Description of the Annual Reproductive Cycle of Wreckfish Polyprion americanus in Captivity

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
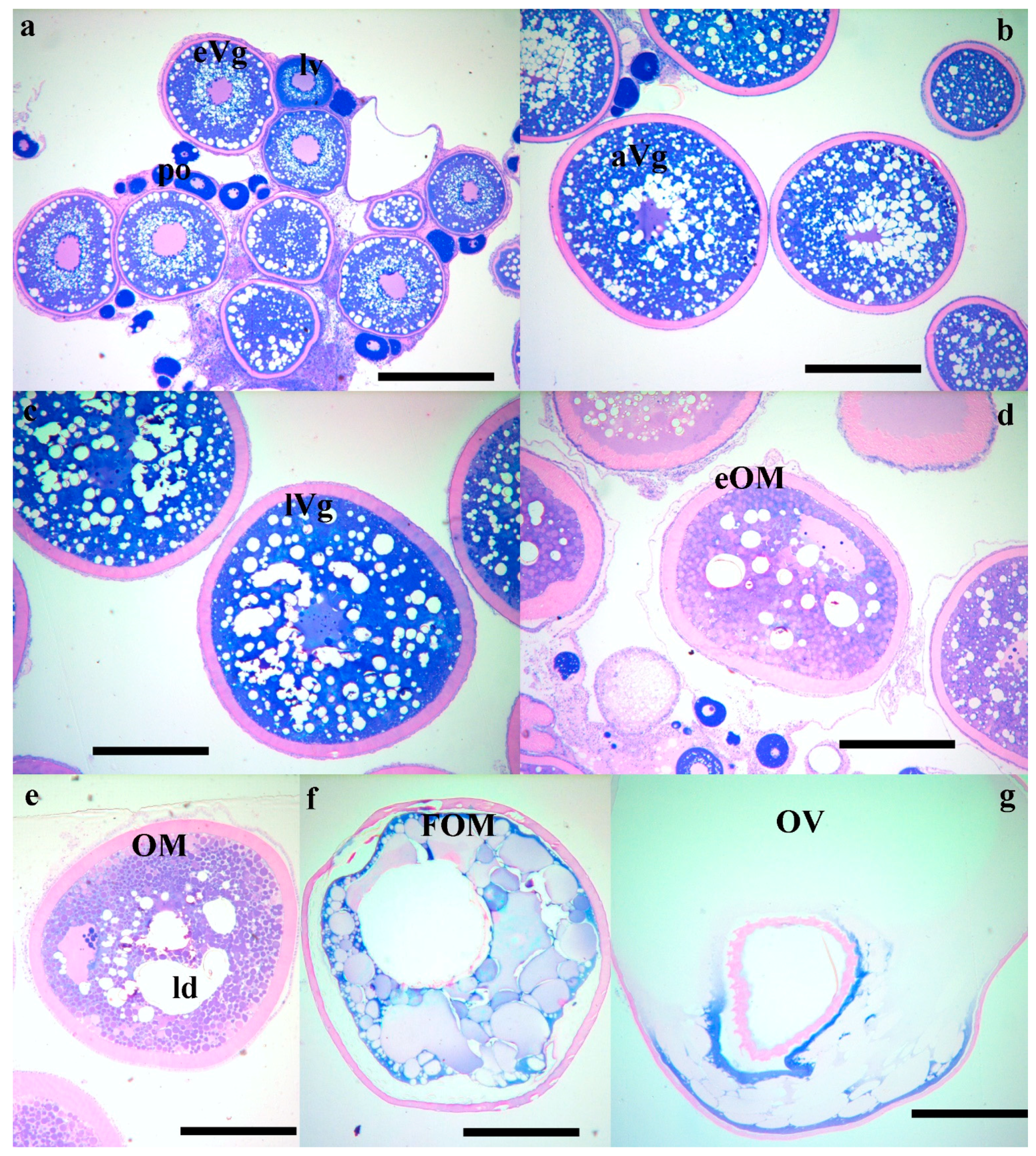
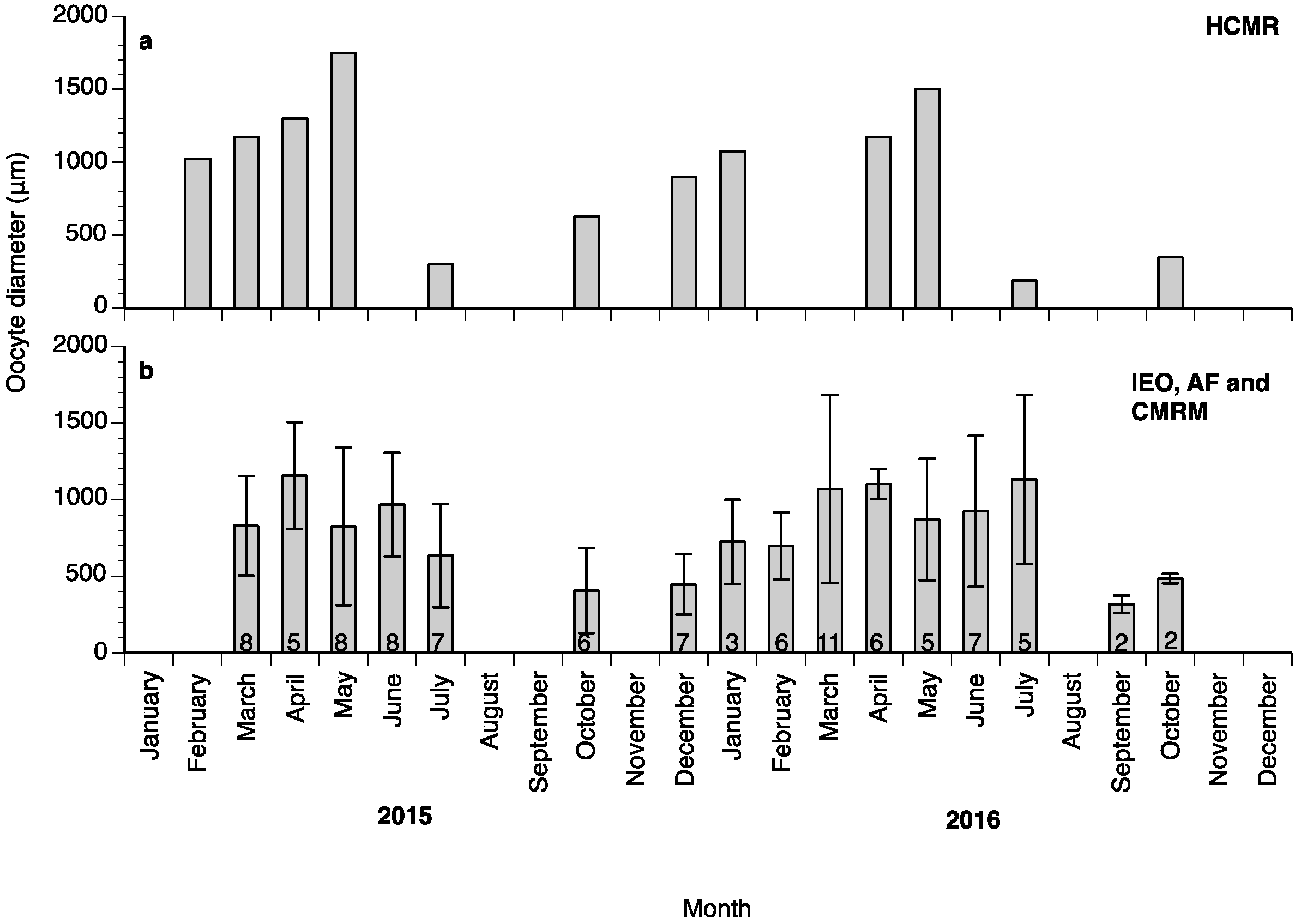
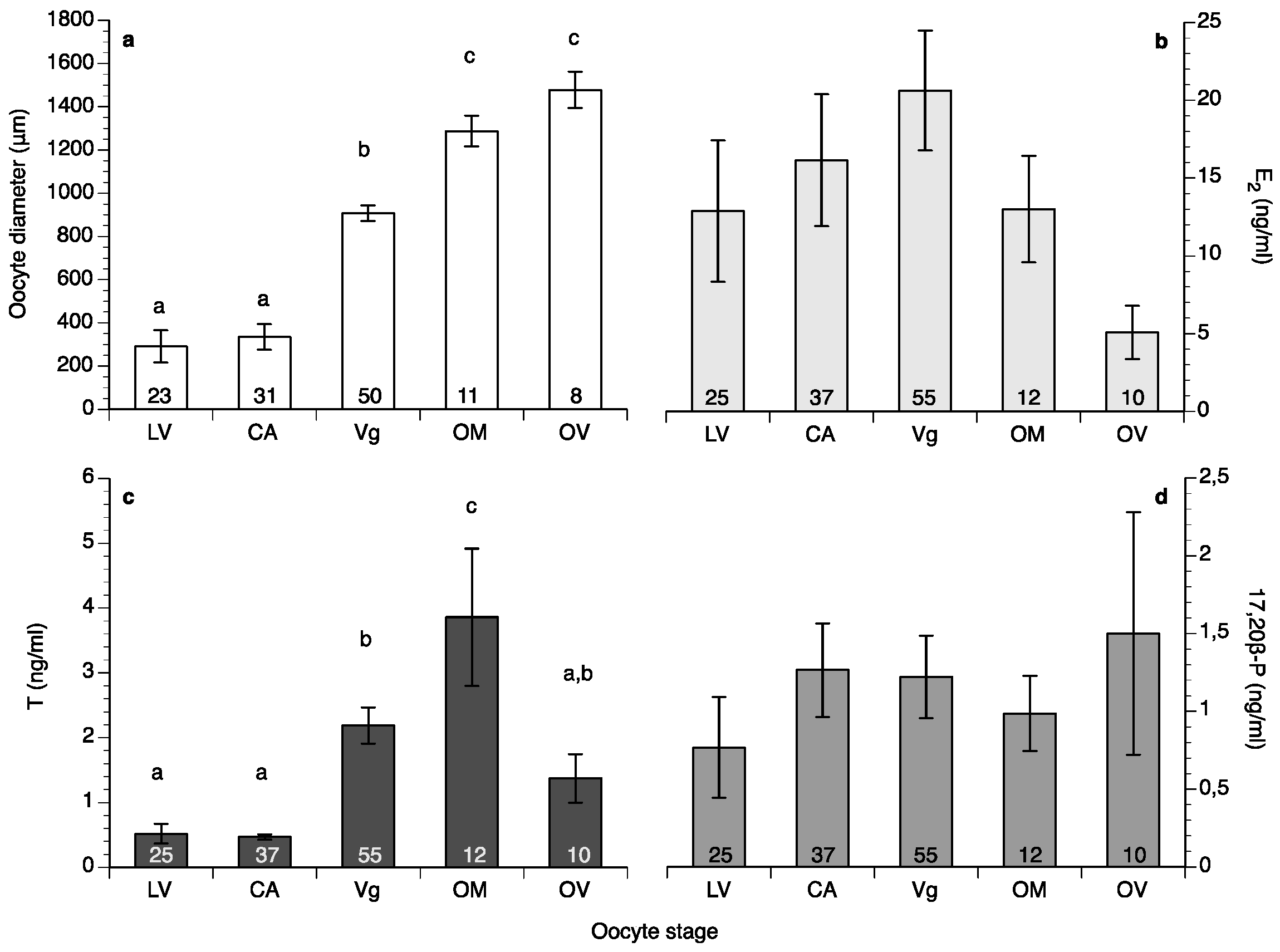
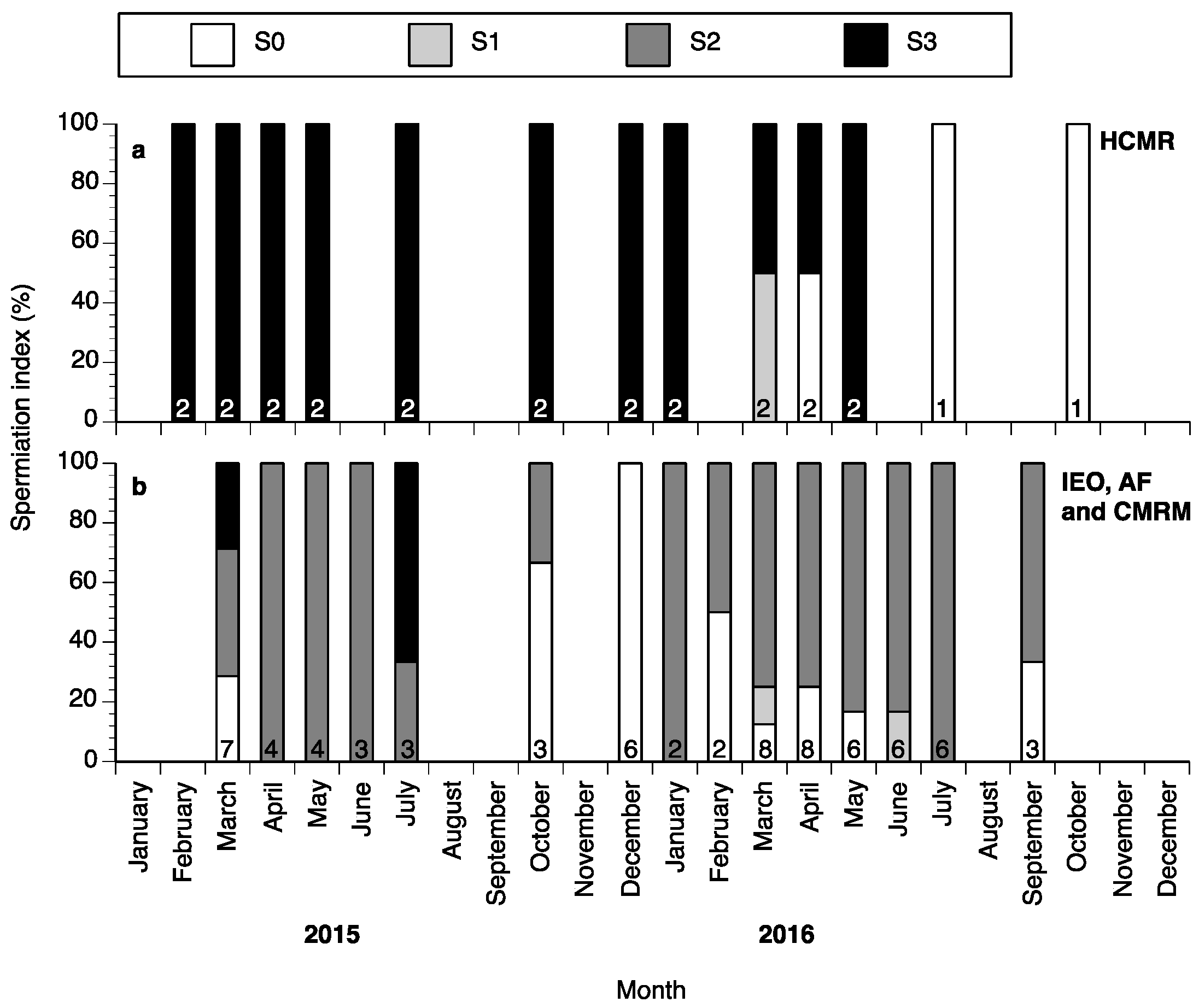
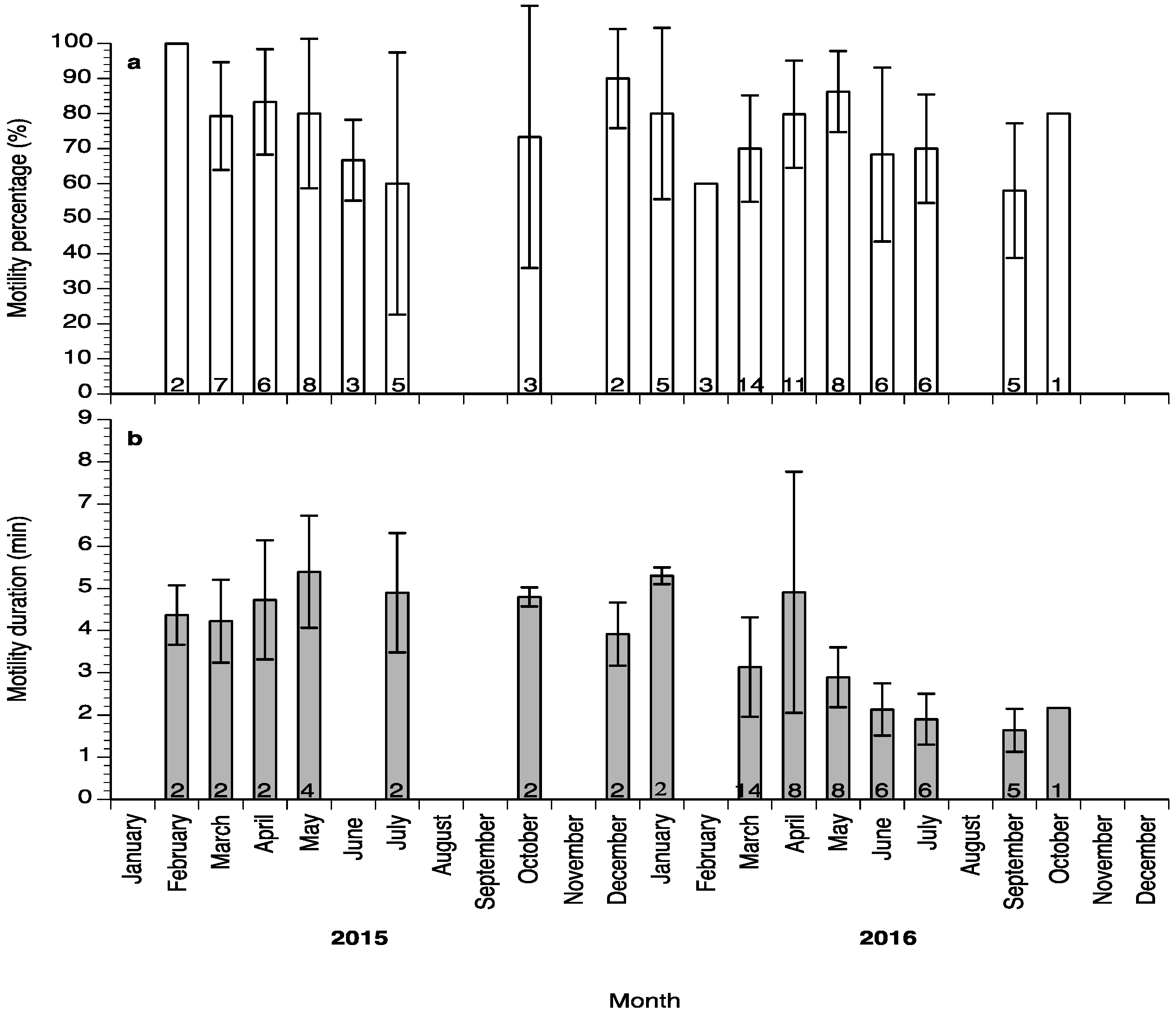
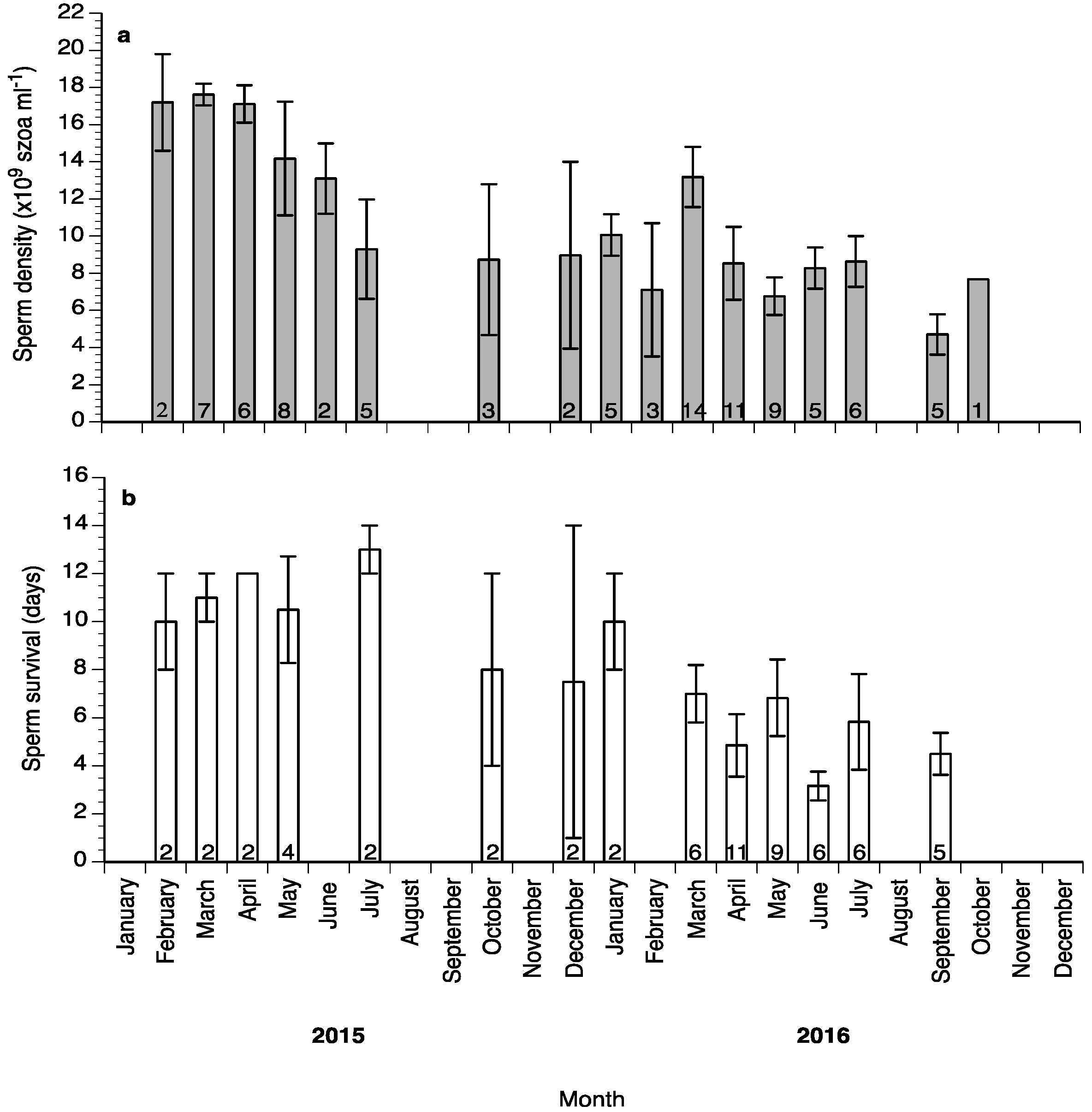
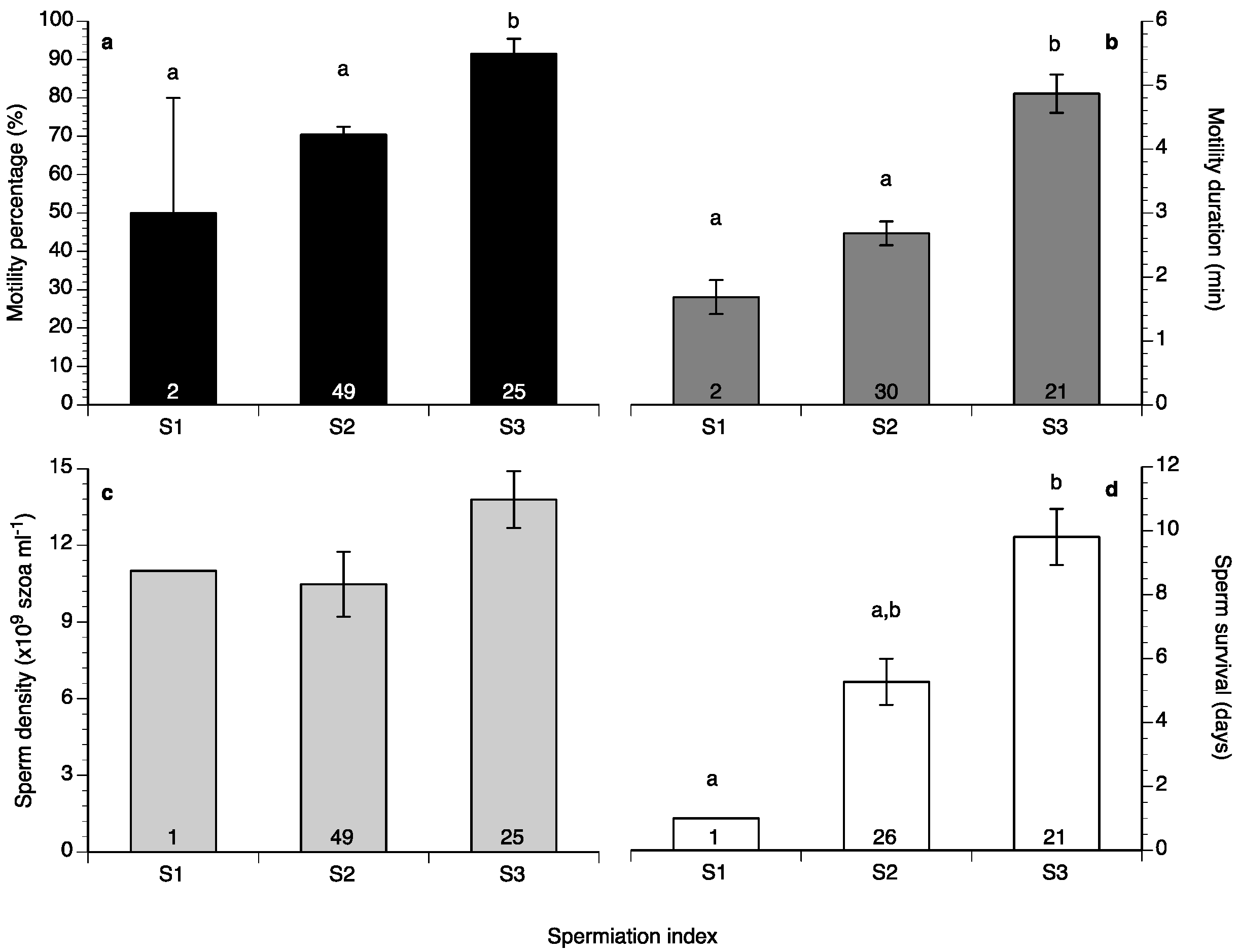
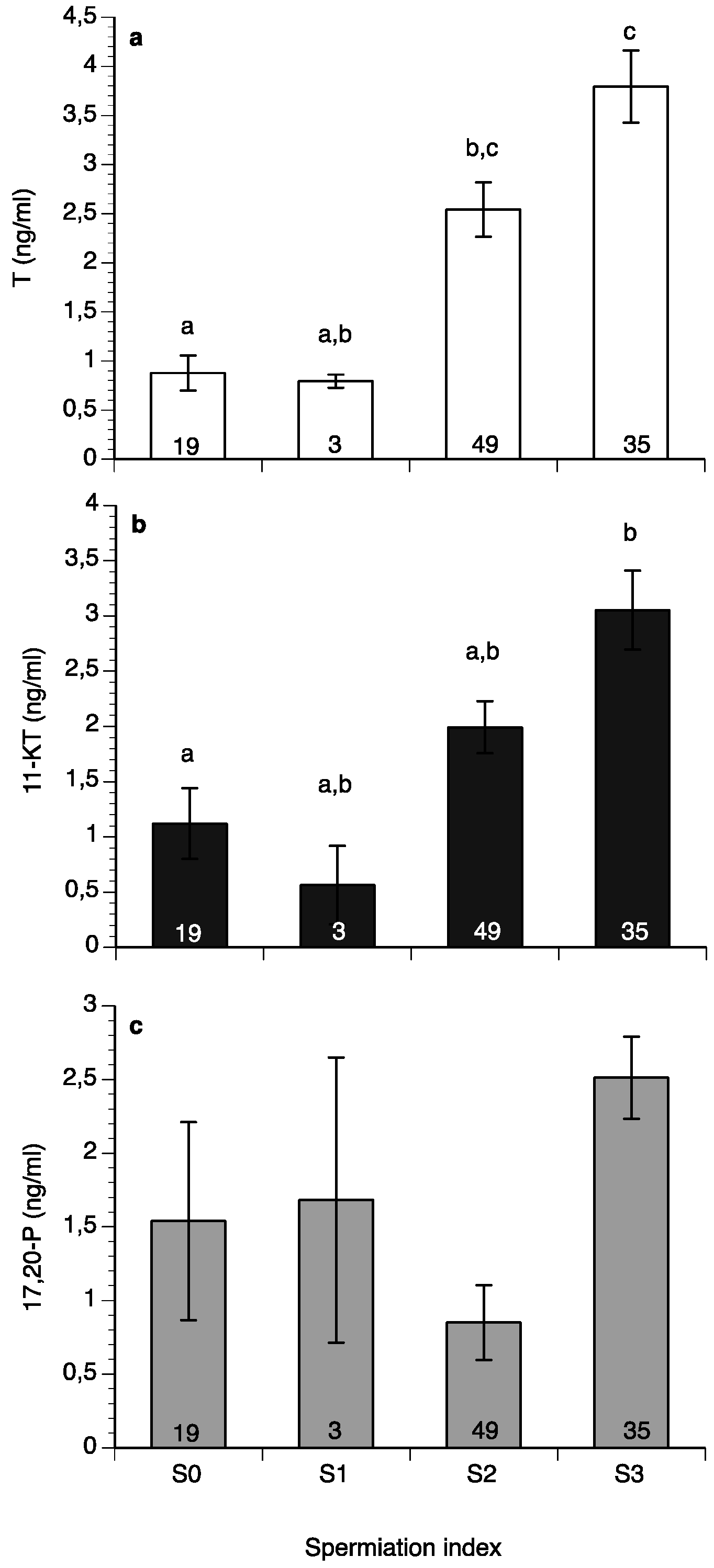
2. Results
3. Discussion
4. Materials and Methods
4.1. Broodstock Maintenance
4.2. Samplings
4.3. Histological Processing
4.4. Hormone Measurements
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Machias, A.; Somarakis, S.; Papandroulakis, N.; Spedicato, M.-T.; Suquet, M.; Lembo, G.; Divanach, P. Settlement of the wreckfish (Polyprion americanus). Mar. Biol. 2003, 142, 45–52. [Google Scholar] [CrossRef]
- Roberts, C.D. Reproductive mode in the percomorph fish genus Polyprion Oken. J. Fish Biol. 1989, 34, 1–9. [Google Scholar] [CrossRef]
- Ball, A.O.; Sedberry, G.R.; Zatcoff, M.S.; Chapman, R.W.; Carlin, J.L. Population structure of wreckfish Polyprion americanus determined with microsatelite genetic markers. Mar. Biol. 2000, 137, 1077–1090. [Google Scholar] [CrossRef]
- Peres, M.B.; Klippel, S. Reproductive biology of southwestern Atlantic wreckfish, Polyprion americanus (Teleostei: Polyprionidae). Environ. Biol. Fishes 2003, 68, 163–173. [Google Scholar] [CrossRef]
- Papandroulakis, N.; Suquet, M.; Spedicato, M.T.; Machias, A.; Fauvel, C.; Divanach, P. Feeding rates, growth performance and gametogenesis of wreckfish (Polyprion americanus) kept in captivity. Aquacult. Int. 2004, 12, 395–407. [Google Scholar] [CrossRef]
- Anderson, S.A.; Salinas, I.; Walker, S.P.; Gublin, Y.; Pether, S.; Kohn, Y.Y.; Symonds, J.E. Early development of New Zealand hapuku Polyprion oxygeneios eggs and larvae. J. Fish Biol. 2012, 80, 555–571. [Google Scholar] [CrossRef] [PubMed]
- Symonds, J.E.; Walker, S.P.; Pether, S.; Gublin, Y.; McQueen, D.; King, A.; Irvine, G.W.; Setiawan, A.N.; Forsythe, J.A.; Bruce, M. Developing yellowtail kingfish (Seriola lalandi) and hāpuku (Polyprion oxygeneios) for New Zealand aquaculture. N. Z. J. Mar. Freshw. Res. 2014, 48, 371–384. [Google Scholar] [CrossRef]
- Wylie, M.J.; Setiawan, A.N.; Irvine, G.W.; Symonds, J.E.; Elizur, A.; Dos Santos, M.; Lokman, P.M. Ovarian development of captive F1 wreckfish (hapuku) Polyprion oxygeneios under constant and varying temperature regimes–Implications for broodstock management. Gen. Comp. Endocrinol. 2018, 257, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Fauvel, C.; Suquet, M.; Sévère, A.; Mylonas, C.C.; Papandroulakis, N. Slow-release GnRHa therapy prevented atresia during vitellogenesis and induced ovulation of captive wreckfish (Polyprion americanus). Cybium 2008, 32, 191. [Google Scholar]
- Mylonas, C.C.; Zohar, Y. Controlling reproduction in aquaculture. In New Technologies in Aquaculture: Improving Production Efficiency, Quality and Environmental Management; Burnell, G., Allan, G., Eds.; Woodhead Publishing Ltd.: Cambridge, UK, 2009; pp. 109–142. [Google Scholar]
- Mylonas, C.C.; Fostier, A.; Zanuy, S. Broodstock management and hormonal manipulations of fish reproduction. Gen. Comp. Endocrinol. 2010, 165, 516–534. [Google Scholar] [CrossRef] [PubMed]
- Perez, L.; Penaranda, D.S.; Dufour, S.; Baloche, S.; Palstra, A.P.; Van Den Thillart, G.E.; Asturiano, J.F. Influence of temperature regime on endocrine parameters and vitellogenesis during experimental maturation of European eel (Anguilla anguilla) females. Gen. Comp. Endocrinol. 2011, 174, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Mylonas, C.C.; Papandroulakis, N.; Smboukis, A.; Papadaki, M.; Divanach, P. Induction of spawning of cultured greater amberjack (Seriola dumerili) using GnRHa implants. Aquaculture 2004, 237, 141–154. [Google Scholar] [CrossRef]
- Zupa, R.; Rodríguez, C.; Mylonas, C.C.; Rosenfeld, H.; Fakriadis, I.; Papadaki, M.; Pérez, J.A.; Pousis, C.; Basilone, G.; Corriero, A. Comparative study of reproductive development in wild and captive-reared greater amberjack Seriola dumerili (Risso, 1810). PLoS ONE 2017, 12, e0169645. [Google Scholar] [CrossRef] [PubMed]
- Mylonas, C.C.; Mitrizakis, N.; Papadaki, M.; Sigelaki, I. Reproduction of hatchery-produced meagre Argyrosomus regius in captivity I. Description of the annual reproductive cycle. Aquaculture 2013, 414–415, 309–317. [Google Scholar] [CrossRef]
- Micale, V.; Perdichizzi, F.; Basciano, G. Aspects of the reproductive biology of the sharpsnout seabream Diplodus puntazzo (Cetti, 1777). I. gametogenesis and gonadal cycle in captivity during the third year of life. Aquaculture 1996, 140, 281–291. [Google Scholar] [CrossRef]
- Papadaki, M.; Mazzella, D.; Santinelli, V.; Fakriadis, I.; Sigelaki, I.; Mylonas, C.C. Hermaphroditism and reproductive function of hatchery-produced sharpsnout seabream (Diplodus puntazzo) under attenuated annual thermal cycles. Aquaculture 2018, 482, 231–240. [Google Scholar] [CrossRef]
- Guzmán, J.M.; Norberg, B.; Ramos, J.; Mylonas, C.C.; Mañanos, E. Vitellogenin, steroid plasma levels and spawning performance of cultured Senegalese sole (Solea senegalensis). Gen. Comp. Endocrinol. 2008, 156, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Mylonas, C.C.; Duncan, N.J.; Asturiano, J.F. Hormonal manipulations for the enhancement of sperm production in cultured fish and evaluation of sperm quality. Aquaculture 2017, 472, 21–44. [Google Scholar] [CrossRef]
- Peñaranda, D.S.; Pérez, L.; Gallego, V.; Jover, M.; Tveiten, H.; Baloche, S.; Dufour, S.; Asturiano, J.F. Molecular and physiological study of the artificial maturation process in European eel males: From brain to testis. Gen. Comp. Endocrinol. 2010, 166, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Tsagarakis, K.; Başusta, A.; Başusta, N.; Biandolino, F.; Bostanci, D.; Buz, K.; Djodjo, Z.; Dulčić, J.; Gökoğlu, M.; Gücü, A.C.; et al. New Fisheries-related data from the Mediterranean Sea (October 2015). Mediterr. Mar. Sci. 2015, 16, 703–713. [Google Scholar] [CrossRef]
- Carbonara, P.; Costantino, G.; Giovine, G.; Lembo, G.; Spedicato, M.T.; Machias, A. Some aspects of the life history of Polyprion americanus (Schneider, 1801) along the coasts of the North Western Ionian Sea. Biol. Mar. Medit. 2003, 10, 102–112. [Google Scholar]
- Papandroulakis, N.; Mylonas, C.C.; Syggelaki, E.; Katharios, P.; Divakaran, S. First reproduction of captive-reared wreckfish (Polyprion americanus) using GnRH implants. Eur. Aquacult. 2008, 8, 15–18. [Google Scholar]
- Peleteiro, J.B.; Saavedra Penas, C.; Rial, E.P.; Álvarez-Blázquez, B.; Soares, E.C.; Perón, A.V. Diversificación de especies en acuicultura. Estudio preliminar de la reproducción de la cherna (Polyprion americanus) en cautividad. In Proceedings of the Congreso Nacional de Acuicultura, Castelldefels, Spain, 21–24 November 2011. [Google Scholar]
- Wakefield, C.B.; Newman, S.J.; Molony, B.W. Age-based demography and reproduction of hapuku, Polyprion oxygeneios, from the south coast of Western Australia: Implications for management. ICES J. Mar. Sci. 2010, 67, 1164–1174. [Google Scholar] [CrossRef]
- Martínez-Vázquez, J.M.; Pérez-Rial, E.; Peleteiro, J.B.; Linares, F.; Rodríguez, J.L.; Vilar, A.; Cal, R.; Álvarez-Blázquez, B. Description of the wreckfish (Polyprion americanus) reproductive cycle in captivity. In Proceedings of the European Aquaculture 2016, Edinburgh, UK, 20–23 September 2016. [Google Scholar]
- Duarte, C.M.; Alcaraz, M. To produce many small or few large eggs: A size-independent reproductive tactic of fish. Oecologia 1989, 80, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Mylonas, C.C.; Papadaki, M.; Pavlidis, M.; Divanach, P. Evaluation of egg production and quality in the Mediterranean red porgy (Pagrus pagrus) during two consecutive spawning seasons. Aquaculture 2004, 232, 637–649. [Google Scholar] [CrossRef]
- Cerdá, J.; Carrillo, M.; Zanuy, S.; Ramos, J.; de la Higuera, M. Influence of nutritional composition of diet on sea bass, Dicentrarchus labrax L.; reproductive performance and egg and larval quality. Aquaculture 1994, 128, 345–361. [Google Scholar] [CrossRef]
- Kennedy, J.; Geffen, A.J.; Nash, R.D.M. Maternal influences on egg and larval characteristics of plaice (Pleuronectes platessa L.). J. Sea Res. 2007, 58, 65–77. [Google Scholar] [CrossRef]
- Brown, N.P.; Shields, R.J.; Bromage, N.R. The influence of water temperature on spawning patterns and egg quality in the Atlantic halibut (Hippoglossus hippoglossus L.). Aquaculture 2006, 261, 993–1002. [Google Scholar] [CrossRef]
- Sedberry, G.R.; Andrade, C.A.P.; Carlin, J.L.; Chapman, R.W.; Luckhurst, B.E.; Manooch, C.S.; Menezes, G.; Thomsen, B.; Ulrich, G.F. Wreckfish Polyprion americanus in the North Atlantic: Fisheries, biology and management of a widely distributed and long-lived fish. Am. Fish Soc. Symp. 1999, 23, 27–50. [Google Scholar]
- Papadakis, I.E.; Kentouri, M.; Divanach, P.; Mylonas, C.C. Ontogeny of the eye of meagre (Argyrosomus regius) from hatching to juvenile and implications to commercial larval rearing. Aquaculture 2018, 484, 32–43. [Google Scholar] [CrossRef]
- Papadakis, I.; Kentouri, M.; Divanach, P.; Mylonas, C.C. Ontogeny of the digestive system of meagre Argyrosomus regius reared in a mesocosm, and quantitative changes of lipids in the liver from hatching to juveniles. Aquaculture 2013, 388–391, 76–88. [Google Scholar] [CrossRef]
- Lowerre-Barbieri, S.K.; Ganias, K.; Saborido-Rey, F.; Murua, H.; Hunter, J.R. Reproductive timing in marine fishes: Variability, temporal scales, and methods. Mar. Coast. Fish. 2011, 3, 71–91. [Google Scholar] [CrossRef]
- Gunnarsson, Á.; Hjörleifsson, E.; Thórarinsson, K.; Marteinsdóttir, G. Growth, maturity and fecundity of wolffish Anarhichas lupus L. in Icelandic waters. J. Fish Biol. 2006, 68, 1158–1176. [Google Scholar] [CrossRef]
- Gunnarsson, Á.; Hjörleifsson, E.; Thórarinsson, K.; Marteinsdóttir, G. Growth, maturity and fecundity of female spotted wolffish Anarhichas minorin Icelandic waters. J. Fish Biol. 2008, 73, 1393–1406. [Google Scholar] [CrossRef]
- Rideout, R.M.; Maddock, D.M.; Burton, M.P.M. Oogenesis and the spawning pattern in Greenland halibut from the North-west Atlantic. J. Fish Biol. 1999, 54, 196–207. [Google Scholar] [CrossRef]
- Skjæraasen, J.E.; Nash, R.D.M.; Korsbrekke, K.; Fonn, M.; Nilsen, T.; Kennedy, J.; Nedreaas, K.H.; Thorsen, A.; Witthames, P.R.; Geffen, A.J.; et al. Frequent skipped spawning in the world’s largest cod population. Proc. Natl. Acad. Sci. USA 2012, 109, 8995–8999. [Google Scholar] [CrossRef] [PubMed]
- Rideout, R.M.; Rose, G.A.; Burton, M.P.M. Skipped spawning in female iteroparous fishes. Fish Fish. 2005, 6, 50–72. [Google Scholar] [CrossRef]
- Zupa, R.; Corriero, A.; Deflorio, M.; Santamaria, N.; Spedicato, D.; Marano, C.; Losurdo, M.; Bridges, C.R.; De Metrio, G. A histological investigation of the occurence of non-reproductive female bluefin tuna Thunnus thynnus in the Mediterranean Sea. J. Fish Biol. 2009, 75, 1221–1229. [Google Scholar] [CrossRef] [PubMed]
- Tromp, J.J.; Jones, P.L.; Symonds, J.E.; Walker, S.P.; Pope, A.; Pether, S.M.J.; Afonso, L.O.B. Effects of commercial diets and temperature on the growth performance and stress response of hapuku (Polyprion oxygeneios). Aquaculture 2016, 452, 128–133. [Google Scholar] [CrossRef]
- Zohar, Y.; Muñoz-Cueto, J.A.; Elizur, A.; Kah, O. Neuroendocrinology of reproduction in teleost fish. Gen. Comp. Endocrinol. 2010, 165, 438–455. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, H.; Meiri, I.; Elizur, A. Gonadotropin regulation of oocyte development. In The Fish Oocyte: From Basic Studies to Biotechnological Applications; Babin, P.J., Cerdá, J., Lubzens, E., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2007; pp. 171–198. [Google Scholar]
- Lubzens, E.; Young, G.; Bobe, J.; Cerdá, J. Oogenesis in teleosts: How fish eggs are formed. Gen. Comp. Endocrinol. 2010, 165, 36–389. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.P.; Sumpter, J.P.; Stacey, N. The role of the maturation-inducing steroid, 17,20β-dihydroxypregn-4-en-3-one, in male fishes: A review. J. Fish Biol. 2010, 76, 183–224. [Google Scholar] [CrossRef] [PubMed]
- Nagahama, Y.; Yamashita, M. Regulation of oocyte maturation in fish. Dev. Growth Differ. 2008, 50, S195–S219. [Google Scholar] [CrossRef] [PubMed]
- Tokumoto, T.; Yamaguchi, T.; Ii, S.; Tokumoto, M. In vivo induction of oocyte maturation and ovulation in zebrafish. PLoS ONE 2011, 6, e25206. [Google Scholar] [CrossRef] [PubMed]
- Bobe, J.; Montfort, J.; Nguyen, T.; Fostier, A. Identification of new participants in the rainbow trout (Oncorhynchus mykiss) oocyte maturation and ovulation processes using cDNA microarrays. Reprod. Biol. Endocrinol. 2006, 4, 39. [Google Scholar] [CrossRef] [PubMed]
- Kjesbu, O.S.; Kryvi, H.; Norberg, B. Oocyte size and structure in relation to blood plasma steroid hormones in individually monitored, spawning Atlantic cod. J. Fish Biol. 1996, 49, 1197–1215. [Google Scholar] [CrossRef]
- Jeng, S.R.; Yueh, W.S.; Lee, Y.H.; Yen, H.F.; Chang, C.F. 17,20β,21-Trihydroxy-4-pregnen-3-one biosynthesis and 20β-hydroxysteroid dehydrogenase expression during final oocyte maturation in the protandrous yellowfin porgy, Acanthopagrus latus. Gen. Comp. Endocrinol. 2012, 176, 192–200. [Google Scholar] [CrossRef] [PubMed]
- García-López, A.; Anguis, V.; Couto, E.; Canario, A.V.M.; Cañavate, J.P.; Sarasquete, C.; Martínez-Rodríguez, G. Non-invasive assessment of reproductive status and cycle of sex steroid levels in a captive wild broodstock of Senegalese sole Solea senegalensis (Kaup). Aquaculture 2006, 254, 583–593. [Google Scholar] [CrossRef]
- Fauvel, C.; Suquet, M.; Cosson, J. Evaluation of fish sperm quality. J. Appl. Ichthyol. 2010, 26, 636–643. [Google Scholar] [CrossRef]
- Mylonas, C.C.; Papadaki, M.; Divanach, P. Seasonal changes in sperm production and quality in the red porgy Pagrus pagrus (L.). Aquacult. Res. 2003, 34, 1161–1170. [Google Scholar] [CrossRef]
- Papadaki, M.; Papadopoulou, M.; Siggelaki, I.; Mylonas, C.C. Egg and sperm production and quality of sharpsnout sea bream (Diplodus puntazzo) in captivity. Aquaculture 2008, 276, 187–197. [Google Scholar] [CrossRef]
- Zupa, R.; Fauvel, C.; Mylonas, C.C.; Pousis, C.; Santamaria, N.; Papadaki, M.; Fakriadis, I.; Cicirelli, V.; Mangano, S.; et al. Rearing in captivity affects spermatogenesis and sperm quality in greater amberjack, Seriola dumerili (Risso, 1810). J. Anim. Sci. 2017, 95, 4085–4100. [Google Scholar] [CrossRef] [PubMed]
- Suquet, M.; Cosson, J.; de la Gándara, F.; Mylonas, C.C.; Papadaki, M.; Lallemant, S.; Fauvel, C. Sperm features of captive Atlantic bluefin tuna (Thunnus thynnus). J. Appl. Ichthyol. 2010, 26, 775–778. [Google Scholar] [CrossRef]
- Schulz, R.W.; de França, L.R.; Lareyre, J.-J.; LeGac, F.; Chiarini-Garcia, H.; Nobrega, R.H.; Miura, T. Spermatogenesis in fish. Gen. Comp. Endocrinol. 2010, 165, 390–411. [Google Scholar] [CrossRef] [PubMed]
- Schulz, R.W.; Miura, T. Spermatogenesis and its endocrine regulation. Fish Physiol. Biochem. 2002, 26, 43–56. [Google Scholar] [CrossRef]
- Borg, B. Androgens in teleost fishes. Comp. Biochem. Physiol. 1994, 109, 219–245. [Google Scholar] [CrossRef]
- Dahle, R.; Taranger, G.L.; Karlsen, Ø.; Kjesbu, O.S.; Norberg, B. Gonadal development and associated changes in liver size and sexual steroids during the reproductive cycle of captive male and female Atlantic cod (Gadus morhua L.). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2003, 136, 641–653. [Google Scholar] [CrossRef]
- García-López, A.; Fernandez-Pasquier, V.; Couto, E.; Canario, A.V.; Sarasquete, C.; Martinez-Rodriguez, G. Testicular development and plasma sex steroid levels in cultured male Senegalese sole Solea senegalensis Kaup. Gen. Comp. Endocrinol. 2006, 147, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, J.M.; Ramos, J.; Mylonas, C.C.; Manaños, E. Spawning performance and plasma levels of GnRHa and sex steroids in cultured female Senegalese sole (Solea senegalensis) treated with different GnRHa-delivery systems. Aquaculture 2009, 291, 200–209. [Google Scholar] [CrossRef]
- Guzmán, J.M.; Ramos, A.G.; Mylonas, C.C.; Mananos, E. Comparative effects of human chorionic gonadotropin (hCG) and gonadotropin-releasing hormone agonist (GnRHa) treatments on the stimulation of male Senegalese sole (Solea senegalensis) reproduction. Aquaculture 2011, 316, 121–128. [Google Scholar] [CrossRef]
- Norambuena, F.; Mackenzie, S.; Bell, J.G.; Callol, A.; Estévez, A.; Duncan, N. Prostaglandin (F and E, 2- and 3-series) production and cyclooxygenase (COX-2) gene expression of wild and cultured broodstock of senegalese sole (Solea senegalensis). Gen. Comp. Endocrinol. 2012, 177, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Kohn, Y.Y.; Symonds, J.E. Evaluation of egg quality parameters as predictors of hatching success and early larval survival in hapuku (Polyprion oxygeneios). Aquaculture 2012, 342–343, 42–47. [Google Scholar] [CrossRef]
- Linares, F.; Rodriguez, J.L.; Peleteiro, J.B.; Cal, R.; Pazos, G.; Álvarez-Blázquez, B. Biochemical composition of wild wreckfish (Polyprion americanus). In Proceedings of the Aquaculture Europe 2015, Rotterdam, The Netherlands, 21–23 October 2015. [Google Scholar]
- Linares, F.; Rodriguez, J.L.; Peleteiro, J.B.; Cal, R.; Martínez, J.M.; Pazos, G.; Álvarez-Blázquez, B. Influence of broodstock nutrition of wreckfish (Polyprion americanus) on the oocytes fatty acid composition. In Proceedings of the Aquaculture Europe 2015, Edinbourg, UK, 20–23 September 2016. [Google Scholar]
- Anonymous. Guidelines for the treatment of animals in behavioural research and teaching. Anim. Behav. 1998, 55, 251–257. [Google Scholar]
- Metcalfe, J.D.; Craig, J.F. Ethical justification for the use and treatment of fishes in research: An update. J. Fish Biol. 2011, 78, 393–394. [Google Scholar] [CrossRef] [PubMed]
- EU. Directive 2010/63/EU of the European Parliament and the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes; Official Journal of the European Union L 276/33; Animal Protection: London, UK, 2010. [Google Scholar]
- Mylonas, C.C.; Cardinaletti, G.; Sigelaki, I.; Polzonetti-Magni, A. Comparative efficacy of clove oil and 2-phenoxyethanol as anesthetics in the aquaculture of European sea bass (Dicentrarchus labrax) and gilthead sea bream (Sparus aurata) at different temperatures. Aquaculture 2005, 246, 467–481. [Google Scholar] [CrossRef]
- Bennett, H.S.; Wyrick, A.D.; Lee, S.W.; McNeil, J.H. Science and art in preparing tissues embedded in plastic for light microscopy, with special reference to glycol methacrylate, glass knives and simple stains. Stain Technol. 1976, 51, 71–97. [Google Scholar] [CrossRef] [PubMed]
- Cuisset, B.; Pradelles, P.; Kime, D.E.; Kühn, E.R.; Babin, P.; Davail, S.; Le Menn, F. Enzyme immunoassay for 11-ketotestosterone using acetylcholinesterase as label: Application to the measurement of 11-ketotestosterone in plasma of Siberian sturgeon. Comp. Biochem. Physiol. 1994, 108C, 229–241. [Google Scholar]
- Nash, J.P.; Davail-Cuisset, B.; Bhattacharyya, S.; Suter, H.C.; Le Menn, F.; Kime, D.E. An enzyme linked immunosorbent assay (ELISA) for testosterone, estradiol, and 17,20β-dihydroxy-4-pregnen-3-one using acetylcholinesterase as tracer: Application to measurement of diel patterns in rainbow trout (Oncorhynchus mykiss). Fish Physiol. Biochem. 2000, 22, 355–363. [Google Scholar] [CrossRef]
- Rodríguez, L.; Begtashi, I.; Zanuy, S.; Carrillo, M. Development and validation of an enzyme immunoassay for testosterone: Effects of photoperiod on plasma testosterone levels and gonadal development in male sea bass (Dicentrarchus labrax L.) at puberty. Fish Physiol. Biochem. 2000, 23, 141–150. [Google Scholar] [CrossRef]











© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papadaki, M.; Peleteiro, J.B.; Alvarez-Blázquez, B.; Rodríguez Villanueva, J.L.; Linares, F.; Vilar, A.; Pérez Rial, E.; Lluch, N.; Fakriadis, I.; Sigelaki, I.; et al. Description of the Annual Reproductive Cycle of Wreckfish Polyprion americanus in Captivity. Fishes 2018, 3, 43. https://doi.org/10.3390/fishes3040043
Papadaki M, Peleteiro JB, Alvarez-Blázquez B, Rodríguez Villanueva JL, Linares F, Vilar A, Pérez Rial E, Lluch N, Fakriadis I, Sigelaki I, et al. Description of the Annual Reproductive Cycle of Wreckfish Polyprion americanus in Captivity. Fishes. 2018; 3(4):43. https://doi.org/10.3390/fishes3040043
Chicago/Turabian StylePapadaki, Maria, José Benito Peleteiro, Blanca Alvarez-Blázquez, José Luis Rodríguez Villanueva, Fatima Linares, Antonio Vilar, Evaristo Pérez Rial, Nuria Lluch, Ioannis Fakriadis, Irini Sigelaki, and et al. 2018. "Description of the Annual Reproductive Cycle of Wreckfish Polyprion americanus in Captivity" Fishes 3, no. 4: 43. https://doi.org/10.3390/fishes3040043
APA StylePapadaki, M., Peleteiro, J. B., Alvarez-Blázquez, B., Rodríguez Villanueva, J. L., Linares, F., Vilar, A., Pérez Rial, E., Lluch, N., Fakriadis, I., Sigelaki, I., & Mylonas, C. C. (2018). Description of the Annual Reproductive Cycle of Wreckfish Polyprion americanus in Captivity. Fishes, 3(4), 43. https://doi.org/10.3390/fishes3040043