4.3. Discussion on Path Analysis
In the natural survival environments of fish, path analysis is widely applied to analyze the impacts of morphological traits at different growth stages on body weight. The results of path analysis in this study indicate that
P. schrenkii body length has the strongest direct effect on body weight, while body width, body height, and interorbital width exert indirect effects on body weight. The results obtained concur with findings reported in other fish species. For juvenile
Takifugu obscurus, seven morphological traits were retained: total length, snout length, mouth cleft, interorbital width, trunk length, body height, and caudal peduncle length [
8]. For 6-month-old
Hexagrammos agrammus, five traits with extremely significant differences in partial regression coefficients (
p < 0.01)—body length, body width, total length, head length, and body height—were selected as independent variables [
21]. For 4-month-old
Opsariichthys bidens, five morphological traits, including total length, body length, body height, body width, and head length, are the main factors influencing body mass [
32]. For juvenile
Lateolabrax maculatus, four morphological traits—total length, body height, eye diameter, and body width—are the most critical morphological traits affecting body mass [
33].
The establishment of a multiple regression equation can quantify the relationship between independent and dependent variables. In this study, the multiple linear regression analysis of 13 morphological traits all reached extremely significant levels in the significance test of coefficients. In this experiment, the method of multiple linear regression equations was used. As independent variables were introduced, the correlation coefficient of the equation increased. Subsequently, a four-variable linear regression equation for the
P. schrenkii was established, quantifying the comprehensive relationship between morphological traits and body weight. For the
P. schrenkii, none of the direct effects of morphological traits on body weight were negative. In terms of indirect effects, the order was as follows: body height (0.438) > body width (0.430) > interocular distance (0.157), all acting indirectly on body weight through body length. The sum of the indirect effects of body height, body width, and interocular distance was greater than the direct effects of each morphological trait on body mass. For 4-month-old
Maccullochella peelii juveniles, the indirect effect of caudal peduncle height on body weight through body length was the largest. Moreover, the combined indirect effects of caudal peduncle height and eye diameter were greater than their direct effects on body weight, indicating that these two traits indirectly affect body weight through body length. Therefore, in breeding selection, body length should be considered a key trait, while caudal peduncle height and eye diameter should be considered auxiliary traits [
34]. Previous studies have obtained similar results; for example, for 4-month-old
Rachycentron canadum juveniles, there were differences in the direct effects of body length, head length, and post-eye head length on body mass. The direct effects of these three traits were all smaller than their indirect effects through other morphological traits, indicating that multiple morphological traits collectively influence body mass. Analysis of the indirect effects revealed that the indirect effect of head length on body mass through body length was relatively large, with an indirect path coefficient of 0.381; similarly, the indirect effect of post-eye head length on body weight through body length was also significant, with an indirect path coefficient of 0.360 [
35]. For
Leiocassis longirostris, the direct effect of total length was the largest (
p < 0.01), reaching 0.322. Additionally, the direct effects of total length and head width in farmed long-snouted catfish were extremely significant (
p < 0.01), while the direct effects of body length, snout length, and caudal peduncle height were significant (
p < 0.05). Among these, the direct effects of total length, body length, head width, snout length, and caudal peduncle height were all smaller than their indirect effects [
36].
Among the four morphological traits, body length had the highest individual determination coefficient (0.388), indicating the most significant impact on the body weight of P. schrenkii. Additionally, the combined determination coefficient of body length and body width was the largest (0.182), demonstrating the strongest interactive effect between these two traits in influencing body mass, with a joint contribution significantly higher than other trait combinations. The residual determination coefficient of P. schrenkii was 0.059, corresponding to a residual factor of 0.243, indicating a low degree of unexplained variation in the model.
These results are consistent: the total determination coefficient of five morphological traits for body mass in 2-month-old
Amphiprion ocellaris reached 0.962, among which the sum of the individual and combined determination coefficients of body height was 0.779, confirming that body height is the most critical factor affecting body weight [
37]. In 13-month-old farmed
Schizothorax prenanti, the individual determination coefficients of five morphological traits—body length, body height, body width, snout length, and caudal peduncle length—were 0.369, 0.103, 0.008, 0.005, and 0.005, respectively [
38]. The sum of individual determination coefficients and pairwise combined determination coefficients was 0.925, indicating that these five traits are the main factors influencing body weight. Notably, the combined determination coefficient of body length and body height was the largest (0.337), while the combined determination coefficients of caudal peduncle length with body length, body height, and body width were negative, suggesting extremely weak negative effects on body mass. In 6-month-old
Epinephelus akaara, the combined determination coefficient of total length and head length (0.4137) had the most prominent impact on body mass. The sum of determination coefficients of all phenotypic traits was 0.9241, and the determination coefficient of the error term was only 0.0759, indicating that the model had effectively captured the main phenotypic traits affecting body weight with a low error contribution [
39].
To elucidate the univariate relationships between body weight and major morphological traits in
P. schrenkii, we conducted curve model fitting for body length, caudal peduncle height, eye diameter, and other traits with body weight [
40]. Our results demonstrated that body length exhibited a linear relationship with body weight (
R2 = 0.921,
p < 0.01), whereas body width, body height, and interocular distance showed quadratic relationships with body mass
(R2 values were 0.675, 0.711,
p < 0.01 for body width and body height; interocular distance
R2 = 0.091,
p = 0.091). These findings suggest that body length, body width, and body height have substantial predictive power for body mass in
P. schrenkii. Comparisons with previous studies on other fish species revealed notable differences in the relationships between morphological traits and body mass. In 5-month-old
Epinephelus fuscoguttatus, six curve models fitted for four morphological traits and body weight all reached highly significant levels (
p < 0.01). Specifically, body height and body width exhibited the best fit with body weight using exponential functions, interocular distance showed the optimal fit with a power function, and caudal peduncle height had the highest fit with a linear function [
41]. These results highlight that different growth patterns of morphological traits require distinct curve models for accurate representation. For juvenile barramundi
Lates calcarifer, eight curve models fitted for five morphological traits (X4, X5, X6, X11, X12) and body weight were all significant, validating the effectiveness of these models in capturing the relationships between morphological traits and body mass [
42]. In Murray cod larvae
Maccullochella peelii, body length, caudal peduncle height, and eye diameter were best fitted with quadratic functions. However, the goodness of fit (
R2 < 0.85) for caudal peduncle height and eye diameter indicated limited explanatory power when considered individually, suggesting they are more suitable as auxiliary indicators [
34]. In contrast, body length had a higher goodness of fit (
R2 ≥ 0.85), making it a key morphological indicator for predicting body mass.
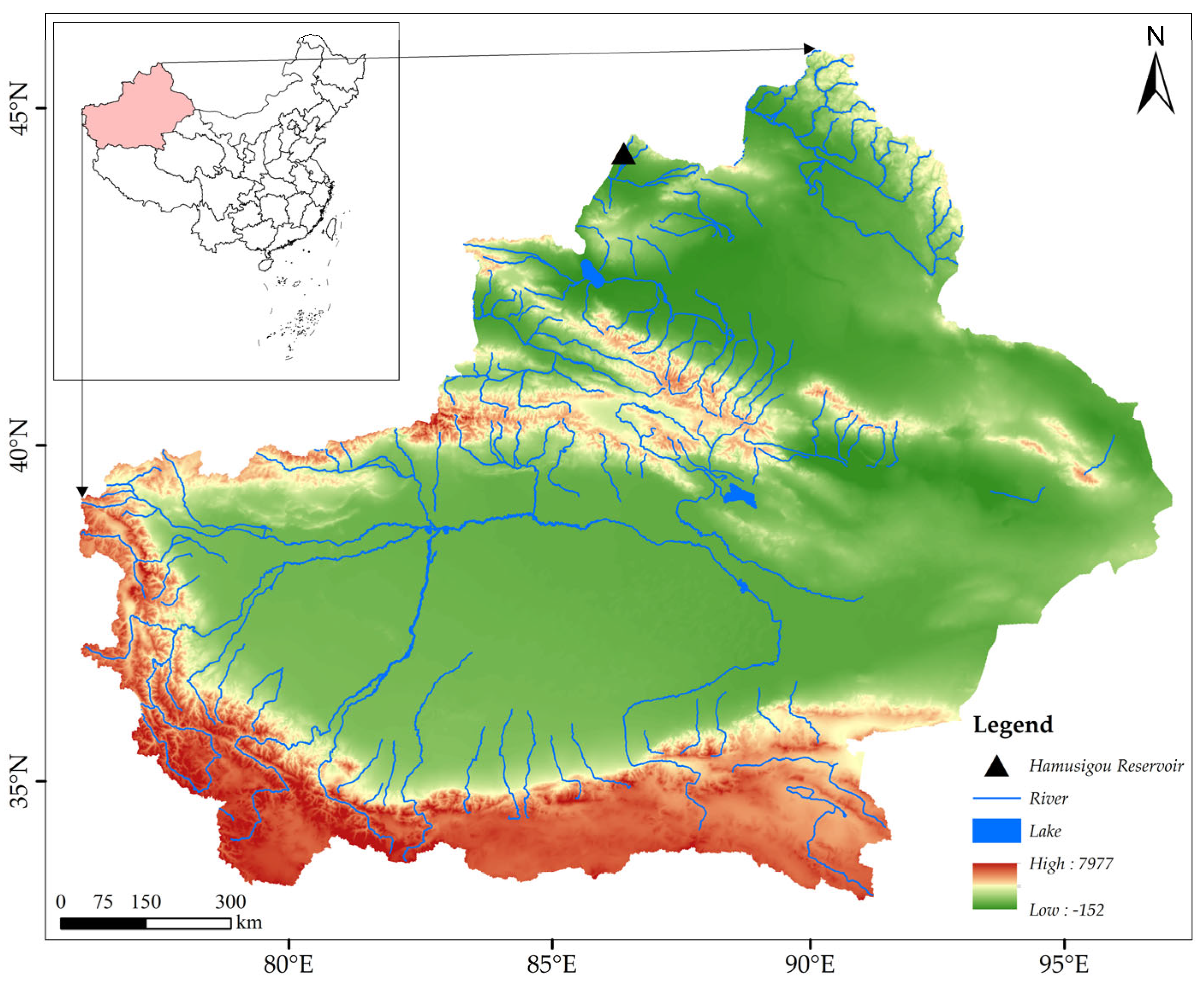
It is important to note that body mass in P. schrenkii is influenced not only by morphological traits but also by environmental factors such as water temperature, dissolved oxygen, food availability, hydrological conditions, and human activities. Future studies should consider these multidimensional variables to provide a more comprehensive understanding of the factors influencing body mass. Additionally, different fish species within the same water body exhibit distinct growth characteristics. Therefore, we recommend further correlation studies on other fish species in the Hamsigou Reservoir in Xinjiang to elucidate species-specific patterns. Given that the samples in this study had not reached sexual maturity and individual sexes could not be distinguished, our conclusions are somewhat limited. We suggest collecting sexually mature samples during the breeding season for future research to address this limitation.


 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}