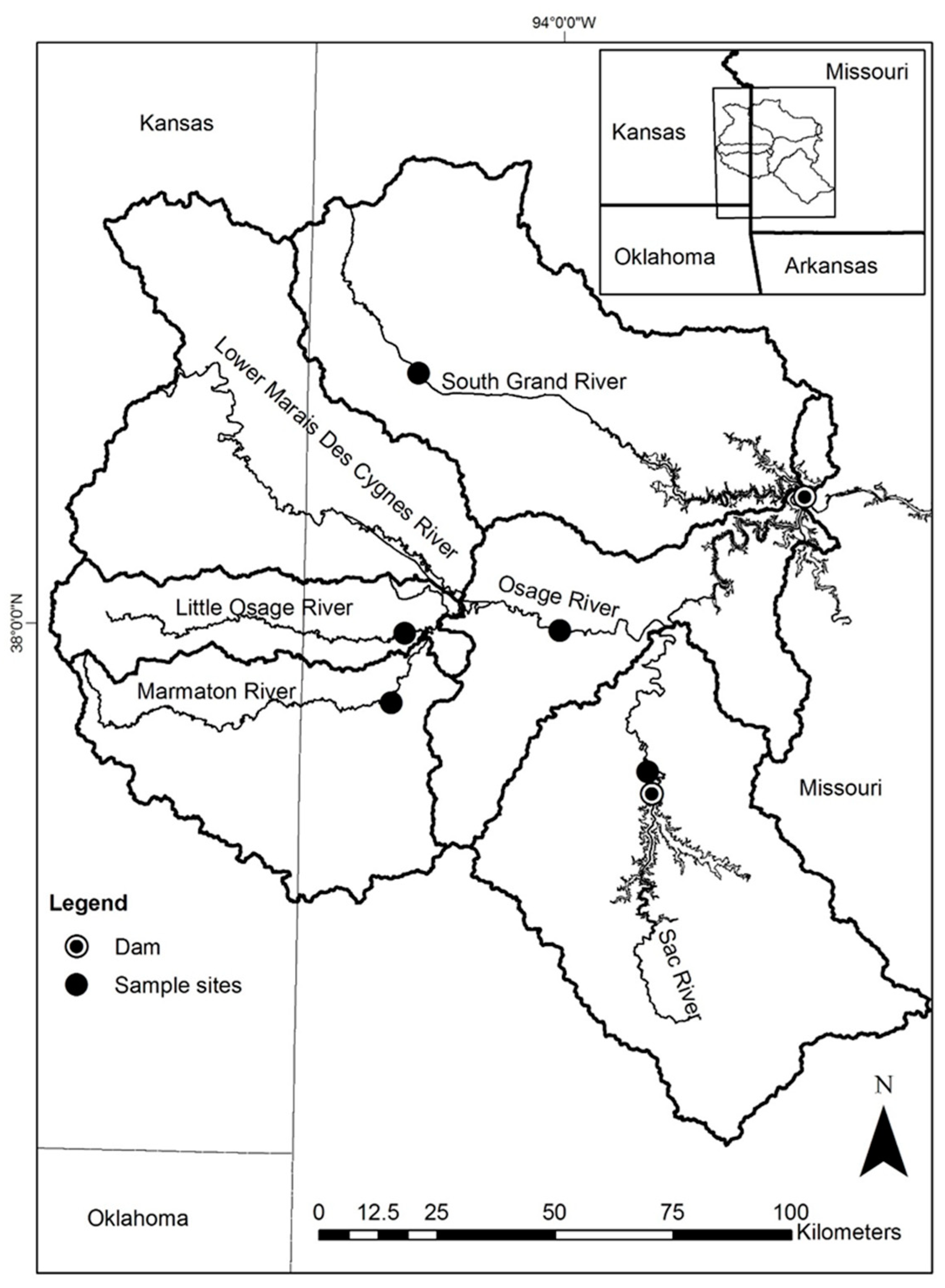
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kolar, C.S.; Chapman, D.C.; Courtenay, W.R., Jr.; Housel, C.M.; Williams, J.D.; Jennings, D.P. Bigheaded Carps: A Biological Synopsis and Environmental Risk Assessment; American Fisheries Society: Bethesda, MD, USA, 2007; p. 204. [Google Scholar]
- George, A.E.; Chapman, D.C. Embryonic and larval development and early behavior in grass carp, Ctenopharyngodon idella: Implications for recruitment in rivers. PLoS ONE 2015, 10, e0119023. [Google Scholar] [CrossRef] [PubMed]
- George, A.E.; Chapman, D.C. Aspects of embryonic and larval development in bighead carp Hypophthalmichthys nobilis and silver carp Hypophthalmichthys molitrix. PLoS ONE 2013, 8, e73829. [Google Scholar] [CrossRef]
- Murphy, E.A.; Jackson, P.R. Hydraulic and Water-Quality Data Collection for the Investigation of Great Lakes Tributaries for Asian Carp Spawning and Egg-Transport Suitability; Report. 2013-5106; U.S. Geological Survey: Reston, VA, USA, 2013; p. 40.
- Garcia, T.; Murphy, E.A.; Jackson, P.R.; Garcia, M.H. Application of the FluEgg model to predict transport of Asian carp eggs in the Saint Joseph River (Great Lakes tributary). J. Great Lakes Res. 2015, 41, 374–386. [Google Scholar] [CrossRef]
- George, A.E.; Garcia, T.; Chapman, D.C. Comparison of size, terminal fall velocity, and density of Bighead Carp, Silver Carp, and Grass Carp eggs for use in drift modeling. Trans. Am. Fish. Soc. 2017, 146, 834–843. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Miaud, C.; Pompanon, F.; Taberlet, P. Species detection using environmental DNA from water samples. Biol. Lett. 2008, 4, 423–425. [Google Scholar] [CrossRef] [PubMed]
- Cudmore, B.; Mandrak, N.E.; Dettmers, J.M.; Chapman, D.C.; Conover, G.; Kolar, C.S. Binational Risk Assessment of Grass Carp in the Great Lakes; Research Document 2016/118; Fisheries and Oceans Canada; Canadian Science Advisory Secretariat: Ottawa, ON, Canada, 2017. [Google Scholar]
- Yi, B.; Yu, Z.; Liang, Z.; Sujuan, S.; Xu, Y.; Chen, J.; He, M.; Liu, Y.; Hu, Y.; Deng, Z.; et al. The distribution, natural conditions, and breeding production of the spawning ground of four famous freshwater fishes on the main stream of the Yangtze River. In Gezhouba Water Control Project and Four Famous Fishes in the Yangtze River; Yi, B., Yi, Z., Liang, Z., Eds.; Hubei Science and Technology Press: Wuhan, China, 1988. [Google Scholar]
- Larson, J.H.; Knights, B.C.; McCalla, S.G.; Monroe, E.; Tuttle-Lau, M.; Chapman, D.C.; George, A.E.; Vallazza, J.M.; Amberg, J. Evidence of Asian Carp spawning upstream of a key choke point in the Mississippi River. N. Am. J. Fish. Manag. 2017, 37, 903–919. [Google Scholar] [CrossRef]
- Klymus, K.E.; Merkes, C.M.; Allison, M.J.; Goldberg, C.S.; Helbing, C.C.; Hunter, M.E.; Jackson, C.A.; Lance, R.F.; Mangan, A.M.; Monroe, E.M.; et al. Reporting the limits of detection and quantification for environmental DNA assays. Environ. DNA 2019. [Google Scholar] [CrossRef]
- Hargrave, C.W.; Gido, K.B. Evidence of reproduction by exotic grass carp in the Red and Washita rivers, Oklahoma. Southwest. Nat. 2004, 49, 89–93. [Google Scholar] [CrossRef]
- Bozkurt, Y.; Öğretmen, F. Sperm quality, egg size, fecundity and their relationships with fertilization rate of grass carp (Ctenopharyngodon idella). Iran. J. Fish. Sci. 2012, 11, 755–764. [Google Scholar]
- Verma, D.K.; Routray, P.; Dash, C.; Dasgupta, S.; Jena, J.K. Physical and biochemical characteristics of semen and ultrastructure of spermatozoa in six carp species. Turk. J. Fish. Aquat. Sci. 2009, 9, 67–76. [Google Scholar]
- Barnes, M.A.; Turner, C.R.; Jerde, C.L.; Renshaw, M.A.; Chadderton, W.L.; Lodge, D.M. Environmental conditions influence eDNA persistence in aquatic systems. Environ. Sci. Technol. 2014, 48, 1819–1827. [Google Scholar] [CrossRef] [PubMed]
- Strickler, K.M.; Fremier, A.K.; Goldberg, C.S. Quantifying effects of UV-B, temperature, and pH on eDNA degradation in aquatic microcosms. Biol. Conserv. 2015, 183, 85–92. [Google Scholar] [CrossRef]
- Eichmiller, J.J.; Best, S.E.; Sorensen, P.W. Effects of temperature and trophic state on degradation of environmental DNA in lake water. Environ. Sci. Technol. 2016, 50, 1859–1867. [Google Scholar] [CrossRef] [PubMed]
- Lance, R.F.; Klymus, K.E.; Richter, C.A.; Guan, X.; Farrington, H.L.; Carr, M.R.; Thompson, N.; Chapman, D.C.; Baerwaldt, K.L. Experimental observations on the decay of environmental DNA from bighead and silver carps. Manag. Biol. Invasions 2017, 8, 343–359. [Google Scholar] [CrossRef]
- Deters, J.E.; Chapman, D.C.; McElroy, B. Location and timing of Asian carp spawning in the Lower Missouri River. Environ. Biol. Fishes 2013, 96, 617–629. [Google Scholar] [CrossRef]
- Coulter, A.A.; Keller, D.; Bailey, E.J.; Goforth, R.R. Predictors of bigheaded carp drifting egg density and spawning activity in an invaded, free-flowing river. J. Great Lakes Res. 2016, 42, 83–89. [Google Scholar] [CrossRef]
- Turner, C.R.; Barnes, M.A.; Xu, C.C.Y.; Jones, S.E.; Jerde, C.L.; Lodge, D.M. Particle size distribution and optimal capture of aqueous macrobial eDNA. Methods Ecol. Evol. 2014, 5, 676–684. [Google Scholar] [CrossRef]
- Michaletz, P.H.; Siepker, M.J. Trends and synchrony in Black Bass and Crappie recruitment in Missouri reservoirs. Trans. Am. Fish. Soc. 2013, 142, 105–118. [Google Scholar] [CrossRef]
- Yi, B.; Liang, Z.; Yu, Z.; Lin, R.; He, M. A study of the early development of grass carp, black carp, silver carp and bighead carp of the Yangtze River. In Early Development of Four Cyprinids Native to the Yangtze River; Chapman, D.C., Ed.; Data Series 239; U.S. Geological Survey: Reston, VA, USA, 2006; pp. 15–51. [Google Scholar]
- Klymus, K.E.; Richter, C.A.; Chapman, D.C.; Paukert, C. Quantification of eDNA shedding rates from invasive bighead carp Hypophthalmichthys nobilis and silver carp Hypophthalmichthys molitrix. Biol. Conserv. 2015, 183, 77–84. [Google Scholar] [CrossRef]
- Coulter, A.A.; Keller, D.; Amberg, J.J.; Bailey, E.J.; Goforth, R.R. Phenotypic plasticity in the spawning traits of bigheaded carp (Hypophthalmichthys spp.) in novel ecosystems. Freshwat. Biol. 2013, 58, 1029–1037. [Google Scholar] [CrossRef]
- Wilson, C.; Wright, E.; Bronnenhuber, J.; MacDonald, F.; Belore, M.; Locke, B. Tracking ghosts: Combined electrofishing and environmental DNA surveillance efforts for Asian carps in Ontario waters of Lake Erie. Manag. Biol. Invasions 2014, 5, 225–231. [Google Scholar] [CrossRef]
- Farrington, H.L.; Edwards, C.E.; Guan, X.; Carr, M.R.; Baerwaldt, K.; Lance, R.F. Mitochondrial genome sequencing and development of genetic markers for the detection of DNA of invasive bighead and silver carp (Hypophthalmichthys nobilis and H. molitrix) in environmental water samples from the United States. PLoS ONE 2015, 10, e0117803. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| River | USGS Gage Number | Distance (Gage to Site rkm 1) | Basin Area (km2) | Sample Site Latitude | Sample Site Longitude | Distance to Truman Reservoir (rkm) | Upstream Barrier Distance (rkm) |
|---|---|---|---|---|---|---|---|
| Little Osage | 06917060 | 8 | 1504 | 38.00658 | −94.31944 | 107 | |
| Marmaton | 06918060 | 26 | 2956 | 37.99797 | −94.31951 | 106 | |
| Osage | 06918250 | 0 | 3117 | 38.003889 | −93.99472 | 48 | |
| Sac | 0691990 | 26 | 5104 | 37.94902 | −93.76917 | 31 | Stockton Dam (40) |
| South Grand | 06921760 | 32 | 5302 | 38.45243 | −94.00636 | 28 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayer, C.-A.; Bayless, M.F.; George, A.; Thompson, N.; Richter, C.A.; Chapman, D.C. Use of Environmental DNA to Detect Grass Carp Spawning Events. Fishes 2020, 5, 27. https://doi.org/10.3390/fishes5030027
Hayer C-A, Bayless MF, George A, Thompson N, Richter CA, Chapman DC. Use of Environmental DNA to Detect Grass Carp Spawning Events. Fishes. 2020; 5(3):27. https://doi.org/10.3390/fishes5030027
Chicago/Turabian StyleHayer, Cari-Ann, Michael F. Bayless, Amy George, Nathan Thompson, Catherine A. Richter, and Duane C. Chapman. 2020. "Use of Environmental DNA to Detect Grass Carp Spawning Events" Fishes 5, no. 3: 27. https://doi.org/10.3390/fishes5030027
APA StyleHayer, C.-A., Bayless, M. F., George, A., Thompson, N., Richter, C. A., & Chapman, D. C. (2020). Use of Environmental DNA to Detect Grass Carp Spawning Events. Fishes, 5(3), 27. https://doi.org/10.3390/fishes5030027