
A New Enigmatic Teleost Fish from the Mid-Cretaceous of Lebanon


Abstract
:1. Introduction
2. Geological Setting
3. Materials and Methods
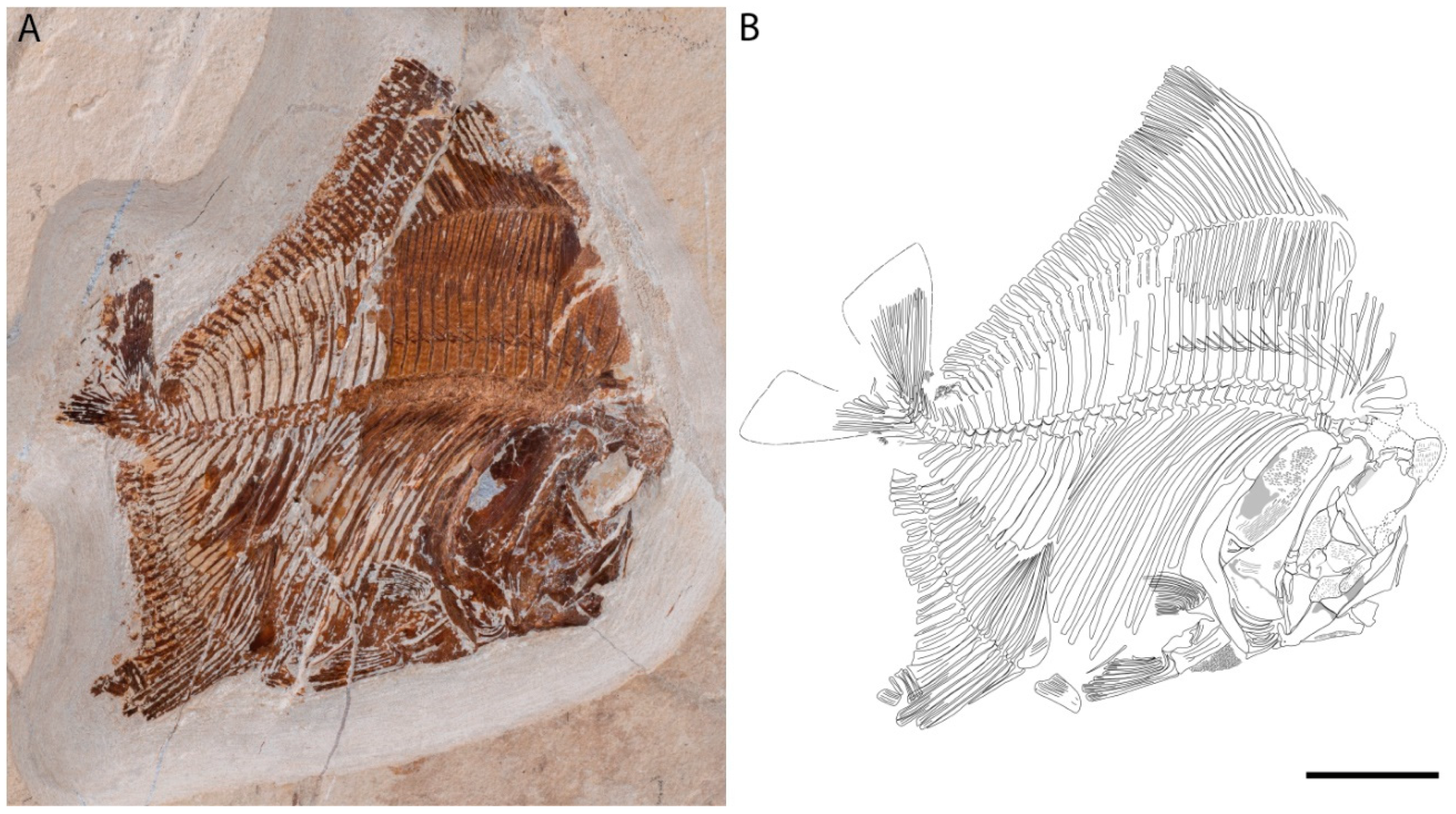
3.1. Described Material
3.2. Comparative Material
3.3. Abbreviations
4. Results
4.1. Systematic Paleontology
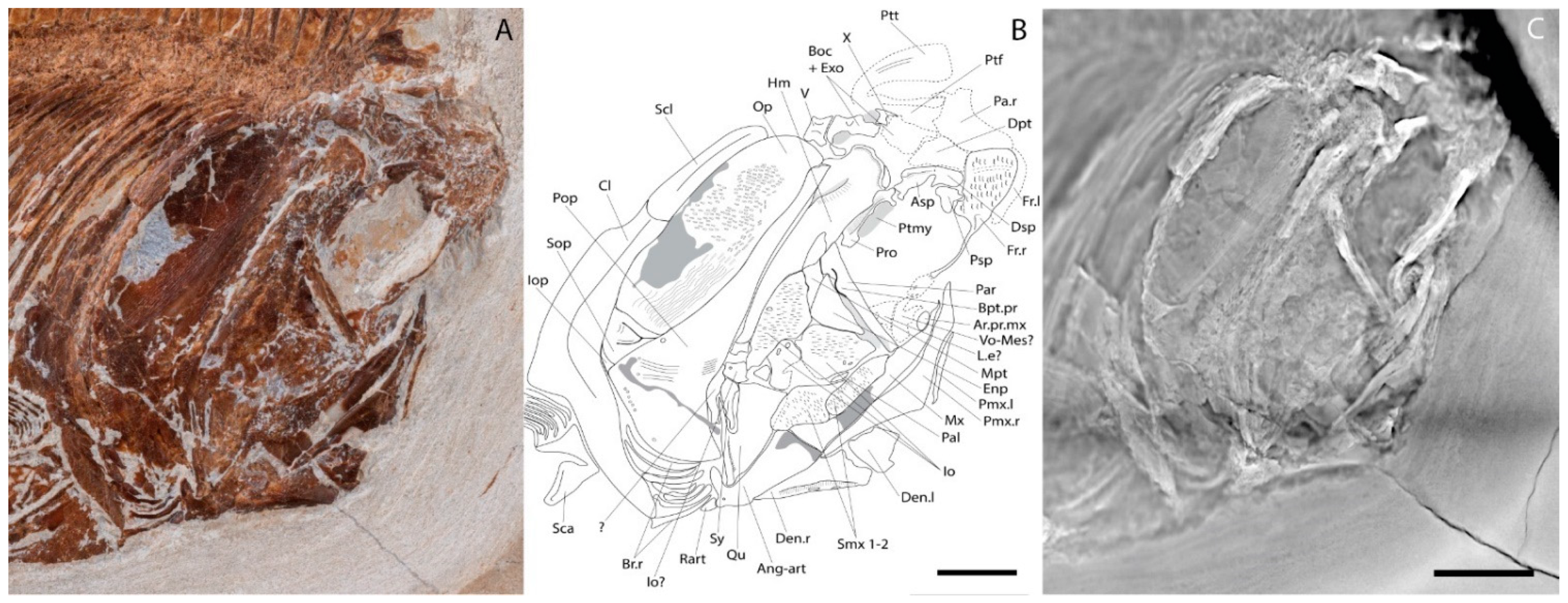
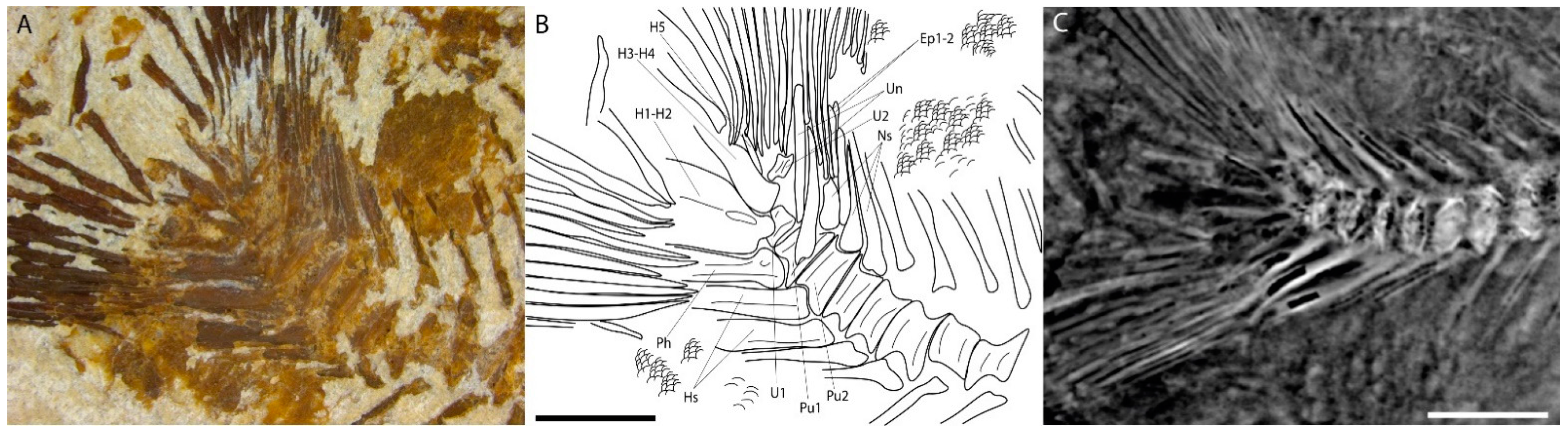
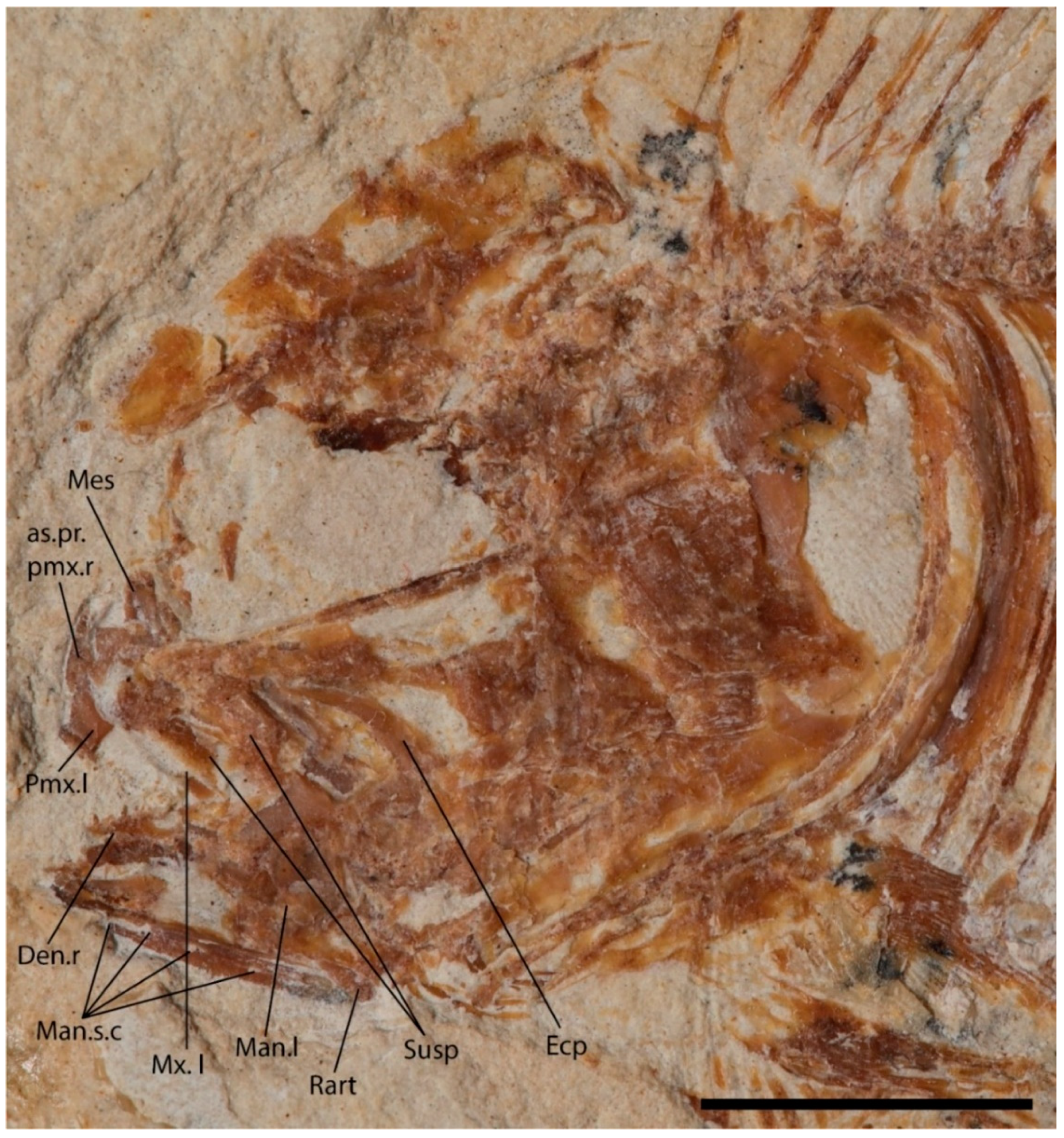
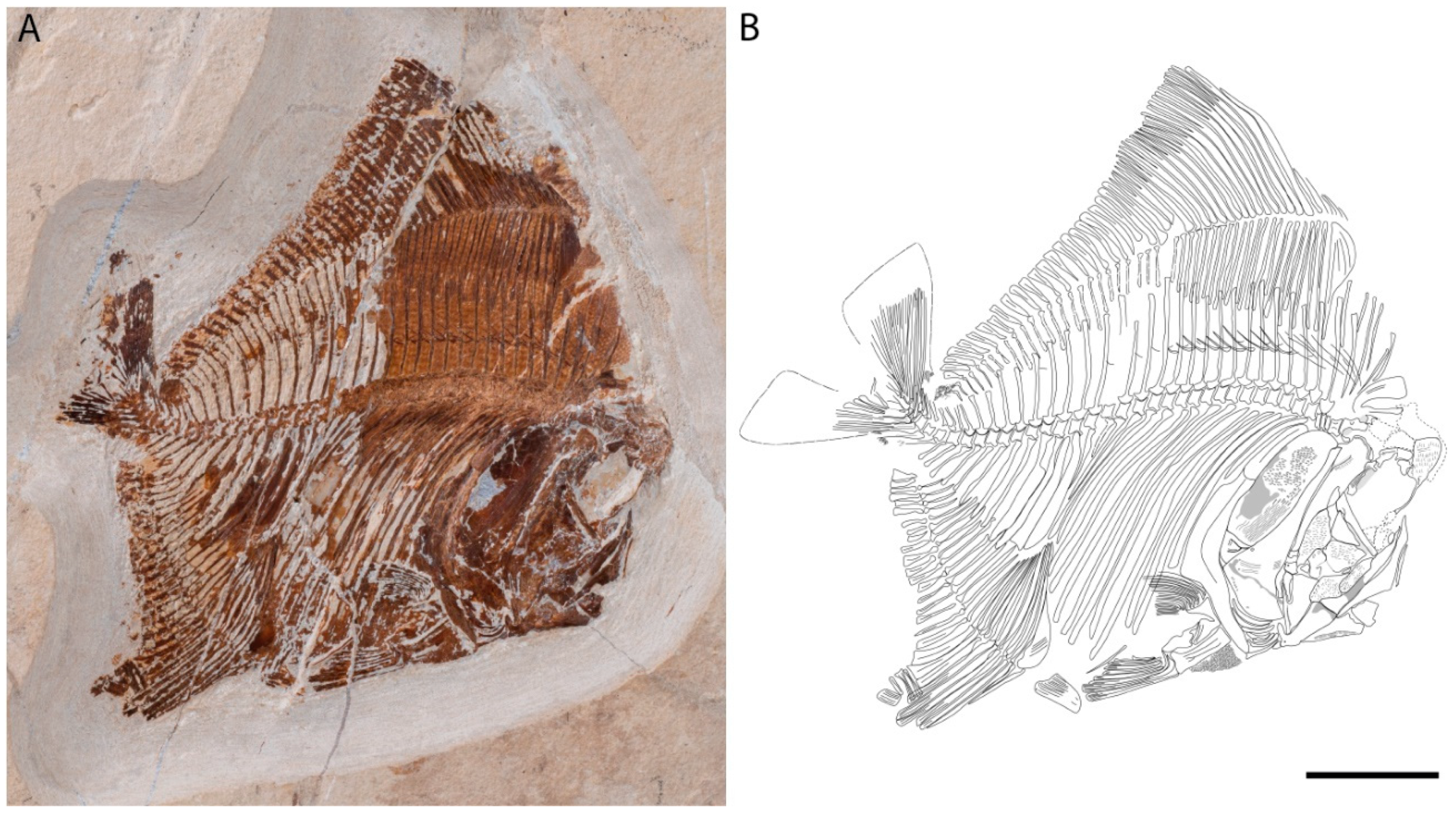
4.2. Description
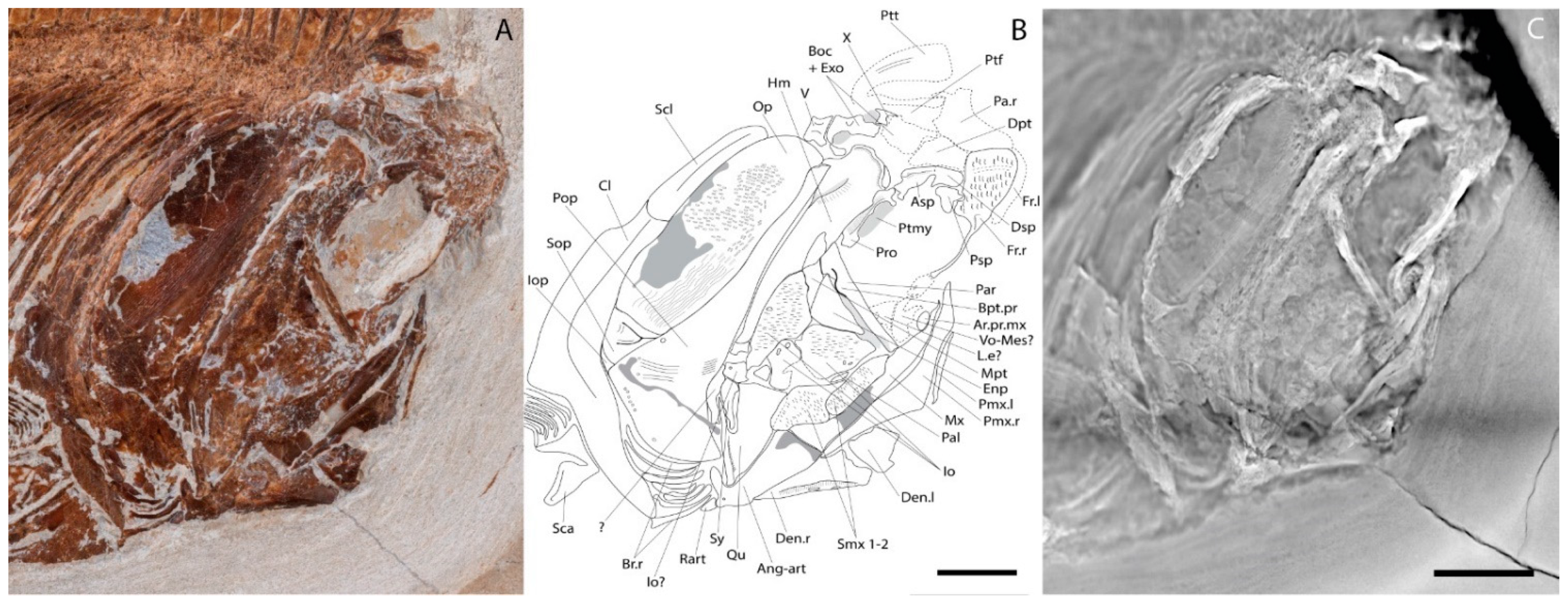
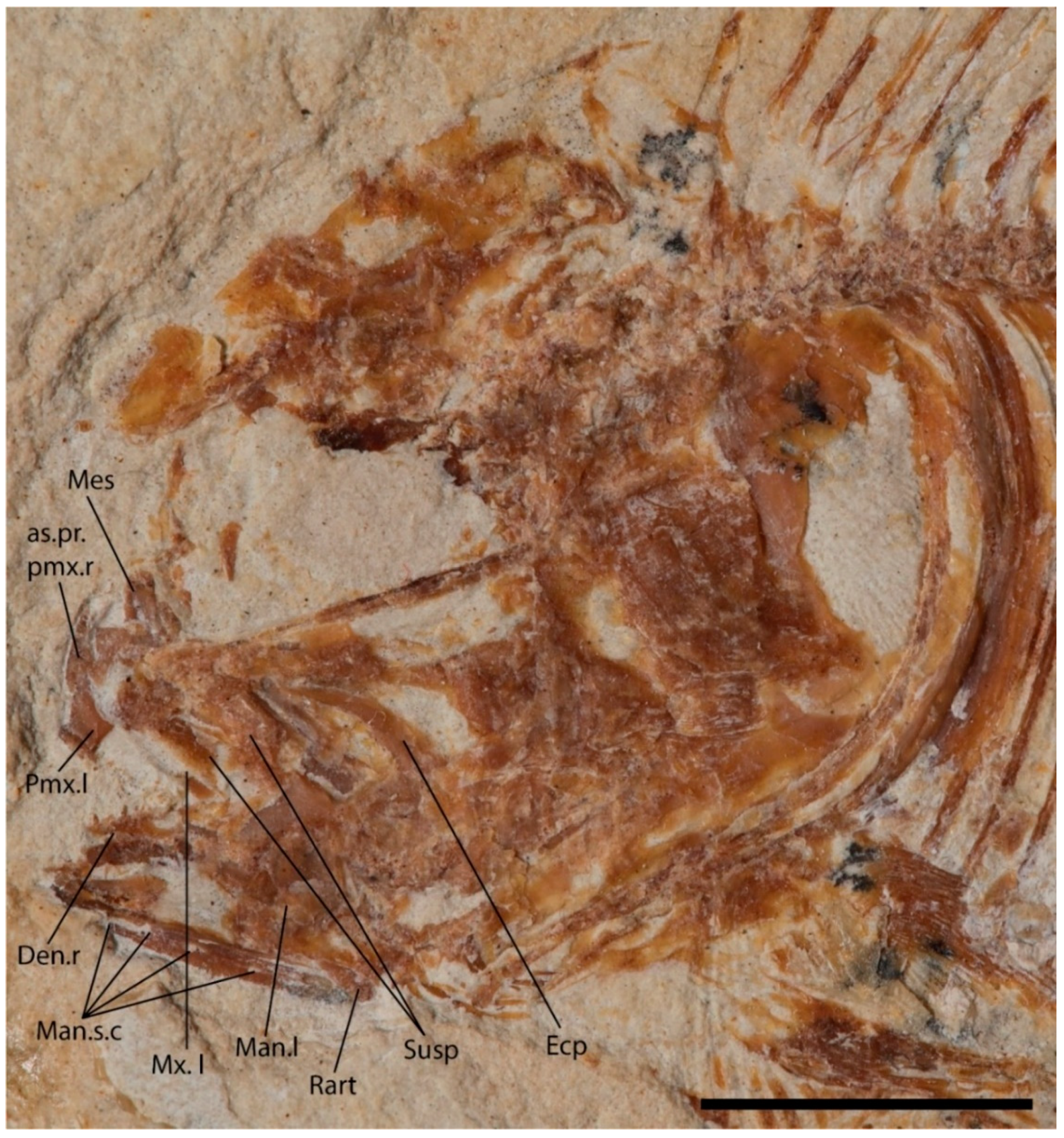
4.2.1. Cranium
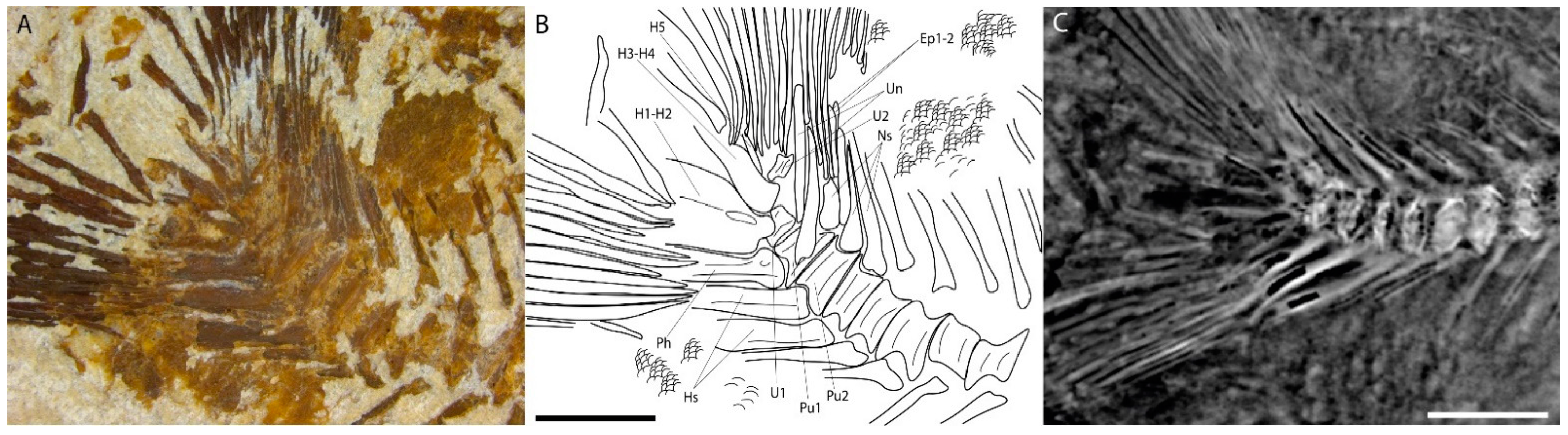
4.2.2. Post-Cranium
5. Systematic Affinities
5.1. The Tselfatiiformes and the Protobramidae
5.2. Araripichthys
6. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taverne, L.; Gayet, M. Phylogenetical relationships and palaeozoogeography of the marine Cretaceous Tselfatiiformes (Teleostei, Clupeocephala). Cybium 2005, 29, 65–87. [Google Scholar]
- Taverne, L. Ostéologie et affinités systématiques de Tselfatia formosa, téléostéen fossile du Crétacé supérieur de la Mésogée eurafricaine. Ann. Soc. Roy. Zool. Belg. 1983, 113, 165–181. [Google Scholar]
- Taverne, L. Révision de Zanclites xenurus, téléostéen (Pisces, Tselfatiiformes) marin du Santonien (Crétacé supérieur) du Kansas (États-Unis). Belg. J. Zool. 1999, 129, 421–438. [Google Scholar]
- Taverne, L. Tselfatia formosa, téléostéen marin du Crétacé (Pisces, Actinopterygii), et la position systématique des Tselfatiiformes ou Bananogmiiformes. Geodiversitas 2000, 22, 5–22. [Google Scholar]
- Taverne, L. Révision du genre Martinichthys, poisson marin (Teleostei, Tselfatiiformes) du Crétacé supérieur du Kansas (États-Unis). Geobios 2000, 33, 211–222. [Google Scholar]
- Taverne, L. Ostéologie et position systématique du genre Plethodus et des nouveaux genres Dixonanogmius et Pentanogmius, poissons marins du Crétacé (Teleostei, Tselfatiiformes). Biologisch Jaarboek Dodonaea 2000, 67, 94–123. [Google Scholar]
- Taverne, L. Eoplethodus chaneti gen. et sp. nov. de l’Albien (Crétacé) marin de Provence (France) et l’évolution du squelette caudal chez les Tselfatiiformes (Osteichthyes, Teleostei). Cybium 2000, 24, 371–381. [Google Scholar]
- Taverne, L. Révision de Niobrara encarsia, téléostéen (Osteichthyes, Tselfatiiformes) du Crétacé supérieur marin du Kansas (États-Unis). Belg. J. Zool. 2001, 131, 3–16. [Google Scholar]
- Taverne, L. Révision du genre Bananogmius (Teleostei, Tselfatiiformes), poisson marin du Crétacé supérieur d’Amérique du Nord et d’Europe. Geodiversitas 2001, 23, 17–40. [Google Scholar]
- Taverne, L. Révision de Syntegmodus altus (Teleostei, Tselfatiiformes), poisson marin du Crétacé supérieur du Kansas (États-Unis). Cybium 2001, 25, 251–260. [Google Scholar]
- Taverne, L. Révision de Luxilites striolatus, poisson marin (Teleostei, Tselfatiiformes) du Crétacé supérieur du Kansas (États-Unis). Belg. J. Zool. 2002, 132, 25–34. [Google Scholar]
- Taverne, L. Étude de Pseudanogmius maiseyi gen. et sp. nov., poisson marin (Teleostei, Tselfatiiformes) du Crétacé supérieur du Kansas (États-Unis). Geobios 2002, 35, 605–614. [Google Scholar]
- Taverne, L. Redescription critique des genres Thryptodus, Pseudothryptodus et Paranogmius, poissons marins (Teleostei, Tselfatiiformes) du Crétacé supérieur des États-Unis, d’Égypte et de Libye. Belg. J. Zool. 2003, 133, 163–173. [Google Scholar]
- Taverne, L. Ostéologie de Pentanogmius evolutus (Cope, 1877) n. comb. (Teleostei, Tselfatiiformes) du Crétacé supérieur marin des États-Unis. Remarques sur la systématique du genre Pentanogmius Taverne, 2000. Geodiversitas 2004, 26, 89–113. [Google Scholar]
- Taverne, L.; Gayet, M. Ostéologie et relations phylogénétiques des Protobramidae (Teleostei, Tselfatiiformes) du Cénomanien (Crétacé supérieur) du Liban. Cybium 2004, 28, 285–314. [Google Scholar]
- Le Danois, E.; Le Danois, Y. L’ordre des Scombres. Mémoires L’ifan 1963, 68, 153–192. [Google Scholar]
- Patterson, C. A second specimen of the Cretaceous teleost Protobrama and the relationships of the sub-order Tselfatioidei. Ark. Fur Zool. 1967, 19, 215–234. [Google Scholar]
- Bardack, D. Fossil vertebrates from the marine Cretaceous of Manitoba. Can. J. Earth Sci. 1968, 5, 145–153. [Google Scholar]
- Taverne, L. À propos de Tselfatia Arambourg, C., 1943 (Cénomanien du Maroc), de Protobrama (Cénomanien du Liban) et du rapport éventuel des Tselfatioïdes fossiles (Crétacé) et des Téléostéens du super-ordre des Ostéoglossomorphes. Ann. Soc. R. Zool. Belg. 1975, 105, 53–72. [Google Scholar]
- Nelson, J.S. Fishes of the World, 3rd ed.; John Wiley & Sons: New York, NY, USA, 1994; p. 600. [Google Scholar]
- Cavin, L. Osteology and phylogenetic relationships of the teleost Goulmimichthys arambourgi Cavin, 1995, from the Upper Cretaceous of Goulmima, Morocco. Ecolog. Geol. Helv. 2001, 94, 509–535. [Google Scholar]
- Santos, R.d.S. Araripichthys castilhoi novo genêro e espécie de Teleostei da Formação Santana, Chapada do Araripe, Brasil. Série Geol. Dep. Nac. Produção Miner. Bras. 1985, 29, 133–139. [Google Scholar]
- Maisey, J.G. Elopomorph fishes from Santana Formation. In Proceedings of the Atas do I Simpósio sobre a Bacia do Araripe e Bacias Interiores do Nordeste, Crato, Brasil, 14–16 June 1990. [Google Scholar]
- Maisey, J.G.; Blum, S. Araripichthys. In Santana Fossils: An Illustrated Atlas; Maisey, J.G., Ed.; TFH Publications: Neptune, NJ, USA, 1991; pp. 208–217. [Google Scholar]
- Patterson, C. An overview of the early fossil record of acanthomorphs. Bull. Mar. Sci. 1993, 52, 29–59. [Google Scholar]
- Arratia, G.; Chorn, J. A new primitive acanthomorph fish from the Greenhorn Formation (Late Cretaceous) of Nebraska. J. Vertebr. Paleontol. 1998, 18, 301–314. [Google Scholar]
- Maisey, J.G.; Moody, J.M. A review of the problematic extinct teleost fish Araripichthys, with description of a new species from the Lower Cretaceous of Venezuela. Am. Mus. Novit. 2001, 3324, 27. [Google Scholar]
- Cavin, L. Nouveaux Teleostei du gisement du Turonien inférieur de Goulmima (Maroc). Comptes Rendus Académie Sci. 1997, 325, 719–724. [Google Scholar]
- Alvarado-Ortega, J.; Brito, P.M. A new species of Araripichthys (Teleostei, Elopocephala) from the Tlauyúa Formation (Cretaceous, Albian), Mexico. J. Vertebr. Paleontol. 2011, 31, 1376–1381. [Google Scholar]
- Mayrinck, D.; Meunier, F.J.; Cupello, C.; Brito, P.M. The paleohistology of †Araripichthys castilhoi from the Lower Cretaceous of Araripe Basin, Northeastern Brazil: A typical case of basal teleost. J. Vertebr. Paleontol. 2023, 42, e2157732. [Google Scholar]
- Hückel, U. Die Fischschiefer von Haqel und Hjoula in der Oberkreide des Libanon. Neues Jahrb. Für Geol. Und Paläontologie Abh. 1970, 135, 113–149. [Google Scholar]
- Maksoud, S.; El Hossny, T.; Piuz, A.; Cavin, L.; Abi Saad, P.; Azar, D.; Samankassou, E. Biostratigraphy and palaeoenvironments of the Upper Cretaceous fossil fish Konservat-Lagerstätten of Lebanon. Department of Earth Sciences, University of Geneva, Rue des Maraîchers 13, Geneva, Switzerland. article in preparation.
- Hückel, U. Vergleich des Mineralbestandes der Plattenkalke Solnhofens und des Libanon mit anderen Kalken. Neues Jahrb. Für Geol. Und Paläontologie Abh. 1974, 145, 153–182. [Google Scholar]
- Dubertret, L. Notice Explicative de la Carte Géologique du Liban au 200.000 ème; Ministère des Travaux Publics: Beyrouth, Liban, 1955; p. 74.
- Botta, P.E. Observations sur le Liban et l’Anti-Liban. Mémoire La Société Géologique Fr. 1833, 1, 135–160. [Google Scholar]
- Fuchs, D.; Bracchi, G.; Weis, R. New octopods (Cephalopoda: Coleoidea) from the Late Cretaceous (Upper Cenomanian) of Hâkel and Hâdjoula, Lebanon. Palaeontology 2009, 52, 65–81. [Google Scholar]
- Walley, C. The lithostratigraphy of Lebanon. Leban. Sci. Bull. 1997, 10, 81–108. [Google Scholar]
- Fraas, O.F. Aus dem Orient: Geologische Beobachtungen am Libanon; Schweizerbart: Stuttgart, Germany, 1878; p. 278. [Google Scholar]
- Pictet, F.-J.; Humbert, A. Nouvelles recherches sur les poissons fossiles du Mont Liban. Arch. Des Sci. Phys. Nat. Genève 1866, 26, 117–133. [Google Scholar]
- Douvillé, H. Études sur les rudistes: Rudistes de Sicile, d’Algérie, d’Égypte, du Liban et de la Perse. Société Géologique Fr. 1910, 1, 84. [Google Scholar]
- Zumoffen, G. Géologie du Liban; Henry Barrère: Paris, France, 1926; p. 165. [Google Scholar]
- Patterson, C. New Cretaceous berycoid fishes from the Lebanon. Bull. Br. Mus. Nat. Hist. Geol. 1967, 14, 69–109. [Google Scholar]
- Saint-Marc, P. Étude géologique de la région d’Hermel (Liban septentrional). Bull. La Société Géologique Fr. 1969, 11, 379–387. [Google Scholar]
- Saint-Marc, P. Étude stratigraphique et micropaléontologique de l’Albien, du Cénomanien et du Turonien du Liban. Notes Mémoires Sur Le Moyen-Orient 1974, 13, 1–342. [Google Scholar]
- Sowerby, J. The Mineral Conchology of Great Britain; or Coloured Figures and Descriptions of Those Remains of Testaceous Animals and Shells, Which Have Been Preserved at Various Times and Depths in the Earth; Biodiversity Heritage Library: London, UK, 1812–[1846]; p. 609. [Google Scholar]
- Hemleben, C. Rote Tiden und die oberkretazischen Plattenkalke im Libanon. Neues Jahrbuchfiir Geol. Und Paläontologie Monatshefte 1977, 77, 239–255. [Google Scholar]
- Wippich, M.E.; Lehmann, H. Allocrioceras from the Cenomanian (mid-Cretaceous) of the Lebanon and its bearing on the palaeobiological interpretation of heteromorphic ammonites. Palaeontology 2004, 47, 1093–1107. [Google Scholar]
- Shumard, B.F. Descriptions of new Cretaceous fossils from Texas. Trans. Acad. Sci. St. Louis 1860, 1, 590–610. [Google Scholar]
- Forey, P.L. A revision of the Elopiformes fishes, fossil and recent. Bull. Br. Mus. 1973, 10, 1–222. [Google Scholar]
- Müller, J. Über den Bau und die Grenzen der Ganoiden und über das natürliche System der Fische. Arch. Für Naturgeschichte 1845, 11, 129. [Google Scholar]
- Nelson, G.J. Notes on the Structure and Relationships of Certain Cretaceous and Eocene Teleostean Fishes. Am. Mus. Novit. 1973, 2524, 1–31. [Google Scholar]
- Patterson, C.; Rosen, D.E. Review of the ichthyodectiform and other Mesozoic teleost fishes and the theory and practice of classifying fossils. Bull. Am. Mus. Nat. Hist. 1977, 158, 83–172. [Google Scholar]
- Arratia, G. The Clupeocephala re-visited: Analysis of characters and homologies. Rev. De Biol. Mar. Y Oceanogr. 2010, 45, 635–657. [Google Scholar]
- Taverne, L.; Capasso, L. A new insight into the Protobramidae (Teleostei, Tselfatiiformes), a fossil fish family from the marine Cenomanian (Upper Cretaceous) of Lebanon. Geo-Eco-Trop 2020, 44, 355–367. [Google Scholar]
- Parey, E.; Louis, A.; Montfort, J.; Bouchez, O.; Roques, C.; Iampietro, C.; Lluch, J.; Castinel, A.; Donnadieu, C.; Desvignes, T.; et al. Genome structures resolve the early diversification of teleost fishes. Science 2023, 379, 572–575. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Counts | Ypsiloichthys sibelleae |
|---|---|
| Vertebrae (Total) | 39–40 |
| Abdominal centra | 18 |
| Caudal centra | 21–22 |
| Hypurals | 5 |
| Dorsal pterygiophores | 48–49 |
| Anal pterygiophores | 26–27 |
| Pectoral fin rays | 20–21 |
| Pelvic fin rays | 8 |
| Dorsal fin rays | 49–51 |
| Anal fin rays | 31–33 |
| Caudal fin rays (Total) | 34 |
| Branchiostegal rays | 6–7 |
| Ribs (in pairs) | 15–16 |
| Measurements (as % of SL) | Ypsiloichthys sibelleae |
|---|---|
| Total length (mm) | 102 |
| Standard length (mm) | 85 |
| Maximum body depth | 84 |
| Head length | 40 |
| Head depth | 49 |
| Predorsal length | 21 |
| Peanal length | 79 |
| Dorsal fin base length | 77 |
| Anal fin base length | 50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Hossny, T.; Cavin, L. A New Enigmatic Teleost Fish from the Mid-Cretaceous of Lebanon. Diversity 2023, 15, 839. https://doi.org/10.3390/d15070839
El Hossny T, Cavin L. A New Enigmatic Teleost Fish from the Mid-Cretaceous of Lebanon. Diversity. 2023; 15(7):839. https://doi.org/10.3390/d15070839
Chicago/Turabian StyleEl Hossny, Tamara, and Lionel Cavin. 2023. "A New Enigmatic Teleost Fish from the Mid-Cretaceous of Lebanon" Diversity 15, no. 7: 839. https://doi.org/10.3390/d15070839
APA StyleEl Hossny, T., & Cavin, L. (2023). A New Enigmatic Teleost Fish from the Mid-Cretaceous of Lebanon. Diversity, 15(7), 839. https://doi.org/10.3390/d15070839