Abstract
Three new species of the rare deep-sea family Haplomunnidae are described from the abyssal of the Northwestern (NW) Pacific adjacent to the Kuril–Kamchatka Trench (KKT) based on material collected during the deep-sea expeditions KuramBio (2012) and SokhoBio (2015). Previously, only three species of the genus Haplomunna Richardson, 1908 were described in the whole North Pacific, including one species from its western part. Haplomunna kurilensis Golovan and Malyutina sp. nov., closely resembling H. japonica (Gamô, 1983) from the upper hadal of the Japan Trench, was the most numerous haplomunnid species in the studied region. Thylakogaster wilsoni Malyutina and Golovan sp. nov., and Abyssaranea minuta Golovan and Malyutina sp. nov., are the first species of their genera described from the Pacific Ocean. For H. kurilensis sp. nov. and T. wilsoni sp. nov., DNA sequences were obtained for 18S rRNA and 28S rRNA genes. The phylogenetic relationship between these species, as well as their position within the Haplomunnidae, were reconstructed using 18S rDNA.
1. Introduction
The Haplomunnidae are a small family of deep-sea janiroidean asellotes. Their members are characterized by a very unusual habitus, including a vaulted body with axially compressed and reduced posterior pereonites, which are overlapped on the dorsal side by a large pleotelson, except for the genus Munella. The compact body is accompanied by a short subchelate pereopod I and long walking pereopods II–VII, as well as small reduced uropods [1,2]. The closest relatives of Haplomunnidae are representatives of another unusual janiroidean family Dendrotionidae [1,3]. Wilson [1] assumed that both families had developed from a long-legged, shallow-water janirid-like ancestor and radiated in the deep sea. During their evolution, representatives of the line leading to Thylalcogaster took the path of specialization in the pereopods and mouthparts and acquired an extremely enlarged pleotelson. The other line, which followed the path of reduction in the size of the antenna and in the seventh pereopod and pereonite, eventually led to very specialized Abyssaranea [1]. It is noticeable that the reduction of the last pereonite and its appendages seems to be a common trend, occurring independently in different taxa of the deep-sea Janiroidea [4,5,6].
Representatives of the Haplomunnidae lead an epibenthic lifestyle, inhabiting differing types of substrates, from soft clays and silts to volcanic gravels, manganese nodules, and rocky outcrops, including extreme conditions of underwater volcanoes and hydrotherms [2,7]. Cunha and Wilson [2] suggested that haplomunnids may prefer biotic substrates. For example, they were found highly aggregated on stalks of hexactinellid sponges [2,8].
The family was erected in 1976 by Wilson [1] to combine four genera of asellotes, i.e., Haplomunna Richardson, 1908, Munella Bonnier, 1896, Abyssaranea Wilson and Hessler, 1974, and Thylakogaster Wilson and Hessler, 1974, previously considered within the families Munnidae and Dendrotionidae. But two years earlier, Wilson and Hessler [9] had already mentioned a “confusing cluster” grouped within these families, which includes the last three genera. In 1983, Gamô described the monotypic genus Aryballurops, including A. japonica Gamô, 1983 [10]. He noted the similarity of the new genus to the genus Thylakogaster but placed it into the family Munnidae. In 2003, Cunha and Wilson [2] provided a comprehensive overview of the family Haplomunnidae, where they synonymized Aryballurops with Haplomunna. In the same year, Just [11] described the monotypic genus Haplodendron Just, 2003, including H. buzwilsoni Just, 2003. Therefore, the family Haplomunnidae currently includes five genera and eleven described species [2,12,13]. In addition, findings of undescribed or unidentified haplomunnid species have been reported from different regions of the World Ocean [2,8,14,15] (see also: https://obis.org, accessed on 2 June 2023).
Although the deep-sea Janiroidea, in general, is a common and abundant component of the abyssal benthic communities, Haplomunnidae are rare in samples and usually presented by a few individuals [16,17,18,19]. Most species are known by one or a few specimens only. In addition, due to their fragility, haplomunnids are often strongly damaged during a sampling process [2,9,11]. Descriptions of some species are incomplete because of the lack of specimens with a full set of appendages in the collections. Therefore, each new finding of Haplomunnidae, especially in good condition, is of great value, adding new information about this group of crustaceans. Moreover, only four specimens of Haplomunnidae were studied with molecular data. All of them were identified as Thylakogaster sp. For one of them, a fragment of mitochondrial COI gene sequence was obtained [20], and for the other three specimens, 18S and 28S gene sequences were obtained [3,21].
Despite their rarity, the Haplomunnidae family, like other deep-sea families of Janiroidea, has a very wide distribution. They are found in all the oceans, except the Indian and the Arctic Oceans [2] (see also https://obis.org, accessed on 2 June 2023). However, hypothetically, the absence of the records of this rare family in some basins may also be due to the fact that the local deep-sea fauna is insufficiently studied or the biotopes preferred by these isopods are difficult to access [22,23]. For example, the hard rock outcrops turned out to be a fairly common biotope, occurring as patchy in the deep sea [24]. They are associated with their own fauna (including organisms on which haplomunnids can settle), which is poorly studied due to the technical difficulties of collecting material [22,23,25]. In addition, due to their fragility, members of the family may remain unidentified in collections. Haplomunnids have been recorded at depths ranging from 950 m (or even from 100 m, according to unconfirmed data) to 6450 m [1,2,10,26]. Most species have been collected from the abyssal (six species) and lower bathyal (four species) depths. The deepest finding was that of Haplomunna japonica (Gamô, 1983), described from the upper boundary of the Japan Trench (6380–6450 m) [10,26].
Data on the haplomunnid fauna of the Pacific Ocean are very fragmentary. Only three species of the genus Haplomunna are described in the northern part, and one species of Haplodendron in the Southern Hemisphere [1,2,10,11,27]. In addition, several members of the genera Haplomunna, Thylakogaster, Munella (unidentified and potentially new species to science), and Abyssaranea cf. rupis have been recorded in the abyssal of the NE Pacific [2,8].
In the NW Pacific, a small collection of haplomunnids was made from several stations in the abyssal zone adjacent to the Kuril–Kamchatka Trench (KKT) during the deep-sea expeditions KuramBio (2012) and SokhoBio (2015) [14,15,28,29]. Further taxonomic processing of the material of these expeditions, available for our examination, revealed three species of Haplomunnidae that turned out to be new to science.
2. Material and Methods
2.1. Material Sampling
The material used in this study was collected in 2012 at the abyssal plain of the NW Pacific Basin to the east of the Kuril–Kamchatka Trench (KKT) during the German–Russian expedition KuramBio (Kurile Kamchatka Biodiversity Studies) on board the RV Sonne [28], and in 2015, in the abyssal to the west of the KKT during the Russian–German expedition SokhoBio (Sea of Okhotsk Biodiversity Studies) on board the RV Akademik M.A. Lavrentyev [29]. During both expeditions, macrobenthos was sampled with the use of a camera epibenthic sledge C-EBS as described by Brandt and coauthors [16,17]. Samples were washed onboard with cold filtered sea water immediately after hauling through sieves of 300 μm mesh and fixed in pre-cooled 96% ethanol; see [30] for the details. The studied specimens of Haplomunnidae were collected at four stations at depths ranging from 4769 to 5380 m (Table 1, Figure 1).

Table 1.
List of the KuramBio (KB) and SokhoBio (SO) stations, where Haplomunnidae were sampled.
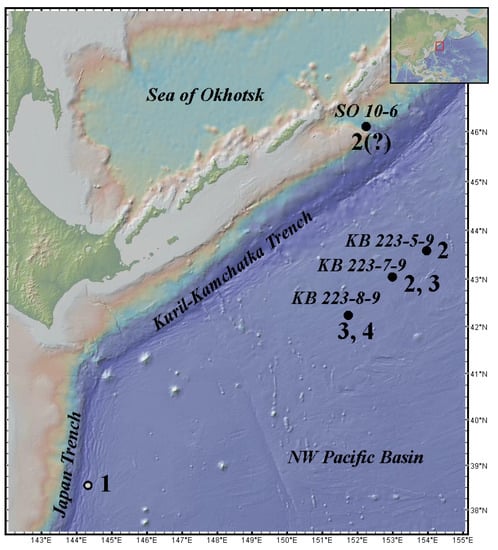
Figure 1.
Distribution of Haplomunnidae species in the Northwest (NW) Pacific: (1) Haplomunna japonica (Gamô, 1983); (2) Haplomunna kurilensis Golovan and Malyutina sp. nov.; (3) Thylakogaster wilsoni Malyutina and Golovan sp. nov.; (4) Abyssaranea minuta Golovan and Malyutina sp. nov.; Black circle—original data; white circle—after Gamô (1983). Data for stations are given in Table 1.
The map was generated using GeoMapApp 3.6.14 (www.geomapapp.org, accessed on 30 August 2022).
2.2. Material Processing and Species Description
Photographs of paramunnids were taken using a Canon EOS 80D camera with an MP-E 65 mm macro lens, installed on a Kaiser Copy Stand RS 2 XA in Figure 2A–D,F–I, and a Nikon D5300 camera mounted on a Zeiss Discovery v.12 stereoscopic dissection microscope with a Zeiss T2-T2 DSLR 1.6x camera adapter in Figure 2E,J.

Figure 2.
Haplomunnidae from the Kuril–Kamchatka trench (KKT) area, collected during the KuramBio expedition. (A–F) Haplomunna kurilensis Golovan and Malyutina sp. nov.: (A,B) male, holotype, lateral and dorsal view (SMF 61311); (C,D) female, paratype, dorsal and lateral view; (E) female pleon, ventral view; (F) manca (MIMB 46614). (G–I) Thylakogaster wilsoni Malyutina and Golovan sp. nov.: (G,H) female, holotype, lateral and dorsal view (SMF 61314); (I) manca (MIMB 46618). (J) Abyssaranea minuta Golovan and Malyutina sp. nov.: female, holotype (SMF 61310). Scale bar = 1 mm.
The best-preserved male specimen of Haplomunna kurilensis Golovan and Malyutina sp. nov. was designated as the holotype. The whole view of the holotype and the pleon of the only female paratype were drawn without preparation. Two male paratypes of this species from the same station as holotype were used for the dissection. Appendages were drawn from the dissected specimens. The best-preserved specimen of Thylakogaster wilsoni Malyutina and Golovan sp. nov. was designated as the holotype. The whole views of the holotype and pereopods were drawn without preparation. Mouth parts and pleopods were drawn from the dissected paratype specimen. Since Abyssaranea minuta Golovan and Malyutina sp. nov. was represented by a single specimen, all appendages that could be studied without destroying the holotype were drawn in situ. For this, the whole animal was placed in a drop of glycerin on a glass slide. Appendages that could not be exposed without dissection were not examined. The exception was the pereopod VI basis, which was carefully cut off and preserved for genetic studies. The drawings were made using a Zeiss Discovery v.12 stereoscopic microscope and an Olympus CX31RTSF microscope, both equipped with a camera lucida, inked using a Wacom Cintiq 22HD interactive display, and processed using Adobe Photoshop CC.
Terminology, measurements, and numeration of body segments and appendages follow [1,9,11,31]. Since in members of Haplomunna and Thylakogaster, the pleotelson is strongly reflected anteriorly, lying above the pereonites. Body length was measured in lateral view from the tip of the clypeus to the posterior lateral margin of the pereonite 7, without pleotelson (as in the cited above studies). Additionally, we gave the total length of the body, including the pleotelson, to make the description more consistent with the descriptions of other Janiroidea. Since the body is curved, the lengths of the pereonites were measured in lateral view (unless otherwise mentioned).
The type material is deposited in the Senckenberg Museum, Frankfurt am Main, Germany (SMF) and in the Museum of the NSCMB FEB RAS, formerly Institute of Marine Biology (IMB) FEB RAS, Vladivostok, Russia (MIMB).
Abbreviations are as follows:
BS—broom seta, DSS—distally setulated seta, SS—simple seta, UBS—unequally bifid seta, UBDS—unequally bifid distally setulated seta, NE—Northeast, NW—Northwest, KKT—Kuril–Kamchatka Trench.
2.3. Molecular Analysis
Total DNA was extracted from whole specimens or one pereopod by heating in 10% solution of Chelex 100 Resin (Chelating Ion Exchange Resin, Bio-Rad) [32] at 95 °C for 20 min. Nuclear small-subunit ribosomal RNA (18S rRNA) and large-subunit ribosomal RNA (28S rRNA) genes were amplified and sequenced using universal invertebrate pairs of primers (Table 2). The PCR (polymerase chain reaction) was performed with Dream Taq Hot Start Green PCR Master Mix (2x) (Thermo Fisher Scientific Baltics UAB, Vilnus, Lithuania) according to the manufacturer’s protocol and annealing temperature 52 °C. PCR products were checked by electrophoresis with 1.5% agarose gel. Amplification products were applied as templates for sequencing, using the same primers as the PCR and BrilliantDye™ Terminator Cycle Sequencing kit v3.1 (NimaGen, Nijmegen, The Netherlands) according to the manufacturer’s protocol. Sequencing reaction products were purified by ethanol precipitation and analyzed on an ABI-3500 Genetic Analyzer (Applied Biosystems, Thermo Fisher Scientific Inc., Foster City, CA, USA). Sequences were verified by forward and reverse comparisons. The contigs were obtained and edited using SeqScape 2.5 (Applied Biosystems). Primer sequences were excluded from the analysis. A BLAST search (https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 2 June 2023) was used to check new sequences against the database for possible contamination and sequence artifacts. All new DNA sequences have been deposited in GenBank (Table 3).
Table 2.
Sequencing primers for polymerase chain reactions.
Table 3.
Genetic analysis information with GenBank accession numbers (acc.) for Haplomunnidae collected during the expeditions KuramBio (KB) and SokhoBio (SO).
Unfortunately, sequences of 28S rRNA, which we obtained in the current analysis, did not overlap with other sequences for this marker for the family Haplomunnidae from GenBank, so we did not use them for phylogenetic reconstruction. In order to construct the 18S phylogram, the sequences of the species of Haplomunnidae from GenBank were comprised in the analysis. Sequences of the related janiroidean families Munnidae and Dendrotionidae were added for phylogenetic reconstructions for this marker. We also included some members of the family Janiridae in the analysis as sister taxa to Dendrotionidae and Haplomunnidae. The sequence of Stenasellus racovitzui Razzauti, 1925 (Stenasellidae, Aselloidea) was selected as the outgroup for reconstructing the trees.
Sequences were aligned using MUSCLE [38] implemented in the MEGA X program [39]. The quality of alignment was checked visually. The model of nucleotide substitution for the tree was selected using jModelTest v. 2.1.6 [40] implemented in CIPRES Science Gateway (http://www.phylo.org/, accessed on 2 June 2023) [41] under Akaike Information Criterion (AIC). The best-fit model was TIM3+I+G. In order to construct the Bayesian (BI) tree, MrBayes 3.2.7a [42] was used and implemented in CIPRES Science Gateway (http://www.phylo.org/, accessed on 2 June 2023) [41] with the following parameters: 10,000,000 generations, with four parallel chains and sample frequencies set to 500 in two separate runs. Based on the convergence of likelihood scores, 25% of the sampled trees were discarded as burn-in. The results tree was visualized using FigTree v. 1.4.4 [42]. Uncorrected genetic distances were calculated using MEGA-X [39]. All estimations of sequences and distances were performed, excluding the outgroup.
3. Results
In the material collected at the four stations in the abyssal adjacent to the Kuril–Kamchatka Trench (KKT) at depths ranging within 4769–5376 m, 28 specimens of the family Haplomunnidae were sampled. They represented three species belonging to three genera. All the species appeared to be new to science (Table 4, Figure 1 and Figure 2).
Table 4.
List of the Haplomunnidae found at the four C-EBS stations during the expeditions KuramBio (KB) and SokhoBio (SO).
Haplomunna kurilensis Golovan and Malyutina sp. nov. was the most numerous species: twenty-one specimens were sampled at three stations located on both sides of the trench. Most of them (19 specimens) were found at the station KB 223-7-9, located in the NW Pacific Basin to the east of the KKT at a depth of 5222–5223 m. Besides these specimens, one male in a good condition was sampled at the nearby station KB 223-5-9 at a depth of 5376–5380 m. In addition, one manca assigned to this species was sampled at the station SO 10-6, located in the abyssal off the KKT western slope at a depth of 4769–4798 m. Considering that this specimen was immature and the attempt to obtain molecular data from it failed, its conspecificity with other specimens of H. kurilensis sp. nov. is not clearly confirmed. It is noteworthy that most of the collected individuals of H. kurilensis sp. nov. were mancas (12) and males (5), while only one female was present at the richest station KB 223-7-9.
Three females and one manca of Thylakogaster wilsoni Malyutina and Golovan sp. nov. and one female of Abyssaranea minuta Golovan and Malyutina sp. nov. were collected at the station KB 223-8-9 to the east of the KKT at a depth of 5125–5126 m. Two mancas of T. wilsoni sp. nov. were sampled at the station KB 223-7-9 at a depth of 5222–5223 m.
The descriptions of the new species are given below.
3.1. Taxonomy
Order Isopoda Latreille, 1817
Suborder Asellota Latreille, 1802
Superfamily Janiroidea G.O. Sars, 1897
Family Haplomunnidae Wilson, 1976
3.1.1. Genus Abyssaranea Wilson and Hessler, 1974
Abyssaranea Wilson and Hessler, 1974: 58.
Type species. Abyssaranea rupis Wilson and Hessler, 1974, by monotypy.
Species included. Abyssaranea minuta Golovan and Malyutina sp. nov., A. rupis Wilson and Hessler, 1974.
Diagnosis. See [9].
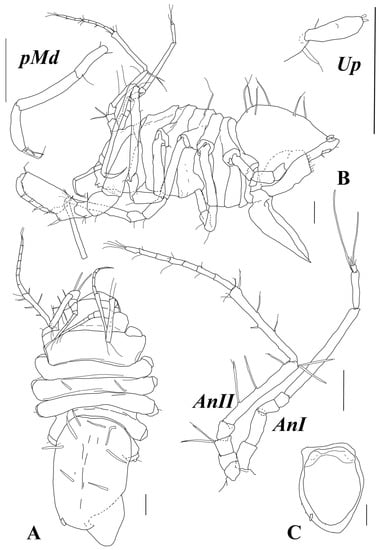
Figure 3.
Abyssaranea minuta Golovan and Malyutina sp. nov., female, holotype (SMF 61310): (A) dorsal view; (B) lateral view; (C) pleotelson, ventral view; (AnI and AnII) antennae I and II; (pMd) mandibular palp; (Up) uropod. Scale bar = 0.1 mm.
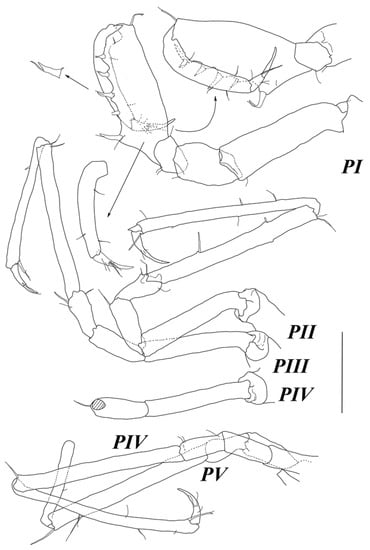
Figure 4.
Abyssaranea minuta Golovan and Malyutina sp. nov., female, holotype (SMF 61310): (PI–V) pereopods I–V. Scale bar = 0.1 mm.
This species is registered in ZooBank under urn:lsid:zoobank.org:act:23F22647-F7C7-4EB5-9B6C-7659C802ACB2.
Material examined. Holotype: (SMF 61310) female (total length 1.1 mm), KB 223-8-9, designated here.
Etymology. The new species name is derived from the Latin adjective “minutus” that means “small, tiny”.
Diagnosis. Female. Dorsal surface lacking spines. Pereonite 1 length 0.3 width and 1.6 of pereonite 2 length. Pleotelson length 1.5 width, with about eight long UBS dorsally and one stout UBS on each lateral margin anteriorly to the place of uropod insertion. Pereopod I carpus is as long as the basis, with length 2.7 width; dactylus and propodus together are about as long as the carpus. Operculum length 1.2 width.
Description. Holotype, female: body (Figure 2J and Figure 3A,B) total length 2.0 width of pereonite 3; body thickest and deepest at pereonite 3; depth 0.38 total body length. Total length 1.1 mm, cephalothorax–pereon length 0.76 mm. Spines absent.
Cephalothorax: length 0.68 width and 0.2 total body length, width 0.7 width of pereonite 1; frons and clypeus rounded in dorsal view.
Pereonite 1 length 0.28 width and 0.12 body length; pereonite 6 visible in dorsal view as a thin belt; pereonites 2–6 lengths to pereonite 1 length ratios: 0.6; 0.7; 0.5; 0.3; 0.3; length to width ratios: 0.15; 0.16; 0.12; 0.09. Dorsal surface of pereonite 3 with pair of long submedial UBS.
Pleotelson length 1.5 width and 0.46 body length, oval in dorsal view, and posterior tip rounded. Dorsal surface with about eight long UBS; lateral margin with one stout UBS anteriorly to the place of uropod insertion.
Antenna I (Figure 3) with seven articles; articles 2–7 lengths to article 1 length ratios: 1.2; 0.64; 0.50; 3.9; 1.2; 0.23; articles 1–7 length to width ratios: 2.0; 2.4; 1.5; 1.2; 10; 4.9; 1.2. Articles 6 and 7, each with one aestetasc; article 7 with five fine setae terminally.
Antenna II (Figure 3) articles 2–6 lengths to article 1 length ratios: 1.1; 1.1; 1.2; 6.4; 6.5; articles 1–6 length to width ratios: 0.86; 0.89; 0.92; 1.1; 7.6; 7.9. Article 3 with two long distoventral UBS; article 5 with three ventral and two distodorsal long UBS; article 6 with four UBS dorsally, one UBS, and one trace of seta insertion ventrally and few fine setae distoventrally. Flagellum length 1.2 length of article 6, with seven articles bearing fine SS.
Mandibular palp (Figure 3) about 0.2 of total body length. Article 1 length about 3.8 width, with one distoventral seta. Articles 2 and 3 lengths about 5 and 4 widths, and 1.6 and 1.1 article 1 length, respectively. Article 3 with a row of few setae.
Maxilliped with two retinaculae.
Pereopod I (Figure 4) basis–dactylus length to width ratios: 3.4; 1.6; 1.4; 2.7; 6.1; 4.1; ischium–dactylus to basis length ratios: 0.47; 0.29; 1.0; 0.75; 0.32. Basis and ischium, each with one fine SS distoventrally. Merus with two setae ventrally and one SS distodorsally. Carpus ventral margin with a row of five strong UBS opposing propodus; proximal UBS largest, located on blunt projection and accompanied with one SS; distodorsal angle with one long UBS; few SS on lateral margin. Propodus with one dorsal and two ventral SS. Dactylus with few SS on lateral margin; claw 0.57 dactylus length, with two slender setae at base of claw.
Pereopods II–V (Figure 2J, Figure 3B and Figure 4) slightly longer than body, similar in shape. Bases length about 5–6 width. Ischia length about 2.5–3 width and about half of bases length, with distoventral seta. Meri length about 2.3 width and 0.4 bases length, with few setae distally. Carpi length about 10 width and about 1.5 bases length, with 1–2 distodorsal setae; in pereopod 2 with two stout setae on ventral margin and one trace of seta insertion on dorsal margin. Propodi length about 14 width and about as long as carpi, with few fine setae, 1–2 UBS dorsally, and one long stout UBS distoventrally at the base of dactylus insertion. Dactyli length about 4–5 width and about 0.3–0.4 bases length; claw length about 0.4 dactylus length. Pereopod VI broken off, except basis.
Operculum (Figure 3B,C) length 1.4 width; convex ventrally.
Pleopods III–V not studied.
Uropod (Figure 3) length about 3.0 width, with two setae distally (broken off).
Remarks. The new species can be distinguished from the only known species of the genus A. rupis by the shape and proportions of the body segments and appendages. In general, A. minuta sp. nov. has a slenderer and less vaulted body than A. rupis. Thus, the new species has a relatively short pereonite 1 (0.3 times longer than width, 1.6 times longer than pereonite 2) compared to that of A. rupis (0.4 times longer than width, 2 times longer than pereonite 2). This character is associated with a slenderer pereopod I in the new species, whose carpus is 2.7 times longer than its width and equal in length to the basis (vs. carpus 2.2 times longer than its width and 1.3 times longer than basis in A. rupis). The new species has more elongated pleotelson and operculum (each about 1.5 times longer than its width) compared to those in A. rupis (each 1.2 times longer than width). Unlike A. rupis, A. minuta sp. nov. has no spines on the dorsal surface and possesses only stout UBS on the pereonite 3 and pleotelson. Furthermore, the new species differs from A. rupis by 7-articled antenna I (vs. 11-articled in the females of A. rupis) with article 5 almost four times longer than article I (about 1.4 times longer in A. rupis). Nevertheless, this may be due to the immaturity stage of the holotype of the new species.
Wilson and Hessler [9] reported a pronounced sexual dimorphism in A. rupis. The females in A. rupis are larger than males (2.1 mm vs. 1.5–1.7 mm) and possess more sturdy-looking bodies. The female holotype of A. minuta sp. nov. (1.1 mm) is smaller than males of A. rupis. Nevertheless, body size cannot be used as a diagnostic feature for the new species, since only one specimen of A. minuta sp. nov. was studied.
Distribution. The species is known only from the type locality in the NW Pacific Basin to the east of the Kuril–Kamchatka Trench, 42.2447° N 151.7351° E 42.2378° N 151.7082° E, at a depth of 5125–5126 m.
3.1.2. Genus Haplomunna Richardson, 1908
Munna Richardson, 1905: 483 (part).
Haplomunna Richardson, 1908: 79; Menzies, 1962: 32; Wilson, 1976: 574; Cunha and Wilson, 2003: 8.
Aryballurops Gamô, 1983: 13–16, figs 8 and 9.
Type species. Munna caeca Richardson, 1905, by monotypy.
Species included. Haplomunna caeca (Richardson, 1905); H. hubbsi Wilson and Hessler, 1976; H. japonica (Gamô, 1983); H. kurilensis Golovan and Malyutina sp. nov.
Diagnosis. See: [2].
Haplomunna kurilensis Golovan and Malyutina sp. nov.
(Figure 2A–F and Figure 5, Figure 6, Figure 7, Figure 8, Figure 9, Figure 10, Figure 11 and Figure 12)
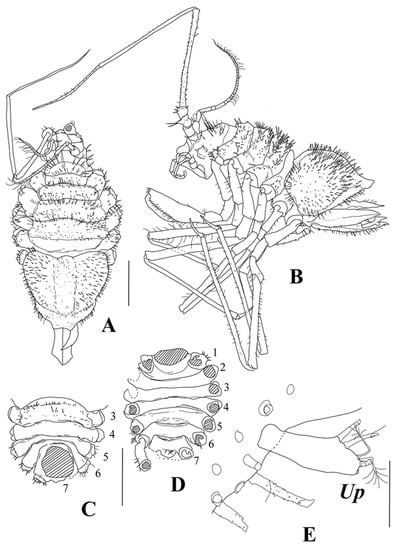
Figure 5.
Haplomunna kurilensis Golovan and Malyutina sp. nov. (A,B,E) male, holotype (SMF 61311): (A,B) dorsal and lateral view, (E) lateral margin of pleotelson with (Up) uropod; (C,D) male 1, paratype (MIMB 46614): (C,D) pereon after dissection, dorsal and ventral view, (1–7) numbers of pereonites. Scale bar = 1 mm (A–D) and 0.1 mm (E).
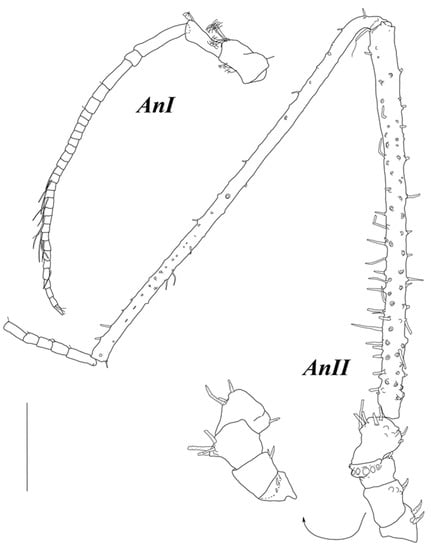
Figure 6.
Haplomunna kurilensis Golovan and Malyutina sp. nov., male 1, paratype (MIMB 46614): (AnI and AnII) antennae I and II. Scale bar = 0.5 mm.
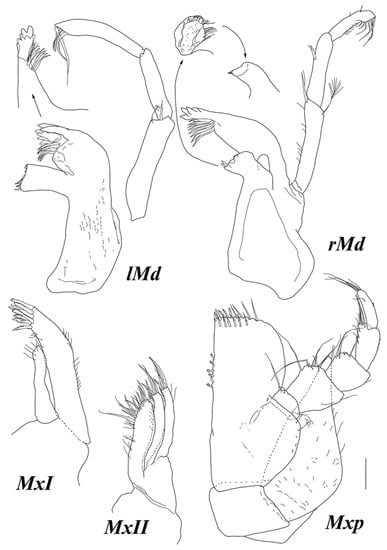
Figure 7.
Haplomunna kurilensis Golovan and Malyutina sp. nov., male paratypes (MIMB 46614). Male 2: (lMd and rMd) left and right mandibles. Male 1: (MxI and MxII) maxillae I and II; (Mxp) maxilliped. Scale bar = 0.1 mm.
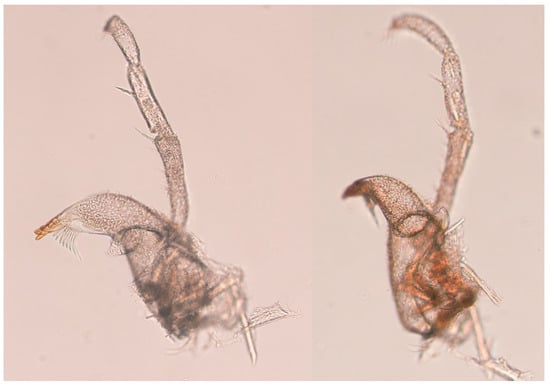
Figure 8.
Haplomunna kurilensis Golovan and Malyutina sp. nov., male 1, paratype (MIMB 46614) right mandible.
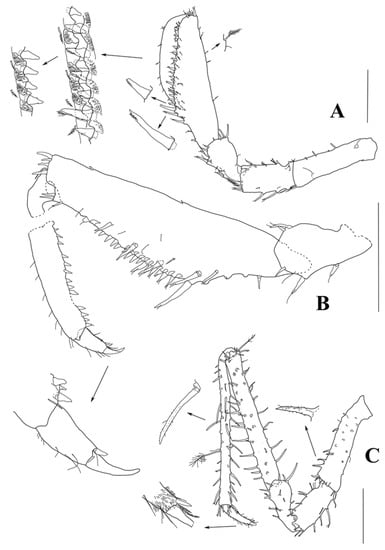
Figure 9.
Haplomunna kurilensis Golovan and Malyutina sp. nov., male paratypes (MIMB 46614). Male 1: (A) pereopod I. Male 2: (B) pereopod I distal part, (C) pereopod II. Scale bar = 0.5 mm.
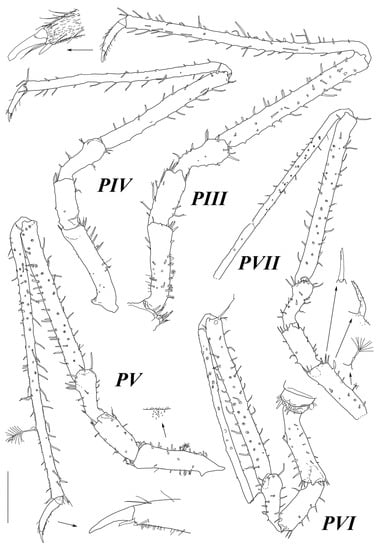
Figure 10.
Haplomunna kurilensis Golovan and Malyutina sp. nov., male paratypes (MIMB 46614). Male 1: (PIII, IV) pereopods III and IV. Male 2: (PV–VII) pereopods V–VII. Scale bar = 0.5 mm.
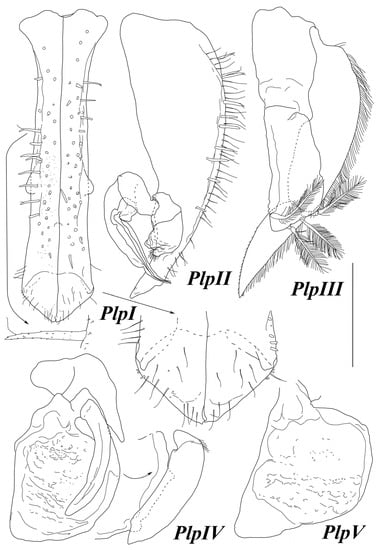
Figure 11.
Haplomunna kurilensis Golovan and Malyutina sp. nov., male 1, paratype (MIMB 46614): (PlpI–V) pleopods I–V. Scale bar = 0.5 mm.
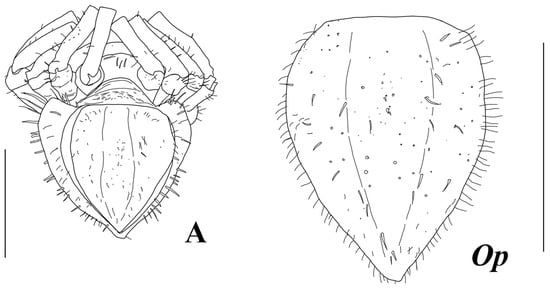
Figure 12.
Haplomunna kurilensis Golovan and Malyutina sp. nov., female, paratype (MIMB 46614): (A) pleotelson, ventral view, (Op) operculum. Scale bar = 1 mm.
This species is registered in ZooBank under urn:lsid:zoobank.org:act:7050DA63-C582-4CC1-AB6C-C978D1D2E8AC.
Material examined. Holotype: (SMF 61311) male (total length 4.4 mm), KB 223-7-9, designated here.
Paratypes: (MIMB 46614) males 1 and 2 (total lengths 4.0 mm), used for dissection, two males (total lengths 3.4 mm), female (total length 3.5 mm), one damaged specimen, six mancas (total lengths 1.3–2.1 mm), the same data as holotype; (SMF 61312) six mancas, the same data as holotype; (SMF 61313) male, KB 223-5-9.
Etymology. The new species is named after the place where it was sampled, i.e., to the east of the Kuril Islands.
Diagnosis. Body surface with numerous thin solitary spines, covered with fine hairs; on dorsomedial parts of each of pereonites 1–3; these spines are grouped on two transverse ridges. Pleotelson inflated, about as long as wide, height 0.8 length, and length 0.6 cephalothorax–pereon length. Maxilliped with 5–6 retinaculae, epipod about as long as basis, narrowing distally. Mandibular palp article 1 with one UBS and one BS distally and about six SS. Pereopod I carpus length about 3 width. Male pleopod I posterior tip pointed, with pronounced distolateral angles.
Description. Holotype, male: body (Figure 2A,B and Figure 5A,B) vaulted, thickest and deepest at pereonite 3, total length 1.9 width, cephalothorax–pereon length 1.4 width, and depth 0.48 cephalothorax–pereon length.
Cephalothorax, pereonites 1–3, and pleotelson dorsal surface with numerous thin spines (Figure 5E), grouped in two transverse rows on medial parts of cephalothorax and pereonites, and evenly arranged on lateral expansions of pereonites and pleotelson. Surface spines of cephalothorax in anterior row are larger than in posterior row. Pereonite 4 with thin spines on dorsal surface of lateral expansions. Pereonites 5–7 with few thin spines laterally. Surface of body and spines with fine setulae, sitting on small tubercle (Figure 5E). In dissected paratype male (Figure 5D), pereonite 7 with a transverse row of small thin spines on ventral surface.
Cephalothorax of holotype length 0.69 width and 0.23 cephalothorax–pereon length, width 0.63 width of pereonite 1; with straight beveled frons.
Pereonites 1–3 lengths about 0.2 widths, pereonites 2 and 3 each 1.1 times longer than pereonite 1. In dissected paratype male with removed pleotelson on dorsal view (Figure 5C), pereonites 4–7 lengths pereonite 3 length ratios: 0.56; 0.14; 0.16; 0.09; on ventral view (Figure 5D), pereonites 2–7 lengths in relation to pereonite 1 length ratios: 0.62; 0.59; 0.71; 1.16; 0.85; 1.2.
Pleotelson about as long as wide, in holotype length 0.61 cephalothorax–pereon length, height 0.80 length, swollen, heart-shaped, medial portion slightly depressed, distal margin flattened, and distal tip bilobed.
Antenna I about 0.84 cephalothorax–pereon length (Figure 5B). In dissected paratype male (Figure 6), articles 1–3 length/width ratios: 1.6; 2.5; 4.1; articles 2–3 lengths to article 1 length ratios: 0.82; 1.0. Article 1 with three small BS medially and five BS distolaterally; article 2 with one UBS and one broken large seta distally. Flagellum with 29 articles, articles 12–23 bearing aestetascs.
Antenna II 2.2 cephalothorax–pereon lengths (Figure 5B). In dissected paratype male (Figure 6), articles 1 and 2 about equal in length, article 3 slightly shorter, and article 4 slightly longer than article 1; articles 5 and 6 lengths 10.5 and 11.7 article 1 length, respectively. Articles 1–4 with thin setulated spines distally; articles 5 and 6 covered with stout setulated setae. Flagellum with more than 15 articles (Figure 5B), in dissected specimen flagellum broken off.
Pereopod I (Figure 5B) length in straightened state about 1.2 cephalothorax–pereon length; pereopods 2–7 lengths about 2–2.3 cephalothorax–pereon length.
Male paratypes:
Mandibles (Figure 7 and Figure 8): incisor process with five cusps. Lacinia mobilis of left mandible with four cusps; spine row with six spines and three SS. Spine row of right mandible with eight spines. Molar process thin, tubular, apex oval, and with sharp edges, producing two lobes bearing setae and scales. Palp length about 0.17 total body length; articles 1–3 lengths about 0.6, 0.4, 0.5 mandibular body length, respectively; article 1 with about six SS on anterior side and one UBS and one BS distally; article 2 with one UBS on anterior side; article 3 with about nine setae, first three of them much longer than other setae.
Maxilla I (Figure 7) lateral endite with seven robust serrate setae distally, with few fine SS laterally and medially; mesial endite width 0.66 lateral endite width, distally with five fine SS.
Maxilla II (Figure 7) lateral and middle endites similar in size, with four robust serrate setae distally and few fine SS laterally and medially; mesial endite width 1.8 lateral endite width, with 14 robust serrate setae distally, and fine SS medially.
Maxilliped (Figure 7) basis length 2 width, with one SS and one BS near palp insertion; endite length 1.4 width, 0.6 basis length, with 5–6 retinaculae, and distal margin with 15 DSS. Palp almost as long as basis, article 1 width 0.36 basis width, lateral length 0.38 width; articles 2–5 to article 1 lateral length ratios: 3.9; 2.7; 3.6; 3.4; articles 1–5 lateral lengths to width ratios: 0.38; 0.99; 0.88; 1.9; 3.6; articles 1–4 with two, eight, five, and six distomedial SS, respectively; article 6 with four terminal SS. Epipod length 2.1 width and 0.91 basis length, narrowing distally, distal tip truncated, with two SS.
Pereopod I (Figure 9) basis–dactylus length to width ratios: 3.5; 1.8; 1.5; 3.1; 4.2; 3.3; ischium–dactylus lengths to basis length ratios: 0.59; 0.52; 1.4; 0.82; 0.29. Basis, ischium, and merus bearing numerous stout-composed setae (UBS, UBDS). Carpus increasing in width towards midlength, with six stout UBS distodorsally and numerous fine DSS dorsally and subventrally; ventral margin convex, proximal part bearing 5–6 UBDS, increasing in size toward occlusal surface, with distal about as long as dactylus; occlusal surface occupying 0.6 of ventral margin, with about 15 strong tooth-like UBDS, which alternate with grooves (fit to same UBDS located on ventral margin of propodus); additionally, about five strong UBDS increasing in size toward occlusal surface, situated subventrally; ventral margin distal to occlusal surface with three small UBS. Propodus with numerous fine DSS on dorsal margin and along occlusal surface of ventral margin; occlusal surface similar to that of carpus, with about 15 tooth-like UBDS and grooves. Dactylus with few fine setae dorsally; dorsal claw length 0.49 dactylus length; ventral claw length 0.28 dorsal claw length; one seta inserted between claws.
Pereopods II–VII (Figure 9C and Figure 10) surface covered with numerous fine setulate; bases dorsal surface bearing few thin setulated spines; all articles except dactyli bearing numerous composed setae (setulated setae, UBS). Proximal articles (bases–meri) and dactyli similar in shape and proportions, while carpi and propodi increasing in length from anterior to posterior pereopods. Dactyli with two claws each; dorsal claw length about 0.4–0.5 dactylus length; ventral claw length about 0.3–0.4 dorsal claw length; two setae inserted between claws.
Pereopod II (Figure 9C) basis–dactylus length to width ratios: 3.5; 2.1; 2.3; 7.7; 12.3; 4.0; ischium–dactylus to basis length ratios: 0.55; 0.58; 1.5; 1.7; 0.41; two setulated setae inserted between claws.
Pereopod III (Figure 10) basis–dactylus length to width ratios: 3.5; 2.4; 2.2; 8.6; 19.8; 6.5; ischium–dactylus length to basis length ratios: 0.66; 0.69; 1.9; 2.8; 0.55.
Pereopod IV (Figure 10) basis–dactylus length to width ratios: 4.3; 2.5; 2.5; 8.6; 18.3; 5.1; ischium–dactylus length to basis length ratios: 0.55; 0.61; 1.6; 2.3; 0.41.
Pereopod V (Figure 10) basis–dactylus length to width ratios: 3.6; 2.4; 2.5; 9.3; 24.8; 6.2; ischium–dactylus length to basis length ratios: 0.63; 0.58; 1.8; 3.2. 0.56.
Pereopod VI (Figure 10) basis–carpus length to width ratios: 3.1; 2.9; 2.6; 11.0; ischium–carpus length to basis length ratios: 0.80; 0.78; 2.3. Distal articles broken off in dissected specimens.
Pereopod VII (Figure 10) basis–carpus length to width ratios: 3.1; 2.4; 2.7; 11.8; ischium–carpus length to basis length ratios: 0.68; 0.78; 2.53. Distal articles broken off in dissected specimens.
Pleopod I (Figure 11) convex ventrally, flattened tip bent up; length 3.5 maximal width; widest basally, narrowest 0.2 length from base, becoming gradually wider distally toward pronounced rounded distolateral angles, located 0.1 length from apex, where pleopod width is 0.2 length; distal tip pointed. Ventral surface covered with fine setulate, proximally to lateral angles bearing stout setulate setae; distal tip with fine SS.
Pleopod II (Figure 11) protopod length 3 width, narrowing distally, with about 10 stout UBS and numerous SS laterally. Endopod basal article length 1.9 width, and 0.19 protopod length; stylet 0.42 protopod length, distal third of stylet abruptly narrowing. Exopod length 1.2 width and 0.14 protopod length, with tuft of fine setae.
Pleopod III (Figure 11) endopod length 2.3 width, with three plumose setae, setae length half of endopod length. Exopod length 5.3 width and 2.1 endopod length, lateral margin with fine SS; basal article length 3.3 width and 1.3 exopod length; distal article length 3.4 width, tapering distally, with straight lateral and convex medial margin.
Pleopod IV (Figure 11) endopod length 1.3 width. Exopod length 3.7 width and 0.86 endopod length.
Pleopod V (Figure 11) endopod length 1.3 width.
Uropod of holotype (Figure 5) length 2.7 width, widest at 0.27 length from apex, distomedial angle oblique; distomedial margin with two stout UBS, distal margin with three BS and one trace of seta incretion.
Female, paratype:
Habitus generally similar with male (Figure 2C–E).
Pleopod II (operculum) (Figure 2E and Figure 12) leaf-shaped, length 1.3 width; with numerous stout-composed setae ventrally; lateral margins with fine SS.
Manca (Figure 2F) differing from adult individuals in less vaulted body, less inflated pleotelson, and fewer spines on body surface.
Remarks. Haplomunna kurilensis sp. nov. is similar to the second West Pacific species H. japonica (Gamô, 1983) from the Japan Trench. Both species significantly differ from the East Pacific species H. caeca (Richardson, 1905) and H. hubbsi Wilson, 1976 in having a large, much more inflated pleotelson.
Nevertheless, H. kurilensis sp. nov. is readily distinguishable from H. japonica (Gamô, 1983) in having the maxilliped with 5–6 retinaculae (in contrast to three retinaculae in H. japonica), and the epipod which is about as long as basis and has abruptly narrowing distal part (in contrast to the epipod reaching about 2/3 of the basis length in H. japonica). In H. kurilensis sp. nov., the first article of the mandibular palp bears a stout UBS distally, while the original illustrations of H. japonica provided by Gamô [10] show no composed setae in this location. In addition, in H. kurilensis sp. nov., the distal tip of the male pleopod I is pointed, with slightly pronounced distolateral angles, in contrast to the broadly rounded tip without angles in H. japonica.
Distribution. The species is known from the type locality in the NW Pacific Basin to the east of the KKT, from 43.0248° N 152.9727° E to 43.5913° N 153.9647° E at a depth of 5222–5380 m. It also possibly occurs to the west of the KKT from 46.10045° N 152.24065° E to 46.097117° N 152°242933 E at a depth of 4769–4798 m, but this information needs to be confirmed.
3.1.3. Genus Thylakogaster Wilson and Hessler, 1974
Type species. Thylakogaster peterpauli Wilson and Hessler, 1974.
Species included. Thylakogaster lobotourus Wilson and Hessler, 1974; T. majusculus Wilson and Hessler, 1974; T. namibiensis Brenke and Buschmann, 2009; T. peterpauli Wilson and Hessler, 1974; T. wilsoni Malyutina and Golovan sp. nov.
Diagnosis. See [2].
Thylakogaster wilsoni Malyutina and Golovan sp. nov.
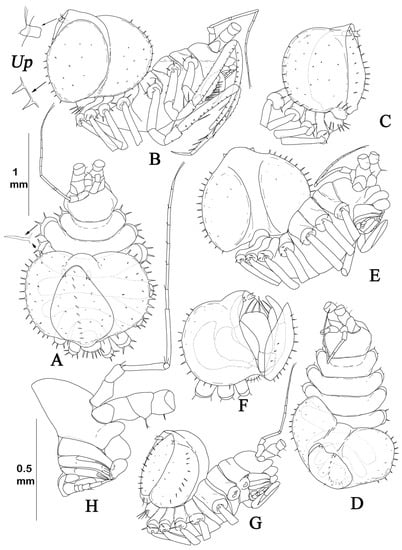
Figure 13.
Thylakogaster wilsoni Malyutina and Golovan sp. nov., female, holotype (SMF 61314): (A) dorsal view; (B) lateral view; (C) pleotelson, lateral view; (Up) uropod. Female paratypes (MIMB 46618). Female 1: (D) dorsal view; (E) lateral view; (F) pleotelson, oblique view, with open operculum; Female 2: (G) lateral view; (H) cephalothorax, lateral view; Scale bar = 0.1 mm.
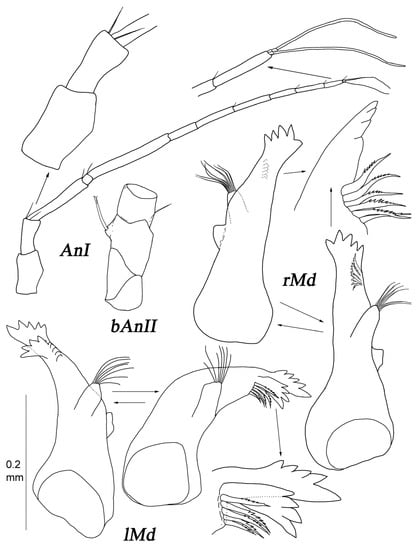
Figure 14.
Thylakogaster wilsoni Malyutina and Golovan sp. nov., female, paratype (MIMB 46618): (AnI) antennae I; (bAnII) basis (four first articles) of antenna II; (rMd and lMd) right and left mandibles in different positions with enlarged incisor process.
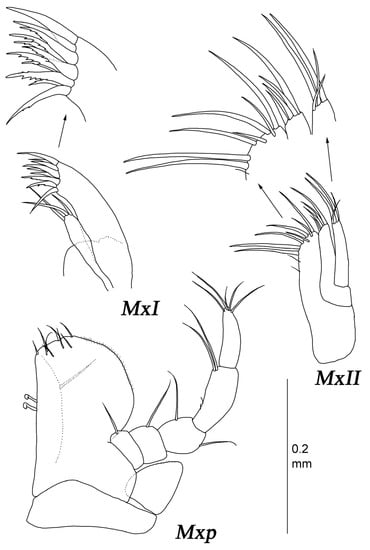
Figure 15.
Thylakogaster wilsoni Malyutina and Golovan sp. nov., female, paratype (MIMB 46618): (MxI and MxII) maxillae I and II with enlarged distal margins; (Mxp) maxilliped.
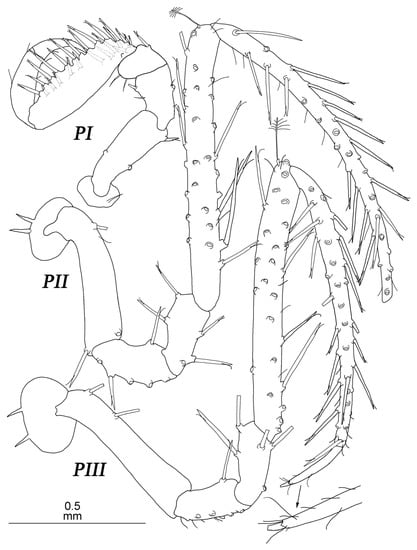
Figure 16.
Thylakogaster wilsoni Malyutina and Golovan sp. nov., female, holotype (SMF 61314): (PI–III) pereopods I–III.
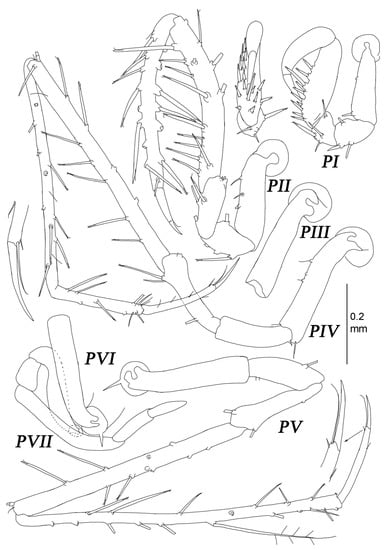
Figure 17.
Thylakogaster wilsoni Malyutina and Golovan sp. nov. female, paratype (MIMB 46618): (PI–VII) pereopods I–VII.
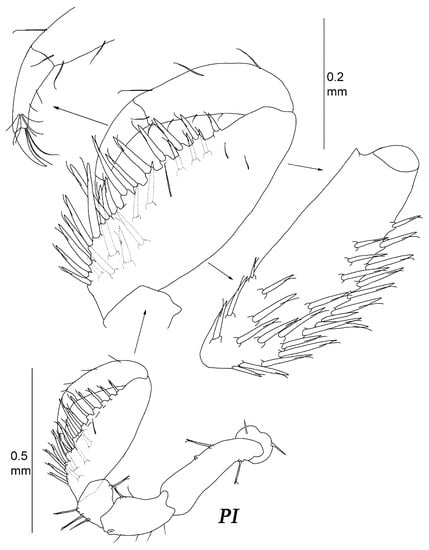
Figure 18.
Thylakogaster wilsoni Malyutina and Golovan sp. nov. female, paratype (MIMB 46618): (PI) pereopod I with enlarged carpus–dactylus, lateral view and carpus, inner view.
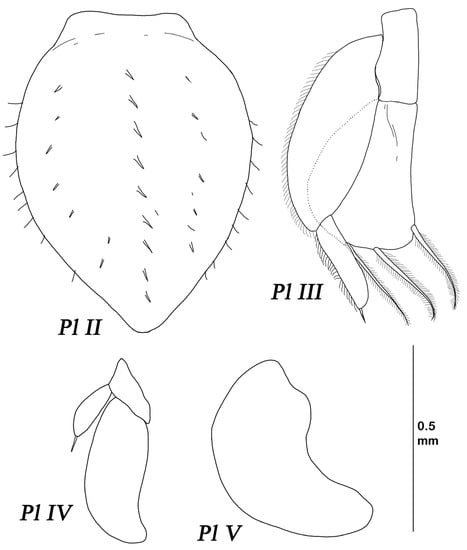
Figure 19.
Thylakogaster wilsoni Malyutina and Golovan sp. nov. female, paratype (MIMB 46618): (Pl II–V) pleopods II–V.
This species is registered in ZooBank under urn:lsid:zoobank.org:act:2B54E2D4-0764-4B15-893B-557CB90AC529.
Material examined. Holotype: (SMF 61314), female (total length 3.0 mm, cephalothorax–pereon length 1.9 mm), KB 223-8-9.
Paratypes: (MIMB 46618) female 1 (total length 2.8 mm), female 2 (total length 2.6 mm), the same data as holotype; (SMF 61315) manca damaged, the same data as holotype; (MIMB 46619) manca (total length 1.7 mm), KB 223-7-9; (SMF 61316) manca damaged, KB 223-7-9.
Etymology. The new species is named after the American isopodologist Dr. George D.F. (Bus) Wilson, for all his contributions to the study of Asellota and, in particular, the genus Thylakogaster.
Diagnosis. Female. Body broadest and deepest at pleotelson, body total length about twice width, and cephalothorax–pereon length 1.5 pereonite 3 width. Cephalothorax weakly recessed into first pereonite. Cephalothorax–pereon dorsal surface without spines. Pleotelson length 1.5 width, 0.6 cephalothorax–pereon length, with fine cuticular spinules and lateral row of spines on dorsolateral convexities, and tip distinctly set off from lateral convexities. Antenna I about 0.5 of body total length, and article 3 about twice longer than article 2. Maxilliped endite length 1.1 width; palp article 3 length 1.1 width; epipod length 0.4 basis length. Pereopod I carpus length 3 width, about 1.3 basis length, with around 25 stout UB setae on ventral margin and medial surface. Propodus–dactylus length 0.85 carpus length. Pereopod III only slightly shorter than pereopod II. Operculum length 1.35 width.
Description. Holotype, female: body (Figure 13A–C) broadest and deepest at pleotelson, total length 2.0 pleotelson width, and cephalothorax–pereon length 1.5 pereonite 3 width.
Cephalothorax: weakly recessed into pereonite 1, length 0.8 width and 0.2 body length, width 0.7 width of pereonite 1, and lateral sides with angular “checks”. Frons and clypeus rounded in dorsal view and protruded forward.
Pereon: lateral expansions of pereonites 1–3 rather tightly packed, directed anteriorly; directed laterally on pereonite 4 and directed posteriorly on pereonites 5–7. Coxae rounded, visible on dorsal view, bearing 2–3 spines. Pereonite 1 length 0.5 cephalothorax length, width 0.86 cephalothorax width; length ratios of pereonites 2–7 to pereonite 1: 1.0, 1.0, 0.87, 0.75, 0.63, 0.56.
Pleon: with one pleonite, pleonite length subequal to pereonite 7 medial length, with a row of long ventral spines. Pleotelson length 0.75 width and 0.36 body total length, dorsolateral convexities highly vaulted, with longitudinal dorsal groove in between, apex rounded, and weakly produced. Pleotelson with fine spinules on dorsal and ventral surface and lateral row of rather long cuticular spines along dorsolateral convexities.
Antenna I (Figure 14) length 0.53 body total length. Articles 1–5 length/width ratios: 1.7, 2.0, 5.7, 1.0, 8.5; length ratios of article 2–5 to article 1: 0.82, 1.41, 0.29, 2.0; following six flagellar articles subequal in length to article 2. Article 1 with ventral bump, article 2 with two distal SS, articles 3 and 4 each with one fine distal SS, some flagellar articles with fine distal seta, and the last article with two aestetascs twice longer than article.
Antenna II (Figure 14) broken off after peduncle article 4. Articles 1–4 of similar length, and article 3 with ventromedial bump bearing one UBS. Article 4 with one small UBS medially.
Mouth parts of small female, paratype:
Mandibles (Figure 14): Incisor process of both mandibles broadened distally, with five cusps. Spine row with five and six serrated setae on left and right mandibles, respectively. Lacinia mobilis of left mandible broadened distally, with four cusps, length 0.83 incisor process length. Molar process almost as long as incisor, cylindrical, with seven long distal setae and condyle length about half of molar length.
Maxilla I (Figure 15) lateral endite distally with 12 differently serrated setae; mesial endite distally with three strong SS.
Maxilla II (Figure 15) lateral and middle endites together almost as broad as mesial endite; all endites with strong distal SS. Lateral endite longest, with two long and two short SS, and middle endite with one long and three short SS. Mesial endite shortest, with five long and two short SS distally and two fine SS medially.
Maxilliped (Figure 15) basis length 1.54 width, endite as long as wide, 0.65 basis length, with two retinacula, and distal margin with six SS. Palp narrow, gradually tapering, length 1.37 basis length; article 1 width 0.42 basis width; article 5 0.4 of article 1 width; articles 1–5 length to width ratios: 0.41, 0.66, 1.12, 2.33, 3.57; articles 2–5 lengths to article 1 length ratios: 1.42, 2.42, 4.0, 3.57. Articles 1–3 with one long medial SS, and article 3 additionally with one distolateral SS; article 4 with two distomedial SS, and article 5 with 5 distal SS. Epipod triangular, width 1.1 length, 0.47 basis width, and length 0.38 basis length.
Pereopod I (Figure 16) robust, subchelate, length 0.46 body length, and length ratios of ischium–dactylus to basis: 0.62, 0.32, 1.28, 0.72, 0.35. Basis to dactylus length/width ratios: 2.68, 1.69, 0.78, 3.05, 3.1, 2.1. Basis with three stout short UBS. Ischium with one ventral, one dorsal stout short UBS, and few SS. Merus with two SS on ventral margin and one UBS on dorsal margin. Carpus with 13–14 stout UBS on outer ventral margin; medial surface with longitudinal row of five UBS near outer ventral margin and six UBS proximally. Propodus with three ventral and two dorsal SS. Dactylus with two fine SS, claw length 0.7 dactylus length, two SS ventrally near claw, and tuft of six SS proximally to claw. Tip of claw reaching proximal quarter of carpus when folded against it.
Pereopod II (Figure 16) with broken dactylus. Length ratios of ischium–propodus to basis: 0.6, 0.55, 2.0, 2.52, 0.35. Basis length 1.44 basis of pereopod I length. Basis to dactylus length/width ratios: 3.87, 1.17, 2.12, 9.54, 17.5. Ischium with one dorsal and three ventral UBS. Merus with two ventral and two dorsal UBS. Carpus with around 25 long UBS. Propodus with >30 UBS.
Pereopod III (Figure 16) length 2.2 pereopod I length, length without dactylus 0.95 P II length without dactylus. Length ratios of ischium–dactylus to basis: 0.52, 0.53, 1.82, 1.86, 0.42. Basis to dactylus length/width ratios: 3.9, 2.26, 2.33, 10.0, 13.7, 8.0. Ischium with five ventral UBS and few small SS. Merus with one ventral and two dorsal UBS. Carpus with about twenty UBS and two distodorsal SS. Propodus with 25 UBS, with about 10 SS distally. Dactylus with few fine SS; tip of claw broken off.
Pereopods IV–VII broken off; bases of subequal length and slightly narrower than basis of pereopod III.
Pereopod I of large female, paratype (Figure 17) length 0.51 body length, length ratios of ischium–dactylus to basis: 0.67, 0.34, 1.28, 0.83, 0.39. Basis to dactylus length/width ratios: 2.68, 1.69, 0.78, 2.75, 3.1, 2.1. Basis length 3 width, with three UBS. Ischium with few SS and one ventral and one dorsal stout short UBS. Merus with two SS on ventral margin and one UBS on dorsal margin. Carpus with fourteen strong UBS on outer ventral margin; medial surface with longitudinal row of five UBS on outer ventral margin and six UBS proximally. Propodus with six ventral and three dorsal SS. Dactylus length approximately 0.5 propodus length. Dactylus with two fine SS ventrally, tuft of six SS proximally to claw, with claw about half of dactylus length and two long ventral SS.
Pereopods of manca, female, paratype (Figure 18). Pereopod I length 0.5 body length, length ratios of ischium–dactylus to basis: 0.55, 0.40, 1.25, 0.82, 0.42. Basis to dactylus length/width ratios: 3.1, 1.57, 1.06, 3.33, 3.0. Basis with two dorsal UBS. Ischium with two ventral and one dorsal stout UBS. Merus with three ventral and one dorsal UBS. Carpus with eight strong UBS on outer ventral margin; medial surface with longitudinal row of five UBS along outer ventral margin and six UBS proximally. Propodus with four ventral and three dorsal SS. Dactylus claw about 1/3 of dactylus length.
Pereopod II length 1.85 pereopod I length. Length ratios of ischium–dactylus to basis: 0.57, 0.72, 1.91, 2.0, 0.61. Basis to dactylus length/width ratios: 2.9, 1.68, 2.26, 7.5, 11.75, 7.25. Basis with three dorsal UBS. Ischium with five ventral UBS and few small SS. Merus with one ventral and two dorsal UBS. Carpus with about twenty UBS and two distodorsal SS. Propodus with 25 UBS, with about 10 SS distally. Dactylus with few fine SS.
Pereopod III broken off after basis. Basis length 1.27 basis II length and 4.0 width.
Pereopod IV length 2.95 pereopod I length and 1.63 pereopod II length. Length ratios of ischium–dactylus to basis: 0.62, 0.76, 1.97, 2.70, 0.90. Basis to dactylus length/width ratios: 4.7, 3.15, 3.12, 14.4, 29.66, 15.0. Basis with three dorsal UBS and two distal SS. Ischium without setae. Merus with one ventral, two distoventral UBS, and one distodorsal UBS. Carpus with about ten UBS and one strong distodorsal BS (broken in specimen). Propodus with about 20 UBS and few SS. Dactylus with few fine SS and one long SS distoventrally; claw slender, with length 0.5 dactylus length.
Pereopod V (Figure 17) length subequal to pereopod IV length (0.99). Length ratios of ischium–dactylus to basis: 0.61, 0.76, 1.88, 2.52, 0.88. Basis to dactylus length/width ratios: 4.85, 3.0, 3.47, 14.22, 28.66, 15.0. Basis without setae. Ischium with one ventral UBS. Merus with one ventral, one distoventral, and one distodorsal UBS. Carpus with about ten UBS and one strong distodorsal BS (broken in specimen). Propodus with eighteen UBS and one long plumose seta dorsally. Dactylus with few fine SS and one long seta distoventrally; claw slender, with length 0.5 dactylus length.
Pereopod VI broken off after basis. Basis length equal to bases IV and V length 5.5 width.
Pereopod VII (Figure 17) not developed, with six separated articles.
Pleopods of female, paratype (Figure 19):
Pleopod II length 1.35 width, ventrally with medial row of spines like on lateral margins of pleotelson, with marginal small setae.
Pleopod III protopod length 2.36 width; endopod length 1.46 width, 1.58 protopod length, width 2.55 protopod width, and distal margin with three plumose setae, which is about 0.65 of endopod length. Exopod is twice longer than endopod, with fine setulae laterally, basal article length 2.71 width, 1.39 endopod length, distal article length 4.33 width, and 0.45 basal article length, with one short terminal SS.
Pleopod IV endopod length 2.87 width. Exopod length 3.2 width and 0.34 endopod length, with one short terminal SS.
Pleopod V endopod length 1.96 width.
Uropod of holotype (Figure 13) length 2 width, distolateral lobe rounded, and twice longer and broader than tiny distomedial lobe, with one short terminal SS and distomedial lobe acute, with three terminal SS.
Remarks. The new species can be distinguished from four known species of the genus: T. lobotourus Wilson and Hessler, 1974, T. majusculus Wilson and Hessler, 1974, T. peterpauli Wilson and Hessler, 1974, and T. namibiensis Brenke and Buschmann, 2009 by the less spiny body, shape, and proportions of segments and appendage articles. In general, T. wilsoni sp. nov. has the broadest pleotelson of all known species from the Atlantic. In contrast to other species, pleotelson tip of the new species distinctly set off from the lateral convexities; by this character, T. wilsoni sp. nov. is close to T. namibiensis. Antenna I article 3 in T. wilsoni sp. nov. is about 2.5 times longer than article 2, while in females of T. namibiensis and T. lobotourus, these articles are subequal in length; in T. peterpauli and T. majusculus, this ratio is 1.6 and 1.8, respectively. In contrast to T. lobotourus, pereopod II of adult specimen of T. wilsoni sp. nov. is not distinctly longer than pereopod III, though T. wilsoni sp. nov. has carpus and propodus of pereopod II longer than these two articles of pereopod III (1.03 and 1.14, respectively). Unfortunately, we could not compare pereopod II with pereopod III in manca as it was broken. Pereopod I of the new species has relatively longer propodus–dactylus concerning carpus: 0.9–0.8, while in T. lobotourus, this ratio is 0.7-0.75; carpus is more elongated in T. wilsoni sp. nov.: length/width ratio is 2.75–3.05, while in T. lobotourus it is 2.1–2.5.
Maxilliped is an important structure for morphological comparison of species. In T. wilsoni sp. nov., the endite length/width ratio is 1.1, and the palp article 3 length/width ratio is also 1.1, while in T. peterpauli and T. majusculus, the ratio of endite is 0.8 and 0.9, respectively, and the ratio of palp article 3 1.9 and 1.7, respectively. By these two ratios, the new species is close to T. namibiensis, though differs from it by the short epipod (length/basis length is 0.4 in T. wilsoni sp. nov. and 0.7 in T. namibiensis).
Distribution. The species is known only from the type locality in the NW Pacific Basin to the east of the Kuril–Kamchatka Trench, 42.2447° N 151.7351° E 42.2378° N 151.7082° E at a depth of 5125–5126 m and 43.0473°N 152.9905° E to 43.0248° N 152.9727° E at a depth of 5222–5223 m.
3.2. Molecular Analysis
Gene extraction was unsuccessful for the specimens of Abyssaranea minuta sp. nov. and Haplomunna cf. kurilensis sp. nov. (Table 3).
According to the obtained phylogenetic results, sequences of the families Haplomunnidae, Dendrotionidae, and Janiridae are clustered in their own unique clades. The family Munnidae also formed a unique clade, but it is distant from other families. Supporting values were high for all clades (posterior probability (pp) = 1), except the Dendrotionidae; for the Dendrotionidae, pp = 0.76. The families Haplomunnidae and Dendrotionidae were more closely related than Janiridae (Figure 20). Genetic distances between families are showed in the Table 5.
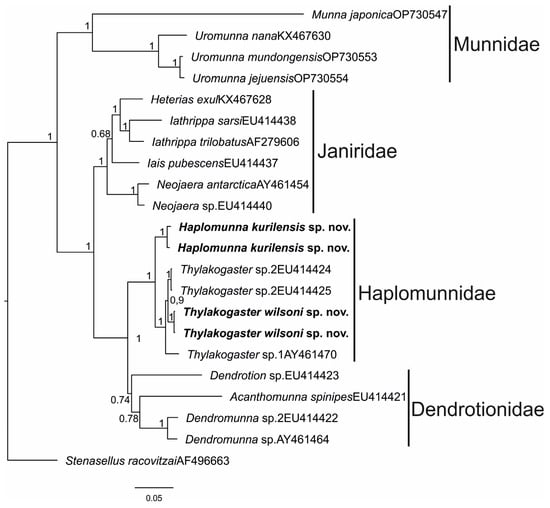
Figure 20.
Bayesian inference analysis of 18S rDNA sequences showing phylogenetic relationships of Haplomunnidae and related groups. Numerals above or below the branches are Bayesian posterior probabilities.
Table 5.
18S rDNA uncorrected genetic distances between the families Haplomunnidae, Dendrotionidae, Janiridae, and Munnidae. Above the diagonal is the standard error (SE).
Sequences of Thylakogaster wilsoni sp. nov. were included in the clade with other Thylakogaster species (pp = 1) and formed well supported clade with Thylakogaster sp. 2 (pp = 0.99). The separations of T. wilsoni and Thylakogaster sp. 2 from Thylakogaster sp. 1 was also well supported (pp = 1). Species Haplomunna kurilensis sp. nov. grouped beside the Thylakogaster clades, together forming a monophyletic clade of Haplomunnidae (Figure 20).
The intraspecific genetic divergence within T. wilsoni sp. nov. and Thylakogaster sp. 1 was 0; within H. kurilensis sp. nov.—0.3 ± 0.1% (mean ± standard error).
The genetic divergences between T.wilsoni sp. nov. and Thylakogaster sp. 2 was 0.5 ± 0.2%; between T. wilsoni sp. nov. and Thylakogaster sp. 1 was 1.1 ± 0.2%. The interspecific genetic divergence between H. kurilensis sp. nov. and T. wilsoni sp. nov. was 1.8 ± 0.3%. Genetic distances for Haplomunnidae species are shown in Table 6.
Table 6.
18S rDNA uncorrected genetic distances between species of the family Haplomunnidae. Above the diagonal is the standard error (SE).
4. Discussion
The discovery of the three species from three genera of Haplomunnidae in the abyssal of the NW Pacific is of great interest.
All hitherto known species of the genera Thylakogaster and Abyssaranea inhabit the bathyal and abyssal of the Atlantic Ocean, where they were sampled at depth ranges of 1135–5415 and 3459–3757 m, respectively [2,12,13]. In addition, Brenke and Buschmann [13] reported several finds of A. rupis Wilson and Hessler, 1974 in the South Atlantic at a depth range of ca. 5000–5500 m. In the Pacific Ocean, these genera have already been encountered before, but this information is limited to reports of several findings of their representatives in open nomenclature in the abyssal of the equatorial and northern subequatorial regions of the Eastern Pacific [2] (see also https://obis.org). Among them, there was a report on the finding of a single specimen of A. cf. rupis at a depth of 4435–4438 m (see [2]), whose belonging to the species A. rupis, described from the equatorial Atlantic, is doubtful and needs to be confirmed. T. wilsoni Malyutina and Golovan sp. nov., and A. minuta sp. nov. Golovan and Malyutina, are the first species of their genera described from the Pacific Ocean.
The genus Haplomunna has, according to currently available data, a North Pacific distribution. H. caeca (Richardson, 1905) and H. hubbsi Wilson, 1976 are known from the abyssal of the NE Pacific (3880–5183 m) [1,27]. In the NW Pacific, one species, H. japonica (Gamô, 1983), was known from the upper part of the eastern (oceanic) slope of the Japan Trench (6380–6450 m) [10,26]. H. kurilensis Golovan and Malyutina sp. nov. was found in the abyssal to the east of the neighboring Kuril–Kamchatka Trench (KKT) at a depth of 5222–5380 m. In addition, one manca, which we assigned to H. kurilensis sp. nov., was found to the west of the KKT at a depth of 4769–4798 m. Both species (H. japonica and H. kurilensis sp. nov.) are very close morphologically while differing markedly from the East Pacific species. Interestingly, the East Pacific species have also been found off the coast of California at close distances (about 100 miles) from each other [1].
The existence of two close species which live nearby, but are separated by the depths of their habitat, is quite a common occurrence. Depths ca. 6000–7000 m are known as a transition zone between abyssal and trench faunas where abyssal species are disappearing, and hadal species are appearing [43,44]. During recent deep-sea studies in various regions of the World Ocean, much evidence has been obtained that for many studied deep-sea species, vertical distributions do not exceed several hundreds of meters [45,46,47,48]. Some species that initially seemed to have a wide bathymetric range turned out to be composed of cryptic (morphologically similar but genetically distinct) species [44,46]. At the same time, according to these studies, the horizontal distribution of abyssal species can extend over several hundred kilometers [44,45,46]. The role of the KKT with its hadal depths as a geomorphological barrier for the bathymetric and geographic distributions of the abyssal species of Isopoda has been discussed previously for other janiroidean families [44,49,50,51,52]. KKT was found to be a barrier in the dispersal of abyssal species of Ischnomesidae [50]. Both the Japan Trench and the KKT, whose boundaries coincide with the 6000 m isobaths, restrict gene flow, creating an isolating effect on populations living on both sides of the trenches. Nevertheless, some species of the families Munnopsidae, Desmosomatidae, Macrostylidae, and Haploniscidae overcome the KKT barrier, occurring in the abyssal on both sides [44,53,54,55,56]. However, given that H. kurilensis sp. nov. was represented by a single immature specimen in our material collected in the abyssal to the west of the KKT, some doubt remains there about the accuracy of the species identification of this individual. Unfortunately, an attempt to obtain genetic data from this specimen failed. Additional collections under scopes of the future studies should help in establishing the range of H. kurilensis sp. nov.
An interesting observation is the fact that H. kurilensis sp. nov. in our collections was almost completely represented by males and mancas, and only one female of this species was sampled at the richest station. In the type material of H. japonica collected during the expedition onboard the Japanese RV Hakuho-Maru at one station in the Japan Trench, the sex ratio was similar [10,26]. On the contrary, the species Thylakogaster wilsoni sp. nov. and Abyssaranea minuta sp. nov. were represented only by females and mancas. The Eastern Pacific species of the genus Haplomunna are known only by females as well [1,27]. In general, the numerical predominance of females over males is typical for deep-sea isopod species ([49,57] and personal observations). Due to the small amount of material, at this stage, it is impossible to give a clear explanation for the predominance of males in the collections of the two Western Pacific species of Haplomunna. However, we can assume that males and females of haplomunnids have differences in behavior, lifestyle, or ecological preferences, contributing to the fact that mainly males of these species were sampled. For example, Riehl and coauthors [49,58] suggested that males and females of another janiroidean family, Macrostylidae, differ in their lifestyle. Females have a more passive infaunal lifestyle. They are larger in size, and their task is to save energy for the production of offspring, whereas males have more actively searching epifaunal lifestyle. Some haplomunnid species (e.g., Abyssaranea) also show sexual dimorphism with the largest females [9].
The phylogenetic reconstruction based on 18S sequences corresponded with previous data based on morphological [1] and genetic analysis [3,21,59]. Families included in the analysis showed their monophyly; however, support for the family Dendrotionidae was not high. This family showed unresolved branches in previous research as well [3,21,59]. Obviously, a single genetic marker was not sufficient to resolve phylogeny between the families.
Two species of Thylacogaster (T. wilsoni and Thylacogaster sp. 2) from the family Haplomunnidae showed low level of interspecific distances (lower 1%) for the 18S marker. Such value of genetic distances between species was described earlier for species Uromunna (Munnidae) [59], and it may be explained by high conservatism of this marker. Our genetic results are the first genetic data for the described Thylakogaster species and for the genus Haplomunna. Further studies exploring additional samplings and a larger set of genetic markers should be used.
Author Contributions
Conceptualization, investigation, resources, data curation, visualization, O.A.G., M.V.M. and S.N.S.; writing of the original text, O.A.G., M.V.M. and S.N.S.; writing and revision, O.A.G., M.V.M. and S.N.S. All authors have read and agreed to the published version of the manuscript.
Funding
The expedition KuramBio was undertaken with financial support of the PTJ (German Ministry for Science and Education), Grant 03G0223A to Prof. Angelika Brandt. The expedition SokhoBio was undertaken with financial support of the Russian Science Foundation, Project No 14-50-00034.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The type material is deposited in the Museum of the NSCMB FEB RAS.
Acknowledgments
We would like to thank the crews of the RV Sonne and Akademik M.A. Lavrentyev for their help on board. We also thank the members of the international scientific team of both the expeditions for their help onboard, support, and fruitful collaboration. We are sincerely grateful to the anonymous reviewers for their valuable comments improving the quality of our manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wilson, G.D.F. The systematics and evolution of Haplomunna and its relatives (Isopoda, Haplomunnidae, new family). J. Nat. Hist. 1976, 10, 569–580. [Google Scholar] [CrossRef]
- Cunha, M.R.; Wilson, G.D.F. Haplomunnidae (Crustacea: Isopoda) reviewed, with a ddescription of an intact specimen of Thylakogaster Wilson & Hessler, 1974. Zootaxa 2003, 326, 1–16. [Google Scholar] [CrossRef]
- Raupach, M.J.; Mayer, C.; Malyutina, M.; Wägele, J.W. Multiple origins of deep-sea Asellota (Crustacea: Isopoda) from shallow waters revealed by molecular data. Proc. R. Soc. B Biol. Sci. 2009, 276, 799–808. [Google Scholar] [CrossRef]
- Brökeland, W.; Brandt, A. Two new species of Ischnomesidae (Crustacea: Isopoda) from the Southern Ocean displaying neoteny. Deep Res. Part II Top. Stud. Oceanogr. 2004, 51, 1769–1785. [Google Scholar] [CrossRef]
- Golovan, O.A.; Malyutina, M.V. A new deep-sea genus of Paramunnidae (Isopoda: Asellota) from the Northwest Pacific adjacent to the Kuril-Kamchatka Trench with remarks on paedomorphosis in deep-sea species. Prog. Oceanogr. 2019, 178, 102200. [Google Scholar] [CrossRef]
- Wilson, G. Compensation improvements in RC-active integrators. IEEE Proc. Part G Electron. Circuits Syst. 1989, 136, 1–8. [Google Scholar] [CrossRef]
- Thistle, D.; Wilson, G.D.F. A Hydrodynamically modified, abyssal isopod fauna. Deep Res. Part I Oceanogr. Res. Pap. 1987, 34, 73–87. [Google Scholar] [CrossRef]
- Beaulieu, S.E. Colonization of habitat islands in the deep sea: Recruitment to glass sponge stalks. Deep Res. Part I Oceanogr. Res. Pap. 2001, 48, 1121–1137. [Google Scholar] [CrossRef]
- Wilson, G.D.F.; Hessler, R.R. Some unusual Paraselloidea (Isopoda, Asellota) from the deep benthos of the Atlantic. Crustaceana 1974, 27, 47–67. [Google Scholar] [CrossRef]
- Gamô, S. Some species of abyssal Asellote isopods (Crustacea) from ast of the Japan Trench, with the descriptions of Janirella (Janirella) aculeata sp. nov., J. (Parajanirella) sedecimtuberculata sp. nov., and Aryballurops japonica gen. et sp. nov. Sci. Rep. Yokohama Natl. Univ. II 1983, 30, 1–18. [Google Scholar]
- Just, J. Haplodendron buzwilsoni gen. nov., sp. nov., the first record of Haplomunnidae from the southern Indo-Pacific (Isopoda: Asellota). Zootaxa 2003, 372, 1–10. [Google Scholar] [CrossRef]
- Boyko, C.B.; Bruce, N.L.; Hadfield, K.A.; Merrin, K.L.; Ota, Y.; Poore, G.C.B.; Taiti, S.; Schotte, M.; Wilson, G.D.F. World Marine, Freshwater and Terrestrial Isopod Crustaceans Database. Available online: http://www.marinespecies.org/aphia.php?p=taxdetails&id=118327 (accessed on 2 June 2023).
- Brenke, N.; Buschmann, A. Thylakogaster namibiensis sp. nov. (Isopoda: Asellota: Janiroidea), a new species of Haplomunnidae from the southeast Atlantic deep sea. Zootaxa 2009, 394, 381–394. [Google Scholar] [CrossRef]
- Elsner, N.O.; Malyutina, M.V.; Golovan, O.A.; Brenke, N.; Riehl, T.; Brandt, A. Deep down: Isopod biodiversity of the Kuril-Kamchatka abyssal area including a comparison with data of previous expeditions of the RV Vityaz. Deep Res. Part II Top. Stud. Oceanogr. 2015, 111, 210–219. [Google Scholar] [CrossRef]
- Golovan, O.A.; Błażewicz, M.; Brandt, A.; Jażdżewska, A.; Jóźwiak, P.; Lavrenteva, A.V.; Malyutina, M.V.; Petryashov, V.V.; Riehl, T.; Sattarova, V.V. Diversity and distribution of peracarid crustaceans (Malacostraca) from the abyss adjacent to the Kuril-Kamchatka Trench. Mar. Biodivers. 2019, 49, 1343–1360. [Google Scholar] [CrossRef]
- Brandt, A.; Elsner, N.; Brenke, N.; Golovan, O.; Malyutina, M.V.; Riehl, T.; Schwabe, E.; Würzberg, L. Epifauna of the Sea of Japan collected via a new epibenthic sledge equipped with camera and environmental sensor systems. Deep Res. Part II Top. Stud. Oceanogr. 2013, 86–87, 43–55. [Google Scholar] [CrossRef]
- Brandt, A.; Elsner, N.O.; Malyutina, M.V.; Brenke, N.; Golovan, O.A.; Lavrenteva, A.V.; Riehl, T. Abyssal macrofauna of the Kuril-Kamchatka Trench Area (Northwest Pacific) collected by means of a camera-epibenthic sledge. Deep Res. Part II Top. Stud. Oceanogr. 2015, 111, 175–187. [Google Scholar] [CrossRef]
- Golovan, O.A.; BŁazewicz-Paszkowycz, M.; Brandt, A.; Budnikova, L.L.; Elsner, N.O.; Ivin, V.V.; Lavrenteva, A.V.; Malyutina, M.V.; Petryashov, V.V.; Tzareva, L.A. Diversity and distribution of peracarid crustaceans (Malacostraca) from the continental slope and the deep-sea basin of the Sea of Japan. Deep Res. Part II Top. Stud. Oceanogr. 2013, 86–87, 66–78. [Google Scholar] [CrossRef]
- Brandt, A.; Alalykina, I.; Fukumori, H.; Golovan, O.; Kniesz, K.; Lavrenteva, A.; Lörz, A.N.; Malyutina, M.; Philipps-Bussau, K.; Stransky, B. First insights into macrofaunal composition from the SokhoBio expedition (Sea of Okhotsk, Bussol Strait and northern slope of the Kuril-Kamchatka Trench). Deep Res. Part II Top. Stud. Oceanogr. 2018, 154, 106–120. [Google Scholar] [CrossRef]
- Janssen, A.; Kaiser, S.; Meißner, K.; Brenke, N.; Menot, L.; Arbizu, P.M. A reverse taxonomic approach to assess macrofaunal distribution patterns in abyssal Pacific polymetallic nodule fields. PLoS ONE 2015, 10, e0117790. [Google Scholar] [CrossRef]
- Raupach, M.J.; Held, C.; Wägele, J.W. Multiple colonization of the deep sea by the Asellota (Crustacea: Peracarida: Isopoda). Deep Res. Part II Top. Stud. Oceanogr. 2004, 51, 1787–1795. [Google Scholar] [CrossRef]
- Golovan, O.A.; Malyutina, M.V. The first record of the family Paramunnidae (Isopoda: Asellota) from the bathyal of the Bering Sea with descriptions of two new species of Munnogonium. Deep Res. Part II Top. Stud. Oceanogr. 2022, 200, 105095. [Google Scholar] [CrossRef]
- Malyutina, M.V.; Golovan, O.A. The first record of Asellota (Isopoda) from hydrothermal vent biotopes of the submarine Piip Volcano, Bering Sea, with descriptions of two new species of Munnopsidae. Deep Res. Part II Top. Stud. Oceanogr. 2022, 202, 105137. [Google Scholar] [CrossRef]
- Riehl, T.; Wölfl, A.C.; Augustin, N.; Devey, C.W.; Brandt, A. Discovery of widely available abyssal rock patches reveals overlooked habitat type and prompts rethinking deep-sea biodiversity. Proc. Natl. Acad. Sci. USA 2020, 117, 15450–15459. [Google Scholar] [CrossRef] [PubMed]
- Kireev, P.A.; Golovan, O.A.; Sharina, S.N. First record of the family Caprellidae (Amphipoda: Senticaudata) from the abyssal zone of the Bering Sea with description of a new species of Cercops. Deep Res. Part II Top. Stud. Oceanogr. 2023, 208, 105238. [Google Scholar] [CrossRef]
- Gamô, S. Systematic study on the benthic small Crustaceans in the Japan Trench and its vicinity. In Preliminary Report of the Hakuho Maru Cruise KH-81-4. 6 July–4 August 1981. Japan Trench (WESTPAC); Ocean Research Institute, University of Tokyo: Tokyo, Japan, 1988; pp. 44–53. [Google Scholar]
- Richardson, H. A Monograph on the isopods of North America. Bull. U. S. Natl. Museum 1905, 54, 1–517. [Google Scholar] [CrossRef]
- Brandt, A.; Malyutina, M.V. The German-Russian deep-sea expedition KuramBio (Kurile Kamchatka biodiversity studies) on board of the RV Sonne in 2012 following the footsteps of the legendary expeditions with RV Vityaz. Deep Res. Part II Top. Stud. Oceanogr. 2015, 111, 1–9. [Google Scholar] [CrossRef]
- Malyutina, M.V.; Chernyshev, A.V.; Brandt, A. Introduction to the SokhoBio (Sea of Okhotsk biodiversity studies) expedition 2015. Deep Res. Part II Top. Stud. Oceanogr. 2018, 154, 1–9. [Google Scholar] [CrossRef]
- Golovan, O.A.; Malyutina, M.V.; Brandt, A. First record of the deep-sea isopod family Dendrotionidae (Isopoda: Asellota) from the Northwest Pacific with description of two new species of Dendromunna. Mar. Biodivers. 2018, 48, 531–544. [Google Scholar] [CrossRef]
- Hessler, R.R. The Desmosomatidae (Isopoda, Asellota) of the Gay Head-Bermuda Transect; University of California Press: Berkeley, CA, USA, 1970; Volume 15, ISBN 0520093194. [Google Scholar]
- Walsh, P.S.; Metzger, D.A.; Higuchi, R. Chelex 100 as a medium for simple extraction of DNA for PCR-based typing from forensic material. Biotechniques 1991, 10, 506–513. [Google Scholar] [CrossRef]
- Giribet, G.; Carranza, S.; Baguna, J.; Riutort, M.; Ribera, C. First molecular evidence for the existence of a Tardigrada + Arthropoda clade. Mol. Biol. Evol. 1996, 13, 76–84. [Google Scholar] [CrossRef]
- Whiting, M.F.; Carpenter, J.C.; Wheeler, Q.D.; Wheeler, W.C. The Strepsiptera problem: Phylogeny of the holometabolous insect orders inferred from 18S and 28S ribosomal DNA sequences and morphology. Syst. Biol. 1997, 46, 1–68. [Google Scholar] [CrossRef]
- Distel, D.L. Phylogenetic relationships among Mytilidae (Bivalvia): 18S rRNA data suggest convergence in mytilid body plans. Mol. Phylogenet. Evol. 2000, 15, 25–33. [Google Scholar] [CrossRef]
- Olson, P.D.; Cribb, T.H.; Tkach, V.V.; Bray, R.A.; Littlewood, D.T.J. Phylogeny and classification of the Digenea (Platyhelminthes: Trematoda). Int. J. Parasitol. 2003, 33, 733–755. [Google Scholar] [CrossRef]
- Williams, S.T.; Reid, D.G.; Littlewood, D.T.J. A molecular phylogeny of the Littorininae (Gastropoda: Littorinidae): Unequal evolutionary rates, morphological parallelism, and biogeography of the Southern Ocean. Mol. Phylogenet. Evol. 2003, 28, 60–86. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES science gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; IEEE: Piscataway, NJ, USA, 2010; pp. 1–8. [Google Scholar]
- Rambaut, A. FigTree 1.4.4 – Produce Figures of Phylogenetic Trees. 2012, pp. 1–8. Available online: https://mybiosoftware.com/figtree-1-3-1-produce-figures-phylogenetic-trees.html (accessed on 2 June 2023).
- Belyaev, G.M. The Deep-Sea Ocean Trenches and Their Fauna; Nauka: Moskow, Russia, 1989. [Google Scholar]
- Jennings, R.M.; Golovan, O.; Brix, S. Integrative species delimitation of desmosomatid and nannoniscid isopods from the Kuril-Kamchatka Trench, with description of a hadal species. Prog. Oceanogr. 2020, 182, 102236. [Google Scholar] [CrossRef]
- Brix, S.; Svavarsson, J. Distribution and diversity of desmosomatid and nannoniscid isopods (Crustacea) on the Greenland-Iceland-Faeroe Ridge. Polar Biol. 2010, 33, 515–530. [Google Scholar] [CrossRef]
- Jennings, R.M.; Etter, R.J.; Ficarra, L. Population differentiation and species formation in the deep sea: The potential role of environmental gradients and depth. PLoS ONE 2013, 8, e77594. [Google Scholar] [CrossRef]
- Havermans, C.; Sonet, G.; d’Udekem d’Acoz, C.; Nagy, Z.T.; Martin, P.; Brix, S.; Riehl, T.; Agrawal, S.; Held, C. Genetic and morphological divergences in the cosmopolitan deep-sea amphipod Eurythenes gryllus reveal a diverse abyss and a bipolar species. PLoS ONE 2013, 8, e74218. [Google Scholar] [CrossRef] [PubMed]
- Brix, S.; Svavarsson, J.; Leese, F. A Multi-gene analysis reveals multiple highly divergent lineages of the isopod Chelator insignis (Hansen, 1916) South of Iceland. Pol. Polar Res. 2014, 35, 225–242. [Google Scholar] [CrossRef]
- Bober, S.; Riehl, T.; Henne, S.; Brandt, A. New Macrostylidae (Isopoda) from the Northwest Pacific Basin described by means of integrative taxonomy with reference to geographical barriers in the abyss. Zool. J. Linn. Soc. 2018, 182, 549–603. [Google Scholar] [CrossRef]
- Bober, J.; Brandt, A.; Frutos, I.; Schwentner, M. Diversity and distribution of Ischnomesidae (Crustacea: Isopoda: Asellota) along the Kuril-Kamchatka Trench—A genetic perspective. Prog. Oceanogr. 2019, 178, 102174. [Google Scholar] [CrossRef]
- Johannsen, N.; Lins, L.; Riehl, T.; Brandt, A. Changes in species composition of Haploniscidae (Crustacea: Isopoda) across potential barriers to dispersal in the Northwest Pacific. Prog. Oceanogr. 2020, 180, 102233. [Google Scholar] [CrossRef]
- Knauber, H.; Silberberg, J.R.; Brandt, A.; Riehl, T. Evolution and biogeography of the Haploniscus belyaevi species complex (Isopoda: Haploniscidae) revealed by means of integrative taxonomy. Syst. Biodivers. 2022, 20, 1–27. [Google Scholar] [CrossRef]
- Golovan, O.A. Desmosomatidae (Isopoda: Asellota) from the abyssal plain to the east of the Kuril-Kamchatka Trench: New data on diversity with the description of two new species. Deep Res. Part II Top. Stud. Oceanogr. 2015, 111, 256–278. [Google Scholar] [CrossRef]
- Golovan, O.A. Description of two ubiquitous species of Desmosomatidae (Isopoda: Asellota) from the Northwest Pacific Basin east of the Kuril-Kamchatka Trench. Zootaxa 2015, 4039, 201–224. [Google Scholar] [CrossRef]
- Riehl, T.; Kühn, M.A.L. Uniting what belongs together—Reevaluation of the isopod species Macrostylis grandis and M. ovata using ontogenetic, morphological and genetic evidence. Prog. Oceanogr. 2020, 181, 102238. [Google Scholar] [CrossRef]
- Malyutina, M.V.; Brandt, A. Munnopsidae (Crustacea, Isopoda, Asellota) from the Kuril–Kamchatka Trench with a regional and inter-ocean comparison of their biogeographic and richness patterns. Prog. Oceanogr. 2020, 183, 102289. [Google Scholar] [CrossRef]
- Elsner, N.O.; Golovan, O.A.; Malyutina, M.V.; Brandt, A. Alone in the dark: Distribution, population structure and reproductive mode of the dominant isopod Eurycope spinifrons Gurjanova, 1933 (Isopoda: Asellota: Munnopsidae) from bathyal and abyssal depths of the Sea of Japan. Deep Res. Part II Top. Stud. Oceanogr. 2013, 86–87, 103–110. [Google Scholar] [CrossRef]
- Riehl, T.; Wilson, G.D.F.; Hessler, R.R. New Macrostylidae Hansen, 1916 (Crustacea: Isopoda) from the Gay Head-Bermuda Transect with special consideration of sexual dimorphism. Zootaxa 2012, 3277, 1–26. [Google Scholar] [CrossRef][Green Version]
- Kim, J.; Kim, J.; Lee, W.; Karanovic, I. Two new Uromunna species (Isopoda: Asellota: Munnidae) from the Korean Peninsula and their phylogenetic position within munnoid groups. Diversity 2022, 15, 20. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).