
A New Basal Neornithischian Dinosaur from the Phu Kradung Formation (Upper Jurassic) of Northeastern Thailand

 , ,
, , 
Abstract
1. Introduction
1.1. The Neornithischia
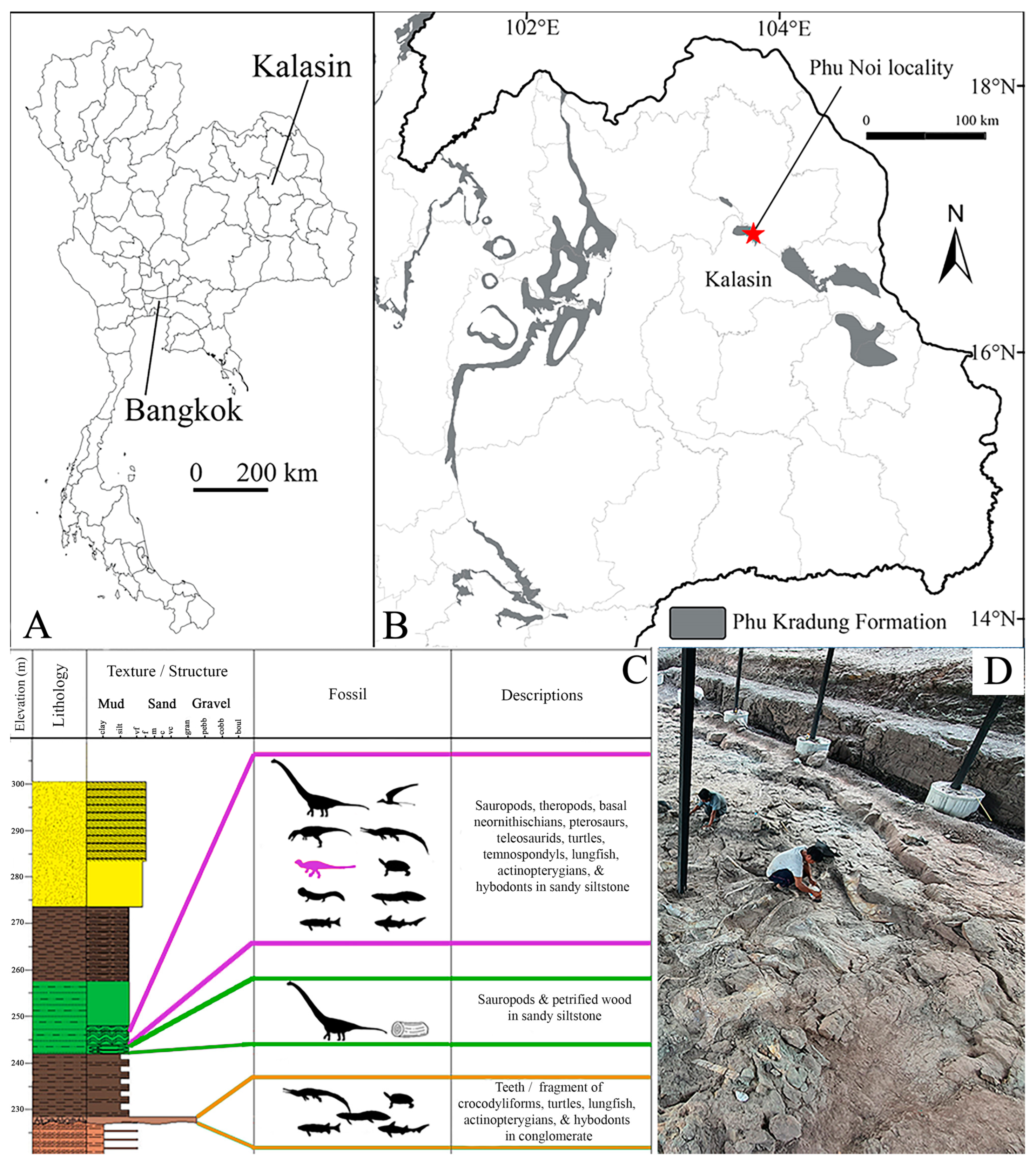
1.2. Geological Settings

2. Materials and Methods
2.1. Examined Material
2.2. Phylogenetic Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Age | Formation | Occurrence | Reference |
|---|---|---|---|---|
| Sanxiasaurus modaoxiensis | Aalenian–Toarcian | Xintiangou | China (Chongqing) | [5] |
| Agilisaurus louderbacki | Middle–Late Jurassic | Lower Shaximiao | China (Sichuan) | [12] |
| Hexinlusaurus multidens | Middle–Late Jurassic | Lower Shaximiao | China (Sichuan) | [10] |
| Xiaosaurus dashanpensis | Middle–Late Jurassic | Lower Shaximiao | China (Sichuan) | [9] |
| Kulindadromeus zabaikalicus | Bathonian | Ukureyskaya | Russia (Cherynyshevsky) | [13] |
| Yandusaurus hongheensis | Late Jurassic | Upper Shaximiao | China (Sichuan) | [41] |
| Minimocursor phunoiensis gen. et sp. nov. | Late Jurassic | Phu Kradung | Thailand (Kalasin) | This study |
| Nanosaurus agilis | Kimmeridgian–Tithonian | Morrison | USA (Colorado, Wyoming) | [42] |
| Hypsilophodon foxii | Barremian | Wessex | England (Isle of Wight) | [43,44] |
| Changmiania liaoningensis | Barremian | Yixian | China (Liaoning) | [45] |
| Jeholosaurus shangyuanensis | Barremian–Aptian | Yixian | China (Liaoning) | [46,47] |
| Changchunsaurus parvus | Aptian–Cenomanian | Quantou | China (Jilin) | [48,49] |
| Leaellynasaura micagraphica | Albian | Eumeralla | Australia (Victoria) | [50] |
- ????? ????? ????? ????? ????? ?0??? ?0?1? ??2?? ?????
- ????? ????? ????? ????? ????? ??130 11011 10?00 0????
- ????? ?1??? ????? ????? ??01? ??0?? ??10? ????1 ??00?
- 0?00? ????? 01100 011?? 0000? ????? ????? ??00? ?00?0
- 00010 01011 11?20 0211? 00??0 ?0001 11000 12120 00-00
- 01221 01000 0?1?? ??0?? 1000? 1100?
- ????? ????? ????? ????? ????? ?0??? ?0?1? ??2?? ?????
- ????? ????? ????? ????? ????? ????? ????? ???00 0????
- ????? ?1??? ????? ????? ????? ????? ????? ????? ?????
- ????? ????? 01100 011?? 0000? ????? ????? ??00? ?00?0
- 00010 01011 11?20 0211? 00?00 ?0001 11000 12120 00-00
- 01221 01000 0?1?? ????? ????? 1100?
- ????? ????? ????? ????? ????? ????? ????? ????? ?????
- ????? ????? ????? ????? ????? ??130 11011 11??? ?????
- ????? ????? ????? ????? ??01? ??0?? ??10? ????1 ??00?
- 0?00? ????? ????? ????? ????? ????? ????? ????? ?????
- ????? ????? ????? ????? ????? ????? ????? ????? ?????
- ????? ????? ????? ????? ????? ?????
2.3. Anatomical Terminologies
2.4. Institutional Abbreviations
3. Results
3.1. Systematic Palaeontology
- Minimocursor gen. nov.
- Minimocursor phunoiensis gen. et sp. nov.
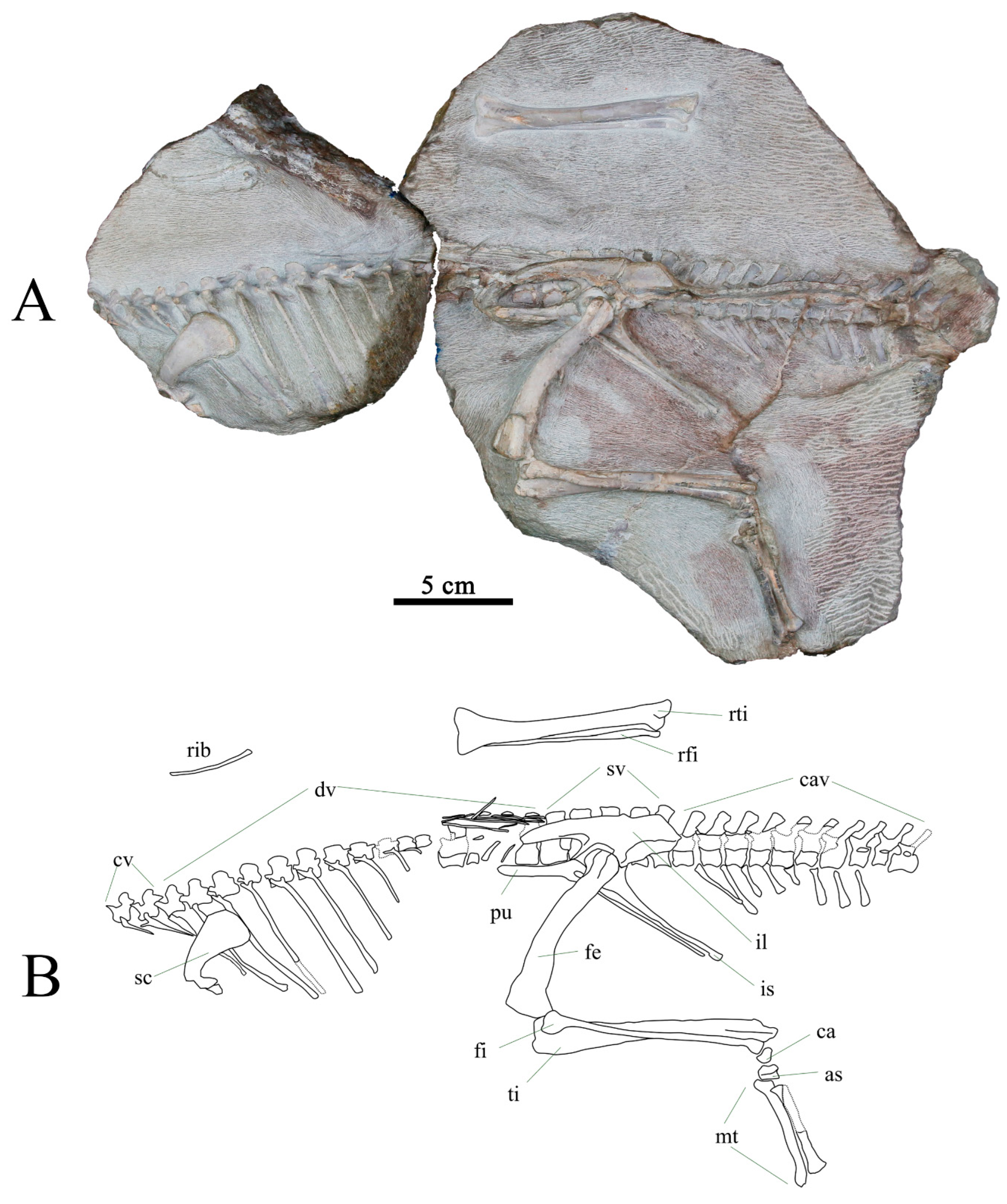
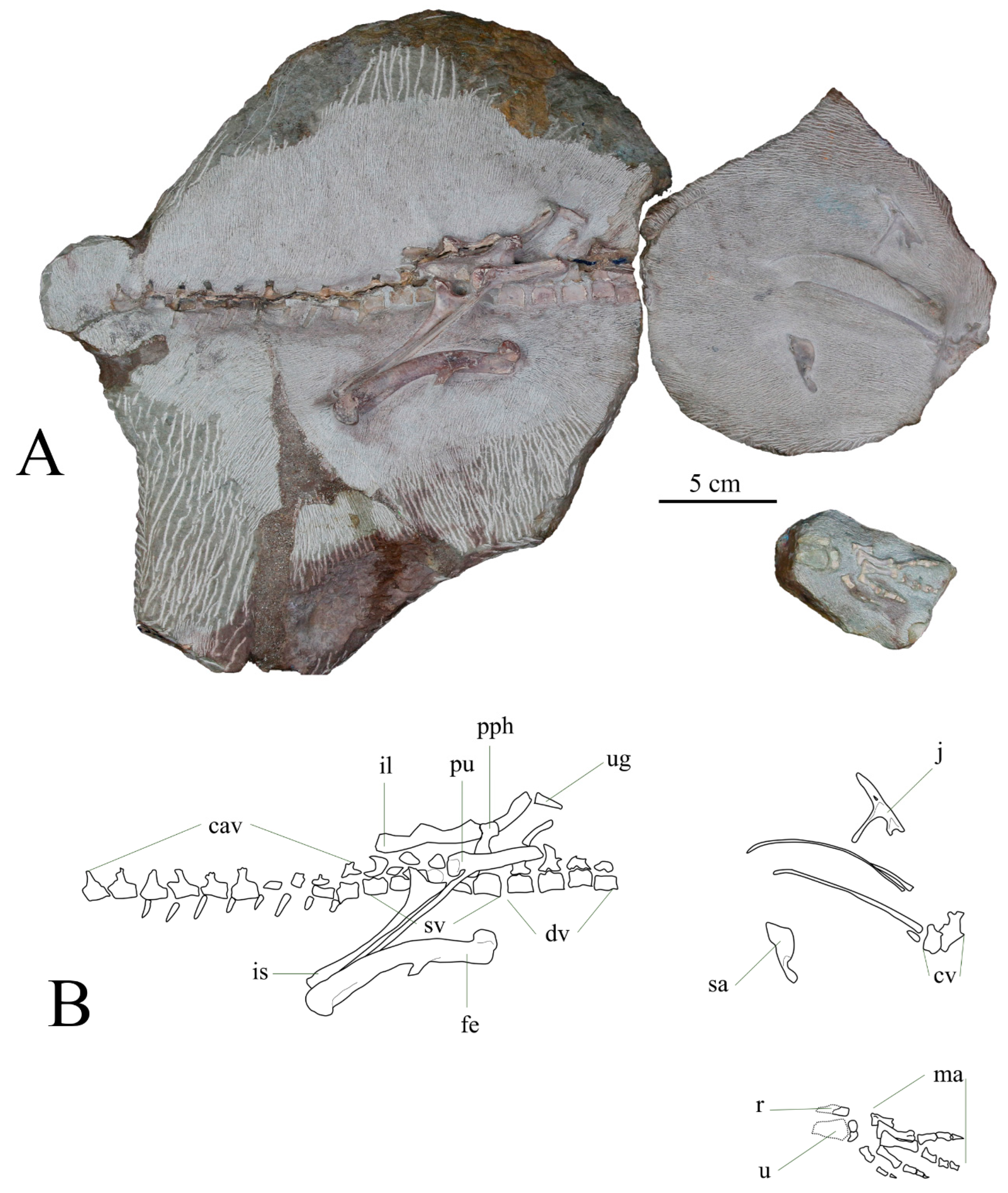
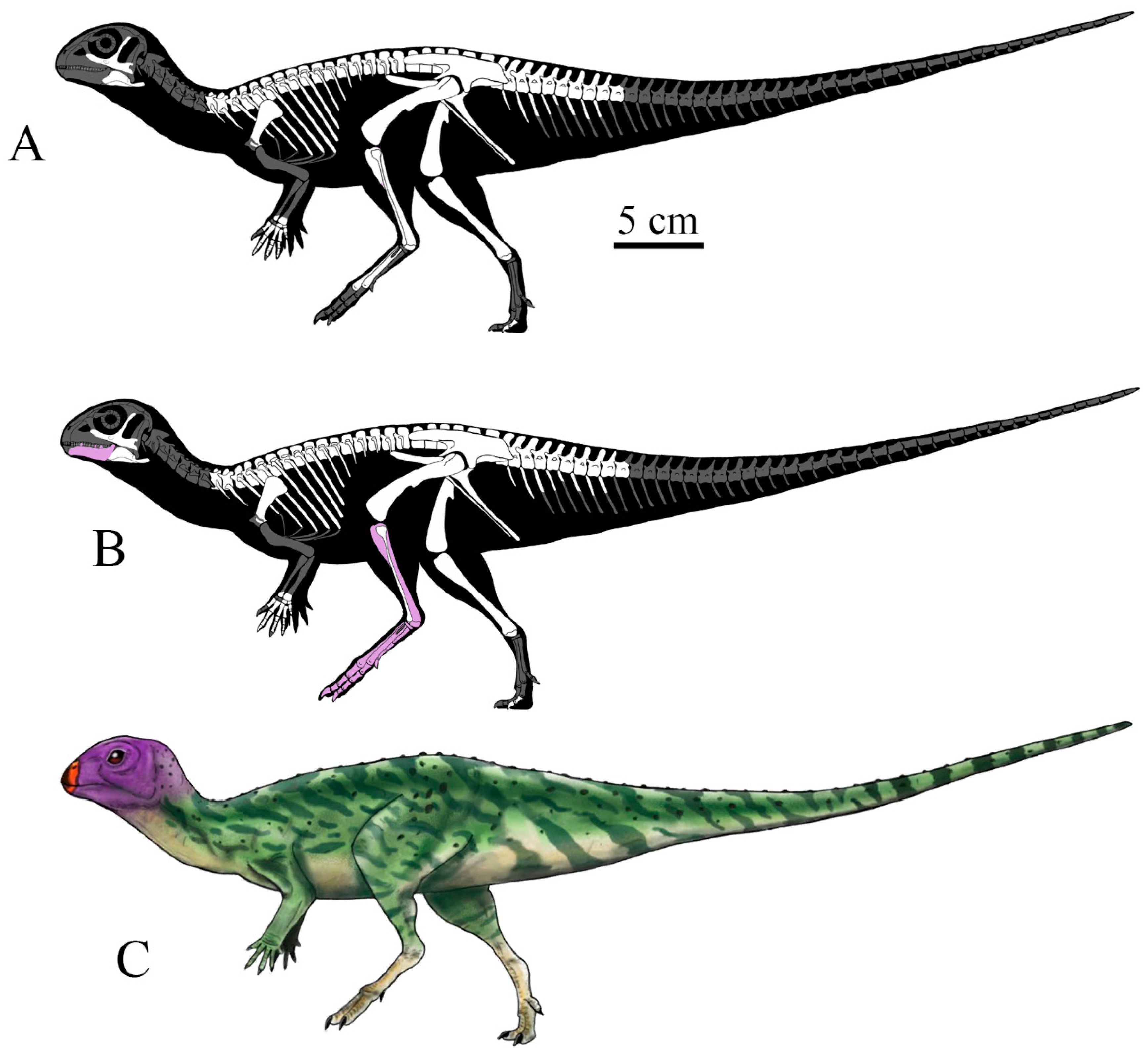
3.2. Anatomical Description
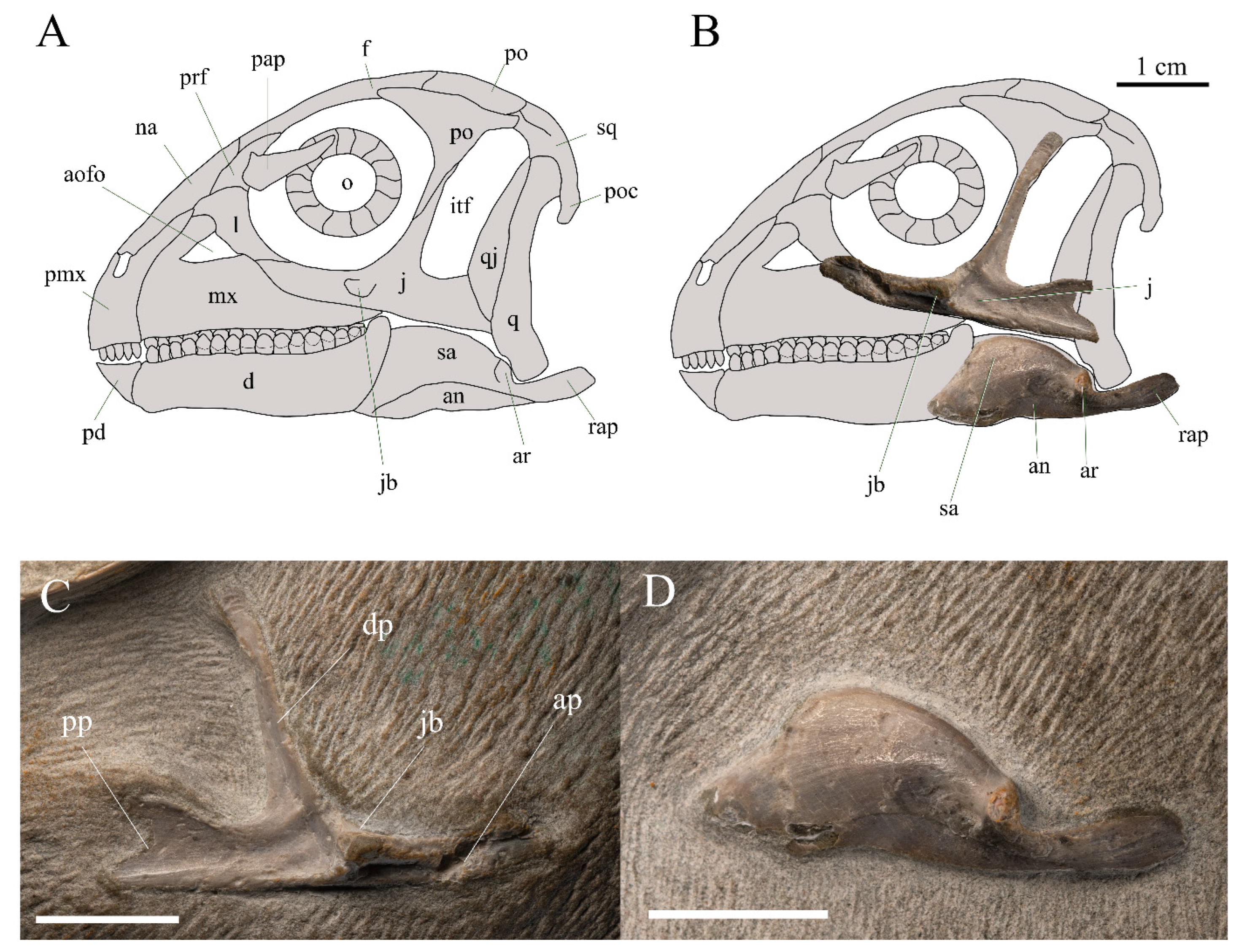
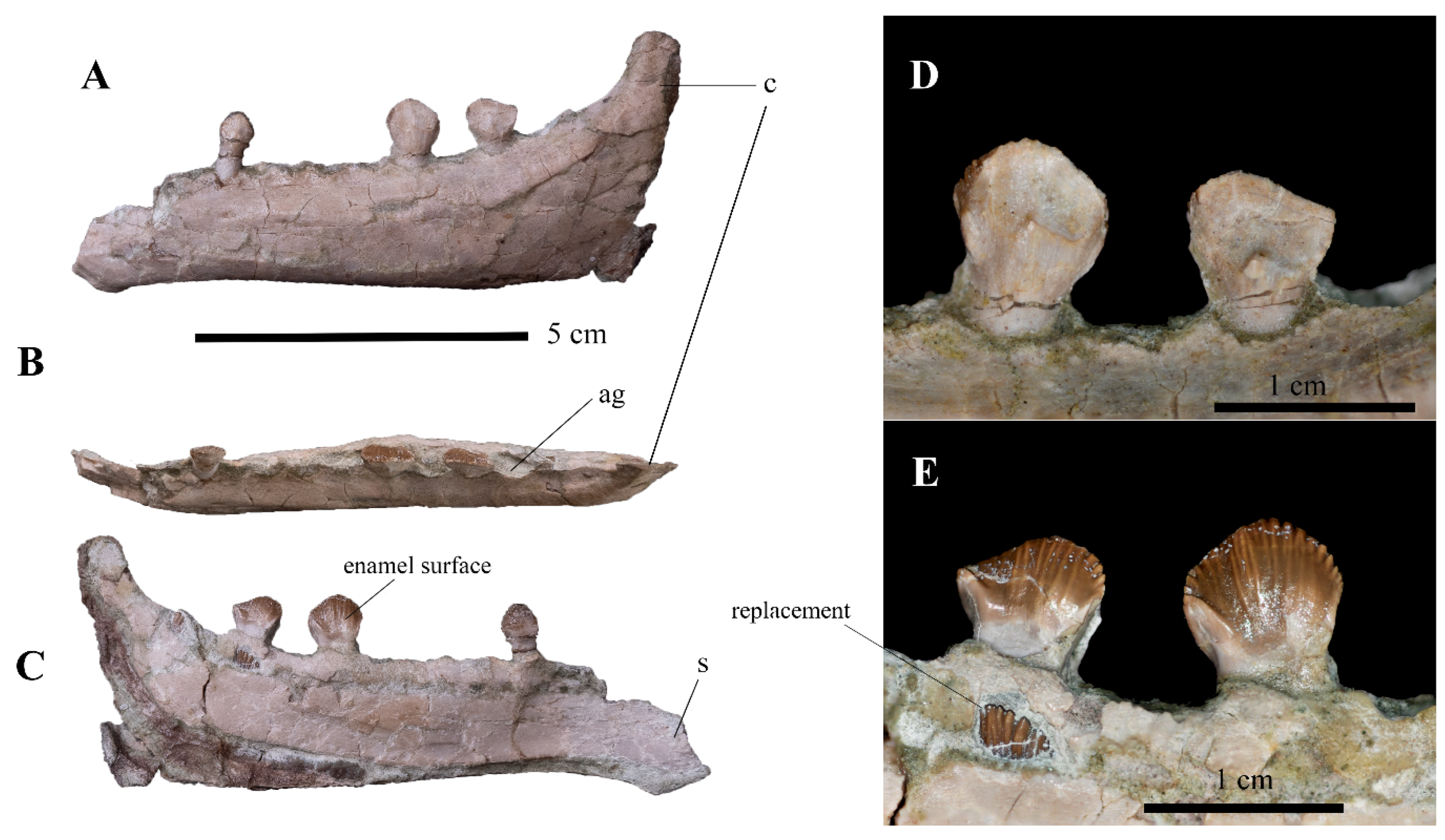
3.2.1. Skull and Mandible

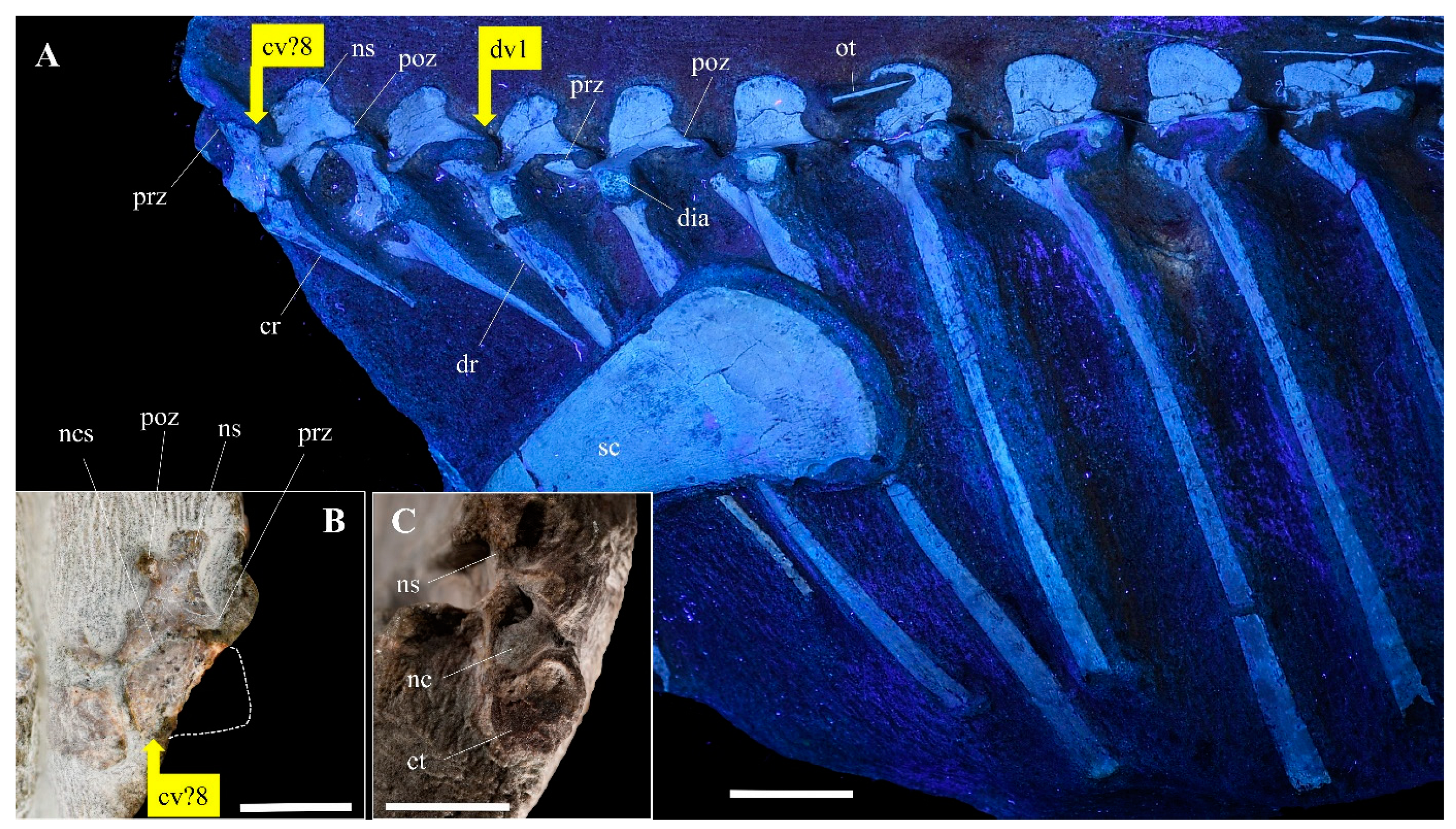
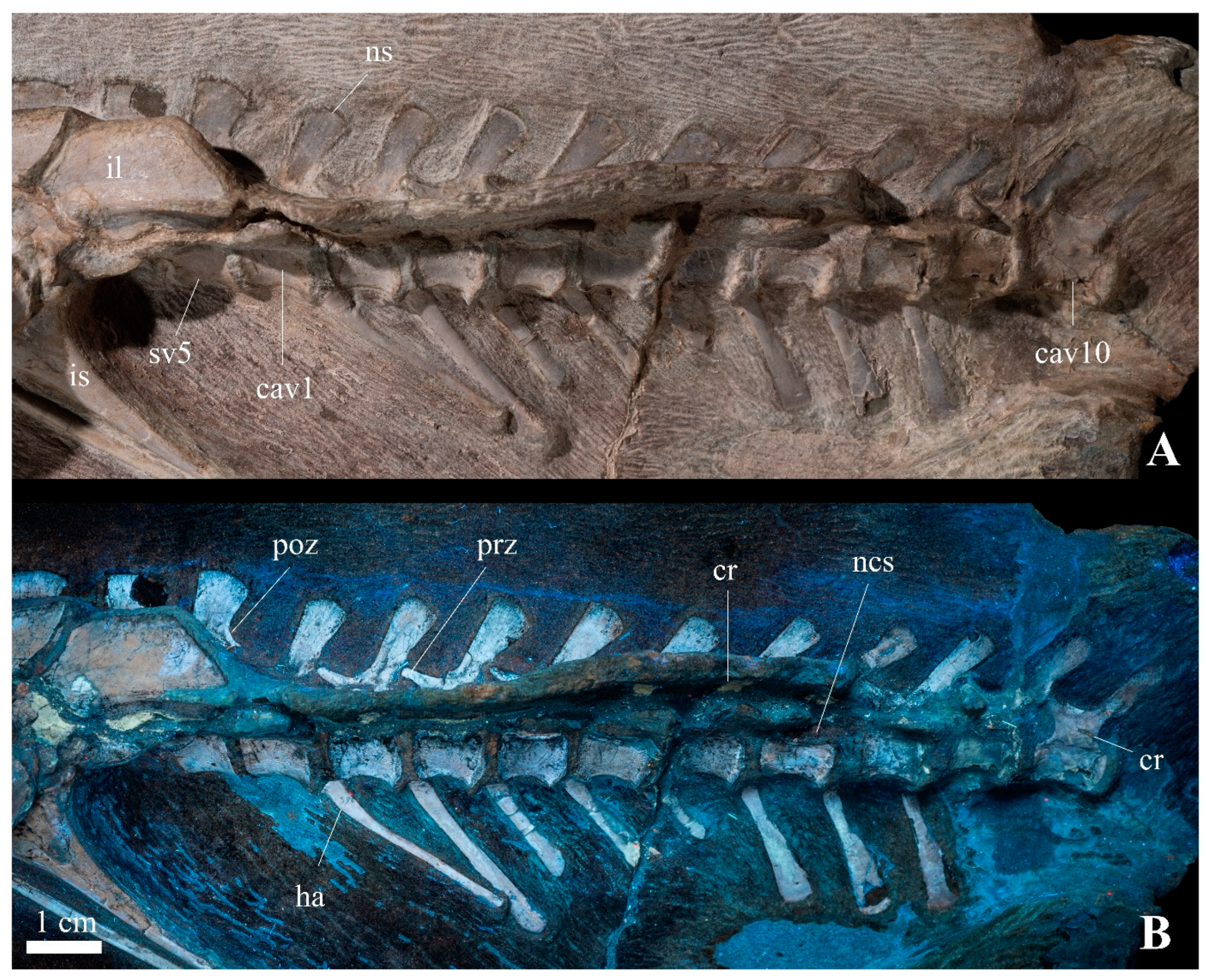
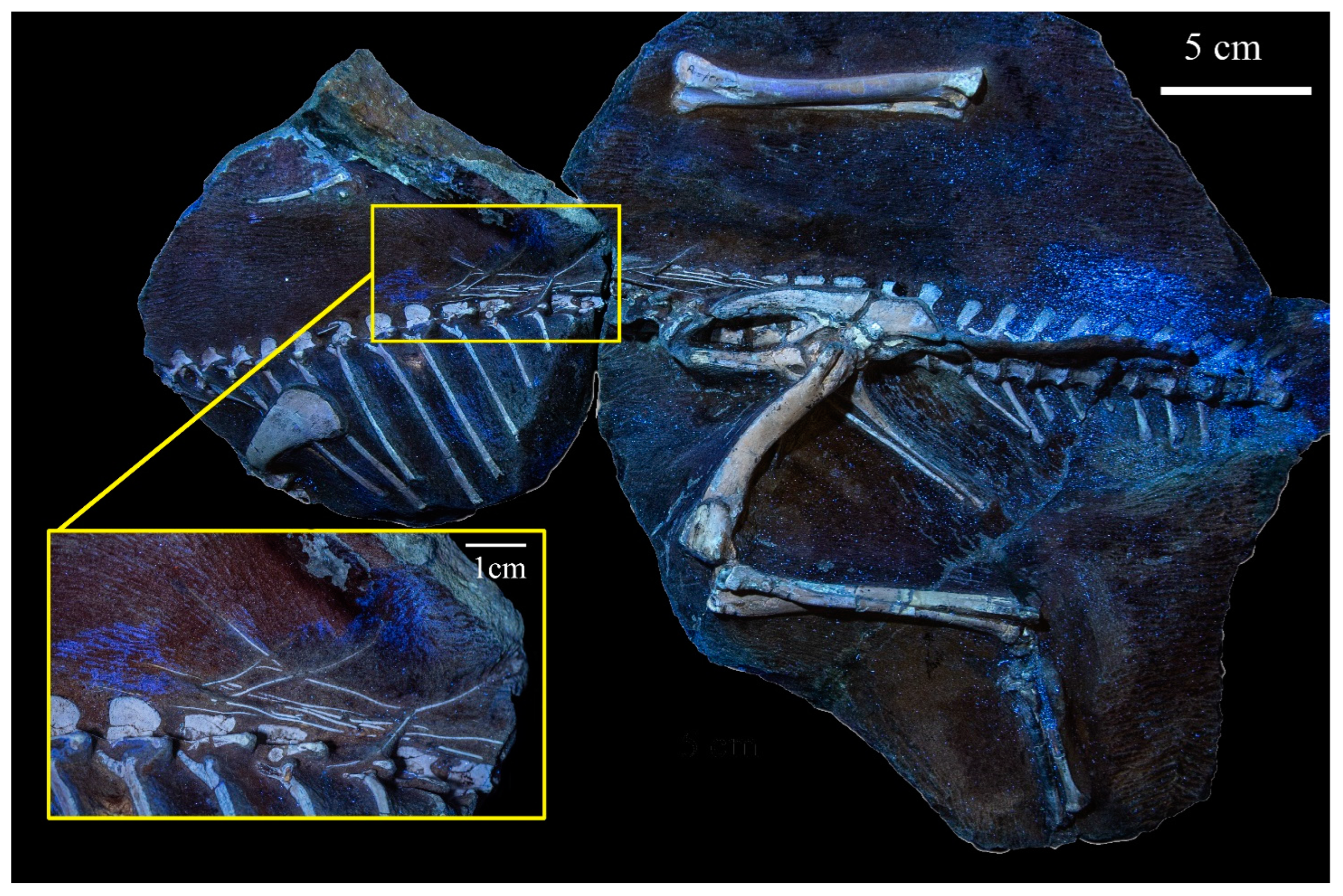
3.2.2. Axial Skeleton

| Taxa | Cervical | Dorsal | Sacral | Caudal | Reference |
|---|---|---|---|---|---|
| Heterodontosaurus tucki | 9 | 12 | 6 | 34–37 | [65] |
| Lesothosaurus diagnosticus | 9 | 15 | 5 | 35–50 | [66] |
| Agilisaurus louderbacki | 9 | 15 | 5 | 44 | [12] |
| Hexinlusaurus multidens | 9 | 15 | 5 | 14+ | [10] |
| Minimocursor phunoiensis | ? | 15 | 5 | 10+ | This study |
| Nanosaurus agilis | 9 | 15 | 6 | ? | [67] |
| Hypsilophodon foxii | 9 | 15–16 | 5–6 | 45–50 | [43] |
| Orodromeus makelai | 9 | 15 | 6 | 30+ | [59] |
| Jeholosaurus shangyuanensis | 9 | 15 | 6 | 15+ | [46,47] |
| Changchunsaurus parvus | 9 | 15 | ? | ? | [48,49] |
| Changmiania liaoningensis | 6 | 15–16 | ? | 36+ | [45] |
| Haya griva | 9 | ?15 | 6 | 19+ | [62,63] |
| Convolosaurus marri | 9 | 15 | 6 | 43 | [68] |
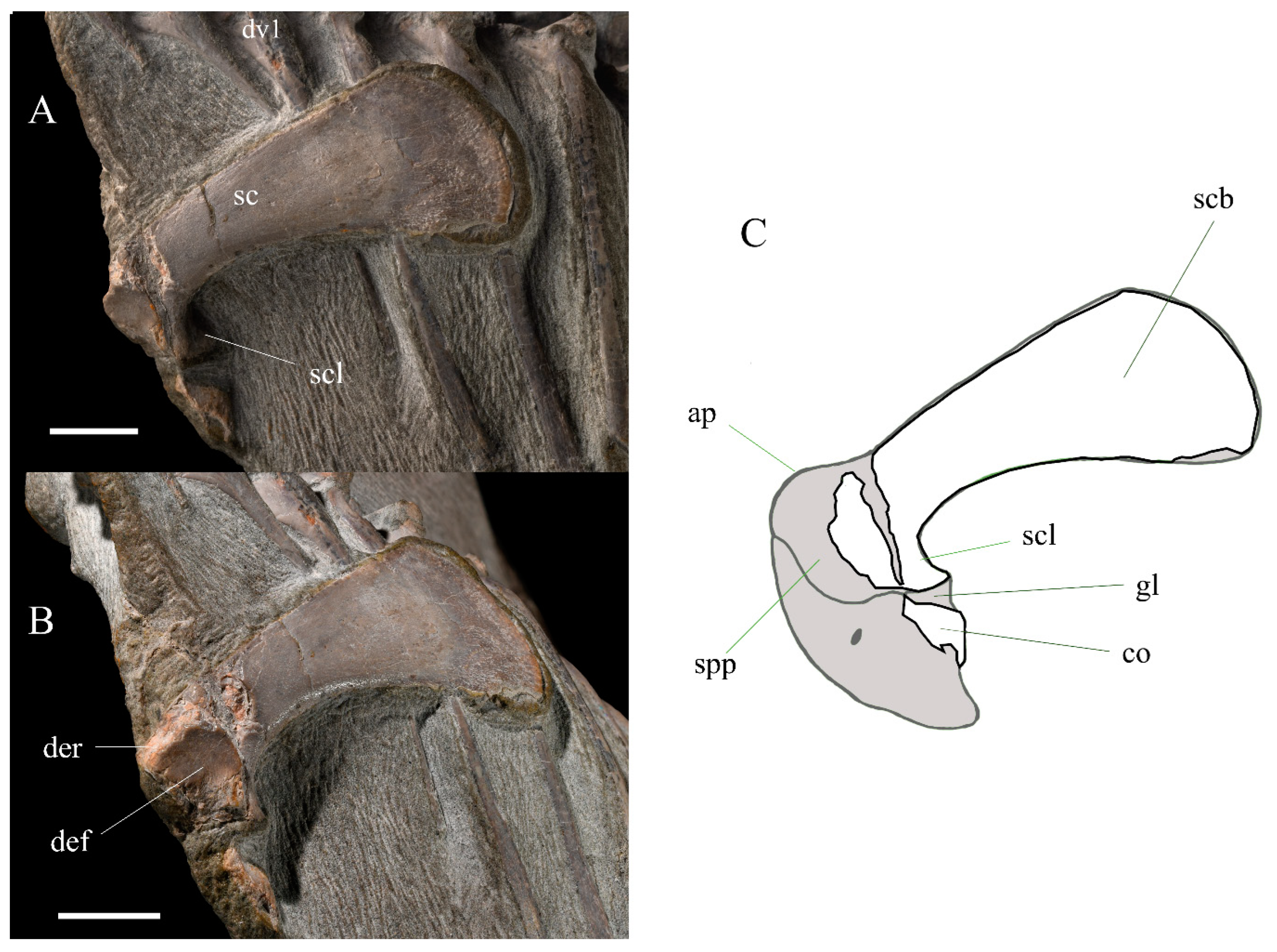
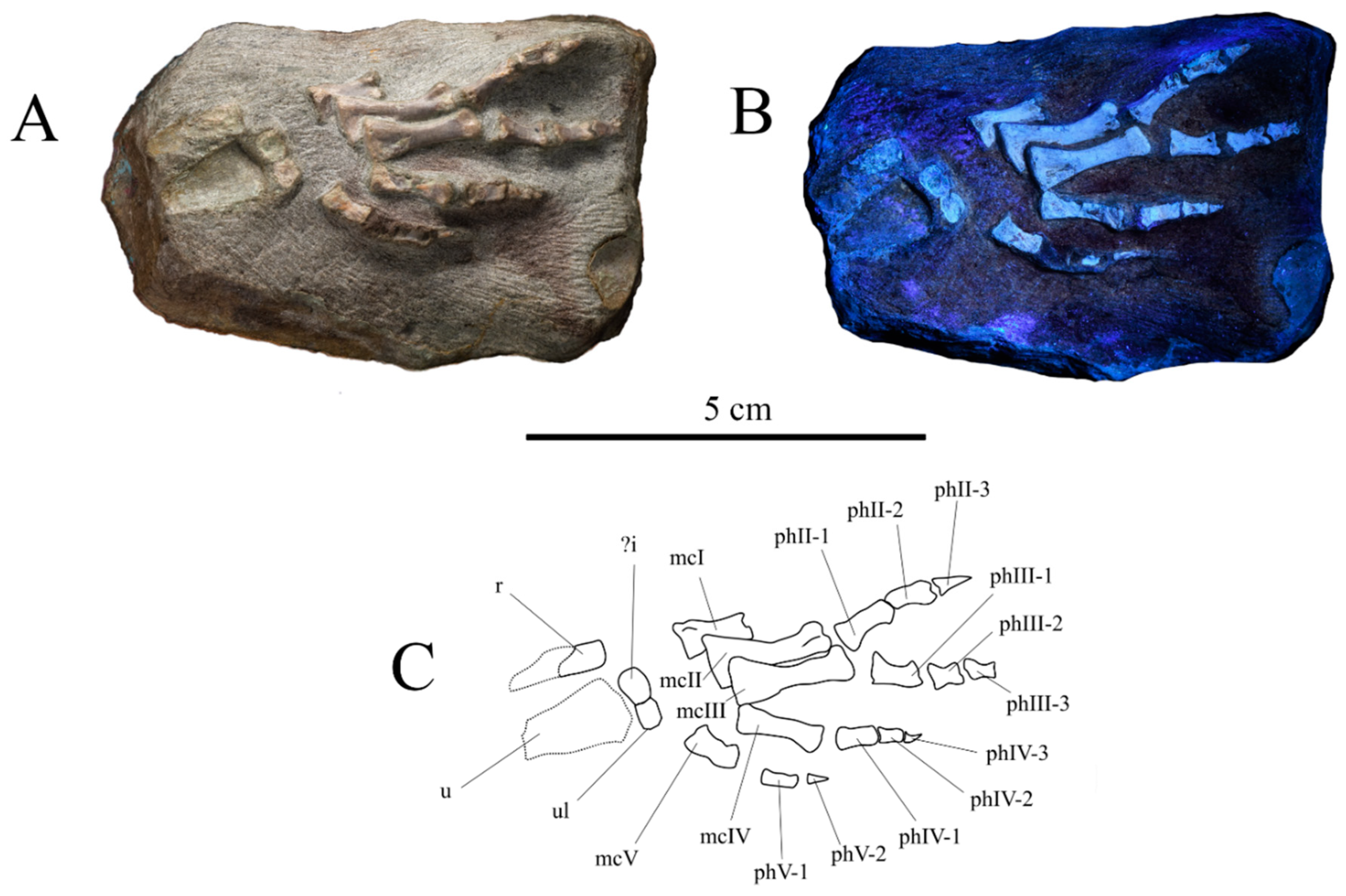
3.2.3. Pectoral Girdle and Forelimb

| Taxa | Family/Clade | Age | Manus | Pes | Source |
|---|---|---|---|---|---|
| Heterodontosaurus tucki | Heterodontosauridae | Early Jurassic, | 2-3-4-3-2 | 2-3-4-5-0 | [60] |
| Xiaosaurusdashanpuensis | Basal neornithischia | Middle Jurassic | ? | 2?-3-4-5-0 | [9] |
| Agilisaurus louderbacki | Basal neornithischia | Middle Jurassic, | ? | 2-3-4-5-0 | [12] |
| Hexinlusaurus multidens | Basal neornithischia | Middle Jurassic | 2-3-4-2?-2 | 2-3-4-5-0 | [10] |
| Minimocursor phunoiensis | Basal neornithischia | Late Jurassic | ? -3-4-3-2 | 2-3-4-5-0 | This study |
| Hypsilophodon foxii | Basal neornithischia | Early Cretaceous, | 2-3-4-3?-1? | 2-3-4-5-0 | [74] |
| Changmiania liaoningensis | Basal neornithischia | Early Cretaceous | 2-3-?-?-? | 2-3-4-5-? | [45] |
| Jeholosaurus shangyuanensis | Jeholosauridae | Early Cretaceous | ? | 2-3-4-5-0 | [46,49] |
| Changchunsaurus parvus | Thescelosauridae | Early Cretaceous | ? | 2-3-4-5-? | [49] |
| Haya griva | Thescelosauridae | Late Cretaceous | 2?-3?-4?-?-?. | 2-3-4-5-0, | [62,63] |
| Orodromeus makelai | Thescelosauridae | Late Cretaceous | ?-3-4-3-? | 2-3-4-5-0 | [60] |
| Dryosaurus altus | Iguanodontia | Late Jurassic, | 2-3-4-3-2 | 0-3-4-5-0 | [60] |
| Tenontosaurus tilletti | Iguanodontia | Early Cretaceous, | 2-3-3-2-1 | 2-3-4-5-0 | [75] |
| Camptosaurus dispar | Iguanodontia | Late Jurassic | 2-3-3-2-1 | 2-3-4-5-0 | [76,77] |
| Iguanodon bemissartensis | Iguanodontidae | Early Cretaceous, | 2-3-3-2-4 | 0-3-4-5-0 | [78] |
| Edmontosaurus regalis | Hadrosauridae | Late Cretaceous, | 0-3-3-3-3 | 0-3-4-5-0 | [60] |
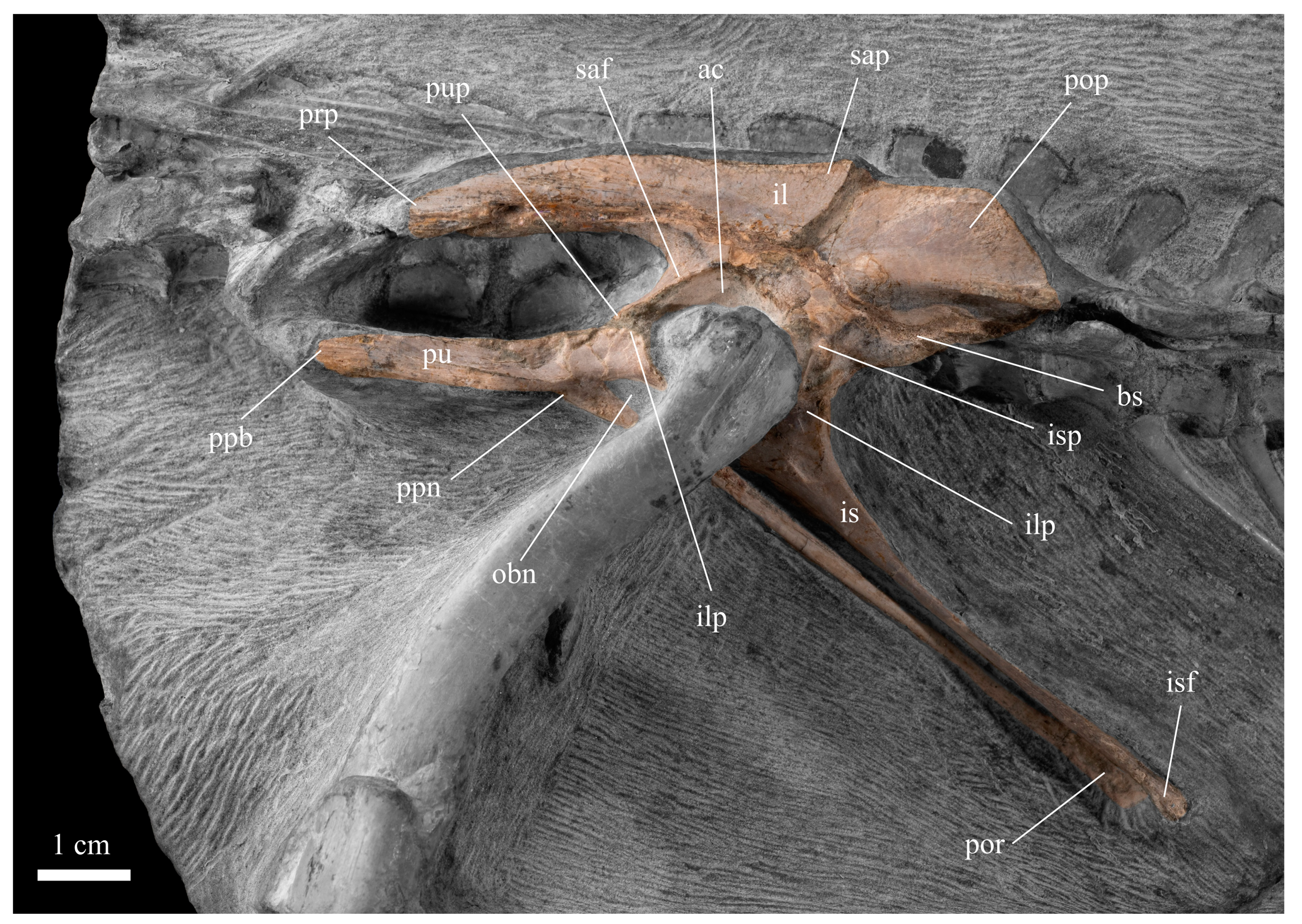
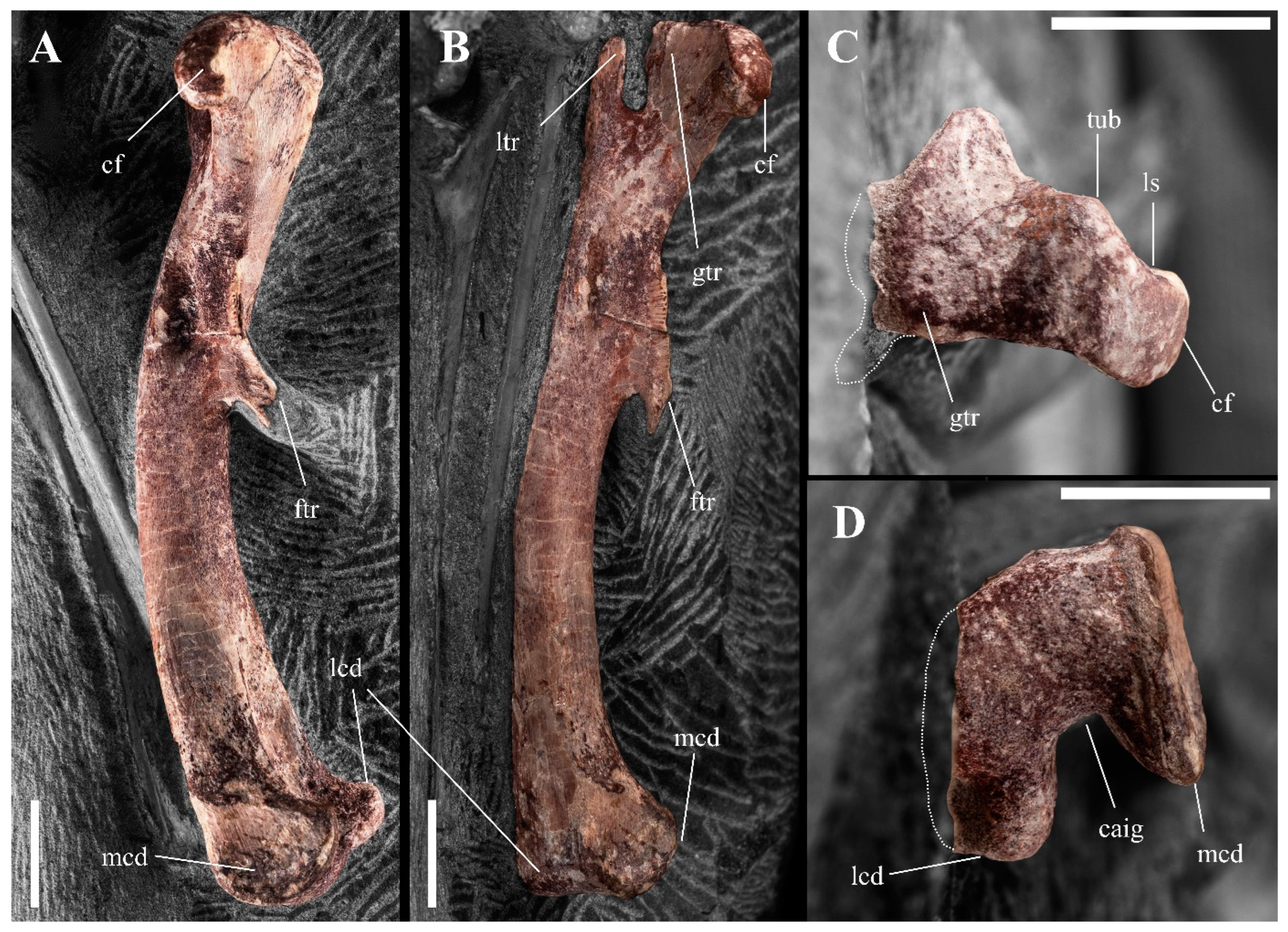
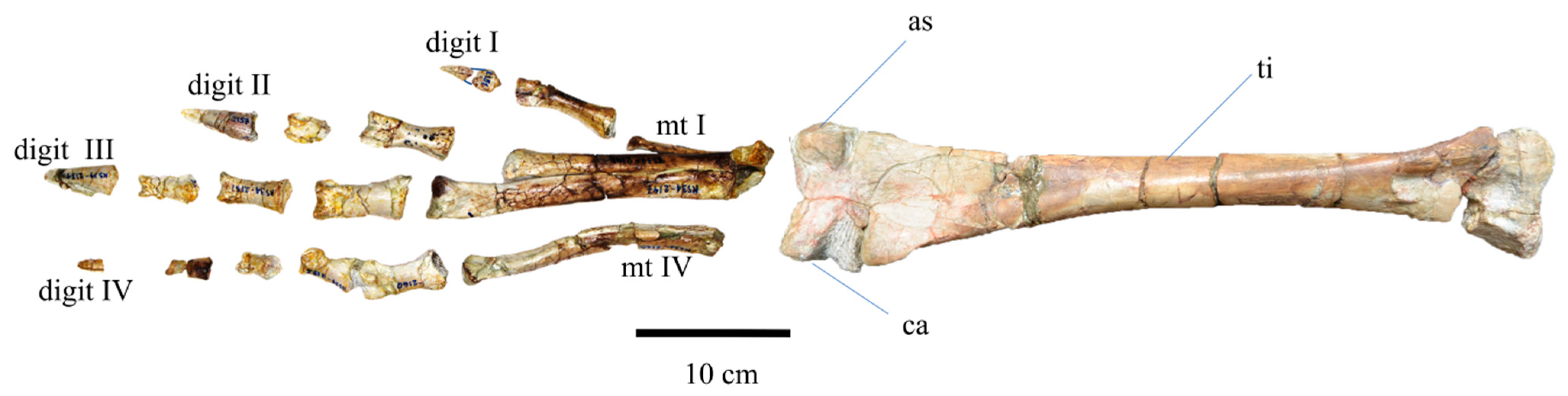
3.2.4. Pelvic Girdle and Hindlimb
3.2.5. Ontogenetic Assessment
3.2.6. Referred Material


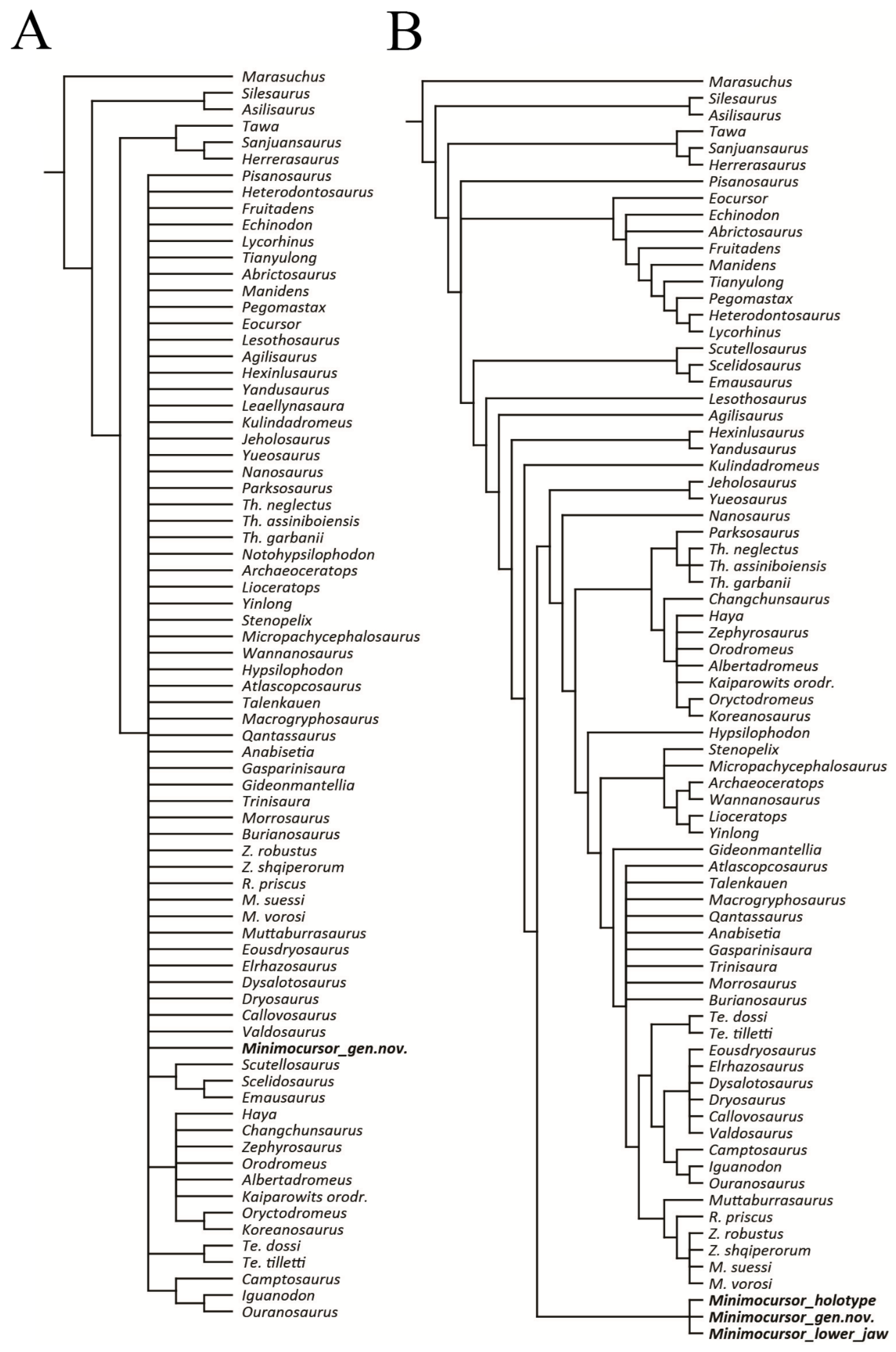
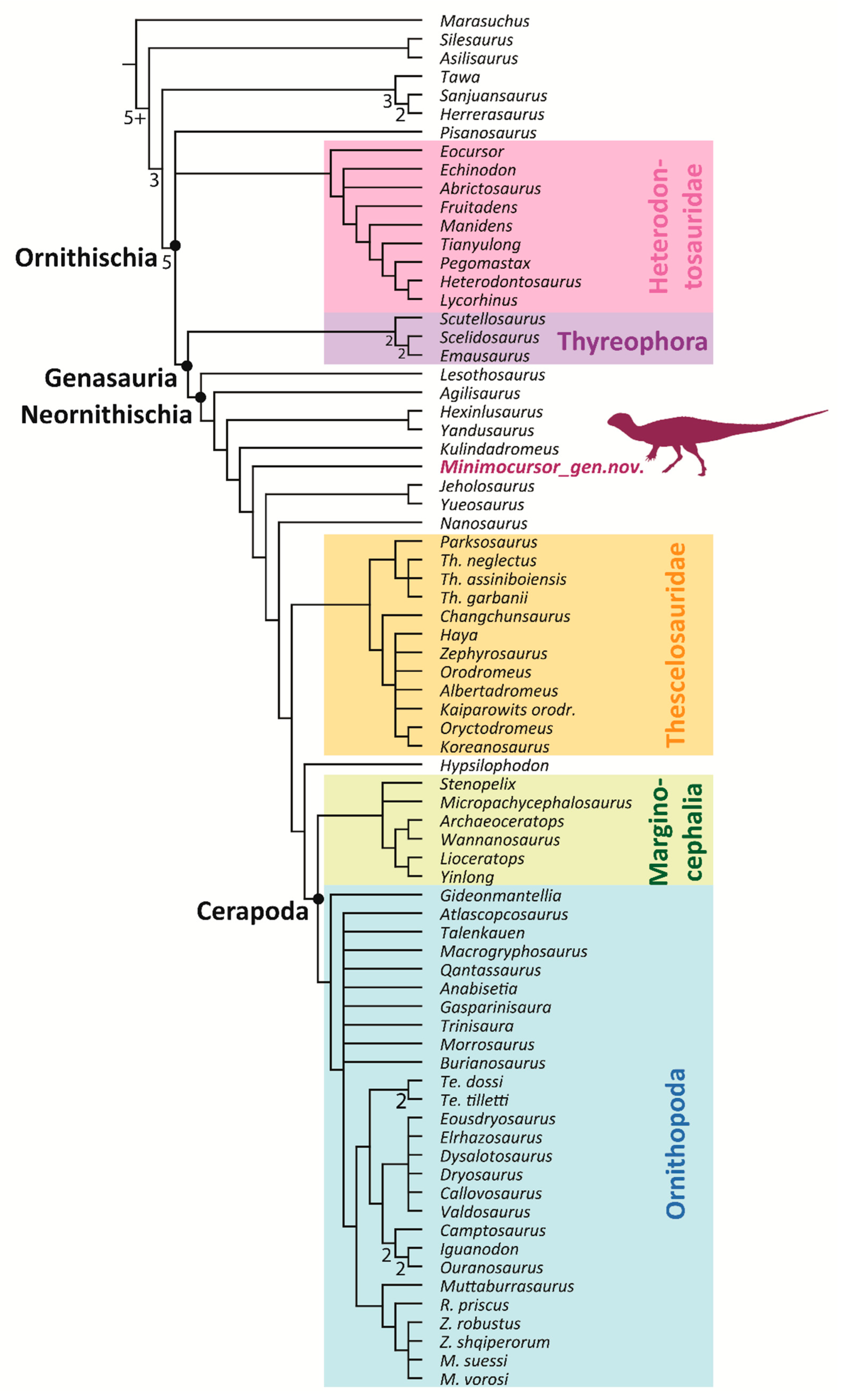
3.3. Phylogenetic Analysis


4. Comparison and Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barrett, P.M.; Butler, R.J.; Knoll, F. Small-Bodied Ornithischian Dinosaurs from the Middle Jurassic of Sichuan, China. Source J. Vertebr. Paleontol. 2005, 25, 823–834. [Google Scholar] [CrossRef]
- Butler, R.J.; Upchurch, P.; Norman, D.B. The Phylogeny of the Ornithischian Dinosaurs. J. Syst. Palaeontol. 2008, 6, 1–40. [Google Scholar] [CrossRef]
- Paul, C. Sereno The Evolution of Dinosaurs. Science 1999, 284, 2137–2147. [Google Scholar] [CrossRef]
- Boyd, C.A. The Systematic Relationships and Biogeographic History of Ornithischian Dinosaurs. PeerJ 2015, 3, e1523. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Dai, H.; Tan, C.; Hu, X.; Wei, Z.; Lin, Y.; Wei, G.; Li, D.; Meng, L.; Hao, B.; et al. A Neornithischian Dinosaur from the Middle Jurassic Xintiangou Formation of Yunyang, Chongqing, China: The Earliest Record in Asia. Hist. Biol. 2019, 33, 1089–1102. [Google Scholar] [CrossRef]
- Madzia, D.; Boyd, C.A.; Mazuch, M. A Basal Ornithopod Dinosaur from the Cenomanian of the Czech Republic. J. Syst. Palaeontol. 2018, 16, 967–979. [Google Scholar] [CrossRef]
- Dieudonné, P.E.; Cruzado-Caballero, P.; Godefroit, P.; Tortosa, T. A New Phylogeny of Cerapodan Dinosaurs. Hist. Biol. 2020, 33, 2335–2355. [Google Scholar] [CrossRef]
- Madzia, D.; Arbour, V.M.; Boyd, C.A.; Farke, A.A.; Cruzado-Caballero, P.; Evans, D.C. The Phylogenetic Nomenclature of Ornithischian Dinosaurs. PeerJ 2021, 9, e12362. [Google Scholar] [CrossRef]
- Dong, Z.; Tang, Z. A Short Report on the Dinosaur Fauna from Dashanpu, Zigong, Sichuan II Ornithopoda. Vertebr. Palasiat. 1983, 21, 168–171. [Google Scholar]
- He, X.; Cai, K. The Middle Jurassic Dinosaurian Fauna from Dashanpu, Zigong, Sichuan. Sichuan Sci. Technol. Publ. House 1984, I, 1–66. [Google Scholar]
- Li, K.; Liu, J.; Yang, C.; Hu, F. Dinosaur Assemblages from the Middle Jurassic Shaximiao Formation and Chuanjie Formation in the Sichuan-Yunnan Basin, China. Vol. Jurass. 2011, IX, 21–42. [Google Scholar]
- Peng, G. Jurassic Ornithopod Agilisaurus louderbacki (Ornithopoda: Fabrosauridae) from Zigong, Sichuan, China. Vertebr. Palasiat. 1992, 30, 39–51. [Google Scholar]
- Godefroit, P.; Sinitsa, S.M.; Dhouailly, D.; Bolotsky, Y.L.; Sizov, A.V.; McNamara, M.E.; Benton, M.J.; Spagna, P. A Jurassic Ornithischian Dinosaur from Siberia with Both Feathers and Scales. Science 2014, 345, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Hoffet, J.H. Description Des Ossements Les plus Caractéristiques Appartenant à Des Avipelviens Du Sénonien Du Bas-Laos [Description of the Most Characteristic Bones Belonging to Bird-Hipped Dinosaurs from the Senonian of Lower Laos]. Comptes Rendus Des Séances Du Cons. Des Rech. Sci. L’indochine 1944, 1944, 179. [Google Scholar]
- Buffetaut, E.; Suteethorn, V. A New Iguanodontian Dinosaur from the Khok Kruat Formation (Early Cretaceous, Aptian) of Northeastern Thailand. Ann. Paleontol. 2011, 97, 51–62. [Google Scholar] [CrossRef]
- Shibata, M.; Jintasakul, P.; Azuma, Y. A New Iguanodontian Dinosaur from the Lower Cretaceous Khok Kruat Formation, Nakhon Ratchasima in Northeastern Thailand. Acta Geol. Sin.-Engl. Ed. 2011, 85, 969–976. [Google Scholar] [CrossRef]
- Shibata, M.; Jintasakul, P.; Azuma, Y.; You, H.-L. A New Basal Hadrosauroid Dinosaur from the Lower Cretaceous Khok Kruat Formation in Nakhon Ratchasima Province, Northeastern Thailand. PLoS ONE 2015, 10, e0145904. [Google Scholar] [CrossRef]
- Buffetaut, E.; Suteethorn, V. A New Species of the Ornithischian Dinosaur Psittacosaurus from the Early Cretaceous of Thailand. Palaeontology 1992, 35, 801–812. [Google Scholar]
- Buffetaut, E.; Suteethorn, V.; Cuny, G.; Khansubha, S.; Tong, H.; Le Loeuff, J.; Cavin, L. Dinosaurs in Thailand. Maha Sarakham Univ. J. Spec. Issue 2003, 22, 69–82. [Google Scholar] [CrossRef]
- Manitkoon, S.; Deesri, U.; Warapeang, P.; Nonsrirach, T.; Chanthasit, P. Ornithischian Dinosaurs in Southeast Asia: A Review with Palaeobiogeographic Implications. Foss. Rec. 2023, 26, 1–25. [Google Scholar] [CrossRef]
- Buffetaut, E.; Suteethorn, S.; Suteethorn, V.; Deesri, U.; Tong, H. Preliminary Note on a Small Ornithopod Dinosaur from the Phu Kradung Formation (Terminal Jurassic—Basal Cretaceous) of Phu Noi, North-Eastern Thailand. J. Sci. Technol. Mahasarakham Univ. 2014, 33, 344–347. [Google Scholar]
- Meesook, A. Cretaceous Environments of North- Eastern Thailand. In Cretaceous Environments of Asia; Okada, H., Mateer, N.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2000; pp. 207–223. ISBN 9780444502766. [Google Scholar]
- Strokes, R.B.; Raksaskulwong, M. (Eds.) Department of Mineral Resources Mesozoic Era. In Geology of Thailand; Bureau of Geological Suevey Department of Mineral Resources: Bangkok, Thailand, 2014; pp. 113–157. [Google Scholar]
- Cuny, G.; Liard, R.; Deesri, U.; Liard, T.; Khamha, S.; Suteethorn, V. Shark Faunas from the Late Jurassic—Early Cretaceous of Northeastern Thailand. Palaontol. Z. 2014, 88, 309–328. [Google Scholar] [CrossRef]
- Martin, J.E.; Suteethorn, S.; Lauprasert, K.; Tong, H.; Buffetaut, E.; Liard, R.; Salaviale, C.; Deesri, U.; Claude, J.; Martin, J.E.; et al. A New Freshwater Teleosaurid from the Jurassic of Northeastern Thailand. J. Vertebr. Paleontol. 2019, 38, 1–28. [Google Scholar] [CrossRef]
- Boonchai, N.; Suteethorn, S.; Sereeprasirt, W.; Suriyonghanphong, C.; Romain, A.; Cuny, G.; Legrand, J.; Thévenard, F.; Philippe, M. Xenoxylon, a Boreal Fossil Wood in the Mesozoic Redbeds of Southeast Asia: Potential for the Stratigraphy of the Khorat Group and the Palinspatic Reconstruction of Southeast Asia. J. Asian Earth Sci. 2019, 189, 104153. [Google Scholar] [CrossRef]
- Ditbanjong, P.; Chanthasit, P. Sedimentary Facies and Depositional Environment of Phu Noi Sita Fro the Late Jurassic Phu Kradung Formation, Khorat Group, Kalasin Province, Northeastern Thailand. In Proceedings of the 4th International Symposium on Asian Dinosaurs in Mongolia, Mongolia, Ulaanbaatar, 24–25 July 2019; pp. 50–51. [Google Scholar]
- Deesri, U.; Lauprasert, K.; Suteethorn, V.; Wongko, K.; Cavin, L. A New Species of the Ginglymodian Fish Isanichthys (Actinopterygii, Holostei) from the Late Jurassic Phu Kradung Formation, Northeastern Thailand. Acta Palaeontol. Pol. 2014, 59, 313–331. [Google Scholar] [CrossRef]
- Cavin, L.; Deesri, U.; Chanthasit, P. A New Lungfish from the Jurassic of Thailand. J. Vertebr. Paleontol. 2020, 40, e1791895. [Google Scholar] [CrossRef]
- Tong, H.; Naksri, W.; Buffetaut, E.; Suteethorn, V.; Suteethorn, S.; Deesri, U.; Sila, S.; Chanthasit, P.; Claude, J. A New Primitive Eucryptodiran Turtle from the Upper Jurassic Phu Kradung Formation of the Khorat Plateau, NE Thailand. Geol. Mag. 2015, 152, 166–175. [Google Scholar] [CrossRef]
- Tong, H.; Naksri, W.; Bu, E.; Suteethorn, S.; Suteethorn, V.; Chanthasit, P.; Claude, J. Kalasinemys, a New Xinjiangchelyid Turtle from the Late Jurassic of NE Thailand. Geol. Mag. 2019, 156, 1645–1656. [Google Scholar] [CrossRef]
- Wilberg, E.W.; Turner, A.H.; Brochu, C.A. Evolutionary Structure and Timing of Major Habitat Shifts in Crocodylomorpha. Sci. Rep. 2019, 9, 514. [Google Scholar] [CrossRef]
- Buffetaut, E.; Suteethorn, V.; Suteethorn, S.; Deesri, U.; Tong, H. An Azhdarchoid Pterosaur Humerus from the Latest Jurassic (Phu Kradung Formation) of Phu Noi, North-Eastern Thailand. Res. Knowl. 2015, 1, 43–47. [Google Scholar] [CrossRef]
- Chanthasit, P.; Suteethorn, S.; Suteethorn, V. Dinosaur Assemblage from Phu Noi Fossil Sited in Kalasin Province, Northeastern Thailand. In Proceedings of the 2nd International Symposium Asian Dinosaurs in Thailand 2015, Bangkok City, Thailand, 20–29 November 2015; pp. 22–23. [Google Scholar]
- Nonsrirach, T.; Manitkoon, S.; Lauprasert, K. First Occurrence of Brachyopid Temnospondyls in Southeast Asia and Review of the Mesozoic Amphibians from Thailand. Foss. Rec. 2021, 24, 33–47. [Google Scholar] [CrossRef]
- Unwin, D.M.; Martill, D.M.; Le, L. Systematic Reassessment of the First Jurassic Pterosaur from Thailand. Geol. Soc. Lond. Spec. Publ. 2018, 455, 181–186. [Google Scholar] [CrossRef]
- Samathi, A.; Chanthasit, P.; Sander, P.M. A Review of Theropod Dinosaurs from the Late Jurassic to Mid-Cretaceous of Southeast Asia. Ann. Paleontol. 2019, 105, 201–215. [Google Scholar] [CrossRef]
- Buffetaut, E.; Suteethorn, V.; Tong, H.Y. The First Thyreophoran Dinosaur from Southeast Asia: A Stegosaur Vertebra from the Late Jurassic Phu Kradung Formation of Thailand. Neues Jahrb. Fur Geol. Und Palaontol.-Mon. 2001, 2001, 95–102. [Google Scholar] [CrossRef]
- Sues, H.; Evans, D.C.; Galton, P.M.; Brown, C.M. Anatomy of the Neornithischian Dinosaur Parksosaurus warreni (Parks, 1926) from the Upper Cretaceous (Lower Maastrichtian) Horseshoe Canyon Formation of Alberta, Canada. Cretac. Res. 2023, 141, 105369. [Google Scholar] [CrossRef]
- Goloboff, P.A.; Morales, M.E. TNT Version 1.6, with a Graphical Interface for MacOS and Linux, Including New Routines in Parallel. Cladistics 2023, 39, 144–153. [Google Scholar] [CrossRef] [PubMed]
- He, X.-L. A Newly Discovered Ornithopod Dinosaur, Yandusaurus from Zigong, Sichuan. In Contributions to International Exchange of Geology, Part 2 (Stratigraphy and Paleontology); Geological Publishing House: Beijing, China, 1979; pp. 116–123. [Google Scholar]
- Marsh, O.C. Principal Characters of American Jurassic Dinosaurs. Part I. Am. J. Sci. Arts 1878, 16, 411–416. [Google Scholar] [CrossRef]
- Galton, P.M. The Ornithischian Dinosaur Hypsilophodon from the Wealden of the Isle of Wight. Bull. Br. Mus. Nat. Hist. Geol. 1974, 25, 157. [Google Scholar] [CrossRef]
- Huxley, T.H. On Hypsilophodon foxii, a New Dinosaurian from the Wealden of the Isle of Wight. Q. J. Geol. Soc. Lond. 1870, 26, 3–12. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, W.; Dieudonné, P. A New Basal Ornithopod Dinosaur from the Lower Cretaceous of China. PeerJ 2020, 8, e9832. [Google Scholar] [CrossRef]
- Han, F.-L.; Barrett, P.M.; Butler, R.J.; Xu, X.; Han, F. Postcranial Anatomy of Jeholosaurus shangyuanensis (Dinosauria, Ornithischia) from the Lower Cretaceous Yixian Formation of China. J. Vertebr. Paleontol. 2012, 32, 1370–1395. [Google Scholar] [CrossRef]
- Xu, X.; Wang, X.; You, H.-L. A Primitive Ornithopod from the Early Cretaceous Yixian Formation of Liaoning. Vertebr. Palasiat. 2000, 38, 318–325. [Google Scholar]
- Zan, S.; Chen, J.; Jin, L.; Li, T. A Primitive Ornithopod from the Early Cretacoeus Quantou Formation of Central Jilin, China. Vertebr. Palasiat. 2005, 43, 182–193. [Google Scholar]
- Butler, R.J.; Liyong, J.; Jun, C.; Godefroit, P. The Postcranial Osteology and Phylogenetic Position of the Small Ornithischian Dinosaur Changchunsaurus parvus from the Quantou Formation (Cretaceous: Aptian-Cenomanian) of Jilin Province, North-Eastern China. Palaeontology 2011, 54, 667–683. [Google Scholar] [CrossRef]
- Rich, T.H.; Galton, P.M.; Vickers-Rich, P. The Holotype Individual of the Ornithopod Dinosaur Leaellynasaura amicagraphica Rich & Rich, 1989 (Late Early Cretaceous, Victoria, Australia). Alcheringa 2010, 34, 385–396. [Google Scholar] [CrossRef]
- Owen, R. Report on British Fossil Reptiles. Part II; John Murray: London, UK, 1842. [Google Scholar]
- Seeley, H.G. On the Classification of the Fossil Animals Commonly Named Dinosauria. Proc. R. Soc. Lond. 1888, 43, 165–171. [Google Scholar]
- Cooper, M.R. A Revision of the Ornithischian Dinosaur Kangnasaurus coetzeei Haughton, with a Classification of the Ornithischia. Ann. S. Afr. Mus. 1985, 95, 281–317. [Google Scholar]
- Sereno, P.C. Taxonomy, Morphology, Masticatory Function and Phylogeny of Heterodontosaurid Dinosaurs. Zookeys 2012, 226, 1–225. [Google Scholar] [CrossRef]
- Pol, D.; Rauhut, O.W.M.; Becerra, M. A Middle Jurassic Heterodontosaurid Dinosaur from Patagonia and the Evolution of Heterodontosaurids. Naturwissenschaften 2011, 98, 369–379. [Google Scholar] [CrossRef]
- Liyong, J.; Jun, C.; Shuqin, Z.; Butler, R.J.; Godefroit, P. Cranial Anatomy of the Small Ornithischian Dinosaur Changchunsaurus parvus from the Quantou Formation (Cretaceous: Aptian-Cenomanian) of Jilin Province, Northeastern China. J. Vertebr. Paleontol. 2010, 30, 196–214. [Google Scholar] [CrossRef]
- Weishampel, D.B.; Heinrich, R.E. Systematics of Hypsilophodontidae and Basal Iguanodontia (Dinosauria: Ornithopoda). Hist. Biol. Int. J. Paleobiol. Hist. Biol. 1992, 63, 159–184. [Google Scholar] [CrossRef]
- Brown, C.M.; Evans, D.C.; Ryan, M.J.; Russell, A.P. New Data on the Diversity and Abundance of Small-Bodied Ornithopods (Dinosauria, Ornithischia) from the Belly River Group (Campanian) of Alberta. J. Vertebr. Paleontol. 2013, 33, 495–520. [Google Scholar] [CrossRef]
- Scheetz, R.D. Osteology of Orodromeus makelai and the Phylogeny of Basal Ornithopod Dinosaurs; Montana State University: Bozeman, MT, USA, 1999. [Google Scholar]
- Norman, D.B.; Sues, H.-D.; Witmer, L.M.; Coria, R.A. Basal Ornithopoda. In The Dinosauria; Weishampel, B.D., Dodson, P., Osmólska, H., Eds.; University of California Press: Oakland, CA, USA, 2004; pp. 393–412. [Google Scholar]
- Barrett, P.M.; Feng-Lu, H.A.N. Cranial Anatomy of Jeholosaurus shangyuanensis (Dinosauria: Ornithischia) from the Early Cretaceous of China. Zootaxa 2009, 55, 31–55. [Google Scholar] [CrossRef]
- Makovicky, P.J.; Kilbourne, B.M.; Sadleir, R.W.; Norell, M.A.; Makovicky, P.J.; Kilbourne, B.M.; Sadleir, R.W.; Norell, M.A. A New Basal Ornithopod (Dinosauria, Ornithischia) from the Late Cretaceous of Mongolia. J. Vertebr. Paleontol. 2011, 31, 626–640. [Google Scholar] [CrossRef]
- Barta, D.E.; Norell, M.A. The Osteology of Haya griva (Dinosauria: Ornithischia) from the Late Cretaceous of Mongolia. Bull. Am. Mus. Nat. Hist. 2021, 445, 1–112. [Google Scholar] [CrossRef]
- Huh, M.; Lee, D.G.; Kim, J.K.; Lim, J.D.; Godefroit, P. A New Basal Ornithopod Dinosaur from the Upper Cretaceous of South Korea. Neues Jahrb. Fur Geol. Und Palaontol.-Abh. 2011, 259, 1–24. [Google Scholar] [CrossRef]
- Galton, P.M. Notes on the Postcranial Anatomy of the Heterodontosaurid Dinosaur Heterodontosaurus tucki, a Basal Ornithischian from the Lower Jurassic of South Africa. Rev. Paleobiol. 2014, 33, 97–141. [Google Scholar]
- Baron, M.G.; Norman, D.B.; Barrett, P.M. Postcranial Anatomy of Lesothosaurus diagnosticus (Dinosauria: Ornithischia) from the Lower Jurassic of Southern Africa: Implications for Basal Ornithischian Taxonomy and Systematics. Zool. J. Linn. Soc. 2017, 179, 125–168. [Google Scholar] [CrossRef]
- Galton, P.M.; Jensen, J.A. Skeleton of a Hypsilophodontid Dinosaur (Nanosaurus (?) rex) from the Upper Jurassic of Utah. Brigh. Young Univ. Geol. Ser. 1973, 20, 137–157. [Google Scholar]
- Andrzejewski, K.A.; Winkler, D.A.; Jacobs, L.L. A New Basal Ornithopod (Dinosauria: Ornithischia) from the Early Cretaceous of Texas. PLoS ONE 2019, 14, e0207935. [Google Scholar]
- Zheng, W.; Jin, X.; Shibata, M.; Azuma, Y.; Yu, F. A New Ornithischian Dinosaur from the Cretaceous Liangtoutang Formation of Tiantai, Zhejiang Province, China. Cretac. Res. 2012, 34, 208–219. [Google Scholar] [CrossRef]
- Brown, C.M.; Boyd, C.A.; Russell, A.P. A New Basal Ornithopod Dinosaur (Frenchman Formation, Saskatchewan, Canada), and Implications for Late Maastrichtian Ornithischian Diversity in North America. Zool. J. Linn. Soc. 2011, 163, 1157–1198. [Google Scholar] [CrossRef]
- Butler, R.J.; Galton, P.M. The “dermal Armour” of the Ornithopod Dinosaur Hypsilophodon from the Wealden (Early Cretaceous: Barremian) of the Isle of Wight: A Reappraisal. Cretac. Res. 2008, 29, 636–642. [Google Scholar] [CrossRef]
- He, Y.; Makovicky, P.J.; Wang, K.; Chen, S.; Sullivan, C.; Han, F.; Xu, X. A New Leptoceratopsid (Ornithischia, Ceratopsia) with a Unique Ischium from the Upper Cretaceous of Shandong Province, China. PLoS ONE 2015, 10, e0144148. [Google Scholar] [CrossRef] [PubMed]
- Varricchio, D.J.; Martin, A.J.; Katsura, Y. First Trace and Body Fossil Evidence of a Burrowing, Denning Dinosaur. Proc. R. Soc. B Biol. Sci. 2007, 274, 1361–1368. [Google Scholar] [CrossRef] [PubMed]
- Galton, P. Hypsilophodon, the Cursorial Non-Arboreal Dinosaur. Nature 1971, 231, 159–161. [Google Scholar] [CrossRef] [PubMed]
- Forster, C.A. The Postcranial Skeleton of the Ornithopod Dinosaur Tenontosaurus tilletti. J. Vertebr. Paleontol. 1990, 10, 273–294. [Google Scholar] [CrossRef]
- Gilmore, C.W. Osteology of the Jurassic Reptile Camptosaurus, with a Revision of the Species of the Genus, and Descriptions of Two New Species. Proc. Natl. Mus. 1909, 36, 197–332. [Google Scholar] [CrossRef]
- Carpenter, K. A New Species of Camptosaurus (Ornithopoda: Dinosauria) from the Morrison Formation (Upper Jurassic) of Dinosaur National Monument, Utah, and a Biomechanical Analysis of Its Forelimb. Ann. Carnegie Mus. 2008, 76, 227–264. [Google Scholar] [CrossRef]
- Norman, D.B. On the Ornithischian Dinosaur Iguanodon bernissartensis of Bernissart (Belgium). Mémoires L’institut R. Des Sci. Nat. Belg. 1980, 1980, 1–103. [Google Scholar]
- Carpenter, K.; Galton, P.M. A Photo Documentation of Bipedal Ornithischian Dinosaurs from the Upper Jurassic Morrison Formation, USA. Geol. Intermt. West 2018, 5, 167–207. [Google Scholar] [CrossRef]
- Butler, R.J. The Anatomy of the Basal Ornithischian Dinosaur Eocursor Parvus from the Lower Elliot Formation (Late Triassic) of South Africa. Zool. J. Linn. Soc. 2010, 160, 648–684. [Google Scholar] [CrossRef]
- Galton, P.M. Notes on Neocomian (Lower Cretaceous) Ornithopod Dinosaurs from England—Hypsilophodon, Valdosaurus, “Camptosaurus”, “Iguanodon”—And Referred Specimens from Romania and Elsewhere. Genève 2009, 28, 211–273. [Google Scholar]
- Chanthasit, P.; Suteethorn, S.; Naksri, W.; Tong, H.; Wongko, K.; Sonoda, T. New Vertebrate Fossil Site from the Early Cretaceous Sao Khua Formation, Sakon Nakhon Province, Northeastern Thailand. Open J. Geol. 2019, 9, 619–622. [Google Scholar] [CrossRef]
- Buffetaut, E.; Suteethorn, V.; Tong, H. Dinosaur Assemblages from Thailand: A Comparison with Chinese Faunas. In Proceedings of the Papers from the 2005 Heyuan International Dinosaur Symposium, Heyuan Shi, China, 11 April 2015; Lu, J.C., Kobayashi, Y., Huang, D., Lee, Y.-N., Eds.; Geological Publishing House: Beijing, China, 2006; pp. 19–37. [Google Scholar]
- Racey, A.; Goodall, J.G.S. Palynology and Stratigraphy of the Mesozoic Khorat Group Red Bed Sequences from Thailand. Geol. Soc. Lond. Spec. Publ. 2009, 315, 69–83. [Google Scholar] [CrossRef]
- Tong, H.; Claude, J.; Naksri, W.; Suteethorn, V.; Buffetaut, E.; Khansubha, S.; Wongko, K.; Yuangdetkla, P. Basilochelys macrobios n. Gen. and n. Sp., a Large Cryptodiran Turtle from the Phu Kradung Formation (Later Jurassic-Earliest Cretaceous) of the Khorat Plateau, NE Thailand. Geol. Soc. Spec. Publ. 2009, 315, 153–173. [Google Scholar] [CrossRef]
- Lauprasert, K.; Laojumpon, C.; Saenphala, W.; Cuny, G.; Thirakhupt, K.; Suteethorn, V. Atoposaurid Crocodyliforms from the Khorat Group of Thailand: First Record of Theriosuchus from Southeast Asia. Palaontol. Z. 2011, 85, 37–47. [Google Scholar] [CrossRef]
- Martin, J.E.; Lauprasert, K.; Buffetaut, E.; Liard, R.; Suteethorn, V. A Large Pholidosaurid in the Phu Kradung Formation of North-Eastern Thailand. Palaeontology 2014, 57, 757–769. [Google Scholar] [CrossRef]












| Element | Measurement (mm) | |
|---|---|---|
| Cervical vertebrae (L) 8?, 9? | Length of ribs | 20.12, +23.10 |
| Dorsal vertebrae (L) 1–10 | Length of ribs | 49.85, 62.06, 66.54, +54.45, 65.53, 58.64, 45.98, 32.50, 17.83, 13.73 |
| Dorsal vertebrae (R) 12–15 | Length of centra | 10.54, 10.16, 10.76, 10.78 |
| Sacral vertebrae (R) 1–5 | Length of centra | 12.03, ?, ?, ?, 10.29 |
| Caudal vertebrae (L) 1–10 | Length of centra | 10.00, 9.98, 9.96, 9.74, 9.25, 9.25, 10.97, 9.02, 9.38, 10.22 |
| Scapula (L) | Length Proximal width Distal width Smallest diameter of shaft | +53.63 18.0 25.70 10.44 |
| Ilium (L) | Length Height Breadth of acetabulum | 70.37 14.66 7.26 |
| Pubis (L) | Length Length of preprocess Length of postprocess | 103.04 42.55 75.66 |
| Ischium (L) | Length Proximal width Distal width Smallest breadth shaft | 65.38 16.41 4.11 3.27 |
| Femur (L) | Length Proximal width Distal width Smallest shaft diameter | 82.08 14.07 19.69 9.78 |
| Tibia (R) | Length Proximal width Distal width Smallest shaft diameter | 99.76 14.72 19.84 7.64 |
| Fibula (L) | Length Proximal width Distal width Smallest shaft diameter | 95.41 9.64 7.38 3.74 |
| Metatarsal (L) ?III,?IV | Length | 53.83, 47.55 |
| Pes phalanx (?R) | Length | +20.55 |
| Pes ungual (?R) | Length | 11.12 |
| Taxa | Premaxilla | Maxilla | Enamel Distribution | Mandible | Enamel Distribution |
|---|---|---|---|---|---|
| Pisanosaurus mertii | ? | 11+ (17–18?) | symmetry | 15 | symmetry |
| Heterodontosaurus tucki | 3 | 12 | asymmetry | ? | asymmetry |
| Echinodon becklessii | 3 | ~11 | symmetry | 10 | symmetry |
| Tatisaurus oehleri | ? | ? | symmetry | 18 | symmetry |
| Lesothosaurus diagnosticus | 6 | ~14 | symmetry | ~14 | symmetry |
| Agilisaurus louderbacki | 5 | 14 | symmetry | 20 | symmetry |
| Hexinlusaurus multidens | ? | 18 | symmetry | 20 | symmetry |
| Xiaosaurus dashanpensis | ? | ? | symmetry | ? | ? |
| Yandusaurus hongheensis | ? | 15 | asymmetry | ? | ? |
| Minimocursor phunoiensis | ? | ? | ? | ≥12 | asymmetry |
| Nanosaurus agilis | ? | ? | symmetry | 13 | symmetry |
| Hypsilophodon foxii | 5 | 10–11 | asymmetry | 13–14 | asymmetry |
| Convolosaurus marri | 4 | 8–10 | asymmetry | 11 | asymmetry |
| Dysalotosaurus lettowvorbecki | 0 | 13 | asymmetry | 11–12 | asymmetry |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manitkoon, S.; Deesri, U.; Khalloufi, B.; Nonsrirach, T.; Suteethorn, V.; Chanthasit, P.; Boonla, W.; Buffetaut, E. A New Basal Neornithischian Dinosaur from the Phu Kradung Formation (Upper Jurassic) of Northeastern Thailand. Diversity 2023, 15, 851. https://doi.org/10.3390/d15070851
Manitkoon S, Deesri U, Khalloufi B, Nonsrirach T, Suteethorn V, Chanthasit P, Boonla W, Buffetaut E. A New Basal Neornithischian Dinosaur from the Phu Kradung Formation (Upper Jurassic) of Northeastern Thailand. Diversity. 2023; 15(7):851. https://doi.org/10.3390/d15070851
Chicago/Turabian StyleManitkoon, Sita, Uthumporn Deesri, Bouziane Khalloufi, Thanit Nonsrirach, Varavudh Suteethorn, Phornphen Chanthasit, Wansiri Boonla, and Eric Buffetaut. 2023. "A New Basal Neornithischian Dinosaur from the Phu Kradung Formation (Upper Jurassic) of Northeastern Thailand" Diversity 15, no. 7: 851. https://doi.org/10.3390/d15070851
APA StyleManitkoon, S., Deesri, U., Khalloufi, B., Nonsrirach, T., Suteethorn, V., Chanthasit, P., Boonla, W., & Buffetaut, E. (2023). A New Basal Neornithischian Dinosaur from the Phu Kradung Formation (Upper Jurassic) of Northeastern Thailand. Diversity, 15(7), 851. https://doi.org/10.3390/d15070851