

The Effects of Early Temperature and Live Feeds on the Development of White Muscle in Greater Amberjack (Seriola dumerili)

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
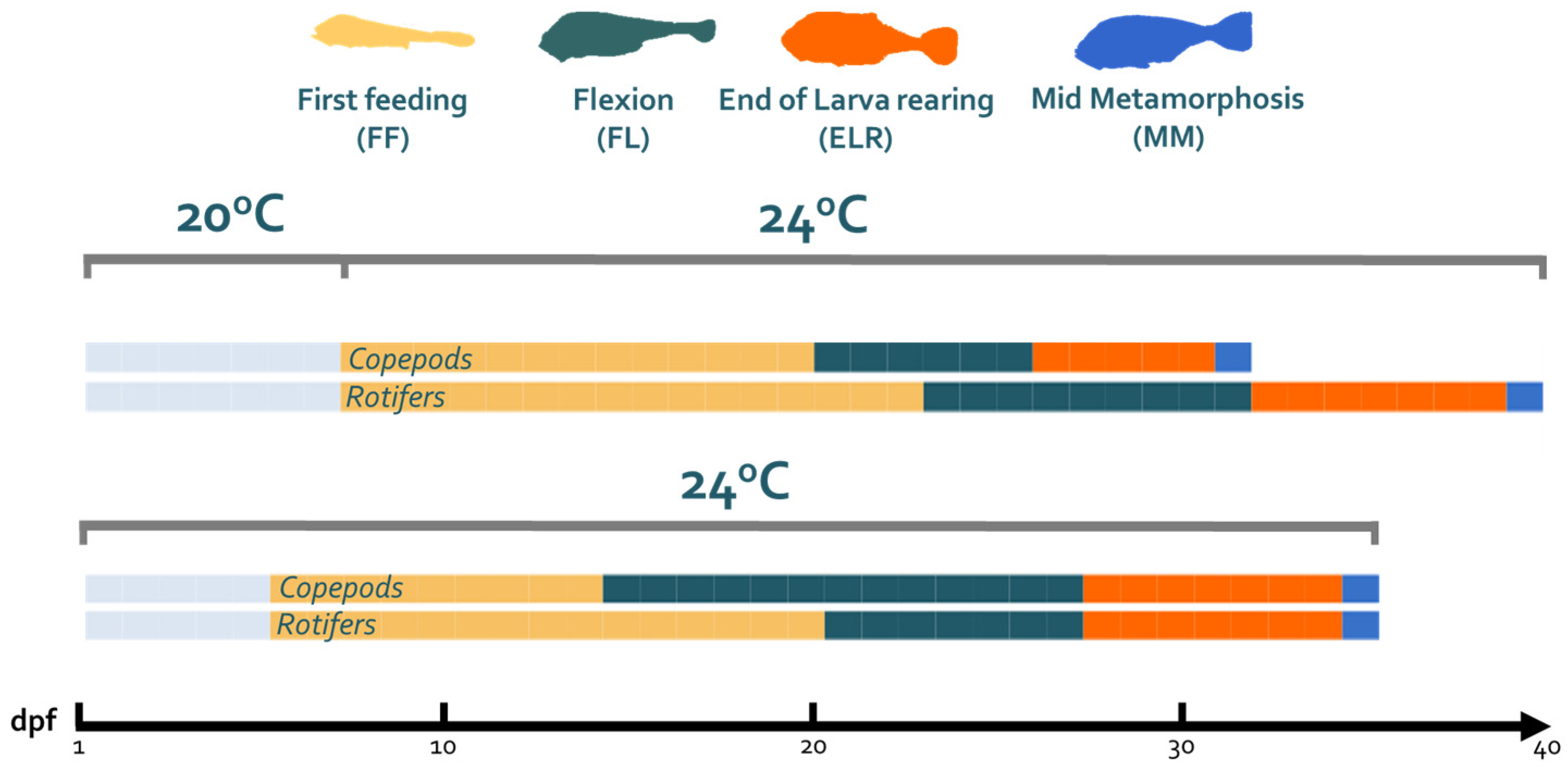
2.2. Experimental Rearing
2.3. Quantitative Histology
2.4. RNA Extraction and Reverse Transcription
2.5. Relative Quantification of Gene Expression Determined Using Real-Time Quantitative Polymerase Chain Reaction Analysis
2.6. Statistical Analysis
3. Results
3.1. Temperature and Live Feed Effect on Larvae Developmental Pace
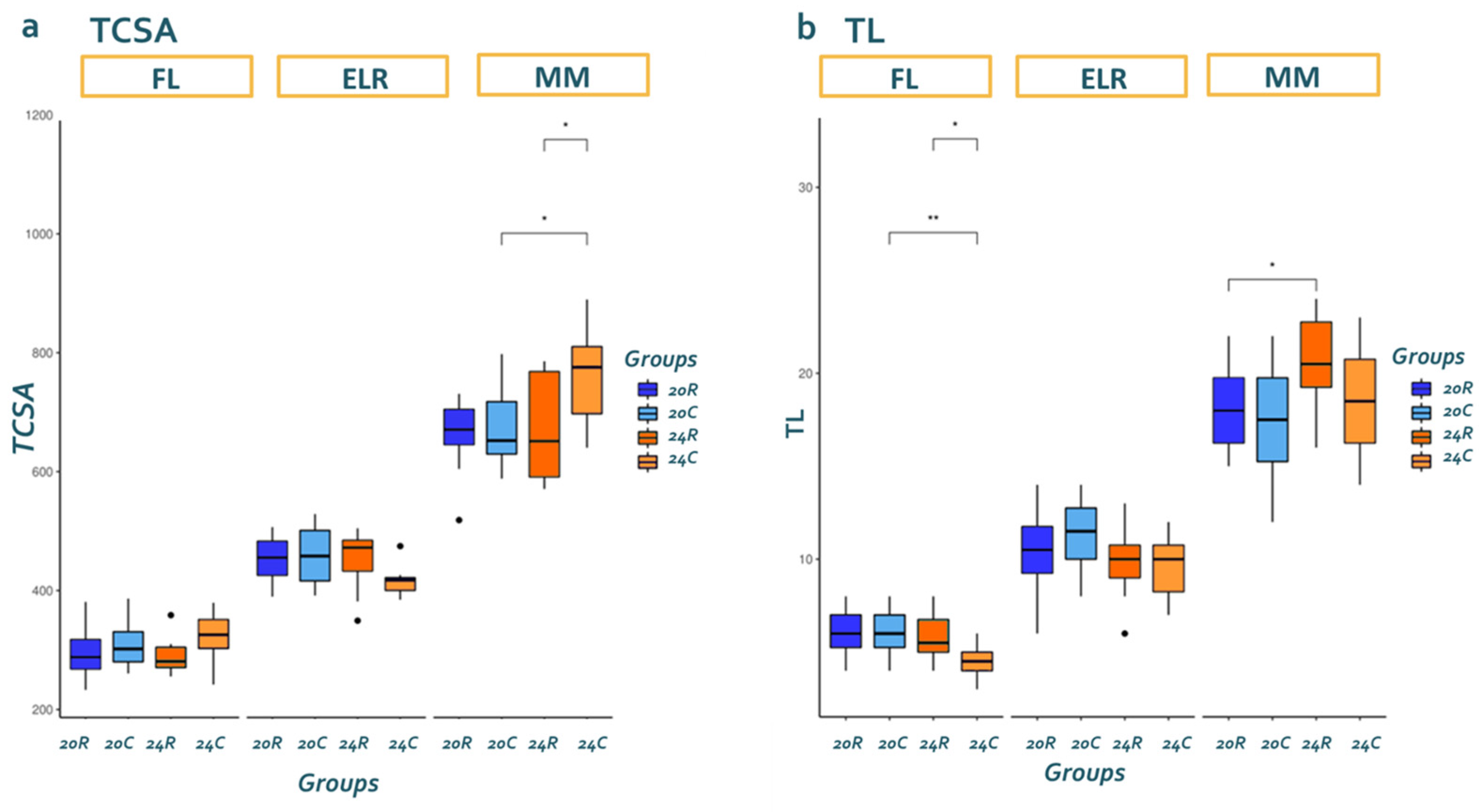
3.2. Stereological Analysis
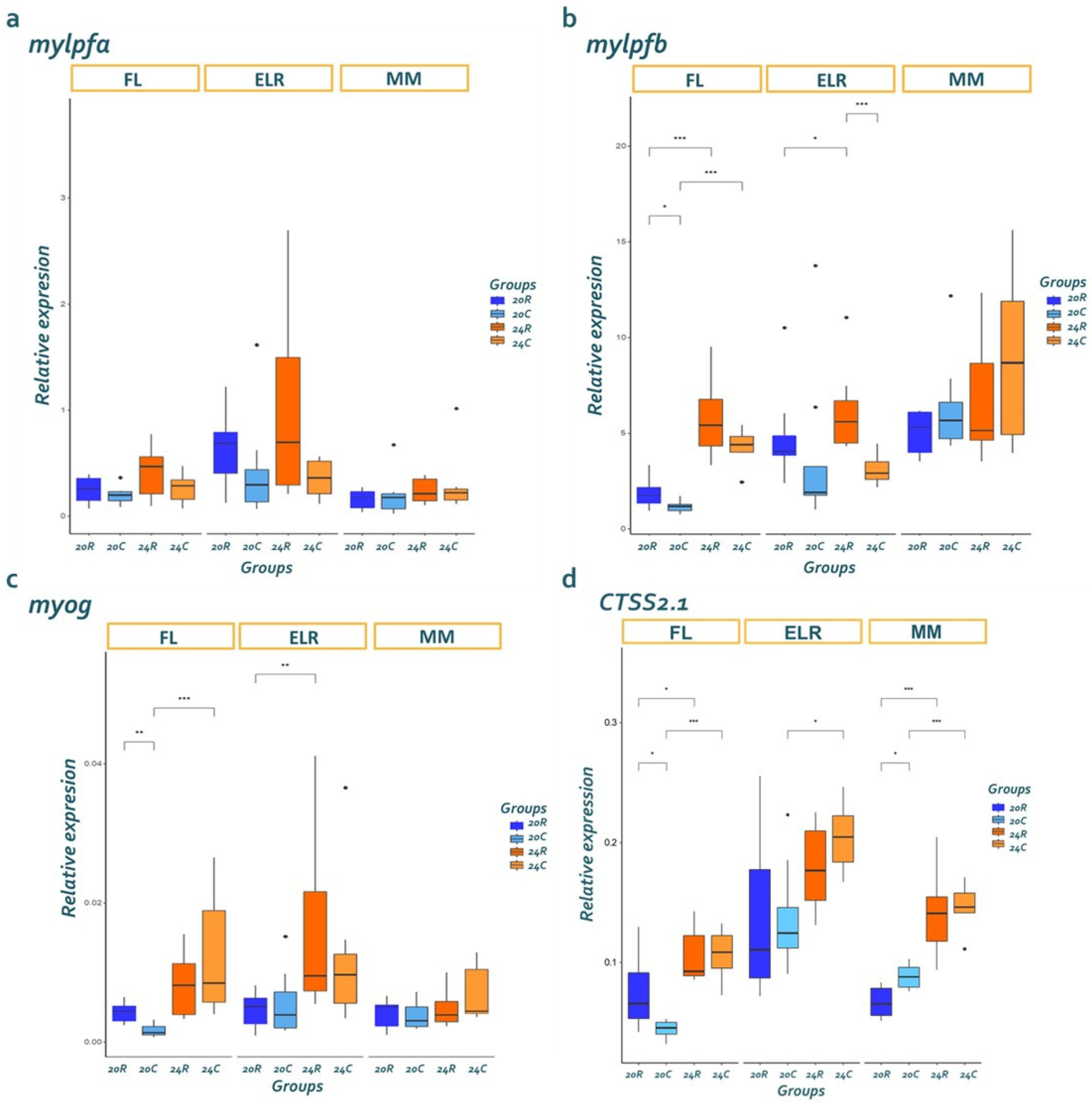
3.3. Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
 |
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Nakada, M. Yellowtail Culture Development and Solutions for the Future. Rev. Fish. Sci. 2010, 10, 559–575. [Google Scholar] [CrossRef]
- Fernández-Montero, A.; Caballero, M.J.; Torrecillas, S.; Tuset, V.M.; Lombarte, A.; Ginés, R.R.; Izquierdo, M.; Robaina, L.; Montero, D. Effect of Temperature on Growth Performance of Greater Amberjack (Seriola dumerili Risso 1810) Juveniles. Aquac. Res. 2018, 49, 908–918. [Google Scholar] [CrossRef]
- Fakriadis, I.; Sigelaki, I.; Papadaki, M.; Papandroulakis, N.; Raftopoulos, A.; Tsakoniti, K.; Mylonas, C.C. Control of Reproduction of Greater Amberjack Seriola dumerili Reared in Aquaculture Facilities. Aquaculture 2020, 519, 734880. [Google Scholar] [CrossRef]
- Pérez, J.A.; Papadakis, I.E.; Papandroulakis, N.; Cruces, L.; Cotou, E.; Gisbert, E.; Lorenzo, A.; Mylonas, C.C.; Rodríguez, C. The Ontogeny of Greater Amberjack Digestive and Antioxidant Defence Systems under Different Rearing Conditions: A Histological and Enzymatic Approach. Aquac. Nutr. 2020, 26, 1908–1925. [Google Scholar] [CrossRef]
- Moran, D. Size Heterogeneity, Growth Potential and Aggression in Juvenile Yellowtail Kingfish (Seriola lalandi Valenciennes). Aquac. Res. 2007, 38, 1254–1264. [Google Scholar] [CrossRef]
- Kestemont, P.; Jourdan, S.; Houbart, M.; Mélard, C.; Paspatis, M.; Fontaine, P.; Cuvier, A.; Kentouri, M.; Baras, E. Size Heterogeneity, Cannibalism and Competition in Cultured Predatory Fish Larvae: Biotic and Abiotic Influences. Aquaculture 2003, 227, 333–356. [Google Scholar] [CrossRef]
- Papandroulakis, N.; Mylonas, C.C.; Maingot, E.; Divanach, P. First Results of Greater Amberjack (Seriola dumerili) Larval Rearing in Mesocosm. Aquaculture 2005, 250, 155–161. [Google Scholar] [CrossRef]
- Johnston, I.A. Environment and Plasticity of Myogenesis in Teleost Fish. J. Exp. Biol. 2006, 209, 2249–2264. [Google Scholar] [CrossRef] [PubMed]
- Alami-Durante, H.; Wrutniak-Cabello, C.; Kaushik, S.J.; Médale, F. Skeletal Muscle Cellularity and Expression of Myogenic Regulatory Factors and Myosin Heavy Chains in Rainbow Trout (Oncorhynchus mykiss): Effects of Changes in Dietary Plant Protein Sources and Amino Acid Profiles. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2010, 156, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Rowlerson, A.; Mascarello, F.; Radaelli, G.; Veggetti, A. Differentiation and Growth of Muscle in the Fish Sparus aurata (L): II. Hyperplastic and Hypertrophic Growth of Lateral Muscle from Hatching to Adult. J. Muscle Res. Cell Motil. 1995, 16, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Pitkänen, T.I.; Xie, S.Q.; Krasnov, A.; Mason, P.S.; Mölsä, H.; Stickland, N.C. Changes in Tissue Cellularity Are Associated with Growth Enhancement in Genetically Modified Arctic Char (Salvelinus alpinus L.) Carrying Recombinant Growth Hormone Gene. Mar. Biotechnol. 2001, 3, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Alami-Durante, H.; Fauconneau, B.; Rouel, M.; Escaffre, A.M.; Bergot, P. Growth and Multiplication of White Skeletal Muscle Fibres in Carp Larvae in Relation to Somatic Growth Rate. J. Fish Biol. 1997, 50, 1285–1302. [Google Scholar] [CrossRef]
- Rauner, M.; Föger-Samwald, U.; Kurz, M.F.; Brünner-Kubath, C.; Schamall, D.; Kapfenberger, A.; Varga, P.; Kudlacek, S.; Wutzl, A.; Höger, H.; et al. Cathepsin S Controls Adipocytic and Osteoblastic Differentiation, Bone Turnover, and Bone Microarchitecture. Bone 2014, 64, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Pacheco-Carlón, N.; Guerrero-Tortolero, D.A.; Cervantes-Montoya, L.B.; Racotta, I.S.; Campos-Ramos, R. The Effects of Constant and Oscillating Temperature on Embryonic Development and Early Larval Morphology in Longfin Yellowtail (Seriola rivoliana Valenciennes). Aquac. Res. 2021, 52, 77–93. [Google Scholar] [CrossRef]
- Pacheco-Carlón, N.; Salgado-García, R.L.; Guerrero-Tortolero, D.A.; Kraffe, E.; Campos-Ramos, R.; Racotta, I.S. Biochemical Composition and Adenylate Energy Charge Shifts in Longfin Yellowtail (Seriola rivoliana) Embryos during Development under Different Temperatures. J. Therm. Biol. 2023, 112, 103470. [Google Scholar] [CrossRef] [PubMed]
- Viader-Guerrero, M.; Guzmán-Villanueva, L.T.; Spanopoulos-Zarco, M.; Estrada-Godínez, J.A.; Maldonado-García, D.; Gracia-López, V.; Omont, A.; Maldonado-García, M. Effects of Temperature on Hatching Rate and Early Larval Development of Longfin Yellowtail Seriola rivoliana. Aquac. Rep. 2021, 21, 100843. [Google Scholar] [CrossRef]
- Campos-Ramos, R.; Vázquez-Islas, G.; Calixto-Heredia, L.M.; Guerrero-Tortolero, D.A. Gene Expression in the Hypothalamic-Pituitary-Thyroid Axis in Seriola rivoliana Early Larvae Development at Different Temperatures. Gen. Comp. Endocrinol. 2024, 358, 114615. [Google Scholar] [CrossRef] [PubMed]
- Watson, S.A.; Allan, B.J.M.; McQueen, D.E.; Nicol, S.; Parsons, D.M.; Pether, S.M.J.; Pope, S.; Setiawan, A.N.; Smith, N.; Wilson, C.; et al. Ocean Warming Has a Greater Effect than Acidification on the Early Life History Development and Swimming Performance of a Large Circumglobal Pelagic Fish. Glob. Chang. Biol. 2018, 24, 4368–4385. [Google Scholar] [CrossRef] [PubMed]
- Lomartire, S.; Marques, J.C.; Gonçalves, A.M.M. The Key Role of Zooplankton in Ecosystem Services: A Perspective of Interaction between Zooplankton and Fish Recruitment. Ecol. Indic. 2021, 129, 107867. [Google Scholar] [CrossRef]
- Conceição, L.E.C.; Yúfera, M.; Makridis, P.; Morais, S.; Dinis, M.T. Live Feeds for Early Stages of Fish Rearing. Aquac. Res. 2010, 41, 613–640. [Google Scholar] [CrossRef]
- Karlsen, Ø.; van der Meeren, T.; Rønnestad, I.; Mangor-Jensen, A.; Galloway, T.F.; Kjørsvik, E.; Hamre, K. Copepods Enhance Nutritional Status, Growth and Development in Atlantic Cod (Gadus morhua L.) Larvae—Can We Identify the Underlying Factors? PeerJ 2015, 2015, e902. [Google Scholar] [CrossRef] [PubMed]
- Miki, T.; Nakatsukasa, H.; Takahashi, N.; Murata, O.; Ishibashi, Y. Aggressive Behaviour and Cannibalism in Greater Amberjack, Seriola Dumerili : Effects of Stocking Density, Feeding Conditions and Size Differences. Aquac. Res. 2011, 42, 1339–1349. [Google Scholar] [CrossRef]
- Ma, Z. Food Ingestion, Prey Selectivity, Feeding Incidence, and Performance of Yellowtail Kingfish Seriola lalandi Larvae under Constant and Varying Temperatures. Aquac. Int. 2014, 22, 1317–1330. [Google Scholar] [CrossRef]
- Georgiou, S.; Alami-Durante, H.; Power, D.M.; Sarropoulou, E.; Mamuris, Z.; Moutou, K.A. Transient Up- and down-Regulation of Expression of Myosin Light Chain 2 and Myostatin MRNA Mark the Changes from Stratified Hyperplasia to Muscle Fiber Hypertrophy in Larvae of Gilthead Sea Bream (Sparus aurata L.). Cell Tissue Res. 2016, 363, 541–554. [Google Scholar] [CrossRef] [PubMed]
- Alami-Durante, H.; Olive, N.; Rouel, M. Early Thermal History Significantly Affects the Seasonal Hyperplastic Process Occurring in the Myotomal White Muscle of Dicentrarchus labrax Juveniles. Cell Tissue Res. 2007, 327, 553–570. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, E45. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, research0034-1. [Google Scholar] [CrossRef] [PubMed]
- Čikoš, Š.; Bukovská, A.; Koppel, J. Relative Quantification of MRNA: Comparison of Methods Currently Used for Real-Time PCR Data Analysis. BMC Mol. Biol. 2007, 8, 113. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Tortolero, D.A.; Vázquez-Islas, G.; Campos-Ramos, R. A Transcriptome Insight During Early Fish Larval Development Followed by Starvation in Seriola rivoliana. Mar. Biotechnol. 2021, 23, 749–765. [Google Scholar] [CrossRef] [PubMed]
- Koganti, P.; Yao, J.; Cleveland, B.M. Molecular Mechanisms Regulating Muscle Plasticity in Fish. Anim. 2021, 11, 61. [Google Scholar] [CrossRef] [PubMed]
- Nemova, N.N.; Kantserova, N.P.; Lysenko, L.A. The Traits of Protein Metabolism in the Skeletal Muscle of Teleost Fish. J. Evol. Biochem. Physiol. 2021, 57, 626–645. [Google Scholar] [CrossRef]
- Johnston, I.A.; Bower, N.I.; Macqueen, D.J. Growth and the Regulation of Myotomal Muscle Mass in Teleost Fish. J. Exp. Biol. 2011, 214, 1617–1628. [Google Scholar] [CrossRef] [PubMed]
- Ayala, M.D.; López-Albors, O.; Gil, F.; Latorre, R.; Vázquez, J.M.; García-Alcázar, A.; Abellán, E.; Ramírez, G.; Moreno, F. Temperature Effect on Muscle Growth of the Axial Musculature of the Sea Bass (Dicentrarchus labrax L.). Anat. Histol. Embryol. 2000, 29, 235–242. [Google Scholar] [CrossRef] [PubMed]
- López-Albors, O.; Ayala, M.D.; Gil, F.; García-Alcázar, A.; Abellán, E.; Latorre, R.; Ramírez-Zarzosa, G.; Vázquez, J.M. Early Temperature Effects on Muscle Growth Dynamics and Histochemical Profile of Muscle Fibres of Sea Bass Dicentrarchus labrax L., during Larval and Juvenile Stages. Aquaculture 2003, 220, 385–406. [Google Scholar] [CrossRef]
- Stickland, N.C.; White, R.N.; Mescall, P.E.; Crook, A.R.; Thorpe, J.E. The Effect of Temperature on Myogenesis in Embryonic Development of the Atlantic Salmon (Salmo salar L.). Anat. Embryol. 1988, 178, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Calvo, J.; Johnston, I.A. Influence of Rearing Temperature on the Distribution of Muscle Fibre Types in the Turbot Scophthalmus Maximus at Metamorphosis. J. Exp. Mar. Bio. Ecol. 1992, 161, 45–55. [Google Scholar] [CrossRef]
- Brooks, S.; Johnston, I.A. Influence of Development and Rearing Temperature on the Distribution, Ultrastructure and Myosin Sub-Unit Composition of Myotomal Muscle-Fibre Types in the Plaice Pleuronectes Platessa. Mar. Biol. 1993, 117, 501–513. [Google Scholar] [CrossRef]
- Vieira, V.I.A.; Johnston, I.A. Influence of Temperature on Muscle-Fibre Development in Larvae of the Herring Clupea Harengus. Mar. Biol. 1992, 112, 333–341. [Google Scholar] [CrossRef]
- Hall, T.E.; Johnston, I.A. Temperature and Developmental Plasticity during Embryogenesis in the Atlantic Cod Gadus morhua L. Mar. Biol. 2003, 142, 833–840. [Google Scholar] [CrossRef]
- Richardson, B.E.; Nowak, S.J.; Baylies, M.K. Myoblast Fusion in Fly and Vertebrates: New Genes, New Processes and New Perspectives. Traffic 2008, 9, 1050–1059. [Google Scholar] [CrossRef] [PubMed]
- García-Pérez, I.; Molsosa-Solanas, A.; Perelló-Amorós, M.; Sarropoulou, E.; Blasco, J.; Gutiérrez, J.; de la Serrana, D.G. The Emerging Role of Long Non-Coding RNAs in Development and Function of Gilthead Sea Bream (Sparus aurata) Fast Skeletal Muscle. Cells 2022, 11, 428. [Google Scholar] [CrossRef] [PubMed]
- Manchanda, M.; Fatima, N.; Chauhan, S.S. Physiological and Pathological Functions of Cysteine Cathepsins. In Proteases in Physiology and Pathology; Springer: Singapore, 2017; pp. 217–256. [Google Scholar] [CrossRef]
- Baeverfjord, G.; Prabhu, P.A.J.; Fjelldal, P.G.; Albrektsen, S.; Hatlen, B.; Denstadli, V.; Ytteborg, E.; Takle, H.; Lock, E.J.; Berntssen, M.H.G.; et al. Mineral Nutrition and Bone Health in Salmonids. Rev. Aquac. 2019, 11, 740–765. [Google Scholar] [CrossRef]
- Suarez-Bregua, P.; Pirraco, R.P.; Hernández-Urcera, J.; Reis, R.L.; Rotllant, J. Impact of Dietary Phosphorus on Turbot Bone Mineral Density and Content. Aquac. Nutr. 2021, 27, 1128–1134. [Google Scholar] [CrossRef]
- Fraser, T.W.K.; Witten, P.E.; Albrektsen, S.; Breck, O.; Fontanillas, R.; Nankervis, L.; Thomsen, T.H.; Koppe, W.; Sambraus, F.; Fjelldal, P.G. Phosphorus Nutrition in Farmed Atlantic Salmon (Salmo salar): Life Stage and Temperature Effects on Bone Pathologies. Aquaculture 2019, 511, 734246. [Google Scholar] [CrossRef]
- Cotti, S.; Huysseune, A.; Koppe, W.; Rücklin, M.; Marone, F.; Wölfel, E.M.; Fiedler, I.A.K.; Busse, B.; Forlino, A.; Witten, P.E. More Bone with Less Minerals? The Effects of Dietary Phosphorus on the Post-Cranial Skeleton in Zebrafish. Int. J. Mol. Sci. 2020, 21, 5429. [Google Scholar] [CrossRef] [PubMed]
- Witten, P.E.; Fjelldal, P.G.; Huysseune, A.; McGurk, C.; Obach, A.; Owen, M.A.G. Bone without Minerals and Its Secondary Mineralization in Atlantic Salmon (Salmo Salar): The Recovery from Phosphorus Deficiency. J. Exp. Biol. 2019, 222, jeb188763. [Google Scholar] [CrossRef] [PubMed]
- Malzahn, A.M.; Ribičić, D.; Hansen, B.H.; Sarno, A.; Kjørsvik, E.; Aase, A.S.N.; Musialak, L.A.; García-Calvo, L.; Hagemann, A. First Feed Matters: The First Diet of Larval Fish Programmes Growth, Survival, and Metabolism of Larval Ballan Wrasse (Labrus bergylta). Aquaculture 2022, 561, 738586. [Google Scholar] [CrossRef]
- Tseng, Y.; Eryalçın, K.M.; Sivagurunathan, U.; Domínguez, D.; Hernández-Cruz, C.M.; Boglione, C.; Philip, A.J.P.; Izquierdo, M. Effects of the Dietary Supplementation of Copper on Growth, Oxidative Stress, Fatty Acid Profile and Skeletal Development in Gilthead Seabream (Sparus aurata) Larvae. Aquaculture 2023, 568, 739319. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Feeding Scheme | ||
|---|---|---|
| Diet Type | Rotifers (R) | Rotifers + Copepods (C) |
| Rotifers | 2–17 dph | 2–17 dph |
| Copepods | - | 2–8 dph |
| AF Artemia | 9–19 dph | 9–19 dph |
| EG Artemia | 12–35 dph | 12–35 dph |
| Microdiet | 13 dph onwards | 13 dph onwards |
| Gene | Gene ID | Forward Primer | Reverse Primer | Product Size | Efficiency (%) |
|---|---|---|---|---|---|
| elf1 | ENSSDUG00000008198 | CACCCCAAGTTCGTCAAGTC | GACCAAGGGGAGCGTAGTTG | 100 bp | 103.7 |
| rps18 | ENSSDUG00000012209 | ATCGCCTTCGCCATCACTG | GCTCCCCAGCCCTCTTGTTG | 100 bp | 99 |
| rpl13 | ENSSDUG00000001262 | TTGCCCCACAAAACCAAGAG | GGACAACCATGCGCTTCCTC | 100 bp | 101.6 |
| actb | ENSSDUG00000018712 | CGTGGCTACTCCTTCACCAC | TACCCATCTCCTGCTCGAAG | 100 bp | 97.2 |
| fau | ENSSDUG00000009474 | TGAGGGGACAGACACCCAAG | ACGTTCACAAAGCGCCTGTT | 100 bp | 99.1 |
| mylpfa | ENSSDUG00000022124 | TCCTCTTCTTCCTCCCTGCT | CCGTCTCCTGTGTCATTGTG | 95 bp | 97.6 |
| mylpfb | ENSSDUG00000022903 | CAAACCCATACCCGCTCCAC | CTGGCAAGGGGAAAGAAAGG | 99 bp | 96.9 |
| myog | ENSSDUG00000001886 | GAGGAGCACCCTGATGAACC | TCGCTTGACGACAACACTCT | 186 bp | 99.3 |
| ctss2.1 | ENSSDUG00000002074 | TTTCAGTGGGGATTGACGCC | GCCCTGGTCTCCGAAAGTAG | 182 bp | 98.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angelakopoulos, R.; Tsipourlianos, A.; Fytsili, A.E.; Mitrizakis, N.; Giannoulis, T.; Papandroulakis, N.; Moutou, K.A. The Effects of Early Temperature and Live Feeds on the Development of White Muscle in Greater Amberjack (Seriola dumerili). Fishes 2025, 10, 360. https://doi.org/10.3390/fishes10070360
Angelakopoulos R, Tsipourlianos A, Fytsili AE, Mitrizakis N, Giannoulis T, Papandroulakis N, Moutou KA. The Effects of Early Temperature and Live Feeds on the Development of White Muscle in Greater Amberjack (Seriola dumerili). Fishes. 2025; 10(7):360. https://doi.org/10.3390/fishes10070360
Chicago/Turabian StyleAngelakopoulos, Rafael, Andreas Tsipourlianos, Alexia E. Fytsili, Nikolaos Mitrizakis, Themistoklis Giannoulis, Nikos Papandroulakis, and Katerina A. Moutou. 2025. "The Effects of Early Temperature and Live Feeds on the Development of White Muscle in Greater Amberjack (Seriola dumerili)" Fishes 10, no. 7: 360. https://doi.org/10.3390/fishes10070360
APA StyleAngelakopoulos, R., Tsipourlianos, A., Fytsili, A. E., Mitrizakis, N., Giannoulis, T., Papandroulakis, N., & Moutou, K. A. (2025). The Effects of Early Temperature and Live Feeds on the Development of White Muscle in Greater Amberjack (Seriola dumerili). Fishes, 10(7), 360. https://doi.org/10.3390/fishes10070360